
Abstract
Background
Kangaroo rats are small mammals that are among the most abundant vertebrates in many terrestrial ecosystems in Western North America and are considered both keystone species and ecosystem engineers, providing numerous linkages between other species as both consumers and resources. However, there are challenges to studying the behavior and activity of these species due to the difficulty of observing large numbers of individuals that are small, secretive, and nocturnal. Our goal was to develop an integrated approach of miniaturized animal-borne accelerometry and radiotelemetry to classify the cryptic behavior and activity cycles of kangaroo rats and test hypotheses of how their behavior is influenced by light cycles, moonlight, and weather.
Methods
We provide a proof-of-concept approach to effectively quantify behavioral patterns of small bodied (< 50 g), nocturnal, and terrestrial free-ranging mammals using large acceleration datasets by combining low-mass, miniaturized animal-borne accelerometers with radiotelemetry and advanced machine learning techniques. We developed a method of attachment and retrieval for deploying accelerometers, a non-disruptive method of gathering observational validation datasets for acceleration data on free-ranging nocturnal small mammals, and used these techniques on Merriam’s kangaroo rats to analyze how behavioral patterns relate to abiotic factors.
Results
We found that Merriam’s kangaroo rats are only active during the nighttime phases of the diel cycle and are particularly active during later light phases of the night (i.e., late night, morning twilight, and dawn). We found no reduction in activity or foraging associated with moonlight, indicating that kangaroo rats are actually more lunarphilic than lunarphobic. We also found that kangaroo rats increased foraging effort on more humid nights, most likely as a mechanism to avoid cutaneous water loss.
Conclusions
Small mammals are often integral to ecosystem functionality, as many of these species are highly abundant ecosystem engineers driving linkages in energy flow and nutrient transfer across trophic levels. Our work represents the first continuous detailed quantitative description of fine-scale behavioral activity budgets in kangaroo rats, and lays out a general framework for how to use miniaturized biologging devices on small and nocturnal mammals to examine behavioral responses to environmental factors.
Similar content being viewed by others


Background
Small mammals are among the most abundant vertebrates in many terrestrial ecosystems, providing numerous linkages between other species as both consumers and resources [1,2,3,4,5,6,7]. Many small mammals act as ecosystem engineers, excavating extensive burrow systems that impact vegetation and provide habitat for many other species [8,9,10,11]. Despite their importance in the broader community, we still lack detailed information on key aspects of the behavior and activity of many small mammals, as numerous species are nocturnal, secretive, semi-fossorial, and thus difficult to study using traditional observation techniques.
Detailed fine-scale data on the activity and behavior of keystone species represents an important component of community ecology, as this information provides invaluable insight into interactions with other species and broader ecological niche parameters [12,13,14]. However, many methods used to quantify behavior, such as direct observation, are limited by expense, time, animal crypsis, physical or temporal barriers, or the impact of a human observer on the natural expression of behavior [15, 16]. In recent years, some of these shortcomings have been addressed by advancements in biologging technology that use miniaturized animal-borne tags to collect data about movement, behavior, physiology, and/or abiotic conditions [17,18,19]. These novel next generation natural history biologging techniques provide a much deeper understanding of the behavior and ecology of species that are both easy to observe in nature and ones that are not amenable to direct observation [18, 20].
The use of accelerometers to quantify behavior and activity has its own set of limitations, including the need to create detailed validation datasets, relatively crude classifications of behavioral states, the lack of broader ecological context for behavioral expression, and the inability of smaller species to carry large enough devices for long enough periods of time [17]. These limitations have led to taxonomic bias in the use of accelerometry, with most studies being restricted to larger-bodied mammals, birds, and charismatic megafauna that are typically diurnal or crepuscular (and thus easier to observe for validation [17]). However, statistical approaches for analyzing animal movements continue to be refined using both supervised and unsupervised machine learning (reviewed in Hoffman et al. [21]), and the continued miniaturization of biologgers is creating opportunities for studying smaller-bodied species.
To date, the smallest mammals that have been studied using advanced biologging techniques to quantify activity and behavioral patterns are Western chipmunks (Tamias alpinus and T. speciosus), diurnal sciurid rodents weighing ~ 50 g. Hammond et al. [22] attached miniaturized accelerometers (1.5–2.5 g) for approximately 2.5 d per individual to measure activity patterns. This study established a general framework for using animal-borne accelerometry on small-bodied animals, but there have not yet been any studies on smaller bodied species that quantify more fine-scaled behavioral states (other than just active versus inactive) or deploy logging devices over longer periods of time. In contrast, the use of accelerometers to study the behavior of large mammals has provided insight on not only activity patterns, but also details of movement type, hunting behaviors, and energy expenditure (e.g., [23,24,25]).
Here, we use animal-borne accelerometry to classify the behavior of free-ranging kangaroo rats (Dipodomys merriami), a nocturnal heteromyid rodent (< 50 g) that is the most abundant terrestrial mammal in many arid regions of North America [26]. Kangaroo rats are semi-fossorial and granivorous, storing and dispersing plant seeds while digging extensive burrow systems. Numerous studies have shown that kangaroo rats can have profound effects on their broader community, acting as both keystone species and ecosystem engineers [27,28,29,30]. Additionally, they are a major prey item for several predators at higher trophic levels such as coyotes and foxes [31, 32], rattlesnakes [3, 6], and birds of prey [33,34,35]. Although there has been significant research on kangaroo rat activity and foraging patterns, it has been limited by the inherent shortcomings associated with direct observation, and we still lack important information on many aspects of the activity and behavior of these critical species (e.g., relative levels of activity during different phases of the diel cycle and how that activity is affected by various environmental parameters).
Our study uses animal-borne accelerometry to classify the behavior of free-ranging individuals, highlighting the utility of this method for collecting comprehensive datasets on the behavior and activity of secretive, nocturnal, and small-bodied terrestrial mammals that play critical roles in ecosystems across the globe. We developed an integrative approach using miniaturized accelerometry and radiotelemetry to identify ecologically relevant behavioral categories such as traveling and foraging. Additionally, we pioneered techniques to validate these behaviors in the field through focal observation and recording of free-ranging individuals, and then explored how different daily and nightly light phases and environmental variables influenced the behavior and activity cycles of Merriam’s kangaroo rats. In particular, our study allowed us to address the question of how variation in moonlight impacts kangaroo rat activity, an important question steeped in uncertainty, with a number of conflicting studies supporting either lunarphilia or lunarphobia [36,37,38,39,40,41].
Methods
Study site, capture procedures, and species
The study was carried out May-August 2021 at Marathon Grassland Preserve (MGP), a 1093-ha Chihuahuan desert grassland habitat located in West Texas, USA. The site was dominated by Trans-Pecos loamy plains grassland and sparse creosote scrub. We captured Merriam’s kangaroo rats (D. merriami) using Sherman live traps and marked captured individuals with 10 mm Biomark pit-tag injected subcutaneously on the dorsum. Kangaroo rats were weighed to the nearest gram, and individuals that were > 40 g in mass (n = 14; 9 females and 5 males; Additional File 1; Table S1) were fitted with a ~ 2.5 g accelerometer/Very High Frequency (VHF) package to the dorsum following a similar harnessing technique to that of Shier & Swaisgood [42]. The package included an accelerometer (model AXY 5, Technosmart Europe Srl., Rome, Italy; dimensions = 20 L x 15 W x 9 H mm; mass = ~ 1.8 g), a VHF microtransmitter (Wildlife Materials model SOM-2011; mass = 0.8 g) allowing for the relocation of individuals and removal of the accelerometer after the ~ 5-day monitoring period (range = 1–6.5 days; some deployments ended early due to early battery discharge or device attachment failure). Accelerometers were triaxial, placed in the same orientation across all individuals, and set to record forces between − 10 and 10 gforce at 25 hz. After attachment individuals were held for ~ 20 min to ensure good fit of bio-loggers (i.e., they were unable to roll over and remove the backpack) and then released at their point of capture. Individuals were then periodically relocated via radiotelemetry (typically twice a day) for validation recording and to verify retention of bio-loggers. If bio-loggers were still retained after 6 days, individuals were re-trapped for removal of the device and then immediately released at point of capture.
Free-ranging behavioral observations and scoring
We used VHF radio tracking (Advanced Telemetry Systems R410 Receiver) and a handheld flexible 3-element Yagi antenna to locate tagged kangaroo rats. Once individuals were located, two observers would work together with a telephoto video camera (10X optical zoom) with infrared (IR) recording capability (Sony Handycam Model DCR-SR80) and an IR flashlight (UniqueFire, 1605 T38 850 nm; invisible to kangaroo rats) to video record behaviors of kangaroo rats at a distance over 10 m, moving slowly and carefully to avoid unnecessary disturbance. Individuals were filmed for as long as they were visible. We spent approximately 3 h per night throughout the sampling period tracking kangaroo rats and recording videos for behavioral validation when they were active on the surface. From these efforts, we scored all videos that showed clear and unambiguous examples of kangaroo rats exhibiting the behaviors described in our ethogram, resulting in 122 m of validated video observations (Additional File 1; Table S2).
Behavioral scores were time matched to accelerometer readings (to within 1 s) to generate annotated acceleration datasets by recording a video of the exact time (using the ExactTime™ software application by ©Neurovat, 2023) as the accelerometry device began logging data (Tecnosmart devices emit a visible signal when powering on). We then also recorded the ExactTime™ (©Neurovat, 2023) to the nearest second at the outset of all validation video recordings of kangaroo rats wearing accelerometry devices [43], and (as a further redundancy) whenever accelerometry devices were turned off after a deployment. All units were calibrated by Technosmart before being used in the field. Additionally, we manually examined accelerometry signatures for any evidence of clock drift when scoring validation videos of individuals many days after the onset of datalogging. We found no evidence of clock drift over the ~ 6.5 d that loggers were deployed. We initially viewed exploratory videos in order to construct a detailed ethogram and determine what potential behavioral classes could be feasibly incorporated into our modeling framework (Figure S1). Additional review determined that several behaviors (e.g., digging, kicking sand, rolling) were too rare for incorporation into the modeling framework, which led us to group behaviors into four broad classes that would be frequent enough to train a model and inclusive of key activities: motionless, traveling, foraging, and grooming (Table 1).
Behavioral classification algorithms
We used the open-access web application AcceleRater to train classification algorithms based on our free-ranging kangaroo rat acceleration datasets [44]. First, because class imbalance can lead to inference issues when using machine-learning [45], all datasets were manually balanced so that each behavioral class was subsampled to the behavior that had the fewest samples from our free-ranging kangaroo rat acceleration datasets (in all cases, this was either the behavior class foraging or grooming). We then ran a linear support vector machine, a decision tree, and a random forest algorithm (as these models are typically used when using acceleration data to predict animal behavior) that either included all summary statistics available in AcceleRater (mean acceleration, standard deviation, maximum, minimum, vector norm, covariance, Pearson correlation, dynamic body acceleration, overall dynamic body acceleration, mean-diff, std-diff, wave amplitude, line crossings, 25 percentile, 50 percentile, and 75 percentile) or just the summary statistics of mean acceleration, standard deviation of acceleration, and overall dynamic body acceleration at what we deemed biologically relevant and distinct window sizes of 1, 2, 3, 6, and 9 s. For each unique model and window size, we trained the model via a (50/50) train-test split, where a random 50% of the data was used to train each model and the remaining 50% of the data set was used to test the model. We implemented the same methods as Clermont et al. [23] and Hanscom et al. [43] to identify which algorithm optimally classified behaviors and we calculated accuracy, precision, and recall from rates of true positives, false positives, and false negatives.
Kangaroo rat diel activity budgets and response to moonlight and weather
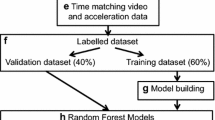
The stepwise methodological workflow for behavioral classification of accelerometry data and assessment of activity budgets is depicted in Fig. 1.We used the top performing algorithm from our model selection procedure to annotate our complete accelerometry dataset. We then determined kangaroo rat activity budgets by calculating proportions of time and expression for each behavior across all individuals.

Stepwise methodological workflow for the behavioural classification of accelerometry data and the development of an activity budget using short duration, high frequency accelerometry on a small mammal < 50 g. More details of each step presented in this workflow can be found in the methods section
Specifically, we determined behavioral activity budgets at different light phases, moon intensity and illumination, and moon position (following several previous ecological studies [46,47,48]). We used the suncalc package in R to retrieve daily light phase times, moonrise and moonset times, and sunrise and sunset times for our study site [49]. Moon illumination levels were calculated on a continuous scale ranging from 0.00 (new moon) to 1.00 (full moon). To further understand how moon illumination impacted activities, we converted moon illumination levels into a three-level categorical variable called moon intensity, where < 0.33 = low, 0.33–0.66 = medium, and > 0.66 = high [48]. The eight light phases we used are defined by the position of the sun relative to the horizon and are the following: evening (last hour above horizon; 1 h), dusk (0–6° below horizon; ~0.5 h), evening twilight (6–18° below horizon; ~1.25 h), night (> 18° below horizon; ~6.5 h), morning twilight (18–6° below horizon; ~1.25 h), dawn (6–0° below horizon; ~0.5 h), morning (first hour above horizon; 1 h), and day (above horizon minus the first and last hour sunlight; ~12 h; Fig. 2; [48]). Our study site is located in an arid and dry environment, and thus almost all nights were clear with no cloud cover. However, to be conservative, we also reran analyses after removing all nights with a precipitation rate > 0, since cloud cover is correlated with precipitation rate. Lastly, because environmental variables such as temperature, wind, and relative humidity have been shown to impact small mammal activity as estimated from trapping data [50,51,52], we investigated how they impacted accelerometry-based estimates of kangaroo rat behavior. We averaged weather data (average nightly air temperature, wind speed, and relative humidity) from two nearby weather stations (KTXMAR8 and KTXALPIN66; both located in similar habitat to our site and ca. 20 km straight-line distance from the field site) included in the Weather Underground network [53, 54].

An illustration of light phases considered during the study period. Artwork for this figure was designed and produced by Alexandra Coots
We examined the effect of moonlight and environmental variables on behaviors in a generalized linear mixed effects modeling (GLMM) framework. First, we investigated the diel activity patterns of kangaroo rats by examining whether the proportion of time spent in each behavioral state (motionless, traveling, grooming, and foraging) was influenced by light phase (all eight light phases as defined above) and sex as fixed effects. Next, we examined whether the proportion of time spent in each behavioral state was influenced by moon intensity (low, medium, or high), moon position (up or down), and their interaction, as well as sex, as fixed effects. This analysis only included light phases when the sun was > 0° below the horizon (i.e., dusk, evening twilight, night, morning twilight, and dawn). Lastly, we used a GLMM to investigate how the abiotic factors of temperature, wind speed, and relative humidity affected kangaroo rat behavioral state at a nightly scale. Additionally, we included moon illumination as a nightly continuous variable in this model to corroborate any patterns found prior regarding the influence of moonlight on kangaroo rat behavior. For each behavior, we explored multiple distribution families, aiming to identify the most suitable family for modeling the data. The candidate distribution families considered were Gaussian, Poisson, Binomial, and Gamma. We employed the Akaike Information Criterion (AIC) as the primary model selection criterion, and then used the top model with the associated best family as the GLMM for each behavior (Additional File 1; Table S3). All environmental variables, as well as sex, were included as fixed effects. To avoid inferential errors associated with collinearity, we calculated the variable inflation factors (VIF) for each predictor variable. All variables had a VIF < 3; thus we retained all variables in our models [55, 56]. All GLMMs included kangaroo rat ID and date as random factors. We calculated model fit using conditional R-squared values, and significance of fixed effects was determined using Wald chi-square (χ2) tests [57].
Results
Behavioral classification using accelerometry data
The random forest model at a window size of 6 s that used all summary statistics available in AcceleRater (see methods above) produced the greatest average accuracy, precision, and recall values compared to all other algorithms across all other window sizes (Overall model accuracy = 85.3%; Additional File 1 (Table S4)). The retained model had high accuracy (> 92.5%), precision (> 85%), and recall (> 87.5%) for both behavioral classes motionless and travel. Accuracy (> 87.5%), precision (> 67.5%), and recall (> 73.0%) were somewhat lower for the behavioral classes foraging and grooming. However, when the model incorrectly classified either foraging or grooming, it typically conflated these two behaviors. Thus, when the model incorrectly classified foraging, it classified foraging as grooming 12% of the time and traveling and motionless at 0%; when the model incorrectly classified grooming as foraging at 22%, traveling at 5%, and motionless at 0%. This is likely because grooming and foraging are similar movement patterns. Consequently, our estimation of activity patterns was highly accurate when kangaroo rats were motionless versus traveling as well as when they were engaged in non-travel activity, and moderately accurate at estimating when that non-travel activity was either foraging or grooming.
Kangaroo rat diel activity patterns
By classifying our complete unlabeled free-ranging kangaroo rat accelerometry data set, we confirmed that kangaroo rats are nocturnal, as they spend less time motionless during nighttime light phases (when the sun is below the horizon) compared to daytime light phases (when the sun is above the horizon) and they rarely travel at all during daytime light phases (Fig. 3). Thus, the typical diel activity cycle of a Merriam’s kangaroo rat consisted of long periods of stillness during the day, with very occasional short bouts of activity, followed by long bouts of activity during the night, punctuated by occasional periods of stillness (Fig. 4).

Proportion of time kangaroo rats spent traveling, foraging, motionless and grooming across eight light phases

An archetypal 24-hour travel activity cycle for two individuals (top = female; bottom = male) illustrating long periods of non-travel activity during the day with long bouts of traveling at night
To further understand the proportion of time kangaroo rats spent performing each behavior, we investigated differences across nighttime light phases (i.e., portion of the diel cycle where sun is below the horizon). The proportion of time spent in each behavioral state during different nighttime light phases differed significantly (traveling: χ2 = 397.82, df = 7, P < 0.001, model R2(c) = 0.60; motionless: χ2 = 470.31, df = 7, P < 0.001, model R2(c) = 0.58; foraging: χ2 = 68.34, df = 7, P < 0.001, model R2(c) = 0.47; and grooming χ2 = 267.13, df = 7, P < 0.001, model R2(c) = 0.51 ). Interestingly, kangaroo rats spent significantly more time traveling during night and morning twilight compared to all other nighttime light phases (P < 0.01 for all comparisons). Additionally, kangaroo rats spent more time traveling at dawn compared to all other nighttime light phases except for night and morning twilight (P < 0.01 for all comparisons). Thus, although the sun is below the horizon, kangaroo rats are not traveling as much at the beginning of the night (dusk and evening twilight) compared to the mirrored light phases at the end of the night (morning twilight and dawn). We found a similar pattern when examining motionless behavior in kangaroo rats across light phases. Kangaroo rats spent significantly less time motionless at night and morning twilight compared to all other light phases (P < 0.01 across all comparisons). Moreover, kangaroo rats spent significantly more time motionless during evening twilight and dawn compared to morning twilight and night (P < 0.01 across both comparisons), but significantly less time motionless during evening twilight and dawn compared to morning, day, evening, and dusk (P < 0.01 across all comparison). Similarly, kangaroo rats spent significantly more time foraging during night, morning twilight, and dawn compared to all other light phases (P < 0.01 across all comparisons). Lastly, kangaroo rats spent more time grooming during evening twilight, night, morning twilight, and dawn compared to all other light phases (P < 0.01 across all comparisons). Corroborating other activity patterns, kangaroo rats spent significantly more time grooming at night and morning twilight compared to all other light phases (P < 0.01 across all comparisons).
Behavioral responses to moonlight and environmental variables
We examined if the proportion of time spent motionless, traveling, foraging, and grooming was influenced by moon intensity and moon position. When we removed all nights (n = 4) with a precipitation rate > 0 to account for cloud cover, no relationships changed—thus we retained all values in the dataset. For the proportion of time kangaroo rats spent traveling, we found that moon intensity and moon position had no impact on their own (χ2 = 5.42, df = 2, P = 0.07 and χ2 = 2.01, df = 1, P = 0.16, respectively; model R2(c) = 0.50), but their interaction was significant (χ2 = 13.33, df = 2, P = 0.001, R2(c) = 0.50). Kangaroo rats typically traveled more during medium moon intensity nights and when the moon was visible (Fig. 5A). Moon position (χ2 = 6.34, df = 1, P = 0.01, R2(c) = 0.52) and the interaction between moon position and moon intensity (χ2 = 17.48, df = 2, P < 0.001, R2(c) = 0.52) significantly impacted the proportion of time kangaroo rats spent motionless. When the moon was visible, kangaroo rats spent significantly less time motionless, and they spent less time motionless during a medium moon intensity compared to high and low moon intensities (Fig. 5B). Moon intensity, moon position, and their interaction had no impact on the proportion of time kangaroo rats spent foraging (χ2 = 5.36, df = 2, P = 0.07, χ2 = 0.40, df = 1, P = 0.53, and χ2 = 1.30, df = 2, P = 0.53, respectively; model R2(c) = 0.85; Fig. 5C). Time spent grooming was significantly greater when the moon was visible (χ2 = 11.5575, df = 1, P < 0.01); however, there was no effect of moon intensity and the moon position/moon intensity interaction on the proportion of time spent grooming (χ2 = 2.28, df = 2, P = 0.31, and χ2 = 3.45, df = 2, P = 0.18, respectively; model R2(c) = 0.65; Fig. 5D).

Proportion of time kangaroo rats spent traveling (A), motionless (B), foraging (C), and grooming (D) during different levels of moon intensity (Low < 0.33; Medium = 0.33–0.66; High > 0.66) and the impact of moon position (up/risen or down/set)
Additionally, we examined the proportion of time kangaroo rats spent traveling, motionless, grooming, and foraging by directly comparing each behavior as a function of temperature, relative humidity, wind speed, and moon illumination. We found that kangaroo rats would spend more time foraging as relative humidity increased (χ2 = 4.19, df = 1, P = 0.04, R2(c) = 0.63), but other behaviors were not influenced by humidity. Temperature, wind speed, and moon illumination had no effect on the proportion of time kangaroo rats spend foraging, traveling, grooming, or motionless (Additional File 1; Table S5).
Discussion
By combining low-mass, miniaturized animal-borne accelerometers with radiotelemetry and advanced machine learning techniques, our study showed that accelerometers can be effectively used to quantify behavioral activity of small-bodied (< 50 g), nocturnal, free-ranging mammals. Our work represents the first continuous detailed quantitative description of fine-scale behavioral activity budgets in kangaroo rats, a diverse group of highly abundant rodents that are widely recognized as keystone species in arid regions of North America. After obtaining highly accurate models for ecologically relevant behaviors such as foraging and traveling, we were able to assess behavioral responses to diel light phases, moonlight, ambient air temperature, average windspeed, and average relative humidity. We confirmed that kangaroo rats are highly nocturnal, but are also much more active in some phases of the night cycle than others. Somewhat surprisingly, although we found that behavioral activity cycles are largely independent of several major abiotic factors (most notably moonlight), kangaroo rats did show a significant increase in time spent foraging on more humid nights, an abiotic factor that has received only limited attention in the ecological literature on heteromyid rodents.
Merriam’s kangaroo rats are not lunarphobic
Much of the ecological literature on nocturnal small mammals indicates that greater moon illumination leads to decreased surface activity and foraging behavior as a result of increased predation risk [58,59,60,61]. However, other studies of nocturnal small mammals have indicated both lunarphilia and lunarphobia in regards to foraging/activity patterns, and sometimes no relationship at all [62]. Prugh and Golden’s [62] synthesis on moonlight and predation risk across nocturnal mammals found mixed results, and although the meta-analysis showed that moonlight suppressed activity of nocturnal mammals overall, moonlight effects were not related to trophic level and could be better explained by visual acuity, habitat cover, and phylogenetic relatedness. Mixed results have also been reported for kangaroo rat (Dipodomys spp.) behavioral responses to moonlight (e.g., lunarphobia: [36,37,38, 40]; lunarphilia: [39, 41]; and no influence: [63]). Because previous studies relied on either limited direct observations (i.e., occasional telemetry fixes) or indirect measures (i.e., trapping data and visual encounter surveys), some of the variation could be related to methodological constraints rather than ecological differences.
Our study is the first to use animal-borne biologging devices, which are arguably the most comprehensive and least invasive tools for measuring activity cycles of kangaroo rats, and we found that increased moonlight did not strongly influence the foraging and traveling patterns of D. merriami. However, we noted some lunarphilic tendencies. In particular, we found that kangaroo rats spent more time traveling and less time motionless when the moon was up during intermediate moon phases (i.e., medium moon intensity), indicating a preference for activity during the lighter, rather than darker, periods of these nights (corroborating with evidence from Lockard and Owings [39] and Prugh and Brashares [41]) that kangaroo rat species are somewhat lunarphilic). Our data indicate that kangaroo rats generally accept this tradeoff, increasing surface activity under conditions when their own heightened visual acuity [64] would enhance their ability to avoid predatory attacks despite them being more visible to predators. Interestingly, we also found that kangaroo rats do not reduce overall activity on either full moon or new moon nights, indicating that they are still willing to travel and forage under a number of different light conditions—but if they have the option (i.e., the moon is up for only part of the night), they favor traveling with some moonlight available rather than none.
This pattern is in contrast to the findings of Daly et al. [36], who used periodic fixes of radiotagged animals to show that D. merriami would make compensatory shifts in foraging behavior on nights with higher moon illumination by foraging at dawn. Our accelerometry data shows that individuals were more active at dawn than other light phases of the day (i.e., morning, day, evening, and dusk) regardless of moon phase and that, when variation in moonlight is available (i.e., during medium moon intensity phases), they prefer to travel more when the moon is visible. However, our study took place during the summer period (May-August), whereas Daly et al. [36] analyzed activity patterns in the winter (December-March), and thus further research would be needed to elucidate any potential interactions between seasonality and lunarphobia. The limited data regarding seasonal ecology of D. merriami indicates that this species is generally active throughout the year, but does shift toward a diet consisting mostly of cached seeds in hotter and drier periods when green vegetation is not available [65]—a shift in foraging ecology that could have consequences for how individuals respond to ambient moonlight and predation risk.
Other studies have also concluded that kangaroo rats avoid moonlight, using various methods across different species (i.e., Dipodomys merriami, D. ordii, D. panamintinus, D. microps, [40]; D. spectabilis, [38]; and D. ordii, [37]), including live trapping, recording with feeding stations, and direct observations and counts while driving along dirt and gravel roads. The methods used in these past studies are generally more indirect or less comprehensive than animal-borne accelerometry, which allows for continual quantification of behavioral states across many individuals and days. For example, human observers using radiotelemetry to locate individuals could influence the results of measuring activity by causing kangaroo rats to hide in their burrows when an observer approaches, live-trapping assumes that all animals that are active on the surface are caught, recording with feeding stations provides a sudden and easily accessible food source for kangaroo rats that could alter their motivation for surface activity, and cruising on roads relies on kangaroo rats traveling adjacent to a potentially dangerous loud and fast moving object. However, using accelerometry to quantify traveling and foraging behavior does not provide details of microhabitat use. Although kangaroo rats do not reduce their surface activity with respect to moonlight, they could shift to using more or less vegetative cover during different lighting conditions. Although we cannot address this possibility, future studies using accelerometry may be able to validate more specific categories of kangaroo rat movement, including generating models that could accurately categorize movement in different habitat types. If movement differs consistently among habitat types (for example, slower and more steady movement profiles in heavily vegetated microhabitat compared to moving across open terrain), overall acceleration signatures for movement through these habitats could be accurately classified. The addition of temperature data logged simultaneously with movement by animal-borne devices could also potentially add information about microhabitat use for habitats that have very different temperature profiles (e.g., underground in burrow system versus on the surface).
Merriam’s kangaroo rats increase foraging effort on humid nights
We expected other major abiotic factors, such as temperature, wind speed, and humidity to potentially influence activity cycles of kangaroo rats (we were unable to examine precipitation due to the very limited rain occurring at our study site). Although temperature and wind speed did not influence activity cycles, increased foraging was associated with higher levels of humidity. In general, kangaroo rats are highly adapted to arid, desert-like conditions and are thought to select microclimates that avoid thermal extremes and evaporative water loss [66,67,68]. Past studies investigating the suppression of evaporative water loss in relation to humidity have mainly focused on the differences between burrow and surface humidity, and the relationship of surface activity to surface relative humidity has been largely overlooked (but see Tracy & Walsberg, [69]). Kangaroo rats lose most of their water through cutaneous evaporative water loss [69], and so they may conserve water by prioritizing longer foraging times on nights with higher surface humidity. Future ecological studies of kangaroo rats should more carefully consider the role of ambient humidity in driving patterns of movement and activity.
Merriam’s kangaroo rats are more active prior to dawn than after dusk
Our analyses of acceleration data to determine diel activity patterns of kangaroo rats confirmed that they are exclusively nocturnal. We relocated individuals using radiotelemetry at least once a day during daylight hours, and because we never observed kangaroo rats outside their burrow systems, we assume that the limited daytime movement logged by accelerometry devices largely represents fossorial activity. This corroborates other studies that used direct observation via radio-telemetry and determined almost exclusively nocturnal/crepuscular activity in kangaroo rats ( [70,71,72,73,74]; but see Boal & Giovanni, [33]). Because accelerometers allow for continuous recording of activity and behavior, we were also able to examine much more fine scale activity levels and, in doing so, we found that kangaroo rats are most active during the latter, cooler portions of the night (i.e., the night, morning twilight, and dawn light phases) compared to the beginning and hotter portions of the night (i.e., dusk and evening twilight). Although there is general consensus in the literature that kangaroo rats are almost exclusively nocturnal with few anecdotal accounts of diurnal activity, there is more variability in estimates of activity across different phases of the night cycle [37, 63, 70, 75,76,77].
Previous studies typically used either observations of captive individuals, or periodic telemetry relocations to estimate activity levels. The most robust study on Dipodomys spp. nightly activity patterns to date was done by Langford [75], using direct observation of a focal individual with a red-cellophane covered spotlight throughout the entire night, a time-intensive method that may still have impacted natural behavior from continuous human presence. In contrast, accelerometers in our study continually measured behavior across individuals for up to 6.5 days without the need for direct human presence beyond the collection of a validation dataset.
Our study also provides new insight into the nocturnal activity patterns of Merriam’s kangaroo rat as other studies have shown that D. merriami is primarily active at dusk and only three to four hours into the night in the winter months [78], and nocturnal activity in captivity peaked right after and right before sunrise and sunset [76]. In contrast, we found that D. merriami were more active later in the night during the summer months. A number of factors could explain this shift in activity, including seasonal availability of resources, temperature, and predation risk. Ectothermic nocturnal predators such as rattlesnakes (three species of rattlesnakes, Crotalus atrox, C. viridis, and C. scutulatus were all abundant at our site) are warmer and more active (and thus likely more dangerous) during the early portion of the night when the temperature is highest. Additionally, ectothermic predators are a seasonal impact, as they are typically inactive during the winter months throughout D. merriami’s range (Hanscom and Clark, unpublished data). Additional research could tease apart the interactive influence of temperature, predation, and seasonality on kangaroo rat activity patterns.
Free-ranging validation techniques
A critical step in using animal-borne accelerometry to quantify difficult-to-observe behaviors is to validate with time-matched and paired datasets of behavioral observations and acceleration data. Many methods have been developed to conduct validation studies, including independent captive studies [22, 79], observing animals in zoo settings [80], using surrogate species [81], and observations of free-ranging animals [24]. Captive validation studies and the use of surrogate species often have more limited utility because the behaviors observed may not show the same acceleration signatures as free-ranging individuals [81]. Additionally, validation studies (whether in captivity or not) using direct observations by humans could alter the behavior of the focal individual, leading to a lack of species-typical behavior in the validation set. Here, we developed a method to create validation datasets that overcome some of these critical limitations. We used only free-ranging individuals, and we relocated focal individuals by attaching miniature (< 1 g) radio transmitters to the accelerometry device. Because our initial presence sometimes caused individuals to enter burrow systems, upon relocating focal animals, we moved away from their immediate vicinity or the vicinity of their burrow system and used a telephoto video camera with IR recording capability and an IR flashlight to video record behaviors of kangaroo rats at a distance over 10 m. We found that kangaroo rats largely resumed natural activity after human observers were more than ~ 10 m away and made no loud sounds, suggesting the observers could not be detected by the kangaroo rats. Although we typically observed individuals for a relatively short amount of time (i.e., 5–10 min) using this method, we attained sufficient validation data for training models by accumulating multiple observations across individuals and nights. To our knowledge, this is the first time an attempt has been made to fully validate a large accelerometry dataset using only observations of free-ranging and nocturnal individuals in situ.
Conclusions
Here, we present the first study using accelerometry to quantify behavior on a free-ranging population of a heteromyid rodent, a keystone species in desert areas across North America (Meriam’s kangaroo rat, Dipodomys merriami). We provide a proof-of-concept approach using combined accelerometry/telemetry devices to record validation videos and assemble large acceleration datasets that allowed us to quantify behavioral patterns across the summer active period. Our study represents one of the most robust behavioral datasets of a keystone species that is estimated to be the most abundant vertebrate in many habitats across southwestern North America [26]. We found that, in summer months Merriam’s kangaroo rats are only active during the nighttime phases of the diel cycle (confirming exclusive nocturnal/crepuscular activity) and are particularly active during later light phases of the night (i.e., late night, morning twilight, and dawn). Additionally, we found little indication that basic activity cycles are impacted by moonlight, but kangaroo rats would spend more time foraging on nights with higher surface relative humidity. Prior studies on the behavioral responses of kangaroo rats to moonlight and weather have been mixed within and across taxa, and our study highlights the utility of using accelerometry methods to address these questions with large datasets that quantify behavioral details of multiple individuals across long time scales while minimizing human impact. Thus, we hope that our study will motivate researchers to adapt and apply accelerometry using a greater diversity of species, as the approaches for using this next generation natural history biologging technique continues to be further developed and improved.
Data Availability
The full data sets used and/or analyzed during the current study are available at: Hanscom, Ryan et al. (2023). Cryptic behavior and activity cycles of a small mammal keystone species revealed through accelerometry: a case study of Merriam’s kangaroo rats [Dataset]. Dryad. https://doi.org/10.6086/D1SQ4Z.
Abbreviations
- VHF:
-
Very High Frequency
- IR:
-
Infrared
- GLMM:
-
Generalized linear mixed modeling
References
Clark RW. Diet of the timber rattlesnake, Crotalus horridus. J Herpetol. 2002;36(3):494–9.
Golley FB, Petrusewicz K, Ryszowski L, editors. Small mammals: their productivity and population dynamics. Cambridge University Press; 1975.
Holycross AT. Movements and Natural History of the Prairie Rattlesnake (Crotalus Viridis Viridis) in the Sand Hills of Nebraska (Doctoral dissertation, University of Nebraska at Omaha).
McCain CM, King SR, Szewczyk T, Beck J. Small mammal species richness is directly linked to regional productivity, but decoupled from food resources, abundance, or habitat complexity. J Biogeogr. 2018;45(11):2533–45.
Reinert HK, MacGregor GA, Esch M, Bushar LM, Zappalorti RT. Foraging ecology of timber rattlesnakes, Crotalus horridus. Copeia. 2011;2011(3):430–42.
Rothe-Groleau C. Diet of a Population of Prairie rattlesnakes (Crotalus viridis) in Kansas. J North American Herpetol. 2022;2022(1).
Reif V, Tornberg R. Using time-lapse digital video recording for a nesting study of birds of prey. Eur J Wildl Res. 2006;52:251–8.
Beca G, Valentine LE, Galetti M, Hobbs RJ. Ecosystem roles and conservation status of bioturbator mammals. Mammal Rev. 2022;52(2):192–207.
Davidson AD, Lightfoot DC. Keystone rodent interactions: prairie dogs and kangaroo rats structure the biotic composition of a desertified grassland. Ecography. 2006;29(5):755–65.
Davidson AD, Detling JK, Brown JH. Ecological roles and conservation challenges of social, burrowing, herbivorous mammals in the world’s grasslands. Front Ecol Environ. 2012;10(9):477–86.
Komonen M, Komonen A, Otgonsuren A. Daurian pikas (Ochotona Daurica) and grassland condition in eastern Mongolia. J Zool. 2003;259(3):281–8.
Breed MD, Moore J. Animal behavior. Academic Press; 2021.
Dugatkin LA. Principles of animal behavior. University of Chicago Press; 2020.
Frey S, Fisher JT, Burton AC, Volpe JP. Investigating animal activity patterns and temporal niche partitioning using camera-trap data: challenges and opportunities. Remote Sens Ecol Conserv. 2017;3(3):123–32.
Bateson M, Martin P. Measuring behaviour: an introductory guide. Cambridge university press; 2021.
Burghardt GM, Bartmess-LeVasseur JN, Browning SA, Morrison KE, Stec CL, Zachau CE, Freeberg TM. Perspectives–minimizing observer bias in behavioral studies: a review and recommendations. Ethology. 2012;118(6):511–7.
Brown DD, Kays R, Wikelski M, Wilson R, Klimley AP. Observing the unwatchable through acceleration logging of animal behavior. Anim Biotelem. 2013;1:1–6.
Tosa MI, Dziedzic EH, Appel CL, Urbina J, Massey A, Ruprecht J, Eriksson CE, Dolliver JE, Lesmeister DB, Betts MG, Peres CA. The rapid rise of next-generation natural history. Front Ecol Evol. 2021;9:698131.
Whitford M, Klimley AP. An overview of behavioral, physiological, and environmental sensors used in animal biotelemetry and biologging studies. Anim Biotelem. 2019;7(1):1–24.
Willson MF, Armesto JJ. Is natural history really dead? Toward the rebirth of natural history. Rev Chil Hist Nat. 2006;79(2):279–83.
Hoffman B, Cusimano M, Baglione V, Canestrari D, Chevallier D, DeSantis DL, Jeantet L, Ladds MA, Maekawa T, Mata-Silva V. Moreno-González V. A benchmark for computational analysis of animal behavior, using animal-borne tags. arXiv preprint arXiv:2305.10740. 2023 May 18.
Hammond TT, Springthorpe D, Walsh RE, Berg-Kirkpatrick T. Using accelerometers to remotely and automatically characterize behavior in small animals. J Exp Biol. 2016;219(11):1618–24.
Clermont J, Woodward-Gagné S, Berteaux D. Digging into the behaviour of an active hunting predator: arctic fox prey caching events revealed by accelerometry. Mov Ecol. 2021;9(1):1–2.
Studd EK, Derbyshire RE, Menzies AK, Simms JF, Humphries MM, Murray DL, Boutin S. The purr-fect catch: using accelerometers and audio recorders to document kill rates and hunting behaviour of a small prey specialist. Methods in Ecol Evol. 2021;12(7):1277–87.
Menzies AK, Studd EK, Seguin JL, Derbyshire RE, Murray DL, Boutin S, Humphries MM. Activity, heart rate, and energy expenditure of a cold-climate mesocarnivore, the Canada lynx (Lynx canadensis). Can J Zool. 2022;100:261–72.
Kelt DA. On the relative importance of history and ecology in structuring communities of desert small animals. Ecography. 1999;22(2):123–37.
Hawkins LK, Nicoletto PF. Kangaroo rat burrows structure the spatial organization of ground-dwelling animals in a semiarid grassland. J Arid Environ. 1992;23(2):199–208.
Kerley GI, Whitford WG, Kay FR. Mechanisms for the keystone status of kangaroo rats: graminivory rather than granivory? Oecologia. 1997;111:422–8.
Longland WS, Dimitri LA. Kangaroo rats: ecosystem engineers on western rangelands. Rangelands. 2021;43(2):72–80.
Longland WS, Ostoja SM. Ecosystem services from keystone species: diversionary seeding and seed-caching desert rodents can enhance Indian ricegrass seedling establishment. Restor Ecol. 2013;21(2):285–91.
Cypher BL, Spencer KA. Competitive interactions between coyotes and San Joaquin kit foxes. J Mammal. 1998;79(1):204–14.
White PJ, Ralls K, White CA. Overlap in habitat and food use between coyotes and San Joaquin kit foxes. SW Nat. 1995;40(3):342–9.
Boal CW, Giovanni MD. Raptor predation on Ord’s kangaroo rats: evidence for diurnal activity by a nocturnal rodent. SW Nat. 2007;291–5.
Marti CD, Kochert MN. Diet and Trophic Characteristics of Great Horned Owls in Southwestern Idaho (Dieta Y Caracteristicas Tróficas De Bubo virginianus En El Suroeste De Idaho). J Field Ornithol. 1996;499–506.
Steenhof K, Kochert MN. Dietary responses of three raptor species to changing prey densities in a natural environment. J Anim Ecol. 1988;37–48.
Daly M, Behrends PR, Wilson MI, Jacobs LF. Behavioural modulation of predation risk: moonlight avoidance and crepuscular compensation in a nocturnal desert rodent, Dipodomys merriami. Anim Behav. 1992;44(1):1–9.
Kaufman DW, Kaufman GA. Effect of moonlight on activity and microhabitat use by Ord’s kangaroo rat (Dipodomys ordii). J Mammal. 1982;63(2):309–12.
Lockard RB, Owings DH. Seasonal variation in moonlight avoidance by bannertail kangaroo rats. J Mammal. 1974;55(1):189–93.
Lockard RB, Owings DH. Moon-related surface activity of bannertail (Dipodomys spectabilis) and Fresno (D. Nitratoides) kangaroo rats. Anim Behav. 1974;22(1):262–73.
O’Farrell MJ. Seasonal activity patterns of rodents in a sagebrush community. J Mammal. 1974;55(4):809–23.
Prugh L, Brashares J. Basking in the moonlight? Effect of illumination on capture success of the endangered giant kangaroo rat. J Mammal. 2010;91(5):1205–12.
Shier DM, Swaisgood RR. Fitness costs of neighborhood disruption in translocations of a solitary mammal. Conserv Biol. 2012;26(1):116–23.
Hanscom RJ, DeSantis DL, Hill JL, Marbach T, Sukumaran J, Tipton AF, Thompson ML, Higham TE, Clark RW. How to study a predator that only eats a few meals a year: high-frequency accelerometry to quantify feeding behaviours of rattlesnakes (Crotalus spp). Anim Biotelem. 2023;11(1):20.
Resheff YS, Rotics S, Harel R, Spiegel O, Nathan R. AcceleRater: a web application for supervised learning of behavioral modes from acceleration measurements. Mov Ecol. 2014;2(1):1–7.
Krawczyk B. Learning from imbalanced data: open challenges and future directions. Prog Artif Intell. 2016;5(4):221–32.
Gigliotti LC, Diefenbach DR. Risky behavior and its effect on survival: snowshoe hare behavior under varying moonlight conditions. J Zool. 2018;305(1):27–34.
Johnson MD, De León YL. Effect of an invasive plant and moonlight on rodent foraging behavior in a coastal dune ecosystem. PLoS ONE. 2015;10(2):e0117903.
Studd EK, Boudreau MR, Majchrzak YN, Menzies AK, Peers MJ, Seguin JL, Lavergne SG, Boonstra R, Murray DL, Boutin S, Humphries MM. Use of acceleration and acoustics to classify behavior, generate time budgets, and evaluate responses to moonlight in free-ranging snowshoe hares. Front Ecol Evol. 2019;7:154.
Agafonkin V, Thieurme B. Suncalc: Computer Sun Position, Sunlight Phases, Moon Position and Lunar Phase. R package version 0.3. 2017.
Bowland AE. The effect of wind on small mammal trapping. Lammergeyer. 1987;38:35–9.
Halle S, Stenseth NC, editors. Activity patterns in small mammals: an Ecological Approach; with 11 tables. Springer Science & Business Media; 2000.
Williams CT, Wilsterman K, Kelley AD, Breton AR, Stark H, Humphries MM, McAdam AG, Barnes BM, Boutin S, Buck CL. Light loggers reveal weather-driven changes in the daily activity patterns of arboreal and semifossorial rodents. J Mammal. 2014;95(6):1230–9.
Goodchild MF. Citizens as sensors: the world of volunteered geography. GeoJournal. 2007;69:211–21.
Sieber R. Public participation geographic information systems: a literature review and framework. Ann Assoc Am Geogr. 2006;96(3):491–507.
Zuur AF, Ieno EN, Elphick CS. A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol. 2010;1(1):3–14.
Thompson CG, Kim RS, Aloe AM, Becker BJ. Extracting the variance inflation factor and other multicollinearity diagnostics from typical regression results. Basic Appl Soc Psychol. 2017;39(2):81–90.
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MH, White JS. Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol. 2009;24(3):127–35.
Bowers MA, Dooley JL. Predation hazard and seed removal by small mammals: microhabitat versus patch scale effects. Oecologia. 1993;94:247–54.
Brown JS, Kotler BP, Smith RJ, Wirtz WO. The effects of owl predation on the foraging behavior of heteromyid rodents. Oecologia. 1988;76:408–15.
Kelt DA, Meserve PL, Karina Nabors L, Forister ML, Gutiérrez JR. Foraging ecology of small mammals in semiarid Chile: the interplay of biotic and abiotic effects. Ecology. 2004;85(2):383–97.
Kotler BP, Brown JS, Hasson O. Factors affecting gerbil foraging behavior and rates of owl predation. Ecology. 1991;72(6):2249–60.
Prugh LR, Golden CD. Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J Anim Ecol. 2014;83(2):504–14.
Braun SE. Home range and activity patterns of the giant kangaroo rat, Dipodomys ingens. J Mammal. 1985;66(1):1–2.
McDonald B, Geiger B, Vrla S. Ultraviolet vision in Ord’s kangaroo rat (Dipodomys ordii). J Mammal. 2020;101(5):1257–66.
Nagy KA, Gruchacz MJ. Seasonal water and energy metabolism of the desert-dwelling kangaroo rat (Dipodomys merriami). Physiological Zool. 1994;67(6):1461–78.
Degen AA. Ecophysiology of small desert mammals. Springer Science & Business Media; 2012.
French AR. Physiological ecology of the Heteromyidae: economics of energy and water utilization. Biology of the Heteromyidae. 1993;10:509–38.
Schmidt-Nielsen B, Schmidt-Nielsen K. Evaporative water loss in desert rodents in their natural habitat. Ecology. 1950;31(1):75–85.
Tracy RL, Walsberg GE. Prevalence of cutaneous evaporation in Merriam’s kangaroo rat and its adaptive variation at the subspecific level. J Exp Biol. 2000;203(4):773–81.
Daly M, Behrends PR, Wilson MI. Activity patterns of kangaroo rats—granivores in a desert habitat. Activity Patterns in Small Mammals: An Ecological Approach. 2000:145 – 58.
Garrison TE, Best TL. Dipodomys ordii. Mammalian species. 1990 Apr 26(353):1 – 0.Kenagy GJ. Field observations of male fighting, drumming, and copulation in the Great Basin kangaroo rat, Dipodomys microps. J Mammal. 1976;57(4):781-5.
Kenagy GJ. Field observations of male fighting, drumming, and copulation in the Great Basin kangaroo rat, Dipodomys microps. J Mammal. 1976;57(4):781–5.
Lockard RB. Seasonal change in the activity pattern of Dipodomys spectabilis. J Mammal. 1978;59(3):563–8.
Reichman OJ, Price MV. Ecological aspects of heteromyid foraging. Biology of the Heteromyidae (HH Genoways and JH Brown, eds.). Special Publication. 1993;10:539 – 74.
Langford A. Pattern of nocturnal activity of male Dipodomys ordii (Heteromyidae). The SW Nat. 1983:341–6.
Reynolds HG. Life history notes on Merriam’s kangaroo rat in southern Arizona. J Mammal. 1960;41(1):48–58.
Schroder GD. Foraging behavior and home range utilization of the bannertial kangaroo rat (Dipodomys spectabilis). Ecology. 1979;60(4):657–65.
Behrends P, Daly M, Wilson MI. Aboveground activity of Merriam’s kangaroo rats (Dipodomys merriami) in relation to sex and reproduction. Behaviour. 1986;210 – 26.
Ladds MA, Salton M, Hocking DP, McIntosh RR, Thompson AP, Slip DJ, Harcourt RG. Using accelerometers to develop time-energy budgets of wild fur seals from captive surrogates. PeerJ. 2018;6:e5814.
Pagano AM, Williams TM. Estimating the energy expenditure of free-ranging polar bears using tri‐axial accelerometers: a validation with doubly labeled water. Ecol Evol. 2019;9(7):4210–9.
Dickinson ER, Twining JP, Wilson R, Stephens PA, Westander J, Marks N, Scantlebury DM. Limitations of using surrogates for behaviour classification of accelerometer data: refining methods using random forest models in Caprids. Mov Ecol. 2021;9(1):28.
Acknowledgements
We thank Sebastian Mendoza and Andrew Powers for field assistance. We thank Matthew Bazzi, Taylor Kallmann, Shannon Michel, and Savannah Silva for assistance with video scoring. We thank Greg Crow and the Marathon Grasslands Nature Conservancy staff for housing and access to their property. We thank Carlo Catoni and Technosmart for invaluable assistance with designing and troubleshooting accelerometers used in this study. Lastly, we thank two anonymous reviewers for numerous suggestions that improved the manuscript and the graduate students of Dr. Rulon Clark’s Lab for helpful insight and discussion.
Funding
Funding was provided by the National Science Foundation under grant IOS 1856404 awarded to Rulon Clark, Jeet Sukumaran, and Tim Higham and through the Kaufman Award given by the American Society of Mammalogists to Ryan Hanscom.
Author information
Authors and Affiliations
Contributions
RJH and RWC conceived the ideas and designed the methodology; RJH, RWC, JLH, and CAP collected the data; RJH, TM, and RWC analysed the data; RJH led the writing of the manuscript; RJH, RWC, TEH, and JS provided supervision. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All procedures were approved by the San Diego State University Institutional Animal Care and Use Committee (22-07-008 C) and followed the guidelines of the American Society of Mammalogists for the use of wild animals in research (Sikes et al., 2016). All field work was conducted under Texas Parks & Wildlife permit number SPR-0521-066.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hanscom, R.J., Hill, J.L., Patterson, C. et al. Cryptic behavior and activity cycles of a small mammal keystone species revealed through accelerometry: a case study of Merriam’s kangaroo rats (Dipodomys merriami). Mov Ecol 11, 72 (2023). https://doi.org/10.1186/s40462-023-00433-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-023-00433-x