
Abstract
Background
Despite widespread distribution and occurrence in the global shark fin trade, information regarding fundamental biology of the silvertip shark, especially vertical and horizontal movement data, is sparse. Its habitat-faithful nature, confined geographical populations, and life history suggest silvertip sharks are vulnerable to overexploitation, particularly in heavily longlined regions with limited offshore management, such as the Fijian archipelago.
Results
Satellite telemetry identified diel depth, temperature, and vertical habitat use differences, previously unreported, in a tracked individual. Although the individual occupied shallower mean depths at night, nighttime excursions below 200 m were recorded. Net displacement from tagging location was estimated <18 km.
Conclusion
Diel depth differences and expanded daytime depth use could indicate foraging behavior, routine predator avoidance, or temperature selection. Typically, pelagic species are more heavily impacted by commercial longline fishing than reef-associated species, particularly in regions such as Fiji, where reefs are locally managed. The apparent mix of reef-associated and pelagic behaviors, coupled with high levels of commercial longlining in the region make the silvertip shark especially vulnerable to exploitation. Limited sample size prevented drawing any conclusions about the species, but these preliminary results suggest in order to be effectively managed the silvertip shark warrants additional movement studies and stock assessment surveys throughout it range.
Similar content being viewed by others

Background
Satellite telemetry is a widely used tool to define horizontal and vertical movements of elasmobranch species across a variety of environments [1, 2]. Studies have described critical habitats, redefined management strategies, and gathered important life-history data about these at-risk predators [3, 4]. However, very little information exists about pelagic sharks in the South Pacific despite the region’s exportation of substantial quantities of shark fins to the Hong Kong market. Fiji exported 76,634 kg of dried and frozen fins in 2011, though these estimates include fish not necessarily caught in Fijian waters [5]. Fijian communities and villages are generally not targeting sharks on nearshore reefs as no domestic market exists [6, 7]. Considering the gear types (longlines and gillnets) used by larger vessels to harvest sharks in large quantities, it is logical that fins harvested in Fijian waters are primarily from pelagic species.
The Republic of Fiji is home to several shark species that spend at least part of their life history associated with reef habitats. Whitetip reef (Triaenodon obesus), grey reef (Carcharhinus amblyrhyncos), blacktip reef (Carcharhinus melanopterus), bull (Carcharhinus leucas), and silvertip (Carcharhinus albimarginatus) sharks are commonly observed species on Fiji’s reefs [8, 9]. The former four species are also known to make nearshore slope or pelagic excursions [10, 11]. There are no current studies defining movements of pelagic sharks in the Fijian archipelago. Gathering fundamental ecological information, such as vertical and horizontal movement data obtained through satellite tagging, is the critical first step towards appropriate management of species in this area.
The silvertip shark is a slow-growing requiem shark, with a maximum recorded total length (TL) of 300 cm. Males and females reach maturity between 160–180 and 160–200 cm TL, respectively [11, 12]. Although widely distributed throughout the tropical Indian and Pacific Oceans, populations appear fragmented with minimal dispersion [10]. The silvertip shark inhabits coastal and offshore waters, and individuals are most commonly observed near coral reefs and nearshore reef slopes from the intertidal zone to depths of 600–800 m [10, 11, 14]. Juveniles inhabit shallow nearshore waters, such as lagoons. In contrast, adults occupy a larger range of habitats [10, 14]. Few studies have examined the horizontal movements of silvertip sharks. At Osprey Reef, Australia, silvertips demonstrated both year-round residency to this isolated seamount and unidirectional migrations, briefly being detected on neighboring reefs (~14 km away) [14].
Silvertip sharks are an active species with a diet consisting of benthic and pelagic teleosts, eagle rays, other sharks, and cephalopods [15]. A calculated trophic position of 4.2 traditionally classifies silvertip sharks as an apex predator [15].
The conservation status of silvertip sharks is undecided throughout most of its range, but based on available data, the species is currently Red-listed and classified as ‘Near Threatened’ by the International Union for Conservation of Nature (IUCN) [14]. Globally, silvertips are one of nine prominent shark species landed by high seas longlines and net fleets, either intentionally or as by-catch, and are identified in the Hong Kong fin trade [14, 16]. Reports of severe localized declines in Australia and Chagos, high catch rates in commercial and artisanal fisheries in South Africa, Indonesia, Thailand, Myanmar, Cambodia and the Philippines, coupled with its habitat-faithful nature suggest isolated populations are vulnerable to overexploitation [14, 17, 18]. A 1996 shark survey conducted in the Chagos Archipelago noted severe declines in sightings of silvertip sharks when compared to 1970 findings [18]. Given the species’ susceptibility to line capture, this decline was attributed to fishing pressure [18]. However, an increase in silvertip shark abundance was recorded during a 2006 survey, demonstrating the potential time-scales at which isolated populations can be reduced by over-fishing and show signs of recovery [19]. Though no current population data exist for silvertip sharks within the Fijian archipelago, the broader western Pacific tuna longline fishery reports high levels of silvertip by-catch [20]. Additional data may provide a basis for the reclassification of this species by the IUCN.
The cultural and governance structure of the Fijian islands offers unique protection to nearshore reefs and their associated species (including several shark species), which are locally managed by coastal villages that reside in the immediate area [7]. Community-based management or locally managed marine areas (LMMAs) have created de facto protection for certain reef-associated shark species, whereas offshore waters are, conversely, heavily fished by foreign longlining fleets [20]. It is unclear how much protection the silvertip shark obtains from Fijian LMMAs, as silvertips are suspected to inhabit shelf or pelagic waters during some life stages and/or routine movements [9, 10, 14]. A long-term assessment of shark relative abundance at an established Fijian provisioning site, observed seasonality to silvertip presence, being most abundant between September and December [8].
The lack of basic life-history data, the prevalence of silvertip sharks in the international fin trade [16], and recent discussions by the IUCN to relist the species as “vulnerable” [14] give merit to an ecology-focused satellite tagging study. Understanding water column use and the potential possibility to correlate such findings with fisheries-dependent data will be a vital step in protecting this species. The aim of this study was to describe the silvertip’s short-term depth and temperature use to obtain critical habitat data of this poorly understood species.
Methods
Ethics statement
All research was carried out under the Fijian Fisheries Department research permit (RA15/14) issued by the Ministry of Education, National Heritage, Culture and Arts, and in accordance with Stony Brook University animal care protocols developed within the guidelines of the Institutional Animal Care and Use Committees (IACUC).
Study site
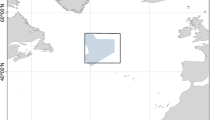
The Great and North Astrolabe Reefs form one of the world’s largest barrier reefs and the second largest barrier reef in the Pacific Ocean. The Great Astrolabe Reef surrounds the Kadavu group of islands including Kadavu, Fiji’s fourth largest island. It is separated from the North Astrolabe Reef by the D’urville Channel roughly 1.5 km at its narrowest (Figure 1). The reefs of Kadavu are in healthy condition with high coral coverage presumably due to the lack of land development. Local chiefs and their communities have actively established ‘tabu’ (no-take) areas throughout Kadavu to manage their fisheries resources [7]. Both the Great and North Astrolabe Reefs offer a broad variety of habitats. Lagoon depths range from very shallow to 50 m and include rock, sand, seagrass, and coral patch reefs. The outer reef drops steeply to 35 m, followed by gentle sediment- and rubble-covered slopes to a depth >65 m. Beyond this, the reef wall drops more than 1,600 m to the ocean floor. The North Astrolabe Reef is a circular atoll barrier reef with multiple reef channels connecting the open ocean to the lagoon.

Field techniques
Tagging was conducted on April 7, 2014. The shark was attracted to the research vessel using a chum crate containing pieces of escolar (Lepidocybium flavobrunneum), oilfish (Ruvettus pretiosus) and yellowfin tuna (Thunnus albacares) heads. The shark was captured using baited handlines, see [1] for a detailed description of capture and handling techniques.
Tag details
This study used one Standard Rate X-Tag (Microwave Telemetry, Inc., Columbia, MD, USA). X-Tags are 12 × 3.2 cm at the maximum diameter with an 18.5 cm antenna and weigh approximately 40 g. Though an X-Tag archives high-resolution 2-min depth, temperature, and light-level data in memory, the tag only transmits a subset of data through the Argos system. For deployments less than approximately 4 months, time-series depth and temperature records collected at 15-min intervals are transmitted. In addition, the transmitted dataset includes daily minimum and maximum depth and temperature records and daily geolocation estimates, with a best-possible error of ±1° for latitude and ±0.5° for longitude [21].
The X-Tag was programmed to release from the animal after 8 months. An X-Tag may release prematurely due to the constant pressure release mechanism, emergency depth release mechanism, or attachment failure. Specifically, this tag was programmed to release due to the constant pressure release mechanism if the tag remained in a 3-m depth band for 1 day. When the release mechanism is initiated, a current is applied to a corrodible link connecting the tether to the X-Tag. Once the link is corroded, the X-Tag is free to float to the surface where transmitted data can reach the Argos satellite system [21].
X-Tags implement data compression techniques which may cause a depth and temperature record to be identified as a ‘delta limited’ value. A depth record marked as a delta limited dive (ascent) may be deeper (or shallower) than indicated. Similarly, a temperature record marked as a delta limited increase (or decrease) may be warmer (or cooler) than indicated. All delta limited values were included in the analyses [21].
Depth and temperature utilization
Daily sunrise and sunset times were calculated from the deployment location. Dawn was defined as 1 h before and after sunrise, and dusk was defined as 1 h before and after sunset. The assignment of diel period allowed for day–night comparisons. Daily diurnal and nocturnal mean depths, mean temperatures, and mean absolute differences between consecutive 15-min depth records were compared with two-tailed t tests with the Welch approximation for unequal variances between groups [22]. The first day period (immediately after tag deployment) and the last night period (immediately prior to tag detachment) were removed from the diel comparisons.
Bathymetry data were obtained from GEBCO gridded bathymetric datasets [23]. Pearson’s product moment correlation was used for all reported correlations. R 2.15.3 (32-bit) was used for all analyses [24], and we specified a significance level of 0.05 for all statistical analyses.
Results
On April 7, 2014, one X-Tag was deployed on an immature female silvertip measuring 155 cm (TL) near Kadavu Island, Fiji (18.698S, 178.530E). The tag and tether prematurely detached from the shark on April 18, 2014, presumably due to attachment failure. After floating at the surface for 1 day, the tag released from the attached tether due to the constant pressure release mechanism and began transmitting on April 19, 2014. The first Argos location (18.556S, 178.442E) was 18.37 km NW from the deployment location (Figure 1). The tag was deployed at a location with bottom depth of approximately 48 m and the first Argos location had a bottom depth of approximately 1,950 m. All (100%) of the transmitted dataset was received through the Argos system providing a total of 2,182 combined depth and temperature records on 15-min intervals and 13 daily position estimates. Five delta limited depth records and seven delta limited temperature records were identified in the dataset.
The silvertip occupied depths between 0 and 381.9 m with a mean (±SD) of 59.9 ± 38.5 m. The silvertip spent the most time (97.0%) in the upper 150 m of the water column and over half of its time (52.9%) above 50 m. The animal spent minimal time near surface waters (3.1% of time in upper 10 m) (Figure 2). This individual recorded a mean temperature of 26.30 ± 1.71°C, and temperatures ranged from 14.16 to 28.57°C. Most temperature records (99.5%) fell between 22 and 29°C, and over half of the records (67.5%) fell between 26 and 29°C. The tag recorded deeper and colder records as the deployment progressed (maximum depth: r 2 = 0.563, P = 0.004913; minimum temperature: r 2 = 0.533, P = 0.006986) (Figure 2).

Depth profile. The silvertip shark’s time-series depth profile colored by diel period with daily minimum and maximum depth records indicated.
The daily daytime mean depths were significantly deeper than the daily nighttime mean depths (t 12.491 = 4.8537, P = 0.000353, Figure 3). Similarly, the daily daytime mean temperatures were significantly cooler than the daily nighttime mean temperatures (t 11.627 = 5.9908, P = 7.19 × 10−5, Figure 4). There was larger variation between consecutive depth records during the day period than the night period (t 17.554 = 4.8456, P = 0.0001389).

Depth distribution. Frequency distribution of the silvertip shark’s diurnal and nocturnal vertical water column use.

Temperature distribution. Frequency distribution of the silvertip shark’s diurnal and nocturnal temperature use.
The three deepest 15-min records (215.2, 220.6, and 199 m) were recorded during the night and were delta limited descents. The daily maximum depth values from these 3 days extend to 263.6, 360.4, and 381.9 m, respectively (Figure 2). There were no consecutive depth records below 200 m, indicating that deep-water excursions were less than 30 min in duration.
Discussion
Tracking a silvertip shark via PSAT provided previously unknown depth, temperature, and habitat information. Daytime and nighttime depth and temperature differences were identified. The shark inhabited deeper and cooler waters during the day. Consecutive daytime depth records exhibited greater variation compared to nighttime depth records, indicating expanded diurnal vertical habitat use. The silvertip’s diel vertical migration (DVM) to deeper daytime depths mimics that of multiple coastal, pelagic and reef-associated elasmobranchs [26–27]. In Palau, grey reef sharks exhibiting a crepuscular pattern of DVM by occupying shallowest depths (<30 m) at dawn and dusk was linked to foraging on shallow reefs [26]. Alternatively, grey reef sharks in Australia exhibited reverse DVM patterns (remaining shallower during the day, but not specifically dawn or dusk) to those tracked in Palau. In both studies, DVM patterns were attributed to foraging [28]. Grey reef sharks have shown to expand horizontal range at night; therefore, these intraspecific differences in DVM patterns may be explained by site-specific bathymetry and/or differences in prey behavior [14, 28]. Furthermore, juvenile white sharks (Carcharodon carcharias) demonstrated similar patterns of deeper daytime depths coupled with increased activity levels, suggesting daytime foraging behavior [27]. Silvertip diet has shown to contain ~13% cephalopods [15], therefore increased daytime vertical habitat use may reflect foraging on species which undergo DVM and seek daytime refuge at depth, such as cephalopods. Furthermore, the DVM pattern may be linked to temperature selection as well as foraging behavior. Small-spotted catsharks (Scyliorhinus canicula) exhibited nocturnal DVMs to warmer, shallower, and prey-rich depths with increased nighttime activity. Laboratory experiments determined this strategy of foraging in warm water and resting in colder water maximized bioenergetic efficiency [29].
Besides foraging behavior, the silvertip’s DVM may be attributed to predator avoidance, similar in function to those observed in juvenile Caribbean reef sharks in Belize. Large Caribbean reef sharks (>110 cm TL) perform DVM presumably for foraging on nocturnally vulnerable prey species (e.g., parrotfish) [26]. Meanwhile, at night, juvenile Caribbean reef sharks moved into shallow waters of the lagoon from the deeper fore-reef to reduce encounters with larger conspecifics [26]. In Fiji, larger conspecifics and other large-bodied shark species such as scalloped hammerhead (Sphyrna lewini), shortfin mako (Isurus oxyrinchus), tiger (Galeocerdo cuvier), and bull (C. leucas) sharks are potential predators of a juvenile silvertip shark.
The tagged animal spent most of its time in the epipelagic zone, although at least four deep-water excursions (>200 m) were identified in the daily maximum depth records. The maximum recorded depth was 381.9 m, but silvertip sharks have previously been recorded to maximum depths between 600 and 800 m [10]. The silvertip did not spend an extended duration (>30 min) at mesopelagic depths, suggesting that the deep dives were short excursions. Though the precise function is unknown, dives would likely be longer in duration if serving the purpose of predator avoidance [30]. The animal recorded increasing depth records as the deployment progressed, potentially corresponding to movement off the shelf and away from the tagging location. Although the tracked individual was tagged on the reef, the deep depth records corresponded to entering pelagic waters. Pelagic species are more likely to encounter commercial longline fishing than reef-associated species, particularly in regions with LMMAs such as Fiji. Commercial tuna longlines are frequently deployed at the surface overnight, increasing the likelihood silvertips will encounter longline gear. The species’ apparent inquisitive disposition (compared with other shark species) make it particularly susceptible to fishing mortality [18, 19].
The fine-scale movements of the silvertip could not be resolved by the tag’s light-based geolocations because the net displacement (<18 km) is considerably less than the best error rate of the X-Tag’s light-based geolocations. The tag detached from the shark due to attachment failure, and subsequently, the tag floated on the surface for 28 h before the first Argos location was estimated. Given that the tag drifted approximately 14 km during the first day of transmission, the tag may have drifted a comparable distance prior to transmission. Consequently, the first Argos location was not likely representative of the shark’s position. It was not expected that an animal this size would travel off the shelf to waters nearly 2,000 m deep given its size and presumed reef association. Site residency is common to many reef-associated shark species, especially in juveniles [14, 26, 31]. However, the deep dives recorded in situ support the use of deeper waters.
Residency to specific locales has been observed in silvertips [14]. Individuals have been repeatedly identified at shark provisioning sites even if not the targeted species of dive operators (B. Paige, pers comm.). Shark provisioning sites are increasing in Fiji as a result of a growing ecotourism sector. Provisioning sharks and the subsequent effect on their behavior is widely debated but must be considered when discussing a species’ movements. Our study area had one commercial dive operator which did not offer a shark feeding experience. Since there are no routine provisioning sites near the study site, the observed behaviors are considered connatural.
Conclusion
The habitat-faithful nature of silvertip shark, confined geographical populations, and life history suggest it is highly vulnerable to overexploitation [14]. Contemporary reports of localized extirpation throughout parts of its range highlight the need for further study of its geographically and genetically distinct populations. Though the data presented here describe a single individual, this pilot study can serve as a model for future silvertip tracking studies. Given the limited 12-day dataset, it is not appropriate to draw conclusions about this species. In retrospect, attaching the PSAT with a first dorsal bridal technique instead of an umbrella dart could have increased tag retention, as has been verified on reef-associated animals in the Caribbean (E. J. Brooks, pers comm.). However, the absence of fundamental data available for this species, especially in this region, gives the results merit. We identified diel differences, previously unreported, in the tracked individual’s vertical use of the water column and temperature use. The deeper depth distribution and increased vertical habitat observed during the day may suggest foraging behavior, but additional studies addressing diet are needed to identify prey composition. Of additional interest, the increased use of the upper water column at night could indicate predator avoidance or particular temperature selection. The silvertip shark’s apparent mix of reef-associated and pelagic behaviors, coupled with anecdotal reports that commercial longliners are increasingly deploying longlines in close proximity to reef systems during the night when local Fijians are less likely to observe or intervene, make silvertips especially vulnerable. Given high levels of intentional or unintentional landings from tuna longlines, specifically, within the Fijian archipelago, this species warrants additional movement studies and stock assessment surveys throughout its range.
References
Howey-Jordan LA, Brooks EJ, Abercrombie DL, Jordan LKB, Brooks A, Williams S et al (2013) Complex movements, philopatry and expanded depth range of a severely threatened pelagic shark, the oceanic whitetip (Carcharhinus longimanus) in the Western North Atlantic. PLoS One 8(2):e56588. doi:10.1371/journal.pone.005658
Weng KC, Castilho PC, Morrissette JM, Landeira-Fernandez AM, Holts DB, Schallert RJ et al. (2005) Satellite tagging and cardiac physiology reveal niche expansion in salmon sharks. Science 310(5745):104–106
Ferretti F, Worm B, Britten GL, Heithaus MR, Lotze HK (2010) Patterns and ecosystem consequences of shark declines in the ocean. Ecol Lett 13(8):1055–1071
Dulvy NK, Fowler SL, Musick JA, Cavanagh RD, Kyne PM, Harrison LR et al (2014) Extinction risk and conservation of the world’s sharks and rays. Elife 3:e00590
Census and Statistic Department (CSD) (2015) Aquaculture fisheries statistics. The Government of the Hong Kong Special Administrative Region
Cooke AJ, Polunin NV, Moce K (2000) Comparative assessment of stakeholder management in traditional Fijian fishing-grounds. Environ Conserv 27(03):291–299
Jupiter SD Cohen PJ, Weeks R, Tawake A, Govan H (2014) Locally-managed marine areas: multiple objectives and diverse strategies. Pac Conserv Biol 20(2):165–179
Brunnschweiler JM, Abrantes KG, Barnett A (2014) Long-term changes in species composition and relative abundances of sharks at a provisioning site. PLoS One 9(1):e86682
Goetze J, Fullwood LAF (2012) Fiji’s largest marine reserve benefits reef sharks. Coral Reefs 32(1):121–125
Compagno LJV, Dando M, Fowler SL (2005) A field guide to the sharks of the world. HarperCollins, UK
Last PR, Stevens JD (2009) Sharks and rays of Australia, 2nd edn. Harvard University Press
Randall JE, Allen GR, Steene RC (1997) Fishes of the great barrier reef and coral sea. University of Hawaii Press, Honolulu
Barnett A, Abrantes KG, Seymour J, Fitzpatrick R (2012) Residency and spatial use by reef sharks of an isolated seamount and its implications for conservation. PLoS One 7(5):e36574
Pillans R, Medina E, Dulvy NK (2009) Carcharhinus albimarginatus. The IUCN red list of threatened species, version 2015.2. http://www.iucnredlist.org. Accessed 5 Mar 2015
Cortés E (1999) Standardized diet compositions and trophic levels of sharks. ICES J Mar Sci J du Conseil 56(5):707–717
Clarke SC, Magnussen JE, Abercrombie DL, McAllister MK, Shivji MS (2006) Identification of shark species composition and proportion in the Hong Kong shark fin market based on molecular genetics and trade records. Conserv Biol 20(1):201–211
Meekan M, Cappo M (2004) Non-destructive techniques for rapid assessment of shark abundance in northern Australia. Australian Institute of Marine Science, Townsville, Queensland, Australia
Anderson C, Sheppard C, Spalding M, Crosby R (1998) Shortage of sharks at Chagos. Shark News 10:1–3
Graham NA, Spalding MD, Sheppard CR (2010) Reef shark declines in remote atolls highlight the need for multi-faceted conservation action. Aquat Conserv Mar Freshw Ecosyst 20(5):543–548
Gilman E, Clarke S, Brothers N, Alfaro-Shigueto J, Mandelman J, Mangel J(2007) Shark depredation and unwanted bycatch in pelagic longline fisheries: industry practices and attitudes, and shark avoidance strategies. Western Pacific Regional Fishery Management Council, Honolulu, Hawaii. ISBN 1-934061-06-9
MicrowaveTelemetry (2015) MicrowaveTelemetry, Columbia, MD. http://www.microwavetelemetry.com/fish. Accessed 15 Dec 2014
Zar HJ (1996) Biostatistical analysis, 3rd edn. Prentice Hall, Upper Saddle River, New Jersey, 07458, London
GEBCO, TOMG (2015) General bathymetric charts of the oceans (GEBCO), TOMG, version 2.0. http://www.gebco.net. Accessed 10 June 2015
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. URL http://www.R-project.org/. 2010
Chapman DD, Pikitch EK, Babcock EA, Shivji MS (2007) Deep-diving and diel changes in vertical habitat use by Caribbean reef sharks Carcharhinus perezi. Mar Ecol Prog Ser 344:271–275
Vianna GM, Meekan MG, Meeuwig JJ, Speed CW (2013) Environmental influences on patterns of vertical movement and site fidelity of grey reef sharks (Carcharhinus amblyrhynchos) at aggregation sites. PLoS One 8(4):e60331
Weng KC, O'Sullivan JB, Lowe CG, Winkler CE, Dewar H, Block BA (2007) Movements, behavior and habitat preferences of juvenile white sharks Carcharodon carcharias in the eastern Pacific. Mar Ecol Prog Ser 338:211–224
Heupel M, Simpfendorfer C (2015) Long-term movement patterns of a coral reef predator. Coral Reefs 34(2):679–691
Sims DW, Wearmouth VJ, Southall EJ, Hill JM, Moore P, Rawlinson K et al. (2006) Hunt warm, rest cool: bioenergetic strategy underlying diel vertical migration of a benthic shark. J Anim Ecol 75(1):176–190
Wirsing AJ, Heithaus MR, Frid A, Dill LM (2008) Seascapes of fear: evaluating sublethal predator effects experienced and generated by marine mammals. Mar Mamm Sci 24(1):1–15
Bond ME, Babcock EA, Pikitch EK, Abercrombie DL, Lamb NF, Chapman DD (2012) Reef sharks exhibit site-fidelity and higher relative abundance in marine reserves on the Mesoamerican barrier reef. PLoS One 7(3):e32983. doi:10.1371/journal.pone.0032983
Google earth V 7.1.5.1557 (May 20, F.S., 179.138394W, Eye alt 741.95 km. SIO, NOAA, U.S. Navy, NGA, GEBCO. Landsat). http://www.earth.google.com. June 17, 2015
Google earth V 7.1.5.1557 (May 20, G.A.R.S., 178.520745 W, Eye alt 50.79 km. LDEO-Columbia, NSF, NOAA, SIO, U.S. Navy, NGA, GEBCO. Landsat). http://www.earth.google.com. June 17, 2015
Authors’ contributions
MEB, LAH and SM participated in the fieldwork. SM facilitated the before expedition site research and attaining of permits. ET participated in the design of the study and performed the statistical analysis. MEB and LAH conceived of the study, and participated in its design and MEB, LAH and SM participated in its coordination. MEB, ET, SM and LAH drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We are grateful for the generous support of the Waitt Foundation and the Moore Charitable Foundation without which this cruise would not have been possible. Demian Chapman, Lance Jordan, and the three anonymous reviewers and associate editor are thanked for comments that greatly improved the content of the case report. Thank you to Trevor Bacon, Sean Williams, Jessica Cramp, Tina Weier, Joey LePore, Andy Mann, the volunteers of Projects Abroad Fiji, Ingrid Sprake, Andy Hill, Wildlife Conservation Society, and for essential logistical and field support, Captain Richard Grissell along with the crew of the Waitt Foundation-provided research vessel. Chris Fallows (hhtp://www.apexpredators.com) is thanked for providing the photograph used for the article cover. We also thank Vicki Devlin for her aid in graphics production.
Compliance with ethical guidelines
Competing interests The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bond, M.E., Tolentino, E., Mangubhai, S. et al. Vertical and horizontal movements of a silvertip shark (Carcharhinus albimarginatus) in the Fijian archipelago. Anim Biotelemetry 3, 19 (2015). https://doi.org/10.1186/s40317-015-0055-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-015-0055-6