
Abstract
Background
Bulinus globosus, the main intermediate snail host of Schistosoma haematobium. The increased contacts between Africa and China could even lead to large-scale dissemination of B. globosus in China. Temperature is the key factor affecting fresh-water snail transmission. This study predicted potential risk of colonization of B. globosus in the mainland of China under climate change.
Methods
We investigated minimum and maximum temperatures for B. globosus eggs, juveniles and adult snails kept under laboratory conditions to find the most suitable range by pinpointing the median effective temperatures (ET50). We also assessed the influence of temperature on spawning and estimated the accumulated temperature (AT). The average air temperatures between 1955 and 2019 in January and July, the coldest and hottest months in China, respectively, were collected from national meteorological monitoring stations and investigated in a geographic information system (GIS) using empirical Bayesian Kriging to evaluate the theoretical possibility for distribution of B. globosus in southern China based on temperature.
Results
The effective minimum temperature (ET50min) for eggs, juveniles, adult snails and spawning were 8.5, 7.0, 7.0, 14.9 °C, respectively, with the corresponding maximum values (ET50max) of 36.6, 40.5, 40.2 and 38.1 °C. The AT was calculated at 712.1 ± 64.9 °C·d. In 1955, the potential B. globosus distribution would have had a northern boundary stretching from the coastal areas of Guangdong Province and Guangxi Autonomous Region to southern Yunnan Province. Since then, this line has gradually moved northward.
Conclusions
Annual regeneration of B. globosus can be supported by the current climate conditions in the mainland of China, and a gradual expansion trend from south to north is shown in the study from 2015 to 2019. Thus, there is a potential risk of colonization of B. globosus in the mainland of China under climate change.
Graphical Abstract

Similar content being viewed by others

Background
Schistosomiasis is a parasitic disease with a life cycle that involves an intermediate snail host. It endangers human health and affects social and economic development in 78 countries and regions in Africa, Asia, South America and the Middle East. About 230 million people are infected with more than 800 million at risk [1,2,3]. Five schistosome species: Schistosoma japonicum, S. mansoni, S. haematobium, S. intercalatum and S. mekongi and a few subgroups can parasitize humans, with the three former species being the most common [4]; S. mansoni is the only species in Latin America, while S. mansoni and S. haematobium dominate in Africa. S. japonicum is the only species in China and the Philippines and some minor pockets in Indonesia; while Cambodia and Laos share a limited endemic area of S. mekongi [5]. All species except S. haematobium cause intestinal symptoms, while the latter produces urogenital symptoms, including bladder cancer [6, 7]. No locally infected schistosomiasis haematobia cases exist in Southeast Asia but imported cases do, particularly in China [8].
Bulinus (Mollusca, Gastropoda, Pulmonata, Basommatophora, Planorbidae) is a hermaphroditic group of freshwater snails of about 30 species that all can transmit S. haematobium [9]. These snails are widely distributed in Africa and countries bordering the Indian and the Mediterranean Seas. According to the shape of the shell and some characteristics of the organism, four main groups of Bulinus species have been named: B. africanus, B. forskalii, B. reticulatus and the B. truncatus/tropicus complex. B. globosus, the main intermediate host of S. haematobium, belongs to the B. africanus group.
Temperature is the key factor governing the distribution of snails, because there are other main factors such as light that affecting snails [10, 11]. According to the 6th Assessment Report (https://www.ipcc.ch/assessment-report/ar6/) of the Intergovernmental Panel on Climate Change (IPCC), the global warming trend is intensifying, which means that it might affect the B. globosus distribution sooner than previously thought, a fact that has attracted widespread attention from scholars around the world [12, 13]. China is the largest prevalent area of schistosomiasis japonica and, so far, no S. haematobium intermediate snail host has been found there [14]. However, with the global warming and the increasing frequency of international exchange, especially that related to advancing the “Belt and Road” initiative, trade between China and Africa has become grown, and many local African aquatic plants and snails can be brought back to China or purchased through the Internet. Many invasive species survive and multiply since individuals first kept at home in their aquariums only to later discard them in the natural environment. These developments have led to an increased risk of the introduction of B. globosus into China.
Empirical Bayesian Kriging (EBK) is a fast and reliable solution for both automatic and interactive data interpolation. It can be used for interpolation of very large datasets, which has resolved the drawbacks of classical geostatistical models [15]. In recent years, EBK has been applied in numerous applied research papers to predict changes in space and climate, for example Kutuzov [16], Nocco [17] and Nogueira [18]. In this research, we used the statistical software of Geostatistical Analysis extension of EBK Regression Prediction of ArcGIS 10.7 (ESRI) to generate maps both for indicating snail survival and for predicting the B. globosus colonization risk.
The aim of this study was to investigate the temperature levels required by the different stages of B. globosus snails (eggs, juveniles and adults), as well as for spawning, and to combine these data with those delivered by the national meteorological monitoring stations (MMSs) in southern China. The study is expected to provide a model that can also be used for monitoring the invasion risk of other snails in other countries (Fig. 1).

The flow chart of this study process
Methods
Snail breeding
B. globosus snails, obtained from the laboratory of Sino-African Cooperation for Schistosomiasis Control in Zanzibar, were collected from an on-site pond on Pemba Island where they had been seeded and kept for 3 years. The snails received daylight at 1,000 lx (General environmental experiment box, 12 h light/12 h darkness photoperiod), while kept at 25 °C in 35 × 24 × 20 cm 2-L containers filled with dechlorinated tap water and fed with fresh lettuce every three days. For snail egg collection, several 5 × 5 cm non-toxic plastic films were kept in suspension on the water surface of the breeding containers to act as receptacles. The plastic films were observed daily and when eggs were found, they were scrutinized microscopically and those with intact internal structures were selected for testing. In the experiments, the snail eggs hatched within 2–3 weeks to become juvenile snails that grew into adult snails after another 5–7 weeks.
Instruments and reagents
The experimental equipment included an MLR-352-PC general environmental experiment box (Panasonic, Osaka, Japan), an ICC50 HD microscope (Leica Camera AG, Wetzlar, Germany), a PolySci 9102 precision temperature water bath (PolyScience, Niles, IL, USA), a MIR-153 biological culture box (Sanyo, Tokyo, Japan), a SB-988 aeration pump (Sobo, Zhongshan Guangdong China). A personal computer equipped with ArcGIS software, v.10.7 (ESRI, Redlands, CA, USA) for geographical information systems (GIS) studies. Dechlorinated water was prepared by adding 0.1 g/L of sodium thiosulfate (Shanghai Aibi Chemical Reagent Co. Shanghai, China) to 50 L of fresh tap water in a plastic tank, stirred until dissolved and left for 12 h before use.
Temperature thresholds for different developmental stages
As the whole scale of temperatures would not be required for testing, we investigated two limited ranges: one with lower temperatures and another with higher ones to find the thresholds restricting survival. We wished to pinpoint the low and high median effective temperatures (ET50min and ET50max) for each stage of snail development (eggs, juveniles and adult snails), i.e., the thresholds at which only 50% of the number of organisms under investigation could survive (or where 50% spawning took place). Each group was set up with 3 replicates, then the mean value of each group is obtained for analysis.
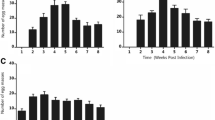
For each group 20‒70 newly deposited snail eggs from aggregated egg collections were investigated by temperature stress testing. The temperature stress testing of snail eggs were divided into 21groups and subjected to different temperatures in experimental water baths containing 10 L of dechlorinated water. The nine low-temperature groups were tested at one-integer increases of temperature from 4.0 °C to 12.0 °C, while the 12 high-temperature groups were tested at temperatures from 34.0 °C to 45.0 °C using the same integer increases. Eggs exposed in different temperatures for 72 h, and then transferred and incubated in a breeding container at 25 °C and observed daily. The hatched eggs in the following 15-day period were counted and those failing to hatch were discarded. The outcome of this experiment revealed the upper and lower thresholds of the range of suitable hatching temperatures. Since the relationship between the mortality rate of the B. globosus and temperature is non-linear [19], we used the statistical software SPSS 20.0 (International Business Machines Corporation) to establish the graph (Fig. 2), which is described by the equations governing the changing hatching rate along the change of water temperatures [20, 21]. This gave the ET50min and ET50max for the snail eggs.

Testing curves of the egg mortality rates at different temperatures
The procedure described above for snail eggs was repeated with 10 juvenile and 10 adult snails in the low-temperature and high-temperature groups with the difference that they were transferred and subjected to 25 °C incubation after 24 h and checked for survival after 72 h using percussion to their head-foot to determine whether they were alive or dead. The ET50min was defined as the minimum water temperature at which the mortality rate reached 50%. The ET50max was defined as the maximum water temperature at which the mortality rate reached 50% (Figs. 3 and 4).

Testing curves of the juvenile mortality rates at different temperatures

Testing curves of the adult mortality rates at different temperatures
Again, 16 groups with different temperatures were compared as described above. This time, however, the water temperature was set to vary from 6.0 °C to 22.0 °C in the water baths in the low-temperature group (6.0, 8.0, 10.0, 12.0, 14.0, 15, 16.0, 18.0, 20.0 and 22.0 °C). The high-temperature group was subjected to vary from 30.0 °C and ending at 40.0 °C (30.0, 32.0, 34.0, 36.0, 38.0 and 40.0 °C). We used three beakers for each temperature group with one adult snail in each, with the snails bred according to the conventional method described above. We observed egg production for each temperature in each beaker and counted the number of eggs over a total of 15 days. Spawning curve equations of the low-temperature group and high-temperature group snails varying with water temperature were established, and the snail spawning extreme at low temperatures and high temperatures were calculated (Fig. 5).

Testing curves of the spawning at different temperatures
Accumulated temperatures
To find the temperature needed to keep an unbroken line of snail generations, there was set up with 3 batches at the same time. Between 50 and 100 snail eggs of each batch were bred indoors using terrariums with ample access to water at prevailing ambient temperatures. We observed the whole chain of snail development (eggs, hatching, juveniles, adult snails and spawning) from a temperature–time point of view. The accumulated temperature (AT) from egg deposition to spawning was calculated using the formula:
where TDA is the daily average temperature and C the threshold temperature of B. globosus. AT is the product of the prevailing temperature and the number of days counted from egg deposition to the first spawning by the snail originating from the egg (°C d). For a full snail generation to be completed, TDA must clearly exceed the threshold temperature C.
Calculation of the colonization risk
In order to find the theoretical limits of B. globosus in southern China, we obtained a set of meteorological information from the MMSs available from the China Natural Resources Database (http://data.cma.cn/) that contains an inventory of temperatures covering the whole country. We selected the daily average temperatures for the coldest month (January) and the hottest (July) covering the period 1955–2019 from 257 MMSs in the 11 provincial-level administration divisions (PLADs)—Chongqing, Zhejiang, Yunnan, Sichuan, Guizhou, Jiangxi, Hunan, Hainan, Guangdong, Fujian and Guangxi Autonomous Region and compared this information with the 72-h ET50 snail values obtained in the laboratory. The combination of these temperatures would tell us whether or not B. globosus would survive to produce consecutive snail generations in China.
We followed the temperatures every ten years to find out which years had recorded temperatures that would have allowed several snail generations. Naturally, we did not get the read-outs from the same number of MMSs each time since their number changed over the long time covered, and the temperature data for the years chosen for the study were not always available. In the years 1955, 1965, 1975, 1985, 1995, 2005, 2015 and 2019, we obtained data from 155, 233, 236, 236, 228, 228, 226 and 197 MMSs, respectively.
As the relationship between temperature and snail mortality [19] could not be expected to be linear, we used SPSS 20.0 software for a variable, non-linear regression analysis based on the T50 measurements discovered to obtain the B. globosus colonization risk. We calculated the potential, annual generation number (N) of B. globosus in the various areas by the formula:
where TDA is the local, average temperature; C the starting temperature of snail development (the time of egg deposition); and AT the accumulated temperature required for snail development to be completed. As for Eq. 1, TDA must clearly exceed the threshold temperature for development to occur. Even so, outcomes of N < 1 indicate that snails cannot reproduce in the area in question.
Application of GIS
We used ArcGIS 10.7 software to establish a spatial database of MMS distribution based on geographical coordinates, imported data for survival, reproduction and other attributes as base values for EBK that would generate maps indicating snail survival as well as maps predicting the B. globosus colonization risk. The MMSs were established from the founding of the People’s Republic of China in 1949, the data from 1951 to 2019 can be downloaded from the Internet. We generated maps for every 10th year of the study period covering 1955 to 2019 [22]. Superimposing geographical temperature maps of southern China with those depicting the distribution of the temperature thresholds required for survival of the different snail stages, including those needed to allow new snail generations to bridge the year-to-year gap, visualized the changing colonization risk over time, similar to what has been previously shown for agricultural pests in southern China [23, 24].
Results
Developmental thresholds of snail
In the 15-day period after 72 h at various temperatures, the egg survival in the low-temperature group was seen to improve along the temperature increases tested. From the 100% mortality rate at 7.0 °C, it reached none (0%) at 11.0 °C with the equation indicating that ET50min were passed at 8.5 °C (Table 1). In the higher end, survival was better than at lower temperatures with 0% mortality rate at 34.0 °C, 100% mortality rate at 40.0 °C and ET50max at 36.6 °C.
The outcomes with respect to the juvenile snails followed this pattern but the survival temperatures were quite different with 100% mortality at 5.0 °C and 0% reached at 8.0 °C with the equation indicating an ET50min of 7.0 °C. At the higher end, the mortality rate was 100%, both at 43.0 °C and 44.0 °C and the ET50max was as high as 40.5 °C. The ET50min and ET50max values of adult snails at 7.0 °C and 40.2 °C, respectively, were very close to those of the juvenile snails. With regard to spawning, the ET50min and ET50max calculated at 14.9 °C and 38.1 °C.
The total development period from egg to adult snail capable of spawning was calculated at 42–65 days (at indoor temperatures varying from 15 °C to 25 °C), with an average of 50.0 ± 5.2 days, while the AT required for completing the development of one snail generation was 607.0–892.3 ℃∙d, with an average of 712.1 ± 64.9 °C∙d.
The survival and reproductive risk
Calculation of the annual number of snail generations allowed by the temperature records (155, 233, 236, 236, 228, 228, 226 and 197) from each decade’s index year (1955, 1965, 1975, 1985, 1995, 2005, 2015 and 2019), based on the AT values, gave us the permitted number of snail generations per index year investigated (Table 2). Importantly, however, too few development generations cannot meet the colonization needs of the snails, so the low figures in the first columns do not suffice for colonization, while the values given in the following columns do.
The northward displacement analysis
The dynamic GIS-based study of the potential distribution of B. globosus showed a gradual expansion from South to North during the period 1955–2019. The northern boundary of the potential geographical distribution of snails in 1955 concerned mainly in the coastal areas of Fujian, Guangxi, Guangdong, northern Hainan and Yunnan’s south-western border area gradually (Fig. 2). Importantly, the overall restriction, i.e., the one represented by 2019 of 31.0% (61/197) as it was governed by the egg incubation restriction, a situation mainly seen in Hainan Province and in the southern parts of the provinces of Yunnan, Sichuan, Guizhou, Guangdong, Fujian and Guangxi Autonomous Region (Fig. 6). With respect to the number of snail generations allowed, 59.4% of the MMSs (117/197) reached more than 5 generations and 5.6% of the MMSs (11/197) reached more than 8 generations, which was mainly distributed in the south-eastern area of Yunnan, the southern parts of Guangxi and Guangdong and all of Hainan Province, while only one (0.5% of the MMs or 1/197) reached 9 generations, which indeed was located in Hainan for the year 2019 (Table 2).

The northward displacement in southern China of the boundary defining areas that could theoretically support colonization of B. globosus. Map approval No. GS(2022)2432
The potential risk of colonization
The analysis of the annual generation based on the temperature data from 1955 to 2019 shows that the snail can complete the reproduction of 1–9 generations in more than 90% of the areas monitored by the MMSs of the 11 PLADs in southern China. The distribution area where the number of annual generations was more than 8 showed a gradual expansion trend from South to North, i.e., the number and proportion of MMSs had increased from 1 to 10 (Table 2). This study comprehensively analyzed the superposition of the survival map and the snail annual developmental GIS map for prediction from 1955 to 2019. The areas that could theoretically support annual regeneration of B. globosus in the mainland of China is showed a gradual expansion trend from South to North. In 1955, the potential snail distribution would have stretched from the coastal areas of Guangdong Province and Guangxi Autonomous Region to southern Yunnan Province via the borders to Vietnam, Laos and Myanmar. Since then, this line has gradually moved northward and the boundary. In 2019, the areas allowing ≥ 7 snail generations have reached north of the tropic of cancer connecting the southernmost coastal area of Fujian Province with the middle of Yunnan Province via Guangxi and Guangdong. The area meeting the development requirements of 5–9 generations of B. globosus is concentrated in parts of Yunnan, Guangdong, Guangxi, Fujian and covers the whole province of Hainan (Fig. 7).

Areas that could theoretically support annual regeneration of B. globosus in the mainland of China on 2019. Map approval No. GS(2022)2432
Discussion
This study depended on China's aid to Zanzibar schistosomiasis prevention and control project that made us investigate the temperature for the growth and development of B. globosus snails in the laboratory. We then went on to use the model developed combined with MMS temperature data to find out if these snails could also colonize southern China, as this would involve the potential of schistosomiasis haematobia take hold in China. In this research, we combined ArcGIS 10.7 software with the Geostatistical Analysis extension to use the EBK model to generate maps indicating snail survival as well as maps predicting the B. globosus colonization risk. As our findings indicate that this could indeed occur, the government will need to strengthen the supervision of accidental introduction of snails.
The relationship between development of organisms and the ambient temperature not only governs the speed of biological development but also sets the boundaries for the distribution of many organisms, particularly those that cannot regulate their body temperature. The AT estimates this time by estimating the amount of heat an organism must accumulate to reach full development. While it must always be within its restricting extreme thresholds, the organism in question obviously needs a longer time at lower temperatures than in a warmer climate. AT was originally used to measure crop maturation, but can also be used for predicting development of parasites, which led to seeing the environment as a set of contiguous niches with diverse ecology as first outlined by Pavlovskii [25] and later developed by Fick and Hijmans [26] into producing the Bioclim environmental records that indicate habitat suitability modelling under past, present or future conditions [27, 28].
Mapping and statistical analysis of climate data contribute to our awareness of how organisms grow and spread. Already in the eighteenth century, René Antoine de Réaumur introduced a unit of AT defined as the amount of heat used an organism needs to accumulate to reach full development [29]. Originally used for predicting the seasonal plant growth in agriculture, this growing-degree-day (GDD) unit has more recently been used for widely different measurements, including the development of parasites and also their vectors [30,31,32]. Here called the AT, this unit accurately expresses the quantitative relationship between snail growth and reproduction and environmental temperature. It is a relatively stable ecological index for the growth and development of snails [33]. The analysis of the annual generation shows that B. globosus can complete more than one generation of development and reproduction in 10 PLADs of southern China, which is a potential geographic distribution area. This study comprehensively analyzed the superposition of the survival risk GIS map and the snail annual developmental GIS map for the different index years, one for each decade.
The dynamic GIS-based study of the potential distribution of B. globosus showed a gradual expansion from South to North during the study period moving the boundary of the potential geographical distribution of snails from the southern borders and the coastal areas northward. As a reflection of the ongoing global warming of the latest 100 years, which has in the past 30 years pushed the average January temperature in China by 1 °C [32], this potential area includes now large parts of Yunnan, Guangdong, Guangxi as well as the whole province of Hainan and a small area in southern Fujian. Along with the prediction a 0.9 °C rise of the average January temperature in China between 2008 and 2030 followed by another one at 1.6 °C by 2050 [34, 35] and the fact that the minimum temperatures have generally risen, it is obvious that the new temperature levels have already created a more suitable condition for the survival, reproduction and development of B. globosus, we face multifaceted risk scenarios. The expansion of the distribution area of snails in general will affect existing geographical schistosomiasis distributions, potentially even lead to the introduction of schistosomiasis haematobium [36, 37].
Although the highest, lowest and average AT have been obtained in the laboratory previously [38,39,40], we felt that a more detailed investigation is required. We not only found the ET50max and ET50min for each developmental stage of B. globosus important but also the thresholds for spawning. The suitable growth temperature for B. globosus is 26–29 °C and if the environment temperature exceeds the maximum or minimum temperature for a period of time, it may not survive [41,42,43]. Knowledge of geographical temperature patterns is very important when attempting to delineate distribution areas and breeding places. Using the ET50 can reduce errors and improve the reliability of the study [44]. The results of this study show very clearly that B. globosus do not propagate by eggs deposition at temperatures below 15 °C or above 38 °C (Table 1), which is consistent with the report by Kalinda et al. [19]. The results of the study showed that the range between ET50min and ET50max for B. globosus in the laboratory varies between 7.0 °C and 40.5 °C. Thus, from the temperature point of view, the spawning and hatching of the snail egg turned out to be the most vulnerable stage and therefore represent the overall strongest restriction of long-term snail survival.
As B. globosus are aquatic snails, the water temperature has a direct impact and sustains the development and reproduction of snails better than air. Since the snails live in shallow waters such as lakes, streams and agricultural irrigation channels, the water and air temperatures are similar. At the same time, there are reports indicating that B. globosus is very sensitive to water temperature. For snails living in deep water bodies, when the water temperature in the deep area is lower than the air temperature, the snails will migrate to the surface of the water body or even climb out of the water surface to escape the low temperature. Therefore, this study was carried out in a constant temperature box in the laboratory. To simulate the impact of meteorological temperature on the snail, the temperature data of the model were collected from the national meteorological monitoring point [45,46,47].
Although there are not yet any signs of B. globosus colonization in China, our examination is warranted as Biomphalaria straminea, an intermediate snail host of S. mansoni in Brazil, was introduced to China in aquatic plants shipments and established habitats in Hong Kong many decades ago. It has not been possible to control that this invasive snail has spread widely in the Pearl River Delta and in Guangdong [48,49,50]. The risk of the introduction of B. globosus into the mainland of China has increased with implementation of “the Belt and Road Initiative” that has opened frequent trade exchange between China and African countries [51]. As the temperature is key for snail survival [52], we focused on this variable leaving other factors, such as light, pH, and oxygen content influence for future studies.
Conclusions
Our study obtained temperature-related parameters of B. globosus through laboratory experiments, and then combined with the meteorological data in the mainland of China to predict the risk of colonization in China via using related models. Our study finds that annual regeneration of B. globosus can be supported by the current climate conditions in the mainland of China, and a gradual expansion trend from south to north is shown in the study from 2015 to 2019. Thus, there is a potential risk of colonization of B. globosus in the mainland of China under climate change. This study provides a model for monitoring the invasion risk of snails in non-endemic countries. To avoid the incidence and prevalence of schistosomiasis haematobia in China, rigorous control of snail importation and patients with active urogenital schistosomiasis are warranted.
Availability of data and materials
All data supporting the findings of this study are included in the article and additional file.
Abbreviations
- ET50:
-
Median effective temperatures
- AT:
-
Accumulated temperature
- GIS:
-
Geographic information system
- ET50min :
-
Effective minimum temperature
- ET50max :
-
Corresponding maximum values
- IPCC:
-
Intergovernmental panel on climate change
- MMSs:
-
Meteorological monitoring stations
- GDD:
-
Growing-degree-day
References
Colley DG, Bustinduy AL, Secor WE, King CH. Human schistosomiasis. Lancet. 2014;368(9936):2253–64.
King CH, Galvani AP. Underestimation of the global burden of schistosomiasis. Lancet. 2018;391(10118):307–8.
Steinmann P, Keiser J, Bos R, Tanner M, Utzinger J. Schistosomiasis and water resources development: systematic review, meta-analysis, and estimates of people at risk. Lancet Infect Dis. 2006;6(7):411–25.
Savioli L, Fenwick A, Rollinson D, Albonico M, Ame SM. An achievable goal: control and elimination of schistosomiasis. Lancet. 2015;386(9995):739.
Abdel-Wahab MF, Yosery A, Narooz S, Esmat G, el Hak S, Nasif S, et al. Is Schistosoma mansoni replacing Schistosoma haematobium in the Fayoum? Am J Trop Med Hyg. 1993;49(6):697–700.
Antoni S, Ferlay J, Soerjomataram I, Znaor A, Jemal A, Bray F. Bladder cancer incidence and mortality: a global overview and recent trends. Eur Urol. 2017;71(1):96–108.
Khurana S, Dubey ML, Malla N. Association of parasitic infections and cancers. Indian J Med Microbiol. 2005;23(2):74–9.
Hua HY, Ren GH, Liang YS. Clinical manifestations and treatment of schistosomiasis hematobia. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2014;26(04):357–61 (in Chinese).
Jarne P, Finot L, Delay B, Thaler L. Self-fertilization versus cross-fertilization in the hermaphroditic freshwater snail Bulinus globosus. Evolution. 1991;45(5):1136–46.
Caldeira RL, Jannotti-Passos LK, Carvalho OS. Molecular epidemiology of Brazilian Biomphalaria: a review of the identification of species and the detection of infected snails. Acta Trop. 2009;111(1):1–6.
Pedersen UB, Stendel M, Midzi N, Mduluza T, Soko W, Stensgaard AS, et al. Modelling climate change impact on the spatial distribution of fresh water snails hosting trematodes in Zimbabwe. Parasit Vectors. 2014;7:536.
Wang YA, Yang K, Liang YS, Qu GL, Shi F, Xing YT, et al. Studies on colonization risk and potential geographical distribution of Biomphalaria glabrata as an intermediate host of Schistosoma mansoni in Mainland of China. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2018;30(03):11-6+21 (in Chinese).
Yang GJ, Bergquist R. Potential impact of climate change on schistosomiasis: a global assessment attempt. Trop Med Infect Dis. 2018;3(4):117.
Zhang LJ, Xu ZM, Dang H, Li YL, Lv S, Xu J, et al. Bulletin of national schistosomiasis epidemic situation in 2019. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2020;32(06):7–14 (in Chinese).
Gribov A, Krivoruchko K. Empirical Bayesian kriging implementation and usage. Sci Total Environ. 2020;722: 137290.
Kutuzov S, Lavrentiev I, Smirnov A, Nosenko G, Petrakov D. Volume changes of Elbrus Glaciers from 1997 to 2017. Front Earth Sci. 2019;7:153.
Nocco MA, Smail RA, Kucharik CJ. Observation of irrigation-induced climate change in the Midwest United States. Global Change Biol. 2019;25(6809):3472–84.
Nogueira ADA, Bragagnolo C, Dasilva MB, Carvalho LS, Benedetti AR, Pinto-Da-Rocha R. Spatial variation in phylogenetic diversity of communities of Atlantic Forest harvestmen (Opiliones, Arachnida). Insect Conserv Diver. 2019;12(5):414–26.
Kalinda C, Chimbari MJ, Mukaratirwa S. Effect of temperature on the Bulinus globosus—Schistosoma haematobium system. Infect Dis Poverty. 2017;6(1):57.
Christie JD, Edward J, Goolaman K, James BO, Simon J, Dugat PS, et al. Interactions between St. Lucian Biomphalaria glabrata and Helisoma duryi, a possible competitor snail, in a semi-natural habitat. Acta Trop. 1981;38(4):395–417.
Kalinda C, Chimbari MJ, Grant WE, Wang HH, Odhiambo JN, Mukaratirwa S. Simulation of population dynamics of Bulinus globosus: Effects of environmental temperature on production of Schistosoma haematobium cercariae. PLoS Negl Trop Dis. 2018;12(8): e0006651.
Liu ZP, He XF, Zhang SH. Application of multi-measure weighted kriging method in high slope deformation stability analysis. J Hydraul Eng. 2009;40(006):709–15.
Chen K, Yao WG, Zhang Z, Xiao YY, Yan J, Xu Y, et al. Study on the colonization risk analysis and regionalization of wheat dwarf bunt in China. Acta Phytopathol Sin. 2002;32(4):312–8 (in Chinese).
Qin YJ. Global population structure, establishment risk and potential geographical distribution of Bactrocera dorsalis. China Agriculture University. 2017 (in Chinese).
Pavlovskii EN. The Russian academy of sciences and faunal investigations in Russia. Izv Akad Nauk SSSR Biol. 1945;2:129–46.
Fick SE, Hijmans RJ. WorldClim 2: new 1-km spatial resolution climate surfaces for global land areas. Int J Climatol. 2017;37(12):4302–15.
Malone JB, Bergquist R, Martins M, Luvall JC. Use of geospatial surveillance and response systems for vector-borne diseases in the elimination phase. Trop Med Infect Dis. 2019;4(1):15.
Malone JB, Nieto P, Tadesse A. Biology-based mapping of vector-borne parasites by geographic information systems and remote sensing. Parassitologia. 2006;48(1–2):77–9.
Bonhomme R. Bases and limits to using ‘degree.day’ units. Eur J Agron. 2000;13(1):1–10.
Bernal J, González D. Experimental assessment of a degree-day model for predicting the development of parasites in the field. J Appl Entomol. 1993;116(1–5):459–66.
Valencia-López N, Malone JB, Carmona CG, Velásquez LE. Climate-based risk models for Fasciola hepatica in Colombia. Geospat Health. 2012;6(3):S67-85.
Zhou XN, Yang GJ, Yang K, Wang XH, Hong QB, Sun LP, et al. Potential impact of climate change on schistosomiasis transmission in China. Am J Trop Med Hyg. 2008;78(2):188–94.
Boitani L, Fuller T. Research techniques in animal ecology: controversies and consequences. Columbia University Press; 2000.
Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, et al. The physical science basis: contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. IPCC Fourth Assessment Report (AR4). 2007;18(2):95–123.
Yang GJ, Vounatsou P, Zhou XN, Tanner M, Utzinger J. A potential impact of climate change and water resource development on the transmission of Schistosoma japonicum in China. Parassitologia. 2005;47(1):127–34.
Aguirre AA, Tabor GM. Global factors driving emerging infectious diseases. Ann N Y Acad Sci. 2008;1149:1–3.
Vonesch N, D’Ovidio MC, Melis P, Remoli ME, Ciufolini MG, Tomao P. Climate change, vector-borne diseases and working population. Ann Ist Super Sanita. 2016;52(3):397–405.
Hong QB, Zhou XN, Sun LP, Yang GJ, Yang K, Huang YX. Research on the impact of global warming on the spread of schistosomiasis in China IV. A study on the accumulated temperature of Oncomelania snail generations development in natural environment. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2003;15(4):269–71 (in Chinese).
Li T, Mo K, Wang J, Chen Q, Zhang J, Zeng C, et al. Mismatch between critical and accumulated temperature following river damming impacts fish spawning. Sci Total Environ. 2021;756: 144052.
University ECN. Animal Ecology. Beijing: Higher Education Press; 1984 (in Chinese).
Joubert PH, Pretorius SJ, Kock KND, Eeden JAV. Survival of Bulinus africanus (Krauss), Bulinus globosus (Morelet) and Biomphalaria pfeifferi (Krauss) at constant high temperatures. Afr Zool. 1986;21(1):85–8.
Marti HP, Tanner M, Degrémont AA, Freyvogel TA. Studies on the ecology of Bulinus globosus, the intermediate host of Schistosoma haematobium in the Ifakara area. Tanzania Acta Trop. 1985;42(2):171–87.
McCreesh N, Booth M. The effect of simulating different intermediate host snail species on the link between water temperature and schistosomiasis risk. PLoS One. 2014;9(7): e87892.
Hong QB, Zhou XN, Sun LP, Yang GJ, Yang K, Huang YX. Impact of global warming on the transmission of schistosomiasis in China II. The aestivation and lethal hyperthermy temperature of Oncomelania hupensis in laboratory. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi. 2003;15(1):24–6 (in Chinese).
Li KF, Hao HS, Zhuang CY, Pu L. A new method for predicting water temperature of river by using meteorological factors. J Sichuan Univ (Eng Sci Ed). 2006;38(1):4–7 (in Chinese).
McCreesh N, Arinaitwe M, Arineitwe W, Tukahebwa EM, Booth M. Effect of water temperature and population density on the population dynamics of Schistosoma mansoni intermediate host snails. Parasit Vectors. 2014;7:503.
Wang HD, Shi FX. A preliminary study on the water temperature of lakes in China. Trans Oceanol Limnol. 1980;3:23–33 (in Chinese).
Habib MR, Lv S, Guo YH, Gu WB, Standley CJ, Caldeira RL, et al. Morphological and molecular characterization of invasive Biomphalaria straminea in southern China. Infect Dis Poverty. 2018;7(1):120.
Wang W, Liang YS, Hong QB, Dai JR. African schistosomiasis in mainland China: risk of transmission and countermeasures to tackle the risk. Parasit Vectors. 2013;6(1):249.
Zeng X, Yiu WC, Cheung KH, Yip HY, Nong W, He P, et al. Distribution and current infection status of Biomphalaria straminea in Hong Kong. Parasit Vectors. 2017;10(1):351.
Hua HY, Wang W, Cao GQ, Tang F, Liang YS. Improving the management of imported schistosomiasis haematobia in China: lessons from a case with multiple misdiagnoses. Parasit Vectors. 2013;6(1):260.
Iijima T, Sugiura S. Studies on the temperature as a limiting factor for the survival of Oncomelania nosophora, the vector snail of Schistosoma japonicum in Japan. Jpn J Med Sci Biol. 1962;15(4):221–5.
Acknowledgements
This study was funded by the grants from National Natural Science Foundation of China, Grant number 82173586; Jiangsu Provincial Department of Science and Technology, grant number BZ2020003. We thank the schistosomiasis field study team, the laboratory staff of Pemba NTD and Ministry of Public Health for their great support in facilitating this work.
Funding
This research was funded by the Grants from National Natural Science Foundation of China, Grant number 82173586; Jiangsu Provincial Department of Science and Technology, Grant number BZ2020003.
Author information
Authors and Affiliations
Contributions
SJ, XW, WL, JZ and KY developed the study protocol and data analysis. XW, MS, MA, JH, MH and DX did the experiments. XW drafted the report. RB and KY revised the report. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was approved by the Ethics Committee in the Jiangsu Institute of Parasitic Diseases.
Consent for publication
Not applicable.
Competing interests
We declare that we have no conflicts of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, X., Juma, S., Li, W. et al. Potential risk of colonization of Bulinus globosus in the mainland of China under climate change. Infect Dis Poverty 11, 52 (2022). https://doi.org/10.1186/s40249-022-00980-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40249-022-00980-2