
Abstract
The holobiont concept was first developed for coral ecosystems but has been extended to multiple organisms, including plants and other animals. Studies on insect-associated microbial communities have produced strong evidence that symbiotic bacteria play a major role in host biology. However, the understanding of these symbiotic relationships has mainly been limited to phytophagous insects, while the role of host-associated microbiota in haematophagous insect vectors remains largely unexplored. Mosquitoes are a major global public health concern, with a concomitant increase in people at risk of infection. The global emergence and re-emergence of mosquito-borne diseases has led many researchers to study both the mosquito host and its associated microbiota. Although most of these studies have been descriptive, they have led to a broad description of the bacterial communities hosted by mosquito populations. This review describes key advances and progress in the field of the mosquito microbiota research while also encompassing other microbes and the environmental factors driving their composition and diversity. The discussion includes recent findings on the microbiota functional roles and underlines their interactions with the host biology and pathogen transmission. Insight into the ecology of multipartite interactions, we consider that conferring the term holobiont to the mosquito and its microbiota is useful to get a comprehensive understanding of the vector pathosystem functioning so as to be able to develop innovative and efficient novel vector control strategies.
Similar content being viewed by others

Background
A holistic view of organisms is necessary to understand the biology of metazoa. The host can no longer be considered as an isolated entity and instead should be considered as a chimera with close interactions with microbial communities. The extent interplay between both partners and its consequences on their evolutionary trajectory has given rise to the holobiont concept (i.e. the host and its community of associated microorganisms as well as their interactions) [1]. Under changing environmental conditions, associated microbiota, i.e. all microorganisms (bacteria, fungi, protists, viruses, etc.), can evolve rapidly and influence the acclimation, adaptation and evolution of host organisms. Changes in microorganism diversity and abundance may have a direct impact on the holobiont’s evolution by modifying the multipartite interaction dynamics.
The holobiont concept—originally developed for coral ecosystems—can also be transposed to other organisms. Most recent advances have shown that human gut microbiota plays a key role in regulating the host’s metabolic functions, immunity, nutrition, physiology and even behaviour [2]. In arthropods, first studies focusing on the molecular mechanisms underlying the role of microbiota such as the host’s immune function, nutrition, physiology and even behaviour concerned phytophagous insects [3]. However, there is still little evidence on the role of microbiota in haematophagous insects of medical importance (pathogen transmission to humans and/or animals) as well as the molecular mechanisms underlying their interactions with the host. The best-known examples concern the mutualistic symbiosis between tsetse flies (Diptera: Glossinidae) and its obligate bacterial symbionts Wigglesworthia spp. [4], even though available data on specific Wolbachia-mosquitoes associations have considerably increased in the past decades [5]. These bacteria participate in nutrient provisioning, insect fitness, host immunity maturation or pathogen transmission [6]. According to the holobiont concept, an arthropod vector should no longer be considered as an isolated organism but rather as a complex system in which the different partners (host and microbiota) interact.
According to the World Health Organization (WHO), mosquito (Diptera: Culicidae) vectors of arthropod-borne pathogens such as Anopheles sp., Aedes sp. and Culex sp. mosquitoes are the greatest threat to public health [7]. Anopheles mosquitoes are able to transmit to humans the causal agent of malaria, which is the deadliest vector-borne disease, with about 212 million cases and an estimated 429,000 deaths reported in 2015 [8]. Culex sp. mosquitoes are able to transmit both arboviruses and parasites [9] and Aedes sp. (mainly Aedes aegypti and Aedes albopictus) can transmit arboviruses of medical importance to animals and humans, including West Nile (WNV) (Flaviviridae, Flavivirus), dengue (DENV) (Flaviviridae, Flavivirus), Zika (ZIKV) (Flaviviridae, Flavivirus) and chikungunya (CHIKV) (Togaviridae, Togavirus) viruses [10, 11]. Dengue is the most serious mosquito-borne viral disease and major chikungunya outbreaks have occurred worldwide over the last decade. Zika virus infection has also recently become a major public health concern, with the global spread of the disease and foetal microcephaly cases arising in women during pregnancy [12]. In addition, some of these mosquito species are invasive and their geographical distribution has been expanding as they take advantage of human activities such as international trade, emphasized by global warming [13, 14]. The lack of effective and preventive treatments against most mosquito-borne diseases restricts control strategies to individual protection and mosquito population control using insecticides. However, intensive and repeated of biocides use leads to the development of mosquito resistance and unwanted effects on non-target species.
There is recent increased interest in studies on mosquito-associated microbiota, which encompass bacteria, fungi, protists and viruses, i.e. both mosquito-specific viruses (MSVs) and the transmitted pathogens. Research on the role of microbial communities in the host biology and pathogen interference has led to the development of new vector control approaches based on the use of “symbiotically” modified mosquitoes [15]. Knowledge on the roles of microbes in the development, physiology or immunity of their hosts, as well as interference with transmitted pathogens, is henceforth essential to be able to develop these alternative strategies. This review summarizes current knowledge on mosquito-associated microbiota. We discuss recent advances and current prospects on the ecology and functions of the mosquito microbiome as well as interactions with the host and transmitted pathogens.
Influence of the breeding site ecology on mosquito bacterial microbiota
The origin of microbes colonizing mosquitoes and the role of the environment in microbial acquisition are issues that have long been debated [16]. This aspect is essential to define the dynamics of microbial communities in the mosquito holobiont. Most studies carried out to date on microbial acquisition in mosquitoes have been focused mainly on the bacterial component.
Recent studies confirmed that a substantial fraction of bacteria colonizing mosquitoes is acquired during the aquatic life stage, through the aquatic larval habitats. The microbial composition and environmental characteristics of breeding sites could partially explain the different colonization patterns of bacteria in immature and adult mosquito stages (Fig. 1). These effects likely accentuate interspecific variations in the microbiota due to the habitat tropism of the different mosquito species. Duguma et al. [17] showed some association patterns between nutrient contents and microbial composition in larval habitats and bacterial communities associated with Culex nigripalpus adults. While mosquitoes originating from high-nutrient habitats were associated with members of the Clostridiales order, those from low-nutrient habitats were instead associated with Burkholderiales order members. A specific community profile depending on environmental factors has also been associated with the bacterial composition in Anopheles gambiae [18]. Interestingly, the difference in bacterial diversity of larvae from different mosquito species sharing the same breeding site was shown to be lower than that of larvae of similar species living in different collection sites [19]. In Anopheles coluzzii and An. gambiae, some bacterial communities are shared among fourth instar larvae, water of the larval habitat and adults [20]. Another interspecific comparison indicated that the bacterial communities present in aquatic larval habitats and in larva guts were similar to each other and differed from the bacterial communities of the adult guts [21]. Similarly, only Firmicutes and Actinobacteria phyla were commonly found in both Ae. aegypti larvae and aquatic ecosystems, with higher bacterial diversity found in water than in larvae [22]. This suggests that although the bacterial community is acquired from water, the insect’s gut is a more selective habitat for bacteria. This selectivity could be explained by the physicochemical conditions present in the gut (e.g. alkaline pH, redox potential, oxygen level below 5%, etc.) as well as other factors such as the immune response, peristalsis or presence of lytic enzymes or microbial interactions. Moreover, the findings of other studies indicate that some larvae-borne bacteria persist in adults (Fig. 1). Interestingly, the Thorsellia genus was detected in both immature (early and late larval instars and pupae) and adult stages in Culex tarsalis [23]. In Ae. albopictus, some bacteria belonging to Micrococcaceae, Pseudomonadaceae and Staphylococcaceae families are common to larvae, adult males, as well as sugar-fed and blood-fed females [24].

Venn diagrams illustrating overlapping of bacterial composition between mosquito species, development stages and habitats. a Number of bacterial taxa specific and common between mosquito larvae, habitats and adults of Aedes japonicus, Aedes triseriatus and Anopheles gambiae [20, 21]. b Number of bacterial taxa specific and common to larvae of Anopheles gambiae, Culex pipiens, Culex nigripalpus, Aedes aegypti and Aedes japonicus [17, 18, 20, 22, 23, 27]. c Number of bacterial taxa specific and common to adults of Anopheles gambiae, Anopheles stephensi, Culex nigripalpus, Aedes albopictus and Aedes aegypti [17, 18, 20, 21, 23, 24, 27, 29,30,31,32,33, 35,36,37, 39]. An additional table shows in more detail the identification of bacterial species/genera in mosquito species [see Additional file 1]
Overall, these results imply that there is a continuum of bacteria from the aquatic environment to immature stages and adult mosquitoes, as shown by the overlap in bacterial composition between water, larvae and adults [20, 25] (Fig. 1). Contrary to previous assumptions [26], bacterial clearance during mosquito metamorphosis from pupae to adults would not be complete, clearly suggesting that a subset of such environmental-acquired bacterial microbiota will intrinsically be part of the holobiont cell components.
Mosquito core- and pan-microbiota
From mosquito core-microbiota…
The concept of a core microbiota in mosquitoes (i.e. shared by mosquito populations belonging to the same species) has been addressed in recent years [20]. Generally defined as being a microbial community associated with hosts from a given group (e.g. individual, population, genus, species, etc.), a more flexible definition considering the microbial species most prevalent in the hosts (≥ 90%) has been proposed [27, 28]. Many studies have described core microbiota shared by different adult mosquito populations and/or species collected in distinct geographic areas [see Additional file 1]. For instance, Pseudomonas, Acinetobacter and Aeromonas spp. bacteria were detected in different Ae. aegypti populations from Brazil [29]. Likewise, An. gambiae populations collected at different sites in Burkina Faso shared Thorsellia, Wolbachia, Massilia and Acinetobacter spp. bacteria, which correspond to the most abundant taxa associated with those populations [18]. In Vietnam, Acinetobacter was found to be the sole core microbiota constituent of 11 Anopheles species [30, 31]. Indigenous and invasive populations of Ae. albopictus from Vietnam and France, respectively, also shared core bacterial microbiota, with Dysgonomonas being the most prevalent and abundant genus [32]. The tissue tropism of core microbiota was also examined. Staphylococcus, Corynebacterium, Geobacillus, Micrococcus, Acinetobacter and Pseudomonas spp. bacteria are present in male and female An. gambiae and An. coluzzii reproductive tissues [27]. Tchioffo et al. [33] detected core microbiota composed of Pseudomonas, Comamonas, Acinetobacter, Rhizobium, Burkholderia and members of the Enterobacteriaceae family in different Anopheles mosquito tissues, i.e. ovaries, salivary glands and midgut. Further studies are needed to investigate whether this tissue tropism is correlated with potential core microbiota functions.
…to mosquito pan-microbiota
As suggested for other organisms, the ecology modulates host-associated microbiota, thus prompting us to apply the pan-microbiota concept to mosquito, e.g. microbiota shared by different mosquito species regardless of their geographic origin [34]. Indeed, environmental factors influence the microbial composition of breeding sites and food resources (plants, sugar, blood). However, as mentioned above, the fact that a common bacterial fraction is shared by different mosquito species at various developmental stages is in favour of the environmental acquisition hypothesis [see Additional file 1]. This repeated-bacterial colonization leads to a stable association between mosquitoes and their environmentally acquired microbiota. For instance, cultivable core microbiota of Ae. aegypti and Ae. albopictus females in north-eastern India was found to be composed of the same bacterial species, i.e. Enterobacter cloacae, Klebsiella michiganensis, Pseudomonas monteilii, Bacillus aryabhattai, Lysinibacillus fusiformis and Staphylococcus hominis [35]. In Culex pipiens and Culex restuans, 44% of operational taxonomic units (OTUs) were common in both species [36]. However, bacterial diversity was reportedly lower in distant mosquito species. Only Bacillus and Escherichia/Shigella spp. were common in Cx. pipiens, Culiseta incidens and Ochlerotatus sierrensis [37]. Similarly, Pseudomonas and Wolbachia were the two genera shared by Culiseta melanura and Coquillettidia perturbans [38]. A broader study targeting 12 mosquito species in the USA, including Aedes, Anopheles and Culex, revealed similarities among their bacterial communities [see Additional file 1]. These communities were dominated by Gluconobacter, Propionibacterium and Staphylococcus bacterial taxa [39]. Part of the microbiota in adult mosquitoes would thus be acquired from the aquatic larval habitat, while the other fraction would be dependent on food resources, through natural plant-based sugar sources or through blood meals for females. Bacterial composition and diversity are modified subsequently to a sugar- and/or blood-meal which increase inter-individual differences [20]. Notably, blood meal leads to a progressive shift in oxidative conditions in the gut through the modification of microbial communities’ composition and structure [40, 41]. Diversity and composition of the bacterial populations are influenced by both infection status and time after the blood meal in Ae. albopictus [42]. The bacterial composition but not its structure is influenced by the blood meal whereas only few taxa varied significantly due to chikungunya virus infection [42]. The bacterial abundance of La Crosse virus (LACV)-infected Aedes japonicus and Aedes triseriatus increased while richness and evenness of resident fungi decreased [43]. Otherwise, the bacterial abundance is reduced in Ae. aegypti infected by DENV [44]. ZIKV infection also modulates the dynamics of the bacterial families Rhodobacteraceae and Desulfuromonadaceae in Ae. aegypti, suggested as potential markers for ZIKV [45]. The associated effects of blood meal and infection tend to accentuate microbiota–mosquito immune interactions, such as immune response as well as redox and detoxifying enzyme metabolisms mentioned below [41]. Similarly, a study conducted by Short et al. [46] showed that part of the host amino acid metabolic pathway, which involves branched chain amino acid degradation, did affect midgut microbial communities in Ae. aegypti. This could partly explain variations in the midgut microbiota of mosquitoes in the field.
To summarize, recent reports have confirmed that the ecology of breeding sites drives environmental bacterial acquisition in mosquitoes. Above all, core and pan-microbiota might represent an assemblage fraction of the mosquito that belongs to the extended genome of the mosquito hologenome and contributes to key features of the holobiont. Further studies are necessary to effectively link these core microbes and their genomes with the functions displayed and to determine which host genetic factors govern host-microbe interactions.
“Neglected” microbes in the mosquito holobiont
Mosquito virobiota
A novel group of insect-specific viruses has recently been described [47]. In contrast to arthropod-borne viruses which have a dual host tropism (i.e. can replicate in mosquitoes and vertebrates), these insect-specific viruses (ISVs) are host restricted and do not replicate in vertebrate cells. Although most have been discovered in mosquitoes, ISV sequences have also been detected in other haematophagous insects such as sandflies [47]. The cell fusing agent virus (CFAV), Kamiti River virus (KRV) and Culex flavivirus (CxFV) were the first endogenous mosquito-specific viruses (MSVs) identified in Aedes and Culex sp. mosquitoes [see Additional file 1] [48,49,50].
In the past decade, next-generation sequencing analysis and increasing interest in both microbiome and arbovirus transmission have led to the description of many MSVs [see Additional file 1]. Despite the host restriction, these RNA viruses are related to mosquito-borne viruses (MBVs) and they essentially belong to the Bunyaviridae, Birnaviridae, Flaviviridae, Mesonoviridae, Negoviridae, Reoviridae, Rhabdoviridae and Togaviridae families [51,52,53]. Most ISVs described to date belong to the Flaviviridae family [54]. Mosquito-specific flaviviruses are divided into two distinct phylogenetic and serologic clades. The first group is closely related to mosquito-borne flaviviruses, including dengue (DENV), yellow fever (YFV) and West Nile (WNV) viruses, while also comprising a dozen virus species [51, 55]. The second one forms a clade distinct from mosquito-borne flaviviruses and includes the previously mentioned viruses CFAV, KRV, CxFV and Aedes flavivirus (AeFV), and others [55]. These mosquito-specific flaviviruses are distributed worldwide and have been isolated or detected in diverse mosquito species [see Additional file 1]. For instance, CFAV, KRV, CxFV or AeFV have been isolated or detected in laboratory reared and field-caught Aedes, Anopheles or Culex populations from Africa, Australia, South America or Japan [55]. Mosquito-specific alphaviruses are less diversified as only two viral species have been described [56, 57]. The first one, i.e. the Eilat virus (EILV), was isolated from collected Anopheles coustani in Israel [56] and the second one, i.e. the Taï Forest virus, was recently detected in Culex decens from Côte d’Ivoire [57].
Despite the high prevalence of MSVs in nature, their acquisition, transmission cycle and maintenance in mosquitoes are poorly known. Like some bacteria, vertical transmission from mother to progeny has been reported. Culex and Aedes flaviviruses are vertically transmitted in Cx. pipiens and Ae. albopictus, respectively [58, 59]. However, horizontal transmission could occur as the same MSVs can be detected in different mosquito species from the same geographical area. Both venereal and ectoparasite transmission have been reported. As for phytophagous insects, MSVs could be acquired from the environment and transmitted through infected plants during nectar feeding [51, 53, 60]. MSVs thus have to infect mosquito salivary glands and/or saliva “injected” during feeding. Mosquito flaviviruses CFAV are absent from saliva and salivary glands of Ae. aegypti and Culex annulirostris, while AeFV was detected in Ae. albopictus saliva [61]. Finally, CxFV was only found in Culex quinquefasciatus saliva bi-infected with WNV [61]. The Eilat alphavirus was detected in salivary glands of Ae. aegypti, Ae. albopictus and Cx. quinquefasciatus [61]. As no horizontal transmission was detected between Cx. pipiens mosquitoes infected or not by CxFv and reared in the same cage with common food resources [62], further investigations are required to corroborate these horizontal modes of transmission. Temperature could modulate their prevalence and/or maintenance in field populations since mosquitoes are ectotherms and some MSVs are temperature sensitive [63].
Reverse genetic tools have revealed that restrictions occurred during cell entry and replication steps and during viral assembly for Eilat and Niénokoué viruses, respectively [56, 64, 65]. Mammal innate immunity could also explain host restriction, as demonstrated for KRV, which can complete its viral replication cycle in cells deficient in some interferon regulatory factors [66].
Mosquito-associated eukaryotes
In addition to viruses and bacteria, the mosquito microbiota is also composed of fungal (mycobiota) and protist communities [see Additional file 1]. However, few studies have focused on these communities in mosquitoes. Recently, Belda et al. [67] developed an efficient method that enabled the identification of eukaryotic microbiota associated with Anopheles mosquito larvae. Ichthyosporeans of the Pseudoperkinsus group were the most abundant protist members. Steyn et al. [68] also provided the first description of yeast microbiota of Cx. pipiens and Culex theileri larvae. The authors identified Candida, Cryptococcus, Galactomyces, Hannaella, Meyerozyma, Pichia, Rhodosporidium, Rhodotorula, Trichosporon and Wickerhamomyces genera. Apart from non-pathogenic fungi, mosquitoes also harbor yeasts of clinical importance. A recent study reported the isolation of the opportunistic pathogen Candida parapsilosis from different developmental stages and organs of several laboratory-reared mosquito species, including An. gambiae, An. stephensi, Cx. quinquefasciatus, Ae. albopictus and Ae. aegypti [69]. Moreover, culturable yeast microbiota analysis from wild Cx. pipiens and Cx. theileri mosquito larvae revealed the presence of clinically relevant species, including the well-known opportunistic human pathogen Candida albicans.
Overall, there has been much consideration of bacterial microbiota in the holobiont concept as it represents the abundant fraction of the host microbiota and consequently has been shown to impact the extended phenotype. The advent of high-throughput sequencing methods has made it possible to describe other communities of microbes including viruses and protists. Some of them, even at lower abundances, established notable interactions with their host. In mammals, retroviral genes are thought to be involved in the placenta formation [70]. We provided here some published data on virobiota and eukaryotic microbiota associated with mosquitoes. We encourage experimental evolutionary and functional researches to be performed on this neglected microbiota and we anticipate, in light of what was demonstrated with bacterial microbiota, that it will build a foundation to extend the list of microbial taxa involved in the mosquito holobiont.
Microbiome-mosquito interactions and mosquito holobiont success
As previously mentioned, most studies on mosquito microbiota have been descriptive and focused on bacteria with the aim of drawing up an inventory of the microbial communities [see Additional file 1] and their variation factors. While recent studies have demonstrated interactions between bacteria and mosquitoes as well as between fungi and mosquitoes, the role of MSVs in the host biology remains to be determined. Cytophatic effects have been observed in cell cultures, but it is still unknown whether and how MSVs influence host life history traits (lifespan, fecundity, oviposition).
Growth and larval development
In recent years, major studies have focused on the role of microbes in mosquito development (Fig. 2). One of the most relevant examples is about the Asaia genus, which is closely associated with An. stephensi and is one of the principal members of its microbiota. Up- and downregulation of genes involved in cuticle synthesis in Asaia-infected larvae promotes the insect’s growth (size and development stages) [71]. Similarly, both bacteria (Klebsiella and Aeromonas) and yeasts (Saccharomyces cerevisiae) promote Cx. pipiens development [72]. Another study showed that the survival and pupation of Cx. pipiens larvae were negatively impacted when they were fed with yeast isolates compared to fish food [68].

Putative functions of mosquito-associated microbiota (bacteria and fungi). Functions related to metabolism are indicated in orange and those related to life history traits are indicated in yellow: (1) Blood digestion (Acinetobacter, Pantoea, Enterobacter, Dysgonomonas), (2) Sugar digestion (Acinetobacter, Elizabethkingia, Thorsellia, Sphingomonadaceae family, Meyerozyma), (3) Supply of vitamins and amino acids (Dysgonomonas, Klebsiella, Aeromonas, Saccharomyces cerevisiae), (4) Survival (Escherichia coli, Beauveria bassiana), (5) Mediating oviposition site choice (Klebsiella, Aeromonas), (6) Egg production (Comamonas), (7) Larval development (Acinetobacter, Asaia, Aeromonas, Chryseobacterium, Paenibacillus, Aquitalea, Escherichia coli) [19, 25, 68, 71,72,73,74,75, 77, 80]. Pictures from JM Hosatte, with permission
Besides experimental infections, antibiotic treatments are the main approaches used to manipulate mosquito bacterial communities for the purpose of studying the functional roles of insect microbiota. However, these methods fail to eliminate all the bacteria. An alternative approach was therefore developed to generate axenic (i.e. without microorganisms) mosquitoes using a combination of ethanol and bleach to sterilize the egg surface. Gnotobiotic (i.e. with known microbiota) individuals can thus be obtained via inoculation of a given bacterium [25] or other microbes. Recent studies using this experimental approach, together with functional assays, clearly demonstrated that larvae rely on bacteria for their development. All axenic Ae. aegypti, An. gambiae and Aedes atropalpus larvae fed with a standard sterile diet died during the first larval instar [25]. However, mono-associated inoculation of Acinetobacter, Aeromonas, Aquitalea, Chryseobacterium or Paenibacillus restored Ae. aegypti larval development. Likewise, Ae. albopictus and Cx. quinquefasciatus require gut colonization by living bacteria for their development [19]. Overall, these results suggest that mosquitoes do not rely on specific bacterial taxa for their development but rather on exchangeable symbiont combinations that could be acquired through larval aquatic habitats.
Some mechanisms involved in larval development have been described. Genes involved in nutrient acquisition, metabolism and stress responses are differentially expressed in the first axenic Ae. aegypti larval stage compared to conventional and gnotobiotic ones, suggesting a putative role of gut bacteria in nutrient acquisition and/or assimilation after hatching [73]. Mechanisms were detected with Ae. aegypti gnotobiotic larvae colonized by different E. coli mutants [74]. The cytochrome bd oxidase gene appeared to be a key component in this interaction by reducing the gut oxygen level. Consequently, hypoxia induces the stabilization of hypoxia-induced transcription factors (HIFs) that enables larval growth and ecdysone-induced molting [74, 75]. To summarize, larvae acquire some of their bacterial microbiota from aquatic habitats and these bacteria contribute to the host development, survival and pupation. Any disturbance of the microbial community in the larval aquatic habitat could therefore impact the mosquito biology and ecology [76].
Egg production and oviposition
Coon et al. [77] demonstrated the contribution of gut bacterial microbiota in egg production by Ae. aegypti and Ae. atropalpus while comparing gnotobiotic larvae to their axenic relatives (Fig. 2). Ae. atropalpus can produce their first clutch of eggs without blood-feeding and depends on specific members of their gut microbiota to produce eggs whereas a blood meal is mandatory for Ae. aegypti. Ae. atropalpus microbiota probably provides nutrient reserves during larval development which are necessary for the first clutch in the absence of a blood meal. Interestingly, Cx. pipiens females were shown to select media containing Klebsiella and Aeromonas bacteria for oviposition [72].
Mosquito pathogens
Nowadays, Bacillus thuringiensis subsp. israelensis (Bti) producing Cry, Cyt, Vip and Sip insecticidal proteins is a safer alternative to chemical insecticides. However, its repeated and intensive use was found to generate strong selection pressure that could promote Bti resistance. A reduction in bacterial microbiota diversity in An. stephensi larvae has been shown to increase their susceptibility to Bti [78]. This finding differs from what was previously known in other insect models where microbiota was a key component in Bti efficiency [79]. In addition, the microaerophilic conditions of the larval gut would also favour the use of insecticidal proteins as a source of nitrogen by bacteria [78]. Intestinal bacteria would increase larval resistance through Bti toxin degradation. There has been recent increased interest in the identification of mosquito-killing fungi to tackle insecticide resistance. It was recently shown that the pathogenic fungus Beauveria bassiana could interact with gut bacterial microbiota and accelerate Anopheles mosquito death [80]. The fungus induced an increase of the opportunistic pathogenic bacterium Serratia marcescens density that lead to its dissemination in the haemocoel and promote death of their host [80].
Published data reported here illustrate that hypothesis- and experimentally- driven researches are key elements to demonstrate the contribution of bacterial microbiota in extended phenotypes of the mosquito holobiont. Notably, the manipulation of microbial assemblages allows to deduce their important impact on the mosquito life history traits and somehow which host-genes were modulated.
Mosquito-microbiota co-evolution
Recent studies have highlighted host-microbiota phylosymbiosis, i.e. a congruency between the host phylogeny and the divergence in its associated microbial community composition [81, 82]. A field study conducted by Novakova et al. [82] in 11 mosquito species from Canada revealed congruency between the phylogeny of hosts and differences in their associated bacterial communities. However, this co-evolutionary pattern was not observed for all related host species. Indeed, in standard laboratory conditions, Ae. aegypti showed more divergent microbiota from the closely related species Ae. atropalpus than from the distantly related species An. gambiae [25]. Conversely, a congruence pattern in microbial communities was also observed in species belonging to the same subgroup. Analysis of gut bacterial microbiota revealed a strong similarity between Ae. albopictus and a cryptic species living in sympatry in Vietnam [81]. Those findings could possibly be explained by recent host-microbe co-adaptation. Additional studies are necessary to clarify the influence of local environmental parameters and host genotypes on the mosquito microbiota composition. Evolutionary prospects of Wolbachia-mosquito associations have also recently been highlighted. Besides, horizontal gene transfers (HGTs) were described between Aedes mosquitoes and Wolbachia [83, 84]. The evolution of Wolbachia-mosquito associations raises the question of the outcome of bacterial-to-mosquito HGTs on adaptive capacities of the holobiont or/and mosquito vector competence, as Wolbachia can modulate the transmission of some pathogens depending on the mosquito species [5]. The adaptive capacities of the holobiont could also be impacted by functional bacterial HGTs. This event was demonstrated for the coffee-crop insect that acquired a gene encoding a polysaccharide enzyme from bacteria belonging to Bacilli class through HGT, that could extend its host-plant range and so promote its adaptation to new ecological niches [85].
Likewise, phylogenetic studies have highlighted that some MSVs belonging to Bunyaviridae and Flaviviridae families have co-evolved and diversified with their mosquito host [63, 86]. Transovarial transmission as well as the detection of integrated viral sequences in mosquito genomes (i.e. endogenous viral elements [EVEs]) [87] supports the hypothesis of an “ancient” association and possible co-evolution. A description of new lineages of mosquito-specific bunyaviruses associated with phylogenetic ancestral reconstruction indicated that pathogenic bunyaviruses have evolved from an arthropod-specific ancestor [63]. Those data suggest possible adaptation of MSVs to vertebrates, following a spillover phenomenon (i.e. crossing of species barrier), and the emergence of other pathogenic viruses.
In this evolutionary context, in addition to bacterial microbiota, attention should be paid on MSVs as they may represent drivers of biological traits linked to the ability of the mosquito holobiont to transmit or not pathogens (see below).
Microbial interactions
Intra-microbial community interactions
Microbial interactions shape mosquito bacterial diversity and structure. To date, few data on interactions of gut microbes are available. Most of studies on microbe-microbe interactions focused on the influence of Wolbachia on the microbiome, or vice versa [88]. Wolbachia are intracellular, vertically transmitted bacterial symbionts that naturally infect many mosquito species and are known to manipulate their reproduction through cytoplasmic incompatibility (e.g. the offspring of infected males and uninfected females are not viable) [89]. In Anopheles mosquitoes, the identification of new factors modulating Wolbachia transmission in artificially transfected adults highlighted the importance of native mosquito microbiota and interaction with Wolbachia [90]. The disturbance of bacterial microbiota by antibiotic treatment reduced the ability of Anopheles to transmit the Wolbachia wAlbB strain (from Ae. albopictus) to offspring. Additional experiments combining high-throughput sequencing and oral infection of bacteria revealed that their native microbiota, especially Asaia, impeded vertical transmission of Wolbachia. This co-exclusion pattern between Wolbachia and Asaia is also found in Ae. albopictus and Cx. quinquefasciatus naturally bi-infected by both bacteria for which Asaia tissue tropism is restrained to the gut. Conversely, Asaia is also able to colonize reproductive organs and salivary glands in species uninfected by Wolbachia such as An. gambiae, An. stephensi and Ae. aegypti [91]. These observations suggest co-exclusion or competition between the two bacterial genera for reproductive organ colonization.
Bacteria could also interact with mosquito-specific viruses but so far only Wolbachia-ISV interactions have been considered [92]. An Ae. aegypti derived-cell line (Aag2) transfected with a Drosophila melanogaster-derived Wolbachia strain (wMelpop) was infected with the mosquito-specific CFAV flavivirus or with the Phasi Charoen-like bunyavirus. Molecular analysis revealed inhibition only for CFAV in Aag2-wMelpop cells, possibly related to the production of CFAV-specific small RNAs [92]. Indeed, MSVs could induce the RNA interference (RNAi) pathway by producing small interfering RNAs (siRNAs), as demonstrated for MBV [93]. The extent to which these interactions are genotype-by-genotype dependant is unknown. Moreover, it is also unknown whether Wolbachia can inhibit other single negative- or positive-strand RNA viruses using in vitro and/or in vivo systems. Wolbachia and MSV interactions in mosquitoes are nevertheless conceivable, as both are vertically transmitted intracellular organisms, suggesting co-localisation in reproductive organs. Their co-localisation at the cellular level still needs to be determined as it could impact the maintenance and transmission of both microbes in field bi-infected populations.
Microbial interference with transmitted pathogens
Mosquito infections with pathogens, including transmitted pathogens, trigger a complex crosstalk between different metabolic and immune pathways. Innate immune systems such as immune deficiency (Imd), the Toll, Janus kinases and signal transducers and activators of transcription (JAK-STAT) are activated in response to diverse microbes (viruses, bacteria, fungi or parasites) whilst RNAi modulates virus replication [41, 94, 95]. This general immune homeostatic response can be associated with the induction of autophagy, apoptosis as well as oxidative stress [40]. Transmitted pathogens can alter metabolisms through the modulation of stress-inducible genes involved for instance in redox and detoxifying enzyme metabolisms. However, molecular responses to transmitted pathogens are more complex if we consider the holobiont. The interplay between microbiota and mosquito immune system may result in an enhanced synergistic effect on the expression of effector molecules of the mosquito immune system, as previously reviewed [96].
Bacteria-mosquito-borne pathogen interference
As shown in Table 1, recent reports have stressed the importance of microbiota in the modulation of vector competence (ability of a susceptible mosquito to get infected by a pathogen, to support the pathogen’s replication and/or development and to transmit the pathogen to a susceptible vertebrate host)—this phenomenon is called microbial interference [97]. Multipartite interactions between the pathogen, the mosquito and its bacterial microbiota have become a major target for developing new control strategies in order to stop pathogen transmission and related epidemics.
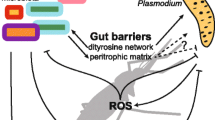
In An. gambiae mosquitoes, the presence of Escherichia coli, Serratia marcescens or Pseudomonas stutzeri resulted in a significant reduction in the prevalence and intensity of Plasmodium falciparum infection [98]. The abundance of Serratia was positively correlated with P. falciparum infection in both the midgut and salivary glands, suggesting a potential interaction between bacteria and the malaria parasite. The bacterium Chromobacterium was shown to increase An. gambiae resistance against P. falciparum [99]. It was suggested that the underlying mechanism behind interference was the production of cyanide by the bacterium. An. gambiae mosquitoes are also naturally colonized by the Enterobacter Esp_Z bacterial strain which inhibits the development of Plasmodium parasites prior to midgut colonization [100]. Specific genes associated with reactive oxygen species (ROS) production were found to be involved in mosquito midgut colonization by Esp_Z bacteria [101]. Recent studies have demonstrated the ability of gut bacteria to produce antiparasitic effectors that inhibit parasite growth. For instance, Chromobacterium was suggested to be a powerful immune elicitor since it increases mosquito immune gene expression. This feature, combined with its ability to rapidly invade the mosquito gut and reduce the lifespan of immature stages and adult mosquitoes, makes this bacterium a promising candidate for vector control applications [99]. Another study showed that the E. coli clone 444ST95 previously isolated from Anopheles mosquito midgut is able to markedly decrease the survival of these mosquitoes as well as the development of their Plasmodium parasites [102]. Hemolysin F or other toxins released by the bacterium are virulence factors associated with this effect [102]. Finally, a positive correlation between intensive antibiotic therapy in humans and increased risk of malaria transmission by An. gambiae mosquitoes has been suggested [103]. Indeed, antibiotics ingested by humans and circulating in their blood would enhance the susceptibility of blood-sucking An. gambiae females to malaria infection by disturbing their gut microbiota [103]. Despite the overall decrease in the microbial load in the mosquito, some specific changes have occurred in the microbial community, including a reduction in Serratia density, associated with an increase in Asaia abundance. Gendrin et al. [104] subsequently showed that critical parameters for the mosquito vector capacity, such as lifespan, permissiveness to P. falciparum, the mosquito microbiota composition and gut homeostasis were specific to the antibiotic treatment used. Moreover, microbiota disruption is closely related to gut homeostasis regulation [105]. For instance, the peritrophic matrix (PM) is a membrane that physically separates the blood meal from epithelium cells and plays a key role in regulation of mosquito gut homeostasis. Rodgers et al. [105] demonstrated that PM synthesis and integrity are related to gut microbiota. Importantly, microbial metabolites trigger the host oxidative response in mosquitoes, while maintaining redox homeostasis in the midgut [40].
As previously mentioned, the immune system is enhanced during microbial infections in mosquitoes. Stathopoulos et al. [106] characterized the molecular processes driving the mosquito immune response following infection by the enterobacterium S. marcescens as well as its consequences on transmission of the parasite. Peptidoglycan recognition proteins (PGRPs) are key regulators of the innate immune response [107]. These proteins specifically recognize microbe-associated molecular patterns (MAMPs) and therefore are influenced by microbiota variations. S. marcescens infections were found to result in the activation of some host genes, including PGRP-LC which activates the IMD/REL2 immune pathway involved in a reduction in Plasmodium infection. The authors also showed that some effectors of Anopheles immunity could impact the microbiota composition and load [107]. Similarly, Gendrin et al. [108] demonstrated that PGRP are important regulators of mosquito epithelial immunity and vector competence. PGRP-LA and PGRP-S2/ PGRP-S3 would be involved in the antiparasitic defense system, while PGRP-LB would promote mosquito permissiveness to P. falciparum. Immune system modulation by gut microbiota has also been demonstrated in Anopheles dirus mosquitoes naturally resistant to Plasmodium yoelii infection [109]. Its microbiota regulates the expression of a thioester-containing protein 1 (TEP1) following parasite infection. tep1 inactivation prevents microbiota from protecting the mosquito against parasitic infections [109]. This result suggests an important role of TEP1 related to microbiota in the refractoriness to P. yoelii infection. The immune regulation through microRNAs (miRNAs) can also modulate anti-Plasmodium defense and midgut microbiota [110]. The use of transgenic mosquitoes engineering to express miRNAs targeting endogenous-miRNAs offers new perspectives for the development of alternative malaria control.
There is abundant literature on Wolbachia-mediated interference of arbovirus transmission, as previously reviewed [111]. Here we will mainly focus on mosquito-borne pathogen interference with other members of bacterial microbiota. Indeed, few studies have examined the role of the microbiota in the modulation of arbovirus replication and transmission in mosquitoes. Interference mechanisms evidenced include production of bacterial metabolites with an anti-viral activity or nutrient competition between arbovirus and resident microbiota. As for Plasmodium, the bacterium Chromobacterium produces a metabolite with an anti-DENV activity in Ae. aegypti [99]. By comparing untreated and antibiotic-treated mosquitoes, Audsley et al. [112] demonstrated that the microbiota composition was not essential for blocking DENV in laboratory-reared Ae. aegypti. However, it seems that this assessment cannot be generalized since larval exposure to an Enterobacterium isolate Esp_ivi (genus-level classification undetermined, alternatively Salmonella, Escherichia or Shigella) resulted in decreased antibacterial activity in the hemolymph of Ae. aegypti females and reduced DENV dissemination titers [113]. Conversely, Serratia odorifera enhanced the susceptibility of Aedes mosquitoes to dengue and chikungunya viruses [114, 115]. A recent study also showed that Enterobacter ludwigii, Pseudomonas rhodesiae and Vagococcus salmoninarium isolated from Ae. albopictus could have an anti-viral effect on the La Crosse virus in vitro [116]. Interestingly, Novakova et al. [82] demonstrated that bacterial microbiota could be an important factor in the variability of vector competence in mosquitoes for WNV. The findings of other studies have also confirmed this observation. Ae. albopictus populations from France were shown to be more efficient in chikungunya virus dissemination compared to Vietnamese autochthonous relatives [117, 118]. In parallel, these invasive populations exhibited a reduction in their gut bacterial diversity compared to the Vietnamese populations [32].
Evidences underscore the importance of RNAi pathways in antiviral defense by the modulation of the expression of host or virus RNA-derived small RNAs, including siRNAs, PIWI interacting RNAs (piRNAs) and miRNAs. The siRNAs are the predominant virus-derived RNAs detected in infected mosquitoes, but piRNAs can also be detected in late/persistent infection [119, 120]. Zika virus modulates expression of both virus-derived siRNAs and piRNAs and host-derived miRNAs in Ae. aegypti [121]. Silencing of RNAi enzyme effectors such as Argonaute-2 (Ago-2) or Dicer 2/ R2D2 complex can promote flaviviruses (DENV) or alphaviruses (CHIKV, O'nyong'nyong virus [ONNV], Sindbis virus [SINV]) replication and/or transmission by Aedes and Anopheles mosquitoes [119, 120]. In a same way, RNAi-mediated knockdown of Imd and JAK-STAT pathways increased DENV replication in some Ae. aegypti lines [119]. Arboviruses can also induce protein synthesis involved in ROS production, carbohydrate or lipid metabolisms. In particular, midgut infection by DENV-2 and CHIKV triggered an antioxidant response through the production of proteins involved in detoxification. Other anti-viral responses such as apoptosis and autophagy can also contribute to innate antiviral immunity [119, 120], but detailed mechanisms involved remains poorly understood. Overall, the relative implications and crosstalk of these metabolic and immune pathways remain to be clarified and seem to be dependent on multiple factors, notably the transmitted virus-mosquito combination of the holobiont.
These crosstalk pathways become more complex as mosquito immune responses to transmitted pathogens can influence resident microbiota and vice versa. Only few data on anti-arbovirus responses in the context of holobiont are available. One of the most documented “system” is the Wolbachia-transfected mosquito. Wolbachia transfection in Wolbachia-free mosquitoes induces oxidative stress that activates the Toll pathway through the production of ROS [122]. The subsequent production of the antimicrobial peptides (AMPs) (cecropin and defensin) modulates DENV replication [122]. Wolbachia can also modify host-derived miRNA expression in wMelPop-CLA-transfected mosquitoes resulting in DENV interference [123]. In An. gambiae, Carissimo et al. [124] demonstrated that the siRNA pathway is not involved in midgut antiviral defense, but instead protects the post-midgut systemic compartment, which is the site of subsequent disseminated viral infection. While Anopheles microbiota hampers ONNV multiplication, viral infection is positively related to the microbiota. These data indicate distinct protective mechanisms that would allow an adapted response specific to each body compartment, infection stages and pathogens.
Overall, these results highlight the need to decipher genetic and molecular mechanisms of interactions in vector pathosystems and their impacts on pathogen transmission. Complex and potentially conflicting interactions in the gut mentioned above might have some direct implications in the design of new vector control strategies based on host microbiota manipulation. Caution is necessary to avoid using mosquito colonies that could potentially host new or circulating pathogenic agents in nature.
In accordance with the recent “pathobiome” concept, i.e. the pathogenic agent integrated within its biotic environment, disease transmission is modulated by interactions between host-transmitted pathogens and commensal and mutualistic microbes [125]. The gut microbiota would not be simply a passive commensal population with limited functions but an active sensor that would contribute to a local or systemic immune response, as previously demonstrated in Drosophila and Anopheles [126,127,128]. The mechanisms underlying these complex multipartite interactions [host-microbial community-environment] that modulate persistence, transmission and evolution of infectious pathogens remain to be deciphered. Understanding these interactions can open new avenues for controlling transmitted pathogen infection in vector insects.
Mosquito-associated eukaryotes and pathogen interactions
Some yeasts can directly, or via the host, interfere with parasites. For instance, Wickerhamomyces produces an anti-plasmodial toxin in vitro, while Penicillium chrysogenum promotes Plasmodium infection by suppressing the host innate immune response [129, 130]. Muturi et al. [43] recently showed that the Meyerozyma yeast dominated fungal communities in response to LACV infections in field-collected Ae. triseriatus and Ae. japonicus females. Secreted factors by the fungus Talaromyces downregulate digestive enzymes of its natural host Ae. aegypti that modulate DENV infection [131].
Mosquito-borne and mosquito-specific virus interference
Evidences of interference between MSVs and MBVs are inconsistent between studies. For instance, WNV replication was reported to decrease in the Ae. albopictus C6/36 cell line when co-infected with a CxFV strain isolated from Colorado [62] but not with one from Guatemala [132]. The CxFV Japan strain promotes dengue and Japanese encephalitis (JEEV) virus infection in Culex tritaeniorhynchus cells [133]. However, replication of JEEV and WNV decreased when co-infected with the Nhumirim flavivirus (NHUV) in C6/36 cells [134, 135]. Superinfection exclusion, which corresponds to an infected cell that is refractory to be secondarily infected with another closely related (or not) virus, could explain the observed interference effects. Different combinations of mosquito cell lines, MBV and MSV strains could also explain the disparities in the reported results.
In vivo experiments are essential to corroborate the interference between MSVs and MBVs, especially since in vitro studies have recurrently used the C6/36 cell line that is deficient in the RNAi immune pathway. Nasar et al. [64] demonstrated negative effects of EILV on CHIKV replication or dissemination in both in C7/10 Ae. albopictus cells and Ae. aegypti mosquitoes, respectively (Table 1) [64].
Most other in vivo studies have focused on CFxV and WNV interactions in Culex sp. but with contrasting results (Table 1). West Nile virus dissemination decreased when Cx. pipiens were intrathoracically infected with CxFV but not with NHUV [62, 135]. The same phenotype was observed in a CxFV-positive colony from Colorado in comparison to a CxFV-negative colony from Iowa [62]. Co-inoculation of WNV and CxFV led to a reduction of WNV transmission in Cx. quinquefasciatus populations from Honduras while no effect was observed when the experiment was reproduced with Floridians populations [132]. Moreover, a Cx. quinquefasciatus NHUV-positive colony was found to transmit WNV less efficiently at days 7 and 9 post-infection (pi) (Table 1) [135]. Viral interference in Culex sp. thus depends on the mosquito species, MSV and MBV strain combination and/or the mosquito infection status with ISVs. Overall, genotype-by-genotype-by-genotype interactions (mosquito-MSV-MBV), the status, time and mode (natural, oral or intrathoracic) of infection, as well as intra-microbial interactions in the mosquito holobiont, could explain the differences observed in these studies.
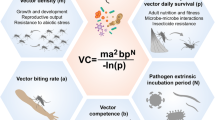
The evolutionary and phylogenetic aspects mentioned above indicate that the function and evolution of the holobiont-associated microbiota can shape keystone phenotypes such as the vectorial capacity, a key component of the dynamic of vector-borne disease transmission. This also underscores the need to decipher factors and mechanisms involved in host restriction or permissiveness and virus interference in the development of new bio-control strategies or vaccines [65].
Microbiota-based control strategies: promising perspectives?
To date, paratransgenesis approach using symbionts to produce molecules that inhibit pathogen development and/or transmission has been mainly restricted to Anopheles mosquitoes to prevent Plasmodium development [136, 137]. The most promising candidates are the bacteria Pantoea agglomerans and Asaia spp. [138, 139]. Paratransgenic strains of Asaia bogorensis were recently engineered using induced-fusion system of antiplasmodial effectors and bacterial secreted putative genes (encoding a siderophore receptor protein and YVTN beta-propeller repeat proteins) [140]. These effectors expressed in An. stephensi significantly inhibit P. berghei development [140]. Mancini et al. [139] evaluated paratransgenesis efficiency in semi-field conditions with genetically modified strains of Asaia and demonstrated their ability to colonize both An. stephensi and An. gambiae mosquito populations. Paratransgenesis still needs to be assessed in Aedes and Culex mosquitoes to impede arbovirus replication and/or transmission. The use and persistence in the environment of microbes that induce mosquito immune response to transmitted viruses (RNA interference) or produce toxins are currently unknown. Complementary data and semi-field studies are needed to evaluate the feasibility of these strategies to control or suppress mosquito populations. The development and implementation of paratransgenesis needs to be carefully evaluated to ensure safety issues for both humans and the environment, as reviewed in Touré et al. [141] and Gabrieli et al. [136].
Currently, phenotypes such as viral and parasite interferences induced by several Wolbachia strains (as wMel and wMelpop of Drosophila transfected in mosquitoes) and cytoplasmic incompatibility are used to control mosquito populations [142, 143]. Field applications highlighted the efficiency of using Wolbachia transfected in Ae. aegypti to block DENV transmission [144]. Development of future Wolbachia-based strategies to prevent malaria transmission are promising, as the first known Wolbachia-Anopheles associations were recently identified [145, 146] and that Plasmodium development seems to be related to the presence of Wolbachia in Anopheles coluzzii field populations [146].
The incompatible insect technique (IIT) can also be combined with the sterile insect technique (SIT) to improve vector control [147]. For instance, Zhang et al. [148] studied the effect of wPip (Wolbachia strain from Cx. pipiens) on Ae. albopictus that is originally naturally bi-infected by wAlbA and wAlbB. They showed that wPip has a low effect on mosquito fitness. Thereby, the authors conclude that the competitiveness with natural populations combined with the strong cytoplasmic incompatibility of this triple Wolbachia-infected Ae. albopictus line supports its use in SIT/IIT strategies to control Ae. albopictus populations.
To conclude, field applications of these different strategies, combined or not, need properly ethical, ecological and social issues, especially if the strategies are related to the release of symbiotically modified mosquitoes [136]. Particular attention should be given to ecological and evolutionary aspects. More data will improve our understanding on the implications, the outcome and the environmental sustainability of these engineering systems, notably for potential gene flows (HGT), the emergence of resistance in mosquito populations and/or the accidental spread to non-target species. In this context, future innovative control strategies should favour the reduction of mosquito density below threshold levels of disease transmission rather than the population-replacement strategy.
Future challenges in mosquito holobiont investigations
Some key components and processes should be determined to gain further insight into the mosquito holobiont. In particular, little is known on the (co)evolutionary aspects of mosquito holobiont functioning, including the involvement of associated-microbiota in adaptation and speciation [149, 150]. Mosquito invasive capacities and global spread could be emphasized through its microbial compartment. Mutualistic symbionts are notably a key factor for the ecological success and adaptation to novel environments of some insect species. Most striking examples are the symbiont-mediated insecticide resistance in the bean bug Riptortus pedestris [151] or thermal tolerance of insect aphids due to a mutation in their bacterial symbiont Buchnera aphidicola [152]. Besides, microbe genomes evolve relatively rapidly through mutations, recombinations, rearrangements, horizontal transfers and hybridizations. Consequently, the microbial hologenome responds differentially and more quickly to selective environmental pressure than the host’s genome. These rapidly evolving characteristics in the symbiotic community could markedly contribute to extending the host and/or their microbiota-associated phenotypes. Moreover, it is also essential to enhance knowledge on holobiont (microbiota and host) genome architecture and expression via high throughput OMICS strategies (genomics, transcriptomics, proteomics, metabolomics, etc.) in order to shape functional networks and obtain a deeper understanding on the multipartite interactions involved. Greater effort will thus be needed in developing ad hoc protocols and tools.
Conclusions
The holobiont concept has raised considerable debate on the functioning and evolution of organisms with the aim of gaining greater insight into important biological issues. Host-associated microbiota involves a complex network of cooperation and competition, which makes it difficult to understand the role of each microorganism. In mosquitoes, recent findings have given rise to new hypotheses on holobiont functioning and dynamics, with symbiotic interactions being the keystone of the vector pathosystem. Intra-community microbial interaction dynamics within the mosquito holobiont as well as the molecular mechanisms involved in these multipartite interactions have yet to be investigated in detail. A technical issue is that deciphering the intricate interactions between microbes, host and environment is challenging. Such investigations could provide new targets and tools for vector-borne pathogen control. Moreover, we believe that studying the mosquito holobiont in the evolutionary context (experimentally, modeling, etc.) could help to predict, for instance, areas suitable for mosquito adaptation or even outbreaks, and thus lead to the development of strategies to avoid outbreaks, epidemics and epizootic diseases.
Abbreviations
- AeFV:
-
Aedes flavivirus
- AMPs:
-
Antimicrobial peptides
- Bti:
-
Bacillus thuringiensis subspecies israelensis
- CFAV:
-
Cell fusion agent virus
- CHIKV:
-
Chikungunya virus
- CxFV:
-
Culex flavivirus
- DENV:
-
Dengue virus
- EILV:
-
Eilat virus
- EVEs:
-
Endogenous viral elements
- HGTs:
-
Horizontal gene transfers
- HIFs:
-
Hypoxia-induced transcription factors
- IIT:
-
Incompatible insect technique
- Imd:
-
Immune deficiency
- ISVs:
-
Insect-specific viruses
- JAK-STAT:
-
Janus kinases and signal transducers and activators of transcription
- JEEV:
-
Japanese encephalitis
- KRV:
-
Kamiti River virus
- LACV:
-
La Crosse virus
- MAMPs:
-
Microbe-associated molecular patterns
- MBVs:
-
Mosquito-borne viruses
- miRNAs:
-
MicroRNAs
- MSVs:
-
Mosquito-specific viruses
- NHUV:
-
Nhumirim virus
- ONNV:
-
O'nyong'nyong virus
- OTUs:
-
Operational taxonomical units
- PGRPs:
-
Peptidoglycan recognition proteins
- pi:
-
Post-infection
- piRNAs:
-
PIWI interacting RNAs
- PM:
-
Peritrophic matrix
- RNAi:
-
RNA interference
- SINV:
-
Sindbis virus
- SIT:
-
Sterile insect technique
- siRNA:
-
Small interfering RNAs
- TEP:
-
Thioester-containing protein
- WHO:
-
World Health Organization
- WNV:
-
West Nile virus
- YFV:
-
Yellow fever virus
- ZIKV:
-
Zika virus
References
Rosenberg E, Koren O, Reshef L, Efrony R, Zilber-Rosenberg I. The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol. 2007;5:355–62.
Cresci GA, Bawden E. Gut microbiome: what we do and don’t know. Nutr. Clin. Pract. Off. Publ. Am. Soc. Parenter. Enter. Nutr. 2015;30:734–46.
Douglas AE. Multiorganismal insects: diversity and function of resident microorganisms. Annu Rev Entomol. 2015;60:17–34.
Snyder AK, Rio RVM. Interwoven biology of the tsetse holobiont. J Bacteriol. 2013;195:4322–30.
Caragata EP, Dutra HLC, Moreira LA. Exploiting intimate relationships: controlling mosquito-transmitted disease with Wolbachia. Trends Parasitol. 2016;32:207–18.
Erkosar B, Storelli G, Defaye A, Leulier F. Host-intestinal microbiota mutualism: “learning on the fly.”. Cell Host Microbe. 2013;13:8–14.
WHO | Executive summary [Internet]. WHO. [cited 2017 Aug 7]. Available from: http://www.who.int/whr/1996/media_centre/executive_summary1/en/index9.html.
WHO | Malaria [Internet]. WHO. [cited 2017 Aug 7]. Available from: http://www.who.int/mediacentre/factsheets/fs094/en/.
Organization WH. Progress report 2000–2009 and strategic plan 2010–2020 of the global programme to eliminate lymphatic filariasis: halfway towards eliminating lymphatic filariasis. Rapport de situation 2000–2009 et plan stratégique 2010–2020 du programme mondial pour l’élimination de la filariose lymphatique: à mi-parcours vers l’élimination de cette maladie [internet]. 2010 [cited 2017 Aug 14]; Available from: http://www.who.int/iris/handle/10665/44473
Patterson J, Sammon M, Garg M. Dengue, Zika and chikungunya: emerging arboviruses in the new world. West J Emerg Med. 2016;17:671–9.
Weaver SC, Charlier C, Vasilakis N, Lecuit M. Zika, chikungunya, and other emerging vectorborne viral diseases. Annu Rev Med. 2018;69:1:395–408.
Baud D, Gubler DJ, Schaub B, Lanteri MC, Musso D. An update on Zika virus infection. Lancet. 2017;390(10107):2099–2109.
Schaffner F, Medlock JM, Van Bortel W. Public health significance of invasive mosquitoes in Europe. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2013;19:685–92.
Sutherst RW. Global change and human vulnerability to vector-borne diseases. Clin Microbiol Rev. 2004;17:136–73.
Ricci I, Damiani C, Capone A, DeFreece C, Rossi P, Favia G. Mosquito/microbiota interactions: from complex relationships to biotechnological perspectives. Curr Opin Microbiol. 2012;15:278–84.
Minard G, Mavingui P, Moro CV. Diversity and function of bacterial microbiota in the mosquito holobiont. Parasit Vectors. 2013;6:146.
Duguma D, Hall MW, Smartt CT, Neufeld JD. Effects of Organic Amendments on Microbiota Associated with the Culex nigripalpus Mosquito Vector of the Saint Louis Encephalitis and West Nile Viruses. mSphere. 2017;2:e00387–16.
Buck M, Nilsson LKJ, Brunius C, Dabiré RK, Hopkins R, Terenius O. Bacterial associations reveal spatial population dynamics in Anopheles gambiae mosquitoes. Sci Rep. 2016;6:22806.
Coon KL, Brown MR, Strand MR. Mosquitoes host communities of bacteria that are essential for development but vary greatly between local habitats. Mol Ecol. 2016;25:5806–26.
Gimonneau G, Tchioffo MT, Abate L, Boissière A, Awono-Ambéné PH, Nsango SE, et al. Composition of Anopheles coluzzii and Anopheles gambiae microbiota from larval to adult stages. Infect Genet Evol J Mol Epidemiol Evol Genet Infect Dis. 2014;28:715–24.
Kim C-H, Lampman RL, Muturi EJ. Bacterial communities and midgut microbiota associated with mosquito populations from waste tires in East-Central Illinois. J Med Entomol. 2015;52:63–75.
Dada N, Jumas-Bilak E, Manguin S, Seidu R, Stenström T-A, Overgaard HJ. Comparative assessment of the bacterial communities associated with Aedes aegypti larvae and water from domestic water storage containers. Parasit Vectors. 2014;7:391.
Duguma D, Hall MW, Rugman-Jones P, Stouthamer R, Terenius O, Neufeld JD, et al. Developmental succession of the microbiome of Culex mosquitoes. BMC Microbiol. 2015;15:140.
Yadav KK, Datta S, Naglot A, Bora A, Hmuaka V, Bhagyawant S, et al. Diversity of cultivable midgut microbiota at different stages of the Asian Tiger mosquito, Aedes albopictus from Tezpur. India PloS One. 2016;11:e0167409.
Coon KL, Vogel KJ, Brown MR, Strand MR. Mosquitoes rely on their gut microbiota for development. Mol Ecol. 2014;23:2727–39.
Moll RM, Romoser WS, Modrzakowski MC, Moncayo AC, Lerdthusnee K. Meconial peritrophic membranes and the fate of midgut bacteria during mosquito (Diptera: Culicidae) metamorphosis. J Med Entomol. 2001;38:29–32.
Segata N, Baldini F, Pompon J, Garrett WS, Truong DT, Dabiré RK, et al. The reproductive tracts of two malaria vectors are populated by a core microbiome and by gender- and swarm-enriched microbial biomarkers. Sci Rep. 2016;6:24207.
Astudillo-García C, Bell JJ, Webster NS, Glasl B, Jompa J, Montoya JM, et al. Evaluating the core microbiota in complex communities: a systematic investigation. Environ Microbiol. 2017;19:1450–62.
David MR, Santos LMBD, Vicente ACP, Maciel-de-Freitas R. Effects of environment, dietary regime and ageing on the dengue vector microbiota: evidence of a core microbiota throughout Aedes aegypti lifespan. Mem Inst Oswaldo Cruz. 2016;111:577–87.
Ngo CT, Aujoulat F, Veas F, Jumas-Bilak E, Manguin S. Bacterial diversity associated with wild caught Anopheles mosquitoes from Dak Nong Province, Vietnam using culture and DNA fingerprint. PLoS One. 2015;10(3):e0118634.
Ngo CT, Romano-Bertrand S, Manguin S, Jumas-Bilak E. Diversity of the bacterial microbiota of Anopheles mosquitoes from Binh Phuoc Province, Vietnam. Front Microbiol. 2016;7:2095.
Minard G, Tran FH, Van VT, Goubert C, Bellet C, Lambert G, et al. French invasive Asian tiger mosquito populations harbor reduced bacterial microbiota and genetic diversity compared to Vietnamese autochthonous relatives. Front Microbiol. 2015;6:970.
Tchioffo MT, Boissière A, Abate L, Nsango SE, Bayibéki AN, Awono-Ambéné PH, et al. Dynamics of bacterial community composition in the malaria mosquito’s epithelia. Front Microbiol. 2016;6:1500.
Leung MHY, Wilkins D, Lee PKH. Insights into the pan-microbiome: skin microbial communities of Chinese individuals differ from other racial groups. Sci Rep. 2015;5:srep11845.
Yadav KK, Bora A, Datta S, Chandel K, Gogoi HK, Prasad GBKS, et al. Molecular characterization of midgut microbiota of Aedes albopictus and Aedes aegypti from Arunachal Pradesh, India. Parasit Vectors. 2015;8:641.
Muturi EJ, Kim C-H, Bara J, Bach EM, Siddappaji MH. Culex pipiens and Culex restuans mosquitoes harbor distinct microbiota dominated by few bacterial taxa. Parasit Vectors. 2016;9:18.
Chandler JA, Liu RM, Bennett SN. RNA shotgun metagenomic sequencing of northern California (USA) mosquitoes uncovers viruses, bacteria, and fungi. Front Microbiol. 2015;6:185.
Andrews ES, Xu G, Rich SM. Microbial communities within field-collected Culiseta melanura and Coquillettidia perturbans. Med Vet Entomol. 2014;28:125–32.
Muturi EJ, Ramirez JL, Rooney AP, Kim C-H. Comparative analysis of gut microbiota of mosquito communities in Central Illinois. PLoS Negl Trop Dis. 2017;11(2):e0005377.
Champion CJ, Xu J. The impact of metagenomic interplay on the mosquito redox homeostasis. Free Radic Biol Med. 2017;105:79–85.
van Tol S, Dimopoulos G. Chapter nine—influences of the mosquito microbiota on vector competence. In: Raikhel AS, editor. Advances in insect physiology, vol. 51; 2016. p. 243–91.
Zouache K, Michelland RJ, Failloux A-B, Grundmann GL, Mavingui P. Chikungunya virus impacts the diversity of symbiotic bacteria in mosquito vector. Mol Ecol. 2012;21:2297–309.
Muturi EJ, Bara JJ, Rooney AP, Hansen AK. Midgut fungal and bacterial microbiota of Aedes triseriatus and Aedes japonicus shift in response to La Crosse virus infection. Mol Ecol. 2016;25:4075–90.
Ramirez JL, Souza-Neto J, Torres Cosme R, Rovira J, Ortiz A, Pascale JM, et al. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl Trop Dis. 2012;6(3):e1561.
Villegas LEM, Campolina TB, Barnabe NR, Orfano AS, Chaves BA, Norris DE, et al. Zika virus infection modulates the bacterial diversity associated with Aedes aegypti as revealed by metagenomic analysis. PLoS One. 2018;13:e0190352.
Short SM, Mongodin EF, MacLeod HJ, Talyuli OAC, Dimopoulos G. Amino acid metabolic signaling influences Aedes aegypti midgut microbiome variability. PLoS Negl Trop Dis. 2017;11:e0005677.
Moureau G, Ninove L, Izri A, Cook S, De Lamballerie X, Charrel RN. Flavivirus RNA in phlebotomine sandflies. Vector Borne Zoonotic Dis Larchmt N. 2010;10:195–7.
Stollar V, Thomas VL. An agent in the Aedes aegypti cell line (Peleg) which causes fusion of Aedes albopictus cells. Virology. 1975;64:367–77.
Sang RC, Gichogo A, Gachoya J, Dunster MD, Ofula V, Hunt AR, et al. Isolation of a new flavivirus related to cell fusing agent virus (CFAV) from field-collected flood-water Aedes mosquitoes sampled from a dambo in Central Kenya. Arch Virol. 2003;148:1085–93.
Hoshino K, Isawa H, Tsuda Y, Yano K, Sasaki T, Yuda M, et al. Genetic characterization of a new insect flavivirus isolated from Culex pipiens mosquito in Japan. Virology. 2007;359:405–14.
Vasilakis N, Tesh RB. Insect-specific viruses and their potential impact on arbovirus transmission. Curr Opin Virol. 2015;15:69–74.
Bolling BG, Weaver SC, Tesh RB, Vasilakis N. Insect-specific virus discovery: significance for the arbovirus community. Viruses. 2015;7:4911–28.
Roundy CM, Azar SR, Rossi SL, Weaver SC, Vasilakis N. Insect-specific viruses: a historical overview and recent developments. Adv Virus Res. 2017;98:119–46.
Calzolari M, Zé-Zé L, Vázquez A, Sánchez Seco MP, Amaro F, Dottori M. Insect-specific flaviviruses, a worldwide widespread group of viruses only detected in insects. Infect Genet Evol. 2016;40:381–8.
Blitvich BJ, Firth AE. Insect-specific flaviviruses: a systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses. 2015;7:1927–59.
Nasar F, Palacios G, Gorchakov RV, Guzman H, Da Rosa APT, Savji N, et al. Eilat virus, a unique alphavirus with host range restricted to insects by RNA replication. Proc Natl Acad Sci U S A. 2012;109:14622–7.
Hermanns K, Zirkel F, Kopp A, Marklewitz M, Rwego IB, Estrada A, et al. Discovery of a novel alphavirus related to Eilat virus. J Gen Virol. 2017;98:43–9.
Saiyasombat R, Bolling BG, Brault AC, Bartholomay LC, Blitvich BJ. Evidence of efficient transovarial transmission of Culex flavivirus by Culex pipiens (Diptera: Culicidae). J Med Entomol. 2011;48:1031–8.
Haddow AD, Guzman H, Popov VL, Wood TG, Widen SG, Haddow AD, et al. First isolation of Aedes flavivirus in the western hemisphere and evidence of vertical transmission in the mosquito Aedes (Stegomyia) albopictus (Diptera: Culicidae). Virology. 2013;440:134–9.
Roossinck MJ. Plants, viruses and the environment: ecology and mutualism. Virology. 2015;479–480:271–7.
Halbach R, Junglen S, van Rij RP. Mosquito-specific and mosquito-borne viruses: evolution, infection, and host defense. Curr Opin Insect Sci. 2017;22:16–27.
Bolling BG, Olea-Popelka FJ, Eisen L, Moore CG, Blair CD. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology. 2012;427:90–7.
Marklewitz M, Zirkel F, Kurth A, Drosten C, Junglen S. Evolutionary and phenotypic analysis of live virus isolates suggests arthropod origin of a pathogenic RNA virus family. Proc Natl Acad Sci U S A. 2015;112:7536–41.
Nasar F, Gorchakov RV, Tesh RB, Weaver SC. Eilat virus host range restriction is present at multiple levels of the virus life cycle. J Virol. 2015;89:1404–18.
Erasmus JH, Auguste AJ, Kaelber JT, Luo H, Rossi SL, Fenton K, et al. A chikungunya fever vaccine utilizing an insect-specific virus platform. Nat Med. 2017;23:192–9.
Tree MO, McKellar DR, Kieft KJ, Watson AM, Ryman KD, Conway MJ. Insect-specific flavivirus infection is restricted by innate immunity in the vertebrate host. Virology. 2016;497:81–91.
Belda E, Coulibaly B, Fofana A, Beavogui AH, Traore SF, Gohl DM, et al. Preferential suppression of Anopheles gambiae host sequences allows detection of the mosquito eukaryotic microbiome. Sci Rep. 2017;7:3241.
Steyn A, Roets F, Botha A. Yeasts associated with Culex pipiens and Culex theileri mosquito larvae and the effect of selected yeast strains on the ontogeny of Culex pipiens. Microb Ecol. 2016;71:747–60.
Bozic J, Capone A, Pediconi D, Mensah P, Cappelli A, Valzano M, et al. Mosquitoes can harbour yeasts of clinical significance and contribute to their environmental dissemination. Environ Microbiol Rep. 2017;9(5):642–48.
Lavialle C, Cornelis G, Dupressoir A, Esnault C, Heidmann O, Vernochet C, et al. Paleovirology of “syncytins”, retroviral env genes exapted for a role in placentation. Philos Trans R Soc Lond Ser B Biol Sci. 2013;368:20120507.
Mitraka E, Stathopoulos S, Siden-Kiamos I, Christophides GK, Louis C. Asaia accelerates larval development of Anopheles gambiae. Pathog Glob Health. 2013;107:305–11.
Díaz-Nieto LM, D Alessio C, Perotti MA, Berón CM. Culex pipiens development is greatly influenced by native bacteria and exogenous yeast. PLoS One. 2016;11:e0153133.
Vogel KJ, Valzania L, Coon KL, Brown MR, Strand MR. Transcriptome sequencing reveals large-scale changes in axenic Aedes aegypti larvae. PLoS Negl Trop Dis. 2017;11:e0005273.
Coon KL, Valzania L, McKinney DA, Vogel KJ, Brown MR, Strand MR. Bacteria-mediated hypoxia functions as a signal for mosquito development. Proc Natl Acad Sci U S A. 2017;114:E5362–9.
Valzania L, Coon KL, Vogel KJ, Brown MR, Strand MR. Hypoxia-induced transcription factor signaling is essential for larval growth of the mosquito Aedes aegypti. Proc Natl Acad Sci U S A. 2018;115:457–65.
Pennington MJ, Prager SM, Walton WE, Trumble JT. Culex quinquefasciatus larval microbiomes vary with instar and exposure to common wastewater contaminants. Sci Rep. 2016;6:21969.
Coon KL, Brown MR, Strand MR. Gut bacteria differentially affect egg production in the anautogenous mosquito Aedes aegypti and facultatively autogenous mosquito Aedes atropalpus (Diptera: Culicidae). Parasit Vectors. 2016;9:375.
Patil CD, Borase HP, Salunke BK, Patil SV. Alteration in Bacillus thuringiensis toxicity by curing gut flora: novel approach for mosquito resistance management. Parasitol Res. 2013;112:3283–8.
Caccia S, Di Lelio I, La Storia A, Marinelli A, Varricchio P, Franzetti E, et al. Midgut microbiota and host immunocompetence underlie Bacillus thuringiensis killing mechanism. Proc Natl Acad Sci U S A. 2016;113:9486–91.
Wei G, Lai Y, Wang G, Chen H, Li F, Wang S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc Natl Acad Sci U S A. 2017;114:5994–9.
Minard G, Tran Van V, Tran FH, Melaun C, Klimpel S, Koch LK, et al. Identification of sympatric cryptic species of Aedes albopictus subgroup in Vietnam: new perspectives in phylosymbiosis of insect vector. Parasit Vectors. 2017;10:276.
Novakova E, Woodhams DC, Rodríguez-Ruano SM, Brucker RM, Leff JW, Maharaj A, et al. Mosquito microbiome dynamics, a background for prevalence and seasonality of West Nile virus. Front Microbiol. 2017;8:526.
Klasson L, Kambris Z, Cook PE, Walker T, Sinkins SP. Horizontal gene transfer between Wolbachia and the mosquito Aedes aegypti. BMC Genomics. 2009;10:33.
Hou Q, He J, Yu J, Ye Y, Zhou D, Sun Y, et al. A case of horizontal gene transfer from Wolbachia to Aedes albopictus C6/36 cell line. Mob Genet Elem. 2014;4:e28914.
Acuña R, Padilla BE, Flórez-Ramos CP, Rubio JD, Herrera JC, Benavides P, et al. Adaptive horizontal transfer of a bacterial gene to an invasive insect pest of coffee. Proc Natl Acad Sci U S A. 2012;109:4197–202.
Li C-X, Shi M, Tian J-H, Lin X-D, Kang Y-J, Chen L-J, et al. Unprecedented genomic diversity of RNA viruses in arthropods reveals the ancestry of negative-sense RNA viruses. eLife. 2015;4:e05378.
Olson KE, Bonizzoni M. Nonretroviral integrated RNA viruses in arthropod vectors: an occasional event or something more? Curr Opin Insect Sci. 2017;22:45–53.
Audsley MD, Seleznev A, Joubert DA, Woolfit M, O’Neill SL, EA MG. Wolbachiainfection alters the relative abundance of resident bacteria in adult Aedes aegypti mosquitoes, but not larvae. Mol Ecol. 2018;27(1):297–309.
Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008;6:741–51.
Hughes GL, Dodson BL, Johnson RM, Murdock CC, Tsujimoto H, Suzuki Y, et al. Native microbiome impedes vertical transmission of Wolbachia in Anopheles mosquitoes. Proc Natl Acad Sci U S A. 2014;111:12498–503.
Rossi P, Ricci I, Cappelli A, Damiani C, Ulissi U, Mancini MV, et al. Mutual exclusion of Asaia and Wolbachia in the reproductive organs of mosquito vectors. Parasit Vectors. 2015;8:278.
Schnettler E, Sreenu VB, Mottram T, McFarlane M. Wolbachia restricts insect-specific flavivirus infection in Aedes aegypti cells. J Gen Virol. 2016;97:3024–9.
Cheng G, Liu Y, Wang P, Xiao X. Mosquito defense strategies against viral infection. Trends Parasitol. 2016;32:177–86.
Hegde S, Rasgon JL, Hughes GL. The microbiome modulates arbovirus transmission in mosquitoes. Curr Opin Virol. 2015;15:97–102.
Bartholomay LC, Michel K. Mosquito immunobiology: the intersection of vector health and vector competence. Annu Rev Entomol. 2018;63:145–67.
Saraiva RG, Kang S, Simões ML, Angleró-Rodríguez YI, Dimopoulos G. Mosquito gut antiparasitic and antiviral immunity. Dev Comp Immunol. 2016;64:53–64.
Weiss B, Aksoy S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011;27:514–22.
Tchioffo MT, Boissière A, Churcher TS, Abate L, Gimonneau G, Nsango SE, et al. Modulation of malaria infection in Anopheles gambiae mosquitoes exposed to natural midgut Bacteria. PLoS One. 2013;8(12):e81663.
Ramirez JL, Short SM, Bahia AC, Saraiva RG, Dong Y, Kang S, et al. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has Entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014;10(10):e1004398.
Cirimotich CM, Dong Y, Clayton AM, Sandiford SL, Souza-Neto JA, Mulenga M, et al. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science. 2011;332:855–8.
Dennison NJ, Saraiva RG, Cirimotich CM, Mlambo G, Mongodin EF, Dimopoulos G. Functional genomic analyses of Enterobacter, Anopheles and Plasmodium reciprocal interactions that impact vector competence. Malar J. 2016;15(1):425.
Tchioffo MT, Abate L, Boissière A, Nsango SE, Gimonneau G, Berry A, et al. An epidemiologically successful Escherichia coli sequence type modulates Plasmodium falciparum infection in the mosquito midgut. Infect Genet Evol. 2016;43:22–30.
Gendrin M, Rodgers FH, Yerbanga RS, Ouédraogo JB, Basáñez M-G, Cohuet A, et al. Antibiotics in ingested human blood affect the mosquito microbiota and capacity to transmit malaria. Nat Commun. 2015;6:5921.
Gendrin M, Yerbanga RS, Ouedraogo JB, Lefèvre T, Cohuet A, Christophides GK. Differential effects of azithromycin, doxycycline, and cotrimoxazole in ingested blood on the vectorial capacity of malaria mosquitoes. Open Forum Infect Dis. 2016;3:ofw074.
Rodgers FH, Gendrin M, Wyer CAS, Christophides GK. Microbiota-induced peritrophic matrix regulates midgut homeostasis and prevents systemic infection of malaria vector mosquitoes. PLoS Pathog. 2017;13(5):e1006391.
Stathopoulos S, Neafsey DE, Lawniczak MKN, Muskavitch MAT, Christophides GK. Genetic dissection of Anopheles gambiae gut epithelial responses to Serratia marcescens. PLoS Pathog. 2014;10(3):e1003897.
Gendrin M, Zaidman-Rémy A, Broderick NA, Paredes J, Poidevin M, Roussel A, et al. Functional analysis of PGRP-LA in Drosophila immunity. PLoS One. 2013;8:e69742.
Gendrin M, Turlure F, Rodgers FH, Cohuet A, Morlais I, Christophides GK. The peptidoglycan recognition proteins PGRPLA and PGRPLB regulate Anopheles immunity to Bacteria and affect infection by Plasmodium. J Innate Immun. 2017;9:333–42.
Wang Y, Wang Y, Zhang J, Xu W, Zhang J, Huang FS. Ability of TEP1 in intestinal flora to modulate natural resistance of Anopheles dirus. Exp Parasitol. 2013;134:460–5.
Dennison NJ, BenMarzouk-Hidalgo OJ, Dimopoulos G. MicroRNA-regulation of Anopheles gambiae immunity to Plasmodium falciparum infection and midgut microbiota. Dev Comp Immunol. 2015;49:170–8.
Kamtchum-Tatuene J, Makepeace BL, Benjamin L, Baylis M, Solomon T. The potential role of Wolbachia in controlling the transmission of emerging human arboviral infections. Curr Opin Infect Dis. 2017;30:108–16.
Audsley MD, Ye YH, McGraw EA. The microbiome composition of Aedes aegypti is not critical for Wolbachia-mediated inhibition of dengue virus. PLoS Negl Trop Dis. 2017;11:e0005426.
Dickson LB, Jiolle D, Minard G, Moltini-Conclois I, Volant S, Ghozlane A, et al. Carryover effects of larval exposure to different environmental bacteria drive adult trait variation in a mosquito vector. Sci Adv. 2017;3:e1700585.
Apte-Deshpande A, Paingankar M, Gokhale MD, Deobagkar DN. Serratia odorifera a midgut inhabitant of Aedes aegypti mosquito enhances its susceptibility to dengue-2 virus. PLoS One. 2012;7:e40401.
Apte-Deshpande AD, Paingankar MS, Gokhale MD, Deobagkar DN. Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for chikungunya virus. Indian J Med Res. 2014;139:762–8.
Dennison NJ, Jupatanakul N, Dimopoulos G. The mosquito microbiota influences vector competence for human pathogens. Curr Opin Insect Sci. 2014;3:6–13.
Vega-Rua A, Zouache K, Caro V, Diancourt L, Delaunay P, Grandadam M, et al. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the southeast of France. PLoS One. 2013;8(3):e59716.
Zouache K, Fontaine A, Vega-Rua A, Mousson L, Thiberge J-M, Lourenco-De-Oliveira R, Caro V, Lambrechts L, Failloux A-B. Three-way interactions between mosquito population, viral strain and temperature underlying chikungunya virus transmission potential. Proc R Soc B Biol Sci. 2014;281(1792).
Sim S, Jupatanakul N, Dimopoulos G. Mosquito immunity against arboviruses. Viruses. 2014;6:4479–504.
Blair CD, Olson KE. The role of RNA interference (RNAi) in arbovirus-vector interactions. Viruses. 2015;7:820–43.
Etebari K, Hegde S, Saldaña MA, Widen SG, Wood TG, Asgari S, et al. Global transcriptome analysis of Aedes aegypti mosquitoes in response to Zika virus infection. mSphere. 2017;2:e00456–17.
Pan X, Zhou G, Wu J, Bian G, Lu P, Raikhel AS, et al. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc Natl Acad Sci U S A. 2012;109:E23–31.
Pan X, Thiem S, Xi Z. Chapter 3—Wolbachia-mediated immunity induction in mosquito vectors. In: Wikel SK, Aksoy S, Dimopoulos G, editors. Arthropod Vector: Controller of Disease Transmission. Vol. 1. Cambridge: Academic Press. 2017. p. 35–58.
Carissimo G, Pondeville E, McFarlane M, Dietrich I, Mitri C, Bischoff E, et al. Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc Natl Acad Sci U S A. 2015;112:E176–85.
Vayssier-Taussat M, Albina E, Citti C, Cosson J-F, Jacques M-A, Lebrun M-H, et al. Shifting the paradigm from pathogens to pathobiome: new concepts in the light of meta-omics. Front Cell Infect Microbiol. 2014;4:29.
Ryu J-H, Kim S-H, Lee H-Y, Bai JY, Nam Y-D, Bae J-W, et al. Innate immune homeostasis by the homeobox gene caudal and commensal-gut mutualism in Drosophila. Science. 2008;319:777–82.
Rodrigues J, Brayner FA, Alves LC, Dixit R, Barillas-Mury C. Hemocyte differentiation mediates innate immune memory in Anopheles gambiae mosquitoes. Science. 2010;329:1353–5.
Wu S-C, Liao C-W, Pan R-L, Juang J-L. Infection-induced intestinal oxidative stress triggers organ-to-organ immunological communication in Drosophila. Cell Host Microbe. 2012;11:410–7.
Valzano M, Cecarini V, Cappelli A, Capone A, Bozic J, Cuccioloni M, et al. A yeast strain associated to Anopheles mosquitoes produces a toxin able to kill malaria parasites. Malar J. 2016;15:21.
Angleró-Rodríguez YI, Blumberg BJ, Dong Y, Sandiford SL, Pike A, Clayton AM, et al. A natural Anopheles-associated Penicillium chrysogenum enhances mosquito susceptibility to Plasmodium infection. Sci Rep. 2016;6:34084.
Angleró-Rodríguez YI, Talyuli OA, Blumberg BJ, Kang S, Demby C, Shields A, et al. An Aedes aegypti-associated fungus increases susceptibility to dengue virus by modulating gut trypsin activity. elife. 2017;6:e28844.
Kent RJ, Crabtree MB, Miller BR. Transmission of West Nile virus by Culex quinquefasciatus say infected with Culex Flavivirus Izabal. PLoS Negl Trop Dis. 2010;4:e671.
Kuwata R, Isawa H, Hoshino K, Sasaki T, Kobayashi M, Maeda K, et al. Analysis of mosquito-borne Flavivirus superinfection in Culex tritaeniorhynchus (Diptera: Culicidae) cells persistently infected with Culex Flavivirus (Flaviviridae). J Med Entomol. 2015;52:222–9.
Kenney JL, Solberg OD, Langevin SA, Brault AC. Characterization of a novel insect-specific flavivirus from Brazil: potential for inhibition of infection of arthropod cells with medically important flaviviruses. J Gen Virol. 2014;95:2796–808.
Goenaga S, Kenney JL, Duggal NK, Delorey M, Ebel GD, Zhang B, et al. Potential for co-infection of a mosquito-specific Flavivirus, Nhumirim virus, to block West Nile virus transmission in mosquitoes. Viruses. 2015;7:5801–12.
Gabrieli P, Smidler A, Catteruccia F. Engineering the control of mosquito-borne infectious diseases. Genome Biol. 2014;15:535.
Wilke ABB, Marrelli MT. Paratransgenesis: a promising new strategy for mosquito vector control. Parasit Vectors. 2015;8:342.
Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, Jacobs-Lorena M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc Natl Acad Sci U S A. 2012;109:12734–9.
Mancini MV, Spaccapelo R, Damiani C, Accoti A, Tallarita M, Petraglia E, et al. Paratransgenesis to control malaria vectors: a semi-field pilot study. Parasit Vectors. 2016;9:140.
Bongio NJ, Lampe DJ. Inhibition of Plasmodium berghei development in mosquitoes by effector proteins secreted from Asaia sp. Bacteria Using a Novel Native Secretion Signal. PloS One. 2015;10:e0143541.
Touré YT, Oduola A, Sommerfeld J, Morel C. Biosafety and risk assessment in the use of genetically modified mosquitoes for disease control. In: Takken W, Scott TW, editors. Ecological Aspects for Application of Genetically Modified Mosquitoes. Wageningen: Wageningen UR Frontis Series. 2002;2:217–22.
Christodoulou M. Biological vector control of mosquito-borne diseases. Lancet Infect Dis. 2011;11:84–5.
Kean J, Rainey SM, McFarlane M, Donald CL, Schnettler E, Kohl A, et al. Fighting arbovirus transmission: natural and engineered control of vector competence in Aedes mosquitoes. Insects. 2015;6:236–78.
Carrington LB, Tran BCN, Le NTH, Luong TTH, Nguyen TT, Nguyen PT, et al. Field- and clinically derived estimates of Wolbachia-mediated blocking of dengue virus transmission potential in Aedes aegypti mosquitoes. Proc Natl Acad Sci U S A. 2018;115:361–6.
Baldini F, Segata N, Pompon J, Marcenac P, Shaw WR, Dabiré RK, et al. Evidence of natural Wolbachia infections in field populations of Anopheles gambiae. Nat Commun. 2014;5:3985.
Shaw WR, Marcenac P, Childs LM, Buckee CO, Baldini F, Sawadogo SP, et al. Wolbachia infections in natural Anopheles populations affect egg laying and negatively correlate with Plasmodium development. Nat Commun. 2016;7:11772.
Zhang D, Lees RS, Xi Z, Bourtzis K, Gilles JRL. Combining the sterile insect technique with the incompatible insect technique: III-robust mating competitiveness of irradiated triple Wolbachia-infected Aedes albopictus males under semi-field conditions. PLoS One. 2016;11:e0151864.
Zhang D, Zheng X, Xi Z, Bourtzis K, Gilles JRL. Combining the sterile insect technique with the incompatible insect technique: I-impact of wolbachia infection on the fitness of triple- and double-infected strains of Aedes albopictus. PLoS One. 2015;10:e0121126.
Dittmer J, van Opstal EJ, Shropshire JD, Bordenstein SR, Hurst GDD, Brucker RM. Disentangling a Holobiont - recent advances and perspectives in Nasonia wasps. Front Microbiol. 2016;7:1478.
Brucker RM, Bordenstein SR. The hologenomic basis of speciation: gut bacteria cause hybrid lethality in the genus Nasonia. Science. 2013;341:667–9.
Kikuchi Y, Hayatsu M, Hosokawa T, Nagayama A, Tago K, Fukatsu T. Symbiont-mediated insecticide resistance. Proc Natl Acad Sci U S A. 2012;109:8618–22.
Dunbar HE, Wilson ACC, Ferguson NR, Moran NA. Aphid thermal tolerance is governed by a point mutation in bacterial symbionts. PLoS Biol. 2007;5:e96.
Bourtzis K, Dobson SL, Xi Z, Rasgon JL, Calvitti M, Moreira LA, et al. Harnessing mosquito-Wolbachia symbiosis for vector and disease control. Acta Trop. 2014;132(Suppl):S150–63.
Chavshin AR, Oshaghi MA, Vatandoost H, Pourmand MR, Raeisi A, Terenius O. Isolation and identification of culturable bacteria from wild Anopheles culicifacies, a first step in a paratransgenesis approach. Parasit Vectors. 2014;7:419.
Sharma P, Sharma S, Maurya RK, De TD, Thomas T, Lata S, et al. Salivary glands harbor more diverse microbial communities than gut in Anopheles culicifacies. Parasit Vectors. 2014;7:235.
Thongsripong P, Chandler JA, Green AB, Kittayapong P, Wilcox BA, Kapan DD, et al. Mosquito vector-associated microbiota: metabarcoding bacteria and eukaryotic symbionts across habitat types in Thailand endemic for dengue and other arthropod-borne diseases. Ecol Evol. 2018;8:1352–68.
Mwadondo EM, Ghilamicael A, Alakonya AE, Kasili RW. Midgut bacterial diversity analysis of laboratory reared and wild Anopheles gambiae and Culex quinquefasciatus mosquitoes in Kenya. Afr J Microbiol Res. 2017;11:1171–83.
Acknowledgements
We thank Jean-Marie Hosatte for pictures of Aedes albopictus.
Funding
Morgane Guégan and Colin Démichel are supported by PhD fellowships from the French Ministère de l’Education Nationale, de la Recherche et des Nouvelles Technologies. Karima Zouache is funded by grant ANR-16-CE35-0004-02. This review was funded in the framework of the ERA-NET BiodivERsA program with the national funders ANR-13-EBID-007-01, FWF I-1437, and DFG KL 2087/6–1.
Availability of data and materials
Not applicable.
Author information
Authors and Affiliations
Contributions
MG, KZ and CVM performed the systematic review and wrote the manuscript. GM, CD and VTV assisted in reviewing literature and writing the manuscript. PM and PP provided critical and intellectual input to the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
List of bacteria, fungi and viruses found in different larva and adult mosquito species [15, 17, 18, 20,21,22, 24, 25, 27, 29, 32, 35,36,37,38, 52, 54, 61, 68, 69, 81, 103, 154,155,156,157]. List of articles mentioned are published since 2013. Previous publications on this research topic are available in our previous review [16]. (XLSX 56 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Guégan, M., Zouache, K., Démichel, C. et al. The mosquito holobiont: fresh insight into mosquito-microbiota interactions. Microbiome 6, 49 (2018). https://doi.org/10.1186/s40168-018-0435-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40168-018-0435-2