
Abstract
Ruminants play a critical role in our food system by converting plant biomass that humans cannot or choose not to consume into edible high-quality food. However, ruminant excreta is a significant source of nitrous oxide (N2O), a potent greenhouse gas with a long-term global warming potential 298 times that of carbon dioxide. Natural phytochemicals or forages containing phytochemicals have shown the potential to improve the efficiency of nitrogen (N) utilization and decrease N2O emissions from the excreta of ruminants. Dietary inclusion of tannins can shift more of the excreted N to the feces, alter the urinary N composition and consequently reduce N2O emissions from excreta. Essential oils or saponins could inhibit rumen ammonia production and decrease urinary N excretion. In grazed pastures, large amounts of glucosinolates or aucubin can be introduced into pasture soils when animals consume plants rich in these compounds and then excrete them or their metabolites in the urine or feces. If inhibitory compounds are excreted in the urine, they would be directly applied to the urine patch to reduce nitrification and subsequent N2O emissions. The phytochemicals' role in sustainable ruminant production is undeniable, but much uncertainty remains. Inconsistency, transient effects, and adverse effects limit the effectiveness of these phytochemicals for reducing N losses. In this review, we will identify some current phytochemicals found in feed that have the potential to manipulate ruminant N excretion or mitigate N2O production and deliberate the challenges and opportunities associated with using phytochemicals or forages rich in phytochemicals as dietary strategies for reducing N excretion and excreta-derived N2O emissions.
Similar content being viewed by others
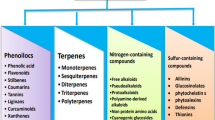

Introduction
Ruminant animals are extremely important not only for producing the highest quantity of milk and meat as essential parts of human diets, but also for their ability to feed on fibrous feeds that cannot be used as human food [1]. However, 75%–90% of consumed nitrogen (N) is excreted as urine and feces [2]. As the excreted N exceeds the plant demand, it can result in considerable N losses via nitrate (NO3−) leaching, ammonia (NH3) volatilization, and nitrous oxide (N2O) production [2]. N2O, a potent greenhouse gas (GHG), is estimated to be 298 times more powerful than carbon dioxide (CO2) in warming power over 20 years [3]. By oxidizing into nitrogen oxides in the stratosphere, N2O plays a vital role in depleting stratospheric ozone [4]. Anthropogenic N2O emissions (2.7 ± 1.6 GtCO2-eq) were 133% higher in 2019 than in 1990 [3]. Approximately 81% of anthropogenic N2O emissions are attributed to the agricultural sector [3], and N2O emissions from ruminant excreta account for 46% of agricultural N2O emissions [5].
Ruminant production is projected to continue growing in the next few decades, driven by fast growth of human population, rising incomes, and dietary preferences towards ‘Western’ diets [6]. Given the growing concerns over the environmental impacts of ruminant farming, there is an impetus to decrease the emissions of ruminant-derived N2O substantially. Over the years, several manure N and N2O mitigation strategies, including dietary or manure management approaches at the herd level along with more targeted approaches, such as reducing dietary protein intake [7], supplementing sodium chloride [8], changing rumen undegradable protein sources [9], alternative forage [10, 11], and nitrification inhibitors [12, 13] have been widely investigated. Contemporary consumer demands orient towards the use of ‘natural products’ to alter ruminant N metabolism and excreta-derived N2O emissions.
On the one hand, regulating the N metabolism of ruminants using nutritional strategies could decrease N excretion and, consequently, reduce N2O emissions. Previous studies indicated that the addition of natural phytochemicals (e.g., tannins, saponins, and essential oils) could reduce ruminal NH3 concentration and alter excreted N partitioning, which helps to lower urinary N excretion and N2O emissions to the atmosphere [14,15,16]. On the other hand, phytochemicals may lessen N2O loss when they are deposited in urine, either as diuretics to lower pasture N loading rates or as natural nitrification inhibitors because of their antibacterial activities [17, 18]. Inhibitory substances found in forage can be applied directly to urine patches after being consumed by ruminant livestock. For example, Plantago aucubin and Brassica isothiocyanates have been shown to inhibit a crucial step in N2O generation from urine patches in ruminants fed these forages [19,20,21].
However, the role and efficacy of phytochemicals in reducing N2O emissions from the excreta of ruminant livestock remain controversial. The reasons for the controversy are multifaceted, mainly including the variable effect of reducing N2O emissions, different assessment methods of N2O emissions, the source of phytochemicals, the number of phytochemicals ingested, the cost of additional feeding, and possible side effects in terms of performance and health. In this review, the potential, mechanisms, and unsolved problems of reducing N2O emissions from ruminant livestock through feeding phytochemicals are also discussed. Our purpose is to provide deeper insights into use of phytochemicals to manipulate N excretion and mitigating N2O emission from ruminants.
N2O production from ruminant excreta
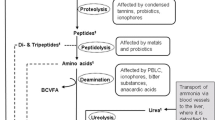
N2O is predominantly generated through two major biological pathways, i.e., nitrification and denitrification [22], but may also be produced by other processes such as nitrifier-denitrification or codenitrification [23]. As shown in Fig. 1, following excreta deposition, the major fraction of organic N or urea N in manure is mineralized or hydrolyzed into ammonium (NH4+) and then converted to NO3− via the activity of nitrifiers under partial aeration. The NO3− produced can then be transformed into dinitrogen (N2) by denitrifying bacteria under anoxic conditions. During nitrification of NH4+ and denitrification of NO3−, N2O gas may escape into the atmosphere as a by-product. The proportion of N released as N2O from ruminant excreta is significantly influenced by feeding regimes, environmental circumstances, farm systems, and manure management practices [24]. We can divide these influencing factors into two facets (i.e., animals and environmental factors) according to the N cycle between ruminants and the environment (Fig. 1).

A brief overview of the N2O production pathways in ruminant excreta applied to soil and the potential influence factors. N Nitrogen
In the latest review, Mancia et al. [25] discussed the factors of disaggregation of N2O emission factors (EF), such as excreta type, animal diet, seasonal variations, and spatial variability. Considering the association between ruminant excreta composition and dietary mitigation strategies, this review focuses on disaggregation by excreta type. In the 2006 Intergovernmental Panel on Climate Change (IPCC) guidelines, the default values of EF were 2% and 1% for cattle and sheep, respectively, with no distinction between urine and dung [26]. The more recent IPCC values of N2O EF have been updated, and the urine-based and dung-based N2O-EF for cattle was 0.77% and 0.13%, respectively, in a wet climate [27]. Most studies reported that the N2O-EF of urine patches were greater than the dung-based N2O-EF [28,29,30,31,32], except the results depicted by Wachendorf et al. [33] and Ma et al. [34].
Compared with a longer time for dung N mineralization, urine could rapidly supply available NH4+ for nitrification and denitrification by hydrolyzing urea, contributing to higher N2O losses [35]. Additionally, the high dry matter (DM) content of the dung also reduced the potential for dung N to infiltrate into the soil, restricting interaction with the soil microbial community [36]. The difference implies that the ratio of dung N to urine N excreted can also influence the amounts of N2O emissions. The benefit of disaggregating emissions into dung and urine is that the effect of diet manipulation on N partitioning and thus on N2O emissions can be accounted for. Therefore, the disaggregation by excreta type offered a better opportunity to regulate N2O emissions through dietary phytochemicals.
Potentials of natural phytochemicals in reducing N excretion and N2O emissions
Diet has a profound effect on the chemical composition and partitioning of excreted N, and may therefore indirectly affect N2O emission from excreta patches [37]. Several phytochemicals (e.g., tannins, essential oils, saponins, and glucosinolates) present in forages and plant extracts have been identified as possible methane (CH4) inhibitors in the rumen [38,39,40,41]. Nonetheless, there are numerous opportunities to simultaneously reduce N waste and CH4 production in ruminants [15]. In this section, recent advances using dietary phytochemicals as N excretion and N2O emission mitigation strategies are described here.
Tannins
Condensed tannins
Conventionally, tannins are usually classified into two groups: condensed tannins (CT) and hydrolyzable tannins (HT). CT or proanthocyanidins are formed via C4–C8 and C4–C6 interflavonoid connections between chatequins, leucoanthocyanidins, and their derivatives; CT is not rapidly degraded in the gastrointestinal tract [42]. CT can interact with plant proteins via hydrogen bonding in the near neutral pH range to form insoluble tannin-protein complexes, which are subsequently dissociated in the abomasum to release protein. CT may hinder the growth of proteolytic microbes, thereby inhibiting proteolysis [43]. Therefore, the formation of tannin-protein complexes and the suppression of microbial proteolytic activity may reduce rumen degradability and increase the flow of protein into the intestine [44] (Fig. 2).

The model of action of dietary tannins in reducing N excretion and N2O emissions from ruminat excreta. N Nitrogen
Numerous studies have reported that dietary supplementation with CT reduced urinary N output and increased fecal N excretion in dairy cows [45], beef cattle [46], sheep [47], lambs [48], and goats [49] (Table 1). The shift in N excretion from urea in the urine to a more stable form of N in feces can increase soil organic N concentration and reduce N2O emissions [50]. A field study demonstrated that feeding beef steers with a tannin‑rich legume (sericea lespedeza hay) effectively reduced the emission of N2O, CH4, and CO2 from the soil for 32 d after the application of manure [51]. However, Hao et al. [52] reported that supplementing CT in the diet of beef cattle did not affect composted manure N2O emissions. The converse effects were obtained by de Souza et al. [53], who observed that Acacia mearnsii tannin extract elevated N2O emissions from excreta patches in the field; however, this unexpected result cannot be well explained. These findings suggested that the effects of CT on ruminant N utilization and N2O emissions can be highly diverse depending on origin, concentration, molecular structure, and tannin dosage.
However, most studies in the past decades investigating the effects of dietary CT on N excretion were conducted with penned ruminant livestock under intensive feeding systems. Only a few articles have been published regarding the effects of fresh forages rich in tannins on excreta-derived N gaseous losses under year-round grazing. Acacia cyanophylla foliage containing 3% CT fed to grazing ewes up to 200 g/d reduced urinary N excretion and increased NUE [55]. Lagrange et al. [63] found that a combination of tanniferous legumes (birdsfoot trefoil and sainfoin) led to reductions in urine N and urinary urea-N concentration that were larger than the decrease observed for the single tanniferous species alone. However, it remains controversial whether tannins can maintain their biological activity during the haymaking process [67]. Stewart et al. [15] reported that feeding CT-containing hays [birdsfoot trefoil (0.6% CT) or sainfoin (2.5% CT)] or HT-containing hay [small burnet (4.5% HT)] to Angus heifers or beef cows also reduced urinary urea N excretion and shifted the partitioning of N from urine to feces, compared to feeding traditional legume and grass hays. The results of Stewart et al. [15] suggested that tannins retain their biological capabilities (i.e., influencing N metabolism) regardless the modifications during the haymaking stage. Furthermore, the potential of adding chestnut and mimosa tannin to grass at ensiling to improve N utilization in sheep was also investigated by Deaville et al. [47], who found that compared to the control silage, both tannins decreased urinary N excretion and increased fecal N output.
By mixing CT into the feed of penned ruminant livestock, enough CT intake can be achieved; however, when cattle and sheep are grazing, it can be difficult to achieve an adequate and consistent consumption of a feed supplement containing CT. Kronberg and Liebig [56] showed that supplementing quebracho tannins to the freshwater of grazing sheep lowered urine urea deposition onto grasslands, and evaluating the feasibility of adding CT to drinking water to minimize N2O emissions from urine patches in pastures is warranted. Tannins have been studied extensively in reducing N or N2O emissions in confined livestock raised in temperate climates. However, the use of tannins can be extended to other production systems by considering their basic biology.
In addition to reducing of urine N2O emissions through decreasing urinary urea excretion, the CT presented in feces may provide a feasible strategy to reduce N2O emissions by applying manure (a combination of dung and urine) to agricultural soils. Over 50% of CT remains undigested in the ruminant gastrointestinal tract [68, 69]. In this light, it should be no surprise that ruminants fed a diet high in CT will excrete CT-rich feces. Tannins from birdsfoot trefoil were shown to be present and potentially active in the feces of dairy cows, as shown by Misselbrook et al. [70]. When dairy calves were fed dietary CT, NH3 emissions from slurries on the barn floor [70] and slurries applied to soil [70, 71] were reduced. Powell et al. [71] observed that urease activity in feces and NH3 emissions from manure were both reduced when dairy cattle were fed chestnut tannin extract or simulated barn floors were applied with tannins. Recent studies confirmed that N2O emissions were reduced from tannin-enriched manure [72, 73].
Additionally, because mineralization of the complex is inhibited, the tannins-protein complexes in feces are more resistant to breakdown in the soil and decompose more slowly than feces without CT [74, 75]. Fagundes et al. [76] reported that feeding Acacia mimosa tannin extract to cattle increased fecal N output, delayed organic matter breakdown, and changed soil microbial dynamics following feces application. However, these researchers did not quantify N2O emissions from the feces of ruminant fed-CT. Larger-scale studies are required to determine the effectiveness of dietary tannin extracts in abating N2O loss from ruminant barn floors and land-applied excreta.
Hydrolyzable tannins
Compared with CT, HT has a weaker affinity for proteins and thus is more easily absorbed by the gastrointestinal tract, increasing potential toxicity to the animal [42, 77]. For this reason, previous studies on the utilization of tannins in ruminant livestock focused on CT instead of HT. Nevertheless, an in vitro investigation revealed no difference between tannin sources in preventing protein degradation [78]. Therefore, HT can also bind to bacteria, modifying their activity, and to proteins, reducing their breakdown in the rumen and consequently altering N output. Supplementing HT extracted from chestnut at 1%–3% DM in sheep [79] or coupled with CT extract (derived from quebracho) at 1.5% DM in steers [80] decreased the ruminal NH3 concentration (Table 2). Chestnut tannins, as a representative HT, have been shown the potential to minimize the environmental impact of ruminants via the N shift from urine to feces [47, 81].
Tannic acid is another typical HT. Yang et al. [83] reported that the supplementation of tannic acid to the diet of beef cattle reduced the ratio of urine N to fecal N and modified the concentrations of nitrogenous compounds in the urine. Gallic acid derives from the hydrolysis of specific HT [87]. Feeding gallic acid to beef cattle altered the pattern of N excretion by increasing the ratio of fecal N to urinary N and decreasing the ratio of urinary urea N to urinary N [82]. Subsequently, laboratory incubation trials demonstrated that adding gallic acid or tannic acid to the diet of steers reduced N2O fluxes after applying urine to the soil [7, 10].
In contrast to CT, it is believed that HT can be degradable in the rumen [88]. Dietary supplementation with gallic acid decreased urine N2O emissions from beef cattle, while urinary N excretion or urea excretion was not influenced [10]. Bao et al. [10] attributed it mainly to the excretion of gallic acid metabolites, such as pyrogallol and resorcinol, which might inhibit the processes of N2O production. However, no direct evidence for this proposed mechanism is currently available. Additionally, a greater amount of urinary hippuric acid excretion was observed in beef cattle fed tannic acid [7, 83]. Hippuric acid excretion correlates with polyphenols consumption because polyphenols are the precursor components to hippuric acid formed during rumen fermentation [89]. The antibacterial compound hippuric acid has been considered for its ability to reduce N2O emissions. Researchers have found that enhancing the percentage of hippuric acid in the urine caused a decrease in N2O fluxes from bovine urine of 50%–54% [90, 91], most likely due to the inhibition of denitrification or a reduction in the ratio of N2O to N2 [90]. Bertram et al. [92] reported that hippuric acid also partially inhibited soil nitrification. These observations suggest that an elevation in urine excretion of hippuric acid in response to the consumption of biodegradable polyphenols (e.g., HT) may represent a possible N2O mitigation strategy. However, conflicting findings have been found in the limited field tests done so far, with no inhibitory effects of hippuric acid on urine N2O fluxes [93, 94]. Further research is needed to identify whether supplementing HT to increase the formation of HT metabolites in urine will decrease subsequent N2O emissions.
In a word, although tannins have shown good potential in reducing urinary N and N2O from ruminant excreta, it does not mean that they can be promoted in livestock production. The nutritional and environmentally sustainable potential of tannins will only be realized when the composition, structure, and biological function of tannins in plant extracts or forages are better defined. Chemical assays should be complemented by measurements of binding capacity to plant proteins and effects on enzyme activity or in vitro digestion, as the relationship between chemical structure and stringency is not well defined. Analytical and experimental efforts will reveal the most effective tannins for expressing temperate legumes by genetic engineering or conventional selection as part of plant breeding projects or for mitigating N2O from ruminant excreta utilizing existing forages.
Glycosides
Glucosinolates
Glucosinolates (GLS) are a large group of plant secondary metabolites with nutritional effects and biologically active compounds. The GLS molecule comprises a β-thioglucose unit, a sulfonated oxime unit, and a side chain derived from an amino acid that varies (Fig. 3A) [95]. There are at least 120 distinct structures of GLS, which are found in 16 families of dicotyledonous plants [95]. Brassica plants are annuals traditionally used to fill feed deficits in temperate ruminant grazing systems [96]. There are five primary degradation products of GLS, with isothiocyanate being the most important, followed by thiocyanate, nitrile, epithionitrile, and oxazolidine-2-thione [97] (Fig. 3A).

The chemical structure of glucosinolate and its metaboites (A), aucubin (B), aucubigenin (C), and saponin (D)
The potential of GLS hydrolysis products to act as biological nitrification inhibitors has been investigated. Studies reported that the application of Brassicaceae tissues and subsequent the generation of various GLS hydrolysis products (isothiocyanate, thiocyanate, and nitriles) could weaken the activity of nitrifying bacteria [98] and inhibit nitrification [99]. The GLS hydrolysis products have been shown to inhibit the nitrification processes in soil incubated with brassica seed meals containing GLS [100].
GLS and their decomposition metabolites have been detected in the urine of animals fed brassicas [101]. Urine from sheep-fed forage rape (Brassica napus L.) had a lower N2O-EF than urine from sheep-fed perennial ryegrass (Lolium perenne L.) when applied to a freely draining pasture soil in early spring (0.11% vs. 0.27%, respectively) [19] (Table 3). According to Hoogendoorn et al. [102], the urine from sheep-fed ryegrass (Lolium perenne L./Trifolium repens) had a higher N2O EF than sheep-fed fodder rape (Brassica napus L.). This decline may be explained by the fact that the brassica-derived chemicals GLS hydrolysis products were transferred from urine to the soil [103, 104], inhibiting nitrification in the urine patch [105]. The promising findings of these experiments indicate that isothiocyanate and other urinary secondary metabolites may function as nitrification inhibitors. In a laboratory study, Balvert et al. [105] demonstrated that several GLS hydrolysis products (isothiocyanate and nitrile) inhibited the nitrification process and lowered N2O fluxes from urinary urea applied to soils (Fig. 4). In a field experiment, however, the application of GLS hydrolysis products to artificial urine patches did not result in any significant differences in N2O emissions [105].

The possible mechanisms for glucosinolates, aucubin, and saponin as ruminant manure N and N2O mitigation strategies. N Nitrogen, BNI Biological nitrification inhibition
Furthermore, several experiments have also examined the response of N2O production to the metabolite thiocyanate. Snyder et al. [118] observed that 2-propenyl isothiocyanate and thiocyanate released from Brassica juncea and Sinapis alba seed meals hindered bacterial metabolism and that thiocyanate release was the cause of nitrification inhibition in the Sinapis alba treatment. Thiocyanate was also detected in the rumen fluid and urine of cattle fed rapeseed meals [106, 119, 120], whereas isothiocyanate and oxazolidine-2-thione remained undetected in the rumen fluids. Thus, thiocyanate may be the primary product of GLS hydrolysis in the rumen, and that the ruminal GLS metabolic route could be distinct from that driven by myrosinase [119]. Contrary to expectations, in a laboratory study, Gao et al. [106] found that the urine N2O emissions and N2O-EF were increased in steers received the diet containing high GLS-rapeseed cake. A significant correlation was found between urinary thiocyanate content and urine N2O fluxes [106]. Additional laboratory incubation confirmed that thiocyanate enhanced the denitrification of soil nitrifiers, which may have led to an increase in N2O emissions [121]. These controversial results suggest that GLS hydrolysis products differ in their effects on N transformations. Therefore, more GLS hydrolysis products excreted in ruminant urine should be assessed for their individual or combination effect on N2O production from urine.
Unfortunately, except for the publication by Gao et al. [106], no study has investigated the potential for the feeds containing GLS and the subsequent effect of the voided urine to reduce soil nitrification and N2O emissions. Before the intake of forage rich in GLS or their hydrolysis compounds can be perceived as a strategy for decreasing N2O emissions, the composition and concentration of GLS hydrolysis products in urine must be determined. More research into the N2O production mechanisms in soils treated with urine from ruminants-fed brassica plants under various environmental situations is required. It should be noted that the action of GLS depends on their activity in fertilizing the soil with excreta (mainly urine). However, its activity is often affected by the environment and is full of uncertainties. At the same time, GLS is one of the common antinutritional factors in ruminant feed. Therefore, targeting GLS as an inhibitor to reduce N2O emissions may be difficult.
Aucubin
Aucubin, an iridoid glycoside found in plantain (Plantago), displays similar inhibitory properties as GLS and their metabolites (Fig. 3B). Aucubin is known to degrade completely into its instability aglycone, aucubigenin (Fig. 3C), within 4 h in the presence of β-glucosidase [122], and β-glucosidase is known to be present in soils [123]. Aucubigenin can be converted into an unsaturated aldehyde that binds permanently to the nucleophilic side chains of nucleic acids and proteins, due to its potent alkylating characteristics [124]. Numerous experiments showed that dairy cows grazing plantain (Plantago lanceolata L.) or mixed pastures containing plantain had a lower N concentration in spot-sampled urine [18, 107,108,109, 125] (Table 3), and these researchers hypothesized that the overall output of urine N might also be lowered. This can offer an opportunity to utilize plantain to minimize N losses in grazing ruminants. The increased fraction of undegradable N is one of the potential causes of the lower urine N content in cows received plantains. The greater undegraded N content allows more N to pass through the rumen to be digested in the small intestine, where more N is partitioned to milk and feces, and less N is excreted into urine. Minnée et al. [109] showed that feeding dairy cows with a diet containing 45% DM of plantain reduced the amount of urinary N while increasing the amount of N partitioned to milk and feces by 14%.
The second possible mechanism for decreasing urinary N concentration and increasing urine volume is aucubin's diuretic action. O'Connell et al. [126] confirmed this effect by observing that penned sheep fed plantain emitted more urine than those fed ryegrass. Additionally, Navarrete et al. [127] found that aucubin reduced NH3 production in vitro and was likely degraded to its active aglycone aucubigenin in the rumen. Therefore, the third possible mechanism for influence of grazing plantain on urinary output could be partly due to aucubin lowering ruminal NH3 losses and reducing urea production in the liver [128]. Aucubin, for instance, is antibacterial due to the aucubin aglycone (aucubigenin), which binds to free amino acids, making them unavailable [122]. The escape from the rumen of these aucubigenin-amino acids complexes could also shift the N partitioning from urine to feces by transferring the digestible N fractions to the intestine. Therefore, aucubin and/or aucubigenin in plantain could potentially change the N partitioning in ruminants (Fig. 4).
Furthermore, soils under plantain showed significantly lower NO3− concentrations [129], mineralization and nitrification rates [129], lower ammonia oxidizer bacteria abundance [130], and lower N2O fluxes [131] than under other plant species. Likely, plantain root-released chemicals (e.g., aucubin) with biological nitrification inhibitory capacity contributed partially to the reduced N2O flux observed in the presence of plantain. Luo et al. [20] observed that comparison of N2O emissions from cattle urine applied to plantain and to perennial ryegrass, plantain had lower emissions in winter but not in other seasons. This result may be due to differences in concentrations of aucubin or other secondary metabolites as they could have been affected by temperature or other environmental variables [132]. In addition, broad-spectrum antibacterial action has been observed with aucubin, and aucubigenin has been shown to inhibit cytochrome P-450, which may be related to its capacity to limit ammonia oxidation by inhibiting the activity of the soil enzyme ammonia monooxygenase [133, 134]. Thus, aucubin and its derivative aucubigenin are potential biological nitrification inhibitors (Fig. 4).
It is uncertain what form or concentration of aucubin is excreated in ruminant urine because Navarrete et al. [127] did not quantify the residence time of aucubin and/or aucubigenin during ruminal fermentation or identify its fate following ruminal metabolism. Additionally, plantain was the only plant species tested (550 total) whose antibacterial activities were detectable in rabbit urine 8–16 h after feeding [135]. Even though a rabbit is not a ruminant, similar results can be obtained with ruminant urine. Judson et al. [136] found higher soil NH4+ contents following the application of urine from dairy cows received 100% plantain compared to the urine of cows fed with ryegrass-white clover. However, the two urine sources had comparable total N contents, suggesting that urine from grazing cows fed plantain can inhibit nitrification. Similarly, Simon et al. [21] observed that the increasing consumption of plantain for grazing cows decreased urinary N loading rates and urine N2O emissions. Thus, another potential route for the aucubin in plantain to enter the soil would be through the urine of ruminants grazing plantain-based pastures.
For a short period of time following soil application, aucubin may act as a nitrification inhibitor; however, its inhibitory actions seem insufficient to produce substantial reductions in total urine patch N2O fluxes [137]. A latest trial revealed that grazing dairy cows on plantain pastures did not lower urine N2O fluxes compared to ryegrass-white clover urine when treated at the same N urine rate [138]. It is hypothesized that aucubin degrades swiftly in soils and that the suppressive action of its decomposition metabolites, notably aucubigenin, persist in soils for no more than a few days [137]. Further studies should determine the metabolic pathways of aucubin in soil, quantify aucubin urination ratios, and investigate the impact of aucubin excretion ratios on the inorganic-N dynamics and N2O emissions of urine patches.
Saponins
Saponins are a large family of amphiphilic glycosides of steroids and triterpenes (Fig. 3D). Saponins is well known for their potential of decreasing rumen CH4 production by decreasing both the number and activity of methanogenic microorganisms [139]. Another important effect of saponins in the rumen appears to be to inhibit the protozoa (defaunation) by affecting cell membrane integrity [140]. Ruminal NH3 concentrations are reduced when protozoal growth is inhibited, presumably due to depressed rumen degradation of feed protein or turnover of bacterial protein [14]. NH3 concentration also will be altered by binding of NH3 to compounds like saponin, as noted by Cheeke [141]. Jouany [142] also assumed that urinary N always decreases with defaunation, due to both the decreased NH3 concentration in the ruminal fluid and the increased capture of urea N for microbial protein synthesis. Hu et al. [143] showed that the addition of 40 g/kg of tea saponin led to the lowest concentration of rumen NH3-N and the maximum microbial protein content in vitro. Overall, it appears that plants or their extracts with high concentrations of saponins may operate as natural rumen manipulators, which can increase the efficiency of microbial protein synthesis and enhance protein flux to the intestine by decreasing microbial protein turnover.
In a review by Wina et al. [144], 14 out of 51 publications indicated that saponins did not affect rumen NH3-N content, whereas 17 indicated an inhibitory effect. Supplementation of Biophytum aqueous extract, up to 26 mg/kg BW of saponin, decreased rumen NH3-N and urinary N output, thereby increasing microbial N supply and retained N as a proportion of N digested in goats [14]. Ramírez-Restrepo et al. [145] reported that adding tea seed saponin reduced blood urea concentration in tropical Brahman cattle. Liu et al. [117] observed that dietary addition with tea saponin decreased rumen NH3-N, fecal N, and urinary N excretion, leading to a significant increase in N retention and NUE in Dorper crossbred ewe. These results indicate that saponins may contribute to mitigating N excretion and N2O emissions from ruminants. However, it was shown that tea saponin did not modify the N balance or N excretion of lactating cows [116] or nonlactating cows [115]. These discrepancies may be due to variations in the experimental diets and saponin dosages.
To test the effect of saponin extracts or saponin-rich forages on N2O emissions from excreta under grazing circumstances, additional animal studies and field experiments are still needed. To achieve sustained beneficial effects of saponins in diets, it is necessary to conduct extensive research on the interactions between saponin chemical structures, dietary nutrition components, and their influence on the rumen microbial ecology. It is essential to identify the most biologically active saponins that inhibit the activity and abundance of protozoa while possibly stimulating beneficial bacteria and fungi. Certain classes of saponins may have toxic effects on the body and must be examined in vivo in long-term studies. If more active saponins can be isolated and identified from plants, or if plant biotechnology techniques can be used to produce target saponin components, the beneficial effects of saponins could be widely exploited in various feeding systems, assuming it can be demonstrated in the future that saponins in ruminants are effective at reducing animal N excretion and excreta-derived N2O emissions.
Essential oils
Aromatic plants can produce essential oils, complex combinations of volatile organic substances. Essential oil can contain up to 60 chemical substances, such as alcohols, aldehydes, hydrocarbons, ketones, esters, and ethers [145]. Essential oils have been demonstrated to possess the ability to affect ruminal protein degradation and amino acid absorption in the small intestines of ruminant livestock. Numerous studies reported that addition with essential oil decreased rumen NH3-N concentration in vitro (e.g., Golbotteh et al. [146]; Patra and Yu [147]; Pawar et al. [148]) or in vivo (e.g., Lin et al. [149]; Toseti et al. [150]; Wu et al. [16]). Carrazco et al. [151] also found that feeding essential oils reduced enteric emissions of N2O and NH3 in mid-lactation dairy cattle. Essential oil can reduce ammonia levels, likely due to direct inhibition of proteolytic and ammonia-producing rumen bacteria [147]. Their antibacterial characteristics are explained by various mechanisms, including chemical structures and physical properties [152]. Essential oils are hydrophobic, partitioning through lipid cell membranes, disrupting their integrity and stability, and resulting in leakage of cell contents [153]. The hydroxyl group and their relative position in the phenolic structures (in the case of thymol and eugenol) were believed to be important attributes that influence the antibacterial properties of essential oil [152].
Reducing ruminal NH3 loss and moving more microbial protein to the small intestine can increase tissue N retention, reducing the urinary N excretion and the potential of N2O emission from manure application. Wanapat et al. [154] observed that feeding garlic powder containing essential oil at 80 g/d with urea-treated rice straw decreased urinary N excretion and improved N retention of steers (Table 4). Ribeiro et al. [155] showed that supplementing thyme essential oil enhanced N retention and reduced urinary N excretion compared to monensin. Specifically, the latest meta-analysis of the effectiveness of essential oils revealed that N retention was greater in beef cattle that received essential oil [156]. However, these results should be interpreted with caution because of the low number of studies that reported these response variables [156].
Other trials using lactating dairy cows have also shown that single or combinations of essential oils containing thymol, eugenol, and/or carvacrol have no effect on N utilization [145, 158, 160]. Muñoz-Cuautle et al. [161] found that including oregano essential oil in the diet did not alter urinary N, fecal N, or N retention in meat lambs. Conversely, Tekippe et al. [158] reported that supplementing 525 mg/d essential oils products containing eugenol and cinnamaldehyde enhanced urinary N excretion in dairy cows. Several parameters, such as trial duration, essential oil chemical composition, and dosages, may account for differences in results among in vivo trials. The contradictory results may also be attributable to variations in the kinds and amounts of dietary protein consumed. Some findings indicate that essential oil can inhibit the colonization and/or subsequent degradation of readily degradable substrates, such as starch and protein, thus impacting the metabolism of amylolytic and proteolytic bacteria [162]. All fermentation processes associated with dietary protein degradation and ruminal NH3 production require further investigation. Animal production indices should be quantitatively and qualitatively correlated with the effects of increasing dietary protein escape from the rumen. Overall, the most promising essential oils and their effective concentrations and combinations can be evaluated further in vivo experiments to determine the essential oils (dose and combination) that can be applied on farms. Before their use on farms, the positive effects of commercial essential oils on animal performance and the environment must be established due to their high cost.
Challenges of phytochemicals as manure N and N2O mitigation strategies
The consistency and comparability of study results
Phytochemicals are highly variable depending on many factors, including plant species, growth environment of the plants (e.g., soil composition, temperature, and moisture stress), stage of plant growth, parts of the plants utilized to extract phytochemicals, and phytochemicals extraction or analysis method [163]. However, currently, there is no standard product composition, structure and purity of any commercial plant extracts for use in livestock production. As a result, there are often considerable variations between research results, making it difficult to ascertain the necessary types and dosages. For example, Cobellis et al. [163] summarized the effects of various single essential oils and essential oil blends on ruminal N metabolism in vivo and in vitro experiments and discovered that in vitro and in vivo results are always inconsistent. As shown in Tables 1 and 2, numerous studies showed that feeding tannins increased fecal N output, which is a result of tannins limiting the absorption of N, and suggests that part of the protein-tannins complexes failing to dissolve in the abomasum, resulting in a loss of dietary protein [164]. However, some studies observed that tannins altered only urinary N. Depending on the chemical structures of tannins and proteins, tannin-protein interactions do not always function optimally and account for these variations among studies.
Numerous in vitro studies have documented the possible ruminal NH3 mitigation effects of phytochemicals in vitro. However, it is generally accepted that in vitro systems, while excellent for screening for bulk inhibitors, are not very representative of responses in N excretion and excreta-derived N2O for live animals. Therefore, it is strongly suggested that in vitro results be confirmed in in vivo experiments. Moreover, future studies should detail the source of phytochemicals, extraction method, chemical composition, purity, and dosage. In vitro experiments and studies of the minimal inhibitory concentration of rumen or soil microbes using pure active compounds can shed light on their action method, reveal their major active components, and assist in establishing an appropriate dosage. A better understanding of structure–activity relationships would be needed to acquire consistent results from phytochemicals on mitigating ruminant manure N and N2O emissions.
The balance between its efficacy and side effects
Despite extensive research conducted in recent years, using phytochemicals in ruminant livestock remains challenging and very limited for side effects. Tannins in the diet can bring significant benefits to ruminant livestock; nevertheless, high dietary contents or CT with the ‘wrong’ compositional features would reduce digestion and utilization of dietary protein and absorption of crucial amino acids by the ruminant [88]. For example, Ahnert et al. [57] found that the ruminal infusion of a moderate level of quebracho tannin extract may significantly shift N excretion from urine towards feces, while high quebracho tannin extract dosages has detrimental effects on crude protein and fiber digestibility. The binding capabilities of tannins may potentially reduce fiber digestibility by inhibiting cellulolytic enzyme or binding to dietary carbohydrates, reducing ruminal turnover rate, and consequently minimize feed intake and animal production performance [59, 165]. Guyader et al. [116] also reported that milk production, DM intake, and feed efficiency of dairy cows were reduced with tea saponin (0.52% DM). Major deleterious effects of GLS ingestion in animals are reduced palatability, decreased growth and production [166]. In addition, nitriles are known to influence the activities of the liver and kidneys. Thiocyanates inhibit the availability of iodine, whereas oxazolidine-2-thione can induce the morphological and physiological alterations in the thyroid [166].
Overall, the challenge is to determine which phytochemical features can reduce N2O production from excreta by improving dietary N utilization and/or exerting the biological nitrification inhibitory activities in the urine, without harming animal health, performance, or farmers' profitability. Throughout the production cycle and across several production cycles (for example, dairy cows), the impact of manure N and N2O mitigation strategies on animal health, welfare, and reproduction must be examined through long-term research. Long-term experiments are also needed to study the mechanism of adaptation of gastrointestinal microbes and animals to phytochemical. A better understanding of how phytochemical mitigation strategies impact ruminant product composition, shelf life, sensory traits, and consumer perceptions of livestock products is also essential.
Systematic research methodology
Although saponins and essential oil have the potential to manipulate ruminant N metabolism, N2O fluxes from ruminant excreta using field plots with the static chamber method have not been investigated previously. In grazing systems, biologically inhibition of nitrification through dietary manipulation with GLS or accubin has not been extensively investigated. The following step is to determine the effect of dietary phytochemicals more closely on N2O emissions and soil N cycles in larger-scale, longer-term experiments that simulate more closely manure management of commercial ruminant production systems.
Accurately estimating emissions from farmers using manure N and N2O mitigation options requires an integrated systems approach. Life cycle assessments (LCA) should evaluate the upstream and downstream impacts of mitigation strategies. Meta-analyses are critical to determine the effectiveness of phytochemicals as mitigation protocol. Additionally, phytochemicals have the potential to decrease rumen CH4 emissions. However, few studies have examined phytochemicals' use to reduce main GHG (i.e., CO2, CH4, and N2O) on either the herd level or in individual animals. The goal of reducing GHG is undermined if a strategy reduces N2O but increases another GHG. If LCA is performed, only then will this be captured. Additionally, dietary manipulations with phytochemicals targeting excreted N or N2O reduction are mostly studied in isolation.
Conclusions and perspectives
Ruminant production systems are significant contributors to global N loss and N2O emissions. As demand for high-quality meat and milk products rises, N2O emissions and global temperature will continue to increase. Phytochemicals, because of their anti-microbial activity and easy availability, may be promising agents to enhance NUE and reduce the environmental impact of ruminant N2O emissions. The proposed mechanisms of N2O reduction using natural phytochemicals include inhibiting rumen NH3 production, increasing N partitioning into feces relative to urine, the diuretic effect phytochemical of leading to more frequent urination, and biologically nitrification inhibitor function of plant secondary metabolites from root exudation and/or animal’s urine. Present results indicate that the dietary inclusion of tannins could considerably reduce N excretion and excreta-derived N2O emissions from cattle excreta, whereas the possible negative effects of tannins on ruminant feed intake and nutrient digestibility are of concern. Compared with tannins, none of the existing studies has provided conclusive evidence of the effectiveness and mechanisms of plant glycosides or essential oils in reducing N excretion and N2O emissions, and these compounds should also be evaluated in long-term in vivo trials for their effect on N metabolism and N2O production. Thus, further studies are required to determine their bioactive compositions, effective doses, mode of action, effect on animal performance and health, and cost–benefit ratio before phytochemicals can be applied as additives on farms to minimize N excretion and N2O emissions from ruminant excreta.
Another major issue is affordability; ruminant farmers need greater information on the cost of natural phytochemicals or forages rich in phytochemicals and their impacts on animal productivity, particularly for concentrated animal feeding operations. Regulatory approval requirements for some promising feed ingredients may slow their adoption, and a lack of consumer acceptance of some of them may preclude their use for N2O mitigation. Incentives and low-cost approaches may be needed to encourage adoption because, in most cases, decreased N excretion and N2O production have not increased ruminant performance.
Additionally, it can be challenging to assess the system-wide effects of N2O reduction practices, even though they may be beneficial at specific stages of the production cycle. Therefore, it is critical to analyze mitigation strategies and procedures based on natural phytochemicals or forages rich in phytochemicals in the context of the whole system and LCA to ensure efficiency gains across all levels. In a word, phytochemicals may have a place in sustainable ruminant production scenarios only if more convincing results of their efficacy and effectiveness in mitigating N excretion and GHG emissions are dependably identified. The old saying “do not put all your eggs in one basket” still applies to phytochemical research.
Availability of data and materials
Not applicable.
Abbreviations
- CH4 :
-
Methane
- CT:
-
Condensed tannins
- DM:
-
Dry matter
- GHG:
-
Greenhouse gas
- GLS:
-
Glucosinolates
- HT:
-
Hydrolyzable tannins
- IPCC:
-
Intergovernmental Panel on Climate Change
- LCA:
-
Life cycle assessments
- N:
-
Nitrogen
- N2O:
-
Nitrous oxide
- NH3 :
-
Ammonia
- NH4 + :
-
Ammonium
- NO3 − :
-
Nitrate
- NUE:
-
Nitrogen utilization efficiency
References
Eisler MC, Lee MRF, Tarlton JF, Martin GB, Beddington J, Dungait JAJ, et al. Agriculture: Steps to sustainable livestock. Nature. 2014;507:32–4.
de Klein CAM, Sherlock RR, Cameron KC, van der Weerden TJ. Nitrous oxide emissions from agricultural soils in New Zealand – a review of current knowledge and directions for future research. J Roy Soc New Zeal. 2001;31:543–74.
IPCC. Climate change 2022: Mitigation of climate change. Working Group III contribution to the sixth assessment report of the intergovernmental panel on climate change. In: Shukla PR, Skea J, Slade R, Al Khourdajie A, van Diemen R, McCollum D, et al., editors. Cambridge and New York: Cambridge University Press; 2022.
Ravishankara AR, Daniel JS, Portmann RW. Nitrous oxide (N2O): the dominant ozone–depleting substance emitted in the 21st century. Science. 2009;326:123–5.
Smith P, Martino D, Cai Z, Gwary D, Janzen H, Kumar P. Agriculture. In: Metz B, Davidson OR, Bosch PR, Dave R, Meyer LA, editors. Climate change 2007: mitigation. Cambridge and New York: Cambridge University Press; 2007.
Pulina G, Francesconi AHD, Stefanon B, Sevi A, Calamari L, Lacetera N, et al. Sustainable ruminant production to help feed the planet. Ital J Anim Sci. 2016;16:140–71.
Zhou K, Bao Y, Zhao G. Effects of dietary crude protein and tannic acid on nitrogen excretion, urinary nitrogenous composition and urine nitrous oxide emissions in beef cattle. J Anim Physiol Anim Nutr (Berl). 2019;103:1675–83.
Liu H, Zhou D. Mitigation of ammonia and nitrous oxide emissions from pasture treated with urine of sheep fed diets supplemented with sodium chloride. Anim Feed Sci Technol. 2014;192:39–47.
Coelho LD, Brito LD, Messana JD, Cardoso AD, Carvalho GM, Torres RDS, et al. Effects of rumen undegradable protein sources on nitrous oxide, methane and ammonia emission from the manure of feedlot–finished cattle. Sci Rep. 2022;12:9166.
Bao Y, Zhou K, Zhao GY. Nitrous oxide emissions from the urine of beef cattle as regulated by dietary crude protein and gallic acid. J Anim Sci. 2018;96:3699–711.
Lombardi B, Alvarado PI, Ricci P, Guzman SA, Gonda HL, Juliarena MP. Methane and nitrous oxide emissions from dung patches deposited by grazing cattle supplemented with maize grain. Anim Feed Sci Technol. 2021;279:115029.
de Klein CAM, Luo J, Woodward KB, Styles T, Wise B, Lindsey S, et al. The effect of nitrogen concentration in synthetic cattle urine on nitrous oxide emissions. Agric Ecosyst Environ. 2014;188:85–92.
Mazzetto AM, Barneze AS, Feigl BJ, Van Groenigen JW, Oenema O, De Klein CAM, et al. Use of the nitrification inhibitor dicyandiamide (DCD) does not mitigate N2O emission from bovine urine patches under Oxisol in Northwest Brazil. Nutrt Cycl Agroecosys. 2015;101:83–92.
Santoso B, Kilmaskossu A, Sambodo P. Effects of saponin from Biophytum petersianum Klotzsch on ruminal fermentation, microbial protein synthesis and nitrogen utilization in goats. Anim Feed Sci Technol. 2007;137:58–68.
Stewart EK, Beauchemin KA, Dai X, MacAdam JW, Christensen RG, Villalba JJ. Effect of tannin–containing hays on enteric methane emissions and nitrogen partitioning in beef cattle. J Anim Sci. 2019;97:3286–99.
Wu P, Liu ZB, He WF, Yu SB, Gao G, Wang JK. Intermittent feeding of citrus essential oils as a potential strategy to decrease methane production by reducing microbial adaptation. J Clean Prod. 2018;194:704–13.
Dietz M, Machill S, Hoffmann HC, Schmidtke K. Inhibitory effects of Plantago lanceolata L. on soil N mineralization. Plant Soil. 2013;368:445–58.
Totty VK, Greenwood SL, Bryant RH, Edwards GR. Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures. J Dairy Sci. 2013;96:141–9.
Luo J, Sun XZ, Pacheco D, Ledgard SF, Lindsey SB, Hoogendoom CJ, et al. Nitrous oxide emission factors for urine and dung from sheep fed either fresh forage rape (Brassica napus L.) or fresh perennial ryegrass (Lolium perenne L.). Animal. 2015;9:534–43.
Luo J, Balvert SF, Wise B, Welten B, Ledgard SF, de Klein CAM, et al. Using alternative forage species to reduce emissions of the greenhouse gas nitrous oxide from cattle urine deposited onto soil. Sci Total Environ. 2018;610:1271–80.
Simon PL, de Klein CAM, Worth W, Rutherford AJ, Dieckow J. The efficacy of Plantago lanceolata for mitigating nitrous oxide emissions from cattle urine patches. Sci Total Environ. 2019;691:430–41.
Saggar S. Estimation of nitrous oxide emission from ecosystems and its mitigation technologies. Sci Total Environ. 2010;136:189–91.
Carter MS. Contribution of nitrification and denitrification to N2O emissions from urine patches. Soil Biol Biochem. 2007;39:2091–102.
Dijkstra J, Oenema O, van Groenigen JW, Spek JW, van Vuuren AM, Bannink A. Diet effects on urine composition of cattle and N2O emissions. Animal. 2013;7:292–302.
Mancia A, Chadwick DR, Waters SM, Krol DJ. Uncertainties in direct N2O emissions from grazing ruminant excreta (EF3PRP) in national greenhouse gas inventories. Sci Total Environ. 2022;803:149935.
Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K. Agriculture, forestry and other land use. In: Eggleston HS, Buendia L, Miwa K, Ngara T, Tanabe K, editors. 2006 IPCC guidelines for national greenhouse gas inventories. Japan: IGES; 2006.
IPCC. Volume 4: agriculture, forestry and other land use. In: Calvo Buendia E, Tanabe K, Kranjc A, Baasansuren J, Fukuda M, Ngarize S, et al., editors. 2019 Refinement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories. Switzerland; 2019.
Bell MJ, Rees RM, Cloy JM, Topp CF, Bagnall A, Chadwick DR. Nitrous oxide emissions from cattle excreta applied to a Scottish grassland: effects of soil and climatic conditions and a nitrification inhibitor. Sci Total Environ. 2015;508:343–53.
Chadwick DR, Cardenas LM, Dhanoa MS, Donovan N, Misselbrook T, Williams JR, et al. The contribution of cattle urine and dung to nitrous oxide emissions: Quantification of country specific emission factors and implications for national inventories. Sci Total Environ. 2018;635:607–17.
Hoeft I, Steude K, Wrage N, Veldkamp E. Response of nitrogen oxide emissions to grazer species and plant species composition in temperate agricultural grassland. Agric Ecosyst Environ. 2012;151:34–43.
Luo JF, Saggar S, van der Weerden T, de Klein C. Quantification of nitrous oxide emissions and emission factors from beef and dairy cattle excreta deposited on grazed pastoral hill lands. Agric Ecosyst Environ. 2019;270:103–13.
Thomas BW, Gao X, Beck R, Hao X. Are distinct nitrous oxide emission factors required for cattle urine and dung deposited on pasture in western Canada? Environ Sci Pollut Res Int. 2017;24:26142–7.
Wachendorf C, Lampe C, Taube F, Dittert K. Nitrous oxide emissions and dynamics of soil nitrogen under 15 N-labeled cow urine and dung patches on a sandy grassland soil. J Plant Nutr Soil Sci. 2008;171:171–80.
Ma X, Wang S, Wang Y, Jiang G, Nyren P. Short-term effects of sheep excrement on carbon dioxide, nitrous oxide and methane fluxes in typical grassland of Inner Mongolia. New Zeal J Agric Res. 2010;49:285–97.
Wecking AR, Wall AM, Liang LL, Lindsey SB, Luo JF, Campbell DI, et al. Reconciling annual nitrous oxide emissions of an intensively grazed dairy pasture determined by eddy covariance and emission factors. Agric Ecosyst Environ. 2020;287:106646.
van der Weerden TJ, Luo JF, de Klein CAM, Hoogendoorn CJ, Littlejohn RP, Rys GJ. Disaggregating nitrous oxide emission factors for ruminant urine and dung deposited onto pastoral soils. Agric Ecosyst Environ. 2011;141:426–36.
Lombardi B, Alvarado PI, Ricci P, Buraschi LM, Viduzzi G, Palladino RA, et al. Combination of cattle urine and dung patches synergically increased nitrous oxide emissions from a temperate grassland under wet conditions. Agric Ecosyst Environ. 2022;340:108147.
Brice RM, Dele PA, Ike KA, Shaw YA, Olagunju LK, Orimaye OE, et al. Effects of essential oil blends on in vitro apparent and truly degradable dry matter, efficiency of microbial production, total short–chain fatty acids and greenhouse gas emissions of two dairy cow diets. Animals. 2022;12:2185.
Cieslak A, Zmora P, Pers-Kamczyc E, Szumacher-Strabel M. Effects of tannins source (Vaccinium vitis idaea L.) on rumen microbial fermentation in vivo. Anim Feed Sci Technol. 2012;176:102–6.
Shilwant S, Hundal JS, Singla M, Patra AK. Ruminal fermentation and methane production in vitro, milk production, nutrient utilization, blood profile, and immune responses of lactating goats fed polyphenolic and saponin–rich plant extracts. Environ Sci Pollut Res. 2022;30:10901–13.
Sun X. Invited Review: glucosinolates might result in low methane emissions from ruminants fed brassica forages. Front Vet Sci. 2020;7:588051.
Hagerman AE, Robbins CT, Weerasuriya Y, Wilson TC, McArthur C. Tannin chemistry in relation to digestion. Rangel Ecol Manag. 1992;45:57–62.
Jones GA, Jakober KD, Bae HD, McAllister TA, Cheng KJ. Some interactions between condensed tannins of forage legumes. Some interactions between condensed tannins of forage legumes, bovine serum albumin, and five strains of proteolytic rumen bacteria. Proceedings of the World Conference on Animal Production, vol. 36. Canada: Edmonton; 1993. p. 68–9.
Patra AK, Saxena J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J Sci Food Agric. 2011;91:24–37.
Gerlach K, Pries M, Tholen E, Schmithausen AJ, Buscher W, Sudekum KH. Effect of condensed tannins in rations of lactating dairy cows on production variables and nitrogen use efficiency. Animal. 2018;12:1847–55.
Koenig KM, Beauchemin KA, McGinn SM. Feeding condensed tannins to mitigate ammonia emissions from beef feedlot cattle fed high–protein finishing diets containing distillers grains. J Anim Sci. 2018;96:4414–30.
Deaville ER, Givens DI, Mueller-Harvey I. Chestnut and mimosa tannin silages: Effects in sheep differ for apparent digestibility, nitrogen utilisation and losses. Anim Feed Sci Technol. 2010;157:129–38.
Pathak AK, Dutta N, Pattanaik AK, Chaturvedi VB, Sharma K. Effect of condensed tannins from Ficus infectoria and Psidium guajava leaf meal mixture on nutrient metabolism, methane emission and performance of lambs. Asian-Australas J Anim Sci. 2017;30:1702–10.
Silveira Pimentel PR, Barcellos de Pellegrini C, Meira Galvão J, Santos Brant LM, Vaz di Mambro Ribeiro C, Mariniello Silva T, et al. Performance, digestibility, nitrogen balance and ingestive behaviour of goat kids fed diets supplemented with condensed tannins from Acacia mearnsii extract. Anim Prod Sci. 2021;61:1534–45.
Mueller-Harvey I, Bee G, Dohme-Meier F, Hoste H, Karonen M, Kölliker R, et al. Benefits of condensed tannins in forage legumes fed to ruminants: importance of structure, concentration, and diet composition. Crop Sci. 2019;59:861–85.
van Cleef FOSV, Dubeux JCJB, Ciriaco FM, Henry DD, Ruiz-Moreno M, Jaramillo DM, et al. Inclusion of a tannin–rich legume in the diet of beef steers reduces greenhouse gas emissions from their excreta. Sci Rep. 2022;12:14220.
Hao X, Benke MB, Li C, Larney FJ, Beauchemin KA, McAllister TA. Nitrogen transformations and greenhouse gas emissions during composting of manure from cattle fed diets containing corn dried distillers grains with solubles and condensed tannins. Anim Feed Sci Technol. 2011;166–167:539–49.
de Souza MN, Bayer C, Lassalas M, Michelon GM, Schaitz LH, Biasiolo R, et al. Effects of ground corn and Acacia mearnsii tannin extract supplementation on nitrogen excretion and nitrous oxide emissions from sheep. Livest Sci. 2021;246:104458.
Grainger C, Clarke T, Auldist MJ, Beauchemin KA, McGinn SM, Waghorn GC, et al. Potential use of Acacia mearnsii condensed tannins to reduce methane emissions and nitrogen excretion from grazing dairy cows. Can J Anim Sci. 2009;89:241–51.
Maamouri O, Atti N, Kraiem K, Mahouachi M. Effects of concentrate and Acacia cyanophylla foliage supplementation on nitrogen balance and milk production of grazing ewes. Livest Sci. 2011;139:264–70.
Kronberg SL, Liebig MA. Condensed tannin in drinking water reduces greenhouse gas precursor urea in sheep and cattle urine. Rangeland Ecol Manag. 2011;64:543–7.
Ahnert S, Dickhoefer U, Schulz F, Susenbeth A. Influence of ruminal Quebracho tannin extract infusion on apparent nutrient digestibility, nitrogen balance, and urinary purine derivatives excretion in heifers. Livest Sci. 2015;177:63–70.
Min BR, Solaiman S, Terrill T, Ramsay A, Mueller-Harvey I. The effects of tannins–containing ground pine bark diet upon nutrient digestion, nitrogen balance, and mineral retention in meat goats. J Anim Sci Biotechnol. 2015;6:25.
Aguerre MJ, Capozzolo MC, Lencioni P, Cabral C, Wattiaux MA. Effect of quebracho–chestnut tannin extracts at 2 dietary crude protein levels on performance, rumen fermentation, and nitrogen partitioning in dairy cows. J Dairy Sci. 2016;99:4476–86.
Gunun P, Wanapat M, Gunun N, Cherdthong A, Sirilaophaisan S, Kaewwongsa W. Effects of condensed tannins in Mao (Antidesma thwaitesianum Muell. Arg.) seed meal on rumen fermentation characteristics and nitrogen utilization in goats. Asian-Australas J Anim Sci. 2016;29:1111–9.
Koenig KM, Beauchemin KA. Effect of feeding condensed tannins in high protein finishing diets containing corn distillers grains on ruminal fermentation, nutrient digestibility, and route of nitrogen excretion in beef cattle. J Anim Sci. 2018;96:4398–413.
Zhang J, Xu X, Cao Z, Wang Y, Yang H, Azarfar A, et al. Effect of different tannin sources on nutrient intake, digestibility, performance, nitrogen utilization, and blood parameters in dairy cows. Animals (Basel). 2019;9:507.
Lagrange S, Beauchemin KA, MacAdam J, Villalba JJ. Grazing diverse combinations of tanniferous and non–tanniferous legumes: Implications for beef cattle performance and environmental impact. Sci Total Environ. 2020;746:140788.
da Silva Aguiar F, Bezerra LR, Cordao MA, Cavalcante ITR, de Oliveira JPF, do Nascimento RR, et al. Effects of increasing levels of total tannins on intake, digestibility, and balance of nitrogen, water, and energy in hair lambs. Animals (Basel). 2023;13:2497.
Uushona T, Chikwanha OC, Katiyatiya CLF, Strydom PE, Mapiye C. Substitution effects of Acacia mearnsii leaf–meal for Triticum aestivum bran on nutrient digestibility, rumen fermentation and nitrogen retention in lambs. Small Ruminant Res. 2023;221:106948.
Oliveira LN, Pereira MAN, Oliveira CDS, Oliveira CC, Silva RB, Pereira RAN, et al. Effect of low dietary concentrations of Acacia mearnsii tannin extract on chewing, ruminal fermentation, digestibility, nitrogen partition, and performance of dairy cows. J Dairy Sci. 2023;106:3203–16.
Heckendorn F, Haring DA, Maurer V, Zinsstag J, Langhans W, Hertzberg H. Effect of sainfoin (Onobrychis viciifolia) silage and hay on established populations of Haemonchus contortus and Cooperia curticei in lambs. Vet Parasitol. 2006;142:293–300.
PerezMaldonado RA, Norton BW. The effects of condensed tannins from Desmodium intortum and Calliandra calothyrsus on protein and carbohydrate digestion in sheep and goats. Brit J Nutr. 1996;76:515–33.
Terrill TH, Waghorn GC, Woolley DJ, McNabb WC, Barry TN. Assay and digestion of 14C-labeled condensed tannins in the gastrointestinal tract of sheep. Brit J Nutr. 1994;72:467–77.
Misselbrook TH, Powell JM, Broderick GA, Grabber JH. Dietary manipulation in dairy cattle: laboratory experiments to assess the influence on ammonia emissions. J Dairy Sci. 2005;88:1765–77.
Powell JM, Aguerre MJ, Wattiaux MA. Tannin extracts abate ammonia emissions from simulated dairy barn floors. J Environ Qual. 2011;40:907–14.
Ingold M, Wachendorf C, Buerkert A. Net–mineralization of organic matter and greenhouse gas emissions from Quebracho tannin–enriched manure applied to acidic and alkaline soils. J Plant Nutr Soil Sci. 2021;184:530–42.
Min BR, Parker D, Casey K, Willis W, Castleberry L, Meyer B, et al. The effect of plant tannins on methane and nitrous oxide emissions from dairy manure under laboratory conditions. J Anim Sci. 2020;98:387.
Fox RH, Myers RJK, Vallis I. The nitrogen mineralization rate of legume residues in soil as influenced by their polyphenol, lignin, and nitrogen contents. Plant Soil. 1990;129:251–9.
Palm CA, Sanchez PA. Nitrogen release from the leaves of some tropical legumes as affected by their lignin and polyphenolic contents. Soil Biol Biochem. 1991;23:83–8.
Fagundes GM, Benetel G, Carriero MM, Sousa RLM, Santos KC, Muir JP, et al. Dietary condensed tannins in bovine faeces and effects on soil microbial dynamics: are there environmental benefits for cattle production systems? Anim Prod Sci. 2021;61:690–7.
Mueller-Harvey I. Unravelling the conundrum of tannins in animal nutrition and health. J Sci Food Agric. 2006;86:2010–37.
Getachew G, Pittroff W, Putnam DH, Dandekar A, Goyal S, DePeters EJ. The influence of addition of gallic acid, tannic acid, or quebracho tannins to alfalfa hay on in vitro rumen fermentation and microbial protein synthesis. Anim Feed Sci Technol. 2008;140:444–61.
Liu H, Vaddella V, Zhou D. Effects of chestnut tannins and coconut oil on growth performance, methane emission, ruminal fermentation, and microbial populations in sheep. J Dairy Sci. 2011;94:6069–77.
Aboagye IA, Oba M, Castillo AR, Koenig KM, Iwaasa AD, Beauchemin KA. Effects of hydrolyzable tannin with or without condensed tannin on methane emissions, nitrogen use, and performance of beef cattle fed a high–forage diet. J Anim Sci. 2018;96:5276–86.
Wischer G, Greiling AM, Boguhn J, Steingass H, Schollenberger M, Hartung K, et al. Effects of long–term supplementation of chestnut and valonea extracts on methane release, digestibility and nitrogen excretion in sheep. Animal. 2014;8:938–48.
Wei C, Yang K, Zhao G, Lin S, Xu Z. Effect of dietary supplementation of gallic acid on nitrogen balance, nitrogen excretion pattern and urinary nitrogenous constituents in beef cattle. Arch Anim Nutr. 2016;70:416–23.
Yang K, Wei C, Zhao G, Xu Z, Lin S. Dietary supplementation of tannic acid modulates nitrogen excretion pattern and urinary nitrogenous constituents of beef cattle. Livest Sci. 2016;191:148–52.
Aboagye IA, Oba M, Koenig KM, Zhao GY, Beauchemin KA. Use of gallic acid and hydrolyzable tannins to reduce methane emission and nitrogen excretion in beef cattle fed a diet containing alfalfa silage. J Anim Sci. 2019;97:2230–44.
Herremans S, Decruyenaere V, Cantalapiedra-Hijar G, Beckers Y, Froidmont E. Effects of hydrolysable tannin–treated grass silage on milk yield and composition, nitrogen partitioning and nitrogen isotopic discrimination in lactating dairy cows. Animal. 2020;14:771–9.
Kapp-Bitter A, Dickhoefer U, Suglo E, Baumgartner L, Kreuzer M, Leiber F. Graded supplementation of chestnut tannins to dairy cows fed protein–rich spring pasture: effects on indicators of protein utilization. J Anim Feed Sci. 2020;29:97–104.
Reed JD. Nutritional toxicology of tannins and related polyphenols in forage legumes. J Anim Sci. 1995;73:1516–28.
Makkar HPS. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin–rich feeds. Small Ruminant Res. 2003;49:241–56.
Martin AK. The origin of urinary aromatic–compounds excreted by ruminants. 2. The metabolism of phenolic cinnamic–acids to benzoic–acid. Brit J Nutr. 1982;47:155–64.
Kool D, Hoffland E, Hummelink E, Vangroenigen J. Increased hippuric acid content of urine can reduce soil N2O fluxes. Soil Biol Biochem. 2006;38:1021–7.
Willem van groenigen J, Palermo V, Kool DM, Kuikman PJ. Inhibition of denitrification and N2O emission by urine–derived benzoic and hippuric acid. Soil Biol Biochem. 2006;38:2499–502.
Bertram JE, Clough TJ, Sherlock RR, Condron LM, O’Callaghan M, Wells NS, et al. Hippuric acid and benzoic acid inhibition of urine derived N2O emissions from soil. Global Change Biol. 2009;15:2067–77.
Clough TJ, Ray JL, Buckthought LE, Calder J, Baird D, O’Callaghan M, et al. The mitigation potential of hippuric acid on N2O emissions from urine patches: An in situ determination of its effect. Soil Biol Biochem. 2009;41:2222–9.
Krol DJ, Forrestal PJ, Lanigan GJ, Richards KG. In situ N2O emissions are not mitigated by hippuric and benzoic acids under denitrifying conditions. Sci Total Environ. 2015;511:362–8.
Fahey JW, Zalcmann AT, Talalay P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry. 2001;56:5–51.
Barry TN. The feeding value of forage brassica plants for grazing ruminant livestock. Anim Feed Sci Technol. 2013;181:15–25.
Prieto MA, López CJ, Simal-Gandara J. Chapter Six – Glucosinolates: Molecular structure, breakdown, genetic, bioavailability, properties and healthy and adverse effects. In: Ferreira ICFR, Barros L, editor. Advances in Food and Nutrition Research, vol. 90. Academic Press; 2019. p. 305–50.
Bending GD, Lincoln SD. Inhibition of soil nitrifying bacteria communities and their activities by glucosinolate hydrolysis products. Soil Biol Biochem. 2000;32:1261–9.
Brown PD, Morra MJ. Brassicaceae tissues as inhibitors of nitrification in soil. J Agric Food Chem. 2009;57:7706–11.
Reardon CL, Strauss SL, Mazzola M. Changes in available nitrogen and nematode abundance in response to Brassica seed meal amendment of orchard soil. Soil Biol Biochem. 2013;57:22–9.
Estell RE. Coping with shrub secondary metabolites by ruminants. Small Ruminant Res. 2010;94:1–9.
Hoogendoorn CJ, Luo J, Lloyd-West CM, Devantier BP, Lindsey SB, Sun S, et al. Nitrous oxide emission factors for urine from sheep and cattle fed forage rape (Brassica napus L.) or perennial ryegrass/white clover pasture (Lolium perenne L./Trifolium repens). Agric Ecosyst Environ. 2016;227:11–23.
Munday R, Mhawech-Fauceglia P, Munday CM, Paonessa JD, Tang L, Munday JS, et al. Inhibition of urinary bladder carcinogenesis by broccoli sprouts. Cancer Res. 2008;68:1593–600.
Ye LX, Dinkova-Kostova AT, Wade KL, Zhang YS, Shapiro TA, Talalay P. Quantitative determination of dithiocarbamates in human plasma, serum, erythrocytes and urine: pharmacokinetics of broccoli sprout isothiocyanates in humans. Clin Chim Acta. 2002;316:43–53.
Balvert SF, Luo J, Schipper LA. Do glucosinolate hydrolysis products reduce nitrous oxide emissions from urine affected soil? Sci Total Environ. 2017;603–604:370–80.
Gao J, Cheng B, Sun Y, Zhao Y, Zhao G. Effects of dietary inclusion with rapeseed cake containing high glucosinolates on nitrogen metabolism and urine nitrous oxide emissions in steers. Anim Nutr. 2022;8:204–15.
Box LA, Edwards GR, Bryant RH. Milk production and urinary nitrogen excretion of dairy cows grazing perennial ryegrass–white clover and pure plantain pastures. Proc N Z Soc Anim Prod. 2016;76:18–21.
Cheng L, Judson HG, Bryant RH, Mowat H, Guinot L, Hague H, et al. The effects of feeding cut plantain and perennial ryegrass–white clover pasture on dairy heifer feed and water intake, apparent nutrient digestibility and nitrogen excretion in urine. Anim Feed Sci Technol. 2017;229:43–6.
Minnée EMK, Leach CMT, Dalley DE. Substituting a pasture–based diet with plantain (Plantago lanceolata) reduces nitrogen excreted in urine from dairy cows in late lactation. Livest Sci. 2020;239:104093.
Marshall CJ, Beck MR, Garrett K, Barrell GK, Al Marashdeh O, Gregorini P. Nitrogen balance of dairy cows divergent for milk urea nitrogen breeding values consuming either plantain or perennial ryegrass. Animals (Basel). 2021;11:2464.
Ineichen S, Marquardt S, Wettstein HR, Kreuzer M, Reidy B. Milk fatty acid profile and nitrogen utilization of dairy cows fed ryegrass–red clover silage containing plantain (Plantago lanceolata L.). Livest Sci. 2019;221:123–32.
Nkomboni D, Bryant RH, Edwards GR. Effect of increasing dietary proportion of plantain on milk production and nitrogen use of grazing dairy cows in late lactation. Anim Prod Sci. 2021;61:770–9.
Al-Marashdeh O, Cameron K, Hodge S, Gregorini P, Edwards G. Integrating plantain (Plantago lanceolata L.) and Italian ryegrass (Lolium multiflorum Lam.) into New Zealand grazing dairy system: The effect on farm productivity, profitability, and nitrogen losses. Animals (Basel). 2021;11:376.
McMurphy CP, Sexten AJ, Mourer GL, Sharman ED, Trojan SJ, Rincker MJ, et al. Effects of including saponins (Micro–Aid®) on intake, rumen fermentation and digestibility in steers fed low–quality prairie hay. Anim Feed Sci Technol. 2014;190:47–58.
Guyader J, Eugène M, Doreau M, Morgavi DP, Gérard C, Loncke C, et al. Nitrate but not tea saponin feed additives decreased enteric methane emissions in nonlactating cows. J Anim Sci. 2015;93:5367–77.
Guyader J, Eugene M, Doreau M, Morgavi DP, Gerard C, Martin C. Tea saponin reduced methanogenesis in vitro but increased methane yield in lactating dairy cows. J Dairy Sci. 2017;100:1845–55.
Liu Y, Ma T, Chen D, Zhang N, Si B, Deng K, et al. Effects of tea saponin supplementation on nutrient digestibility, methanogenesis, and ruminal microbial flora in dorper crossbred ewe. Animals (Basel). 2019;9:29.
Snyder AJ, Johnson Maynard JL, Morra MJ. Nitrogen mineralization in soil incubated with 15N–labeled Brassicaceae seed meals. Appl Soil Ecol. 2010;46:73–80.
Gao J, Sun Y, Bao Y, Zhou K, Kong D, Zhao G. Effects of different levels of rapeseed cake containing high glucosinolates in steer ration on rumen fermentation, nutrient digestibility and the rumen microbial community. Brit J Nutr. 2021;125:266–74.
Subuh AMH, Rowan TG, Lawrence TLJ. Toxic moieties in ruminal and duodenal digesta and in milk, and hepatic integrity in cattle given diets based on rapeseed meals of different glucosinolate contents either untreated or treated with heat or formaldehyde. Anim Feed Sci Technol. 1995;52:51–61.
Gao J, Li MM, Zhao G. Thiocyanate increases the nitrous oxide formation process through modifying the soil bacterial community. J Sci Food Agric. 2022;102:2321–9.
Kim DH, Kim BR, Kim JY, Jeong YC. Mechanism of covalent adduct formation of aucubin to proteins. Toxicol Lett. 2000;114:181–8.
Busto MD, Perez-Mateos M. Characterization of beta–D–glucosidase extracted from soil fractions. European J Soil Sci. 2000;51:193–200.
Marak HB, Biere A, Van Damme JM. Systemic, genotype–specific induction of two herbivore–deterrent iridoid glycosides in Plantago lanceolata L. in response to fungal infection by Diaporthe adunca (Rob.) Niessel. J Chem Ecol. 2002;28:2429–48.
Cheng L, McCormick J, Hussein AN, Logan C, Pacheco D, Hodge MC, et al. Live weight gain, urinary nitrogen excretion and urination behaviour of dairy heifers grazing pasture, chicory and plantain. J Agric Sci. 2017;155:669–78.
O’Connell CA, Judson HG, Barrell GK. Sustained diuretic effect of plantain when ingested by sheep. Proc N Z Soc Anim Prod. 2016;76:14–7.
Navarrete S, Kemp PD, Pain SJ, Back PJ. Bioactive compounds, aucubin and acteoside, in plantain (Plantago lanceolata L.) and their effect on in vitro rumen fermentation. Anim Feed Sci Technol. 2016;222:158–67.
Navarrete S, Rodriguez M, Horne D, Hanly J, Hedley M, Kemp P. Nitrogen excretion by dairy cows grazing plantain (Plantago lanceolata) based pastures during the lactating season. Animals. 2022;12:469.
Massaccesi L, Bardgett RD, Agnelli A, Ostle N, Wilby A, Orwin KH. Impact of plant species evenness, dominant species identity and spatial arrangement on the structure and functioning of soil microbial communities in a model grassland. Oecologia. 2015;177:747–59.
Carlton AJ, Cameron KC, Di HJ, Edwards GR, Clough TJ. Nitrate leaching losses are lower from ryegrass/white clover forages containing plantain than from ryegrass/white clover forages under different irrigation. New Zeal J Agric Res. 2019;62:150–72.
Pijlman J, Berger SJ, Lexmond F, Bloem J, van Groenigen JW, Visser EJW, et al. Can the presence of plantain (Plantago lanceolata L.) improve nitrogen cycling of dairy grassland systems on peat soils? New Zeal J Agric Res. 2019;63:106–22.
Pankoke H, Müller C. Impact of defoliation on the regrowth capacity and the shoot metabolite profile of Plantago lanceolata L. Plant Physiol Biochem. 2013;71:325–33.
Bartholomaeus A, Ahokas J. Inhibition of P-450 by aucubin: is the biological–activity of aucubin due to its glutaraldehyde-like aglycone. Toxicol Lett. 1995;80:75–83.
Davini E, Iavarone C, Trogolo C, Aureli P, Pasolini B. The quantitative isolation and antimicrobial activity of the aglycone of aucubin. Phytochemistry. 1986;25:2420–2.
Stewart AV. Plantain (Plantago lanceolata) – a potential pasture species. Proceedings of the New Zealand Grassland Association. 1996;58:77–86.
Judson H, Fraser P, Peterson M. Nitrification inhibition by urine from cattle consuming Plantago lanceolata. J New Zeal Grasslands. 2019;81:111–6.
Gardiner CA, Clough TJ, Cameron KC, Di HJ, Edwards GR. Efficacy of aucubin as a nitrification inhibitor assessed in two Canterbury field trials. New Zeal J Agric Res. 2019;63:73–86.
Rodriguez MJ, Navarrete S, Horne DJ, Hanly JA, Bishop P, Kemp PD. Can secondary metabolites of plantain reduce N losses from urine patches? New Zeal J Agric Res. 2023;66:83–100.
Zúñiga Serrano A, Barrios García HB, Anderson RC, Hume ME, Ruizalbarrán M, Bautista Martínez Y, et al. Antimicrobial and digestive effects of Yucca schidigera extracts related to production and environment implications of ruminant and non–ruminant animals: A review. Agriculture. 2022;12:1198.
Patra AK, Saxena J. The effect and mode of action of saponins on the microbial populations and fermentation in the rumen and ruminant production. Nutr Res Rev. 2009;22:204–19.
Cheeke PR. Actual and potential applications of Yucca schidigera and Quillaja saponaria saponins in human and animal nutrition. In: Oleszek W, Marston A, editors. Saponins in food, feedstuffs and medicinal plants. Dordrecht: Springer; 2000;45:241–54.
Jouany JP. Effect of rumen protozoa on nitrogen utilization by ruminants. J Nutr. 1996;126:1335S–1346S.
Hu WL, Liu JX, Ye JA, Wu YM, Guo YQ. Effect of tea saponin on rumen fermentation in vitro. Anim Feed Sci Technol. 2005;120:333–9.
Wina E, Muetzel S, Becker K. The impact of saponins or saponin–containing plant materials on ruminant production: A review. J Agric Food Chem. 2005;53:8093–105.
Ramírez-Restrepo CA, O’Neill CJ, López-Villalobos N, Padmanabha J, Wang JK, McSweeney C. Effects of tea seed saponin supplementation on physiological changes associated with blood methane concentration in tropical Brahman cattle. Anim Prod Sci. 2016;56:457–65.
Golbotteh MM, Malecky M, Aliarabi H, Zamani P, Ganjkhanlou M. Dose–response effects of the Savory (Satureja khuzistanica) essential oil and extract on rumen fermentation characteristics, microbial protein synthesis and methane production in vitro. Ann Anim Sci. 2022;22:1001–14.
Patra AK, Yu Z. Effects of vanillin, quillaja saponin, and essential oils on in vitro fermentation and protein–degrading microorganisms of the rumen. Appl Microbiol Biotechnol. 2014;98:897–905.
Pawar MM, Kamra DN, Agarwal N, Chaudhary LC. Effects of essential oils on in vitro methanogenesis and feed fermentation with buffalo rumen liquor. Agric Res. 2014;3:67–74.
Lin B, Lu Y, Salem AZM, Wang JH, Liang Q, Liu JX. Effects of essential oil combinations on sheep ruminal fermentation and digestibility of a diet with fumarate included. Anim Feed Sci Technol. 2013;184:24–32.
Toseti LB, Goulart RS, Gouvêa VN, Acedo TS, Vasconcellos GSFM, Pires AV, et al. Effects of a blend of essential oils and exogenous α–amylase in diets containing different roughage sources for finishing beef cattle. Anim Feed Sci Technol. 2020;269:114643.
Carrazco AV, Peterson CB, Zhao Y, Pan Y, McGlone JJ, DePeters EJ, et al. The impact of essential oil feed supplementation on enteric gas emissions and production parameters from dairy cattle. Sustainability. 2020;12:10347.
Dorman HJ, Deans SG. Antimicrobial agents from plants: antibacterial activity of plant volatile oils. J Appl Microbiol. 2000;88:308–16.
Burt S. Essential oils: their antibacterial properties and potential applications in foods––a review. Int J Food Microbiol. 2004;94:223–53.
Wanapat M, Khejornsart P, Pakdee P, Wanapat S. Effect of supplementation of garlic powder on rumen ecology and digestibility of nutrients in ruminants. J Sci Food Agric. 2008;88:2231–7.
Ribeiro ADB, Junior M, Polizel DM, Miszura AA, Gobato LGM, Barroso JPR, et al. Thyme essential oil for sheep: effect on rumen fermentation, nutrient digestibility, nitrogen metabolism, and growth. Arq Bras Med Vet Zoo. 2019;71:2065–74.
Orzuna-Orzuna JF, Dorantes Iturbide G, Lara Bueno A, Miranda-Romero LA, Mendoza Martínez GD, Santiago-Figueroa I. A meta–analysis of essential oils use for beef cattle feed: rumen fermentation, blood metabolites, meat quality, performance and environmental and economic impact. Fermentation. 2022;8:254.
Wanapat M, Kang S, Khejornsart P, Wanapat S. Effects of plant herb combination supplementation on rumen fermentation and nutrient digestibility in beef cattle. Asian Austral J Anim Sci. 2013;26:1127–36.
Tekippe JA, Tacoma R, Hristov AN, Lee C, Oh J, Heyler KS, et al. Effect of essential oils on ruminal fermentation and lactation performance of dairy cows. J Dairy Sci. 2013;96:7892–903.
Oh J, Harper M, Hristov AN. Effects of lowering crude protein supply alone or in a combination with essential oils on productivity, rumen function and nutrient utilization in dairy cows. Animal. 2019;13:2510–8.
Benchaar C. Diet supplementation with thyme oil and its main component thymol failed to favorably alter rumen fermentation, improve nutrient utilization, or enhance milk production in dairy cows. J Dairy Sci. 2021;104:324–36.
Muñoz-Cuautle A, Ortega-Cerrilla ME, Herrera-Haro JG, Nava-Cuellar C, Gutiérrez-Olvera C, Ramírez-Bribiesca JE, et al. Effect of oregano (Lippia graveolens) essential oil as a phytogenic feed additive on productive performance, ruminal fermentation, and antioxidant activity in lamb meat. Agriculture. 2022;12:973.
Wallace RJ, McEwan NR, McIntosh FM, Teferedegne B, Newbold CJ. Natural products as manipulators of rumen fermentation. Asian Austral J Anim Sci. 2002;15:1458–68.
Cobellis G, Trabalza-Marinucci M, Yu Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci Total Environ. 2016;545–546:556–68.
Naumann H, Muir J, Lambert B, Tedeschi L, Kothmann MM. Condensed tannins in the ruminant environment: a perspective on biological activity. J Agric Sci. 2013;1:8–20.
Waghorn G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production—Progress and challenges. Anim Feed Sci Technol. 2008;147:116–39.
Tripathi MK, Mishra AS. Glucosinolates in animal nutrition: A review. Anim Feed Sci Technol. 2007;132:1–27.
Funding
This study was supported by Beijing Captial Agribusiness & Food Group Co., Ltd. ‘ Integrated Innovation and Industrial Application of "Green Digital Intelligence" Technology in the Whole Dairy Industry Chain’ (SNSPKJ2022) and National Natural Science Foundation of China (32302767).
Author information
Authors and Affiliations
Contributions
YCZ: Conceptualization, Writing-original draft, Funding acquisition. ML: Methodology. LSJ: Supervision, Writing-review & editing. LLG: Writing-review & editing. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhao, Y., Liu, M., Jiang, L. et al. Could natural phytochemicals be used to reduce nitrogen excretion and excreta-derived N2O emissions from ruminants?. J Animal Sci Biotechnol 14, 140 (2023). https://doi.org/10.1186/s40104-023-00942-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-023-00942-0