
Abstract
Intestinal health is essential for the resistance to enteric diseases and for nutrient digestion and absorption to support growth. The intestine of nursery pigs are immature and vulnerable to external challenges, which cause negative impacts on the structure and function of the intestine. Among nutritional interventions, the benefits of milk are significant for the intestinal health of pigs. Milk coproducts have traditionally been used in starter feeds to improve the growth of nursery pigs, but their use is somewhat limited due to the high costs and potential risks of excessive lactose on the intestine. Thus, understanding a proper feeding level of milk carbohydrates is an important start of the feeding strategy. For nursery pigs, lactose is considered a highly digestible energy source compared with plant-based starch, whereas milk oligosaccharides are considered bioactive compounds modulating intestinal immunity and microbiota. Therefore, milk carbohydrates, mainly composed of lactose and oligosaccharides, have essential roles in the intestinal development and functions of nursery pigs. The proper feeding levels of lactose in starter feeds could be variable by weaning age, body weight, or genetic lines. Effects of lactose and milk oligosaccharides have been broadly studied in human health and animal production. Therefore, this review focuses on the mechanisms of lactose and milk oligosaccharides affecting intestinal maturation and functions through modulation of enterocyte proliferation, intestinal immunity, and intestinal microbiota of nursery pigs.
Similar content being viewed by others


Introduction
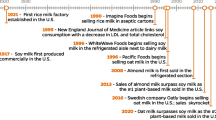
Swine industry has been facing with numerous challenges, including outbreaks of epidemic enteric diseases and removal of antimicrobial growth promoters in feeds which directly affect the efficiency and profitability of pig production. As a result, there have been increased efforts to investigate and find alternative nutraceuticals replacing the use of antibiotics to support intestinal health and growth of nursery pigs [1,2,3]. Intestinal challenges to nursery pigs are primarily caused by their immature intestinal system, which would hard to allow the feeds containing allergenic compounds from plant-based protein supplements as well as microbial infections [4, 5]. In general, sow milk is a natural source of nutrients for sucking piglets during lactation whereas the milk coproducts have been broadly used in milk replacers, creep feeds, and early weaner feeds. Milk coproducts have great potentials in feeds for nursery pigs because nutrients in milk are well balanced and highly digestible, supporting the intestinal health and growth of nursery pigs [6,7,8]. It has also been well documented that milk carbohydrates are most effectively utilized by nursery pigs providing energy and functional properties to support their growth and health [7, 9, 10].
Milk coproducts contain lactose as the major carbohydrate that is highly digestible compared with other carbohydrate sources in cereal grains for nursery pigs [11,12,13]. The main reason for high lactose digestibility would be related to early adaptation of their digestive system to maintain high lactase activity in the intestinal epithelial cells [14, 15]. In general, milk coproducts have been broadly used to supply lactose in nursery feeds. However, feeding lactose over a tolerance level would cause severe digestive problems from massive lactose fermentation in the intestine, causing negative impacts on intestinal development [16,17,18]. There is still a lack of updated information on the recommendation levels of lactose for nursery pigs with the changes in lean gain potential, health status, and weaning ages. Previous studies investigating an optimal lactose level in nursery feeds have shown significant variations due to changes in the genetics of pigs and management environments. However, one of the consistent results is that optimal lactose levels in feeds decrease as a pig grows [7, 10, 19].
Recent evidence suggests that animal milk and milk coproducts contain various forms of milk oligosaccharides with beneficial prebiotic effects on intestinal health and brain development in infants [20,21,22,23]. Milk oligosaccharides are non-digestible carbohydrates and the third-abundant-nutrients next to lactose and fat in animal milk [24]. These milk oligosaccharides are largely diverse and complex with their structure compared with conventional prebiotics such as galacto-oligosaccharides, xylo-oligosaccharides, or fructo-oligosaccharides which are current used in infant formula [25,26,27,28]. Once ingested, milk oligosaccharides would not be absorbed but pass through the gastrointestinal tract (GIT) and then be utilized for fermentation by intestinal microbes resulting in an increased proportion of potentially beneficial bacteria, including Bifidobacterium and Lactobacillus sp. [29,30,31]. Interestingly, a minimal amount of milk oligosaccharides could be absorbed into systemic circulation in young animals, possibly affecting the activation of the immune system [32,33,34]. Due to the diversity and specific structures of milk oligosaccharides, their mode of action for beneficial effects on young animals may be distinctively different from that of conventional prebiotics.
Therefore, there is growing interest in finding effective feeding values of milk carbohydrates for their nutritional and functional roles in the intestinal development and health of nursery pigs. This review focus on the functional role and feeding application of milk carbohydrates in modulating the intestinal health of nursery pigs.
Lactose
Lactose is the most abundant carbohydrate in milk, which consists of galactose and glucose by β-glycosidic linkage. It can be hydrolyzed by lactase (β-D-galactosidase) secreted from epithelial cells lining villi on the small intestine. After the hydrolysis of lactose, galactose and glucose are transported to central circulatory system via Na+-D-glucose cotransporter 1 (SGLT1) and glucose transporter 2 (GLUT2) by facilitated diffusion across brush border and basolateral membranes of enterocytes in the intestine [35]. Once absorbed, these monosaccharides are utilized as carbon donors to generate adenosine triphosphate (ATP) in energy metabolism and assist amino acids (AA) synthesis. In particular, galactose would be converted to glucose-1-phosphate through the Leloir pathway, and then it could be used in the cellular respiration process to produce energy [36]. Despite playing such a direct role as an energy source, lactose could also be involved in AA synthesis and other potential carbon chains indirectly contributing to growth [37,38,39].
Although pigs have a digestive system with a relatively high tolerance level of lactose compared with other species [40], milk coproducts are minimally used in swine feeds due to the high costs compared with the use of other carbohydrates from cereal grains. Thus, feeding a proper amount of lactose in feeds is important for successful pig production, balancing the feed costs and growth of nursery pigs. Nursing piglets can get lactose from sow milk. Lactose composition in sow milk is about 3% (or 13% dry matter (DM) basis) in the colostrum and about 5% (27% DM basis) in the mature milk [9, 11, 41]. The digestive system of nursing piglets, therefore, would maintain high lactase activity [14, 15, 42]. Thus, the use of milk coproducts has been a standard practice to supply effective carbohydrate sources in nursery feeds to improve the growth performance of nursery pigs. However, the suggested optimal inclusion levels of lactose for maximal growth efficiency vary depending on studies. Therefore, a depth review is needed to understand the variability and to provide comprehensive data analysis.
Meta-analysis to estimate lactose requirements for nursery pigs
In this review, a meta-analysis was conducted to determine the optimal dietary need of lactose for nursery pigs based on their growth performance. Data from published articles were obtained from public databases including PubMed, Science Direct, and Web of Science with criteria for filtering. For the criteria, all data from articles consisted of peer-reviewed publications. Secondly, data were obtained from animal experiments testing the effects of different levels of crystalline lactose or milk coproducts as a source of lactose. Lastly, all of the data for growth parameters, including BW, ADG, ADFI, and G:F were obtained or could be calculated from the data.
The comprehensive data provided in this review was obtained from 17 peer-reviewed publications and utilized for the meta-analysis [7, 8, 12, 19, 43,44,45,46,47,48,49,50,51,52,53,54,55]. The data were divided into three groups based on BW of pigs (for 5 to 7 kg BW, published in 10 papers with the 14 experiments; for 7 to 11 kg BW, published in 14 papers with 23 experiments; for 11 to 25 kg BW, published in 12 papers with 15 experiments). Daily lactose intake was calculated using average daily feed intake (g/d) multiplied by the lactose composition in each experimental diet. For statistical analysis, the inclusion level and daily intake of lactose on growth response were evaluated with nonlinear models using the Proc NLMIXED in SAS (SAS. Inc., Cary, USA). The method of nonlinear least squares used to fit the model: Y = a + b (R-x) + e. Where ‘Y’ is the response of feed efficiency, ‘a’ is the ordinate of the breakpoint, ‘b’ is the slope of the line when x < R, ‘x’ is if the inclusion level or average daily lactose intake, ‘R’ is the breakpoint, ‘e’ is a residual error among experiments, and ‘(R − x)’ is defined as zero when x ≥ R. Using a broken-line analysis, optimal levels of nutrient for the parameters of interest can be obtained [7, 56, 57]. Regressions were obtained between lactose levels (%) or intake (g/d) in nursery feeds and the feed efficiency of pigs. Optimal levels (breakpoints) of lactose to maximize feed efficiency were obtained at 20.0% in feeds for 5 to 7 kg body weight (BW) and 13.0% in feeds for 7 to 11 kg BW of nursery pigs (Fig. 1) or at 46 g/d lactose intake for 5 to 7 kg BW and 57 g/d lactose intake for 7 to 11 kg BW of pigs (Fig. 2), respectively. As expected, optimal inclusion levels of lactose for maximal feed efficiency decreased, whereas optimal daily lactose intakes were increased as pig BW increased. A recent review by Zhao et al. [58] demonstrated that nursery pigs need 20% lactose during d 0 to 7 after weaning, 15% lactose during d 7 to 14, and 0% lactose during d 14 to 35. Following these estimations, a previous study by Jang et al. [7] demonstrated that the growth responses to lactose supplementation was gradually decreased as pigs increased with age.

Changes in G:F of nursery pigs fed diets by inclusion level of lactose in diets during 5 to 7 kg (A) and 7 to 11 kg (B) BW using a broken-line analysis. The meta-analysis is conducted by Proc NLMIXED in SAS 9.4 to determine break-points on the regression of feed efficiency of nursery pigs calculated based on the data from 10 published studies (Phase 1 for 5 to 7 kg BW: 14 experiments) and 14 published studies (Phase 2 for 7 to 11 kg BW: 23 experiments). The breakpoints (one-slope broken-line model) were inclusion levels of lactose at 20% during 5 to 7 kg BW and 13% during 7 to 11 kg BW when G:F were 0.80 and 0.77, respectively. One-slope broken-line models; The equation for G:F during 5 to 7 kg BW was G:F = 0.80–0.39 × 10− 2 × z1, R2 = 0.90. The equation for G:F during 7 to 11 kg BW was G:F = 0.77–0.24 × 10− 2 × z1, R2 = 0.76 was; if lactose supplementation is ≥ breakpoint, then z1 = 0; if lactose supplementation is < breakpoint, then z1 = lactose supplementation - breakpoint

Changes in G:F of nursery pigs fed diets by average daily lactose intake during 5 to 7 kg (A) and 7 to 11 kg (B) BW using a broken-line analysis. The meta-analysis is conducted by Proc NLMIXED in SAS 9.4 to determine break-points on the regression of feed efficiency of nursery pigs calculated based on the data from 10 published studies (Phase 1 for 5 to 7 kg BW: 14 experiments) and 14 published studies (Phase 2 for 7 to 11 kg BW: 23 experiments). The breakpoints (one-slope broken-line model) were average daily lactose intake at 45.61 g/d during 5 to 7 kg BW and 57.13 g/d during 7 to 11 kg BW when G:F were 0.79 and 0.77, respectively (P < 0.05). One-slope broken-line models; The equation for G:F during 5 to 7 kg BW was G:F = 0.79–0.12 × 10− 3 × z1, R2 = 0.89. The equation for G:F during 7 to 11 kg BW was G:F = 0.77–0.83 × 10− 4 × z1, R2 = 0.76 was; if average daily lactose intake is ≥ breakpoint, then z1 = 0; if average daily lactose intake is < breakpoint, then z1 = average daily lactose intake - breakpoint
Milk oligosaccharides
Milk oligosaccharides are the third most abundant nutrient in mammalian milk after lactose and lipids [5, 43]. Milk oligosaccharides are composed of the five monosaccharides fucose, glucose, galactose, N-acetylglucosamine, and sialic acid [59, 60]. It has also been reported that milk oligosaccharides have over 200 distinct structures with high complexity due to branches and elongated oligosaccharide structures [32]. These milk oligosaccharides contain core units having a formation of a β1–4 glycosidic linkage between galactose and glucose, and it can be further extended via β1–3 or β1–6 linkages with additional galactose or N-acetylglucosamine [61]. The core units in milk oligosaccharides could be linear or branched and it can be further linked with fucoses and/or sialic acid residues [61]. There are three major types of oligosaccharides found in milk such as neutral, neutral N-containing, and acidic oligosaccharides and classified based on the residue located at the terminal position among fucose (neutral oligosaccharides), N-acetylglucosamine (neutral N-containing oligosaccharides), and sialic acid (acidic oligosaccharides) [60].
The diversity and composition of oligosaccharides in milk are widely variable between species or period of lactation. Human milk contains the highest abundance and structural complexity, with about 20 to 25 g/L in colostrum and 5 to 20 g/L in subsequent mature milk [32]. Bode [21] described that the concentration and complexity of milk oligosaccharides in human milk have been investigated and appear to be 10 to 100 folds higher than the milk of farm animals such as cows, goats, and pigs. The concentration of bovine milk oligosaccharides are about 0.7 to 1.2 g/L in colostrum and 0.05 to 0.10 g/L in the mature milk, with about 100 identified structures, or about 15%, overlapping with human milk oligosaccharides. Accordingly, bovine milk oligosaccharides could serve as an excellent potential source for commercially available and cost-effective milk oligosaccharides in animal production compared with milk oligosaccharides from other species [60]. Dairy production and processing are equipped with sophisticated filtration systems to separate valued milk components resulting in whey permeate with high level lactose and oligosaccharides that can provide nutritional and health benefits to animal species of interest [60, 62]. Thus, nursery pigs could potentially obtain benefits from milk oligosaccharides through supplementation of whey permeate. Jang et al. [7] found that whey permeate contained about 0.4% milk oligosaccharides and showed positive effects on the intestinal health of nursery pigs when whey permeate was supplemented into their feeds. In contrast, little is known about porcine milk oligosaccharides in regards to their concentration and structural characterization in sow milk compared with human and bovine milks. There are increasing attention to the investigation of the functional properties of porcine milk oligosaccharides and their effects on the intestinal health and development of nursing piglets. Upon farrowing, intestine of piglets would initially face with lactose and milk oligosaccharides from sow milk. Previous studies have shown that the profile of oligosaccharides in porcine milk would have a higher similarity to human milk compared with bovine milk [63, 64]. This may help to define specific benefits of milk oligosaccharides to improve intestinal health and growth of nursing piglets during lactation.
Mammalian animals do not have specific enzymes to hydrolyze the milk oligosaccharides. However, milk oligosaccharides received significant attentions for their functional benefits on intestinal development and health in animals (Table 1). Milk oligosaccharides are mostly considered as prebiotics that was defined for the first time in 1995 as “a non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and/or activity of one or a limited number of bacteria in the colon, and thus improves host health” [65]. According to the International Scientific Association of Probiotics and Prebiotics (ISAPP) [66], milk oligosaccharides are also dietary prebiotics, because milk oligosaccharides are also selectively utilized by host microorganisms providing health benefits. Furthermore, milk oligosaccharides are also meeting the other criteria for prebiotics as 1) resistance to gastric pH acidity, absorption, and hydrolysis of mammalian enzymes, 2) fermentable by intestinal microflora, 3) selectively interaction with intestinal microbiota such as Lactobacillus and Bifidobacterium in the small intestine and colon [66,67,68]. Milk oligosaccharides can also directly or indirectly interact with enterocytes, leading to host protection by improving intestinal health [21]. Milk oligosaccharides can also protect the intestine from damages by increasing enterocyte proliferation [69, 70]. Interestingly, once arrived into the intestine, milk oligosaccharides would possibly bind to immune receptors on the innate immune cells or enterocytes, leading to the immune-modulatory and protective functions in the animal body [72, 73]. In addition, the small portion of milk oligosaccharides can also be absorbed into the enterocytes of infants by active transport, and it could possibly affect the systemic immune response [71]. Therefore, milk oligosaccharides provide a spectrum of protective and immune-modulatory functions mediated either by their prebiotic role in enriching specific beneficial microbiota or by direct interaction with immune cells.
Mode of action of milk carbohydrates benefiting immature intestine of nursery pigs
Previous studies indicated that milk carbohydrates played functional roles in intestinal health of nursery pigsby regulating nutrient metabolism, immune response, enterocyte proliferation, and intestinal microbiota [7, 74]. However, the functional importance of milk oligosaccharides has not been discussed thoroughly, whereas the role of lactose has been highlighted well. Therefore, the major objective of this section is to review the published data focusing on the functional effects of both lactose and milk oligosaccharides on intestinal development and health in nursery pigs.
Recently, increasing supplemental levels of milk carbohydrates in feeds improved the growth performance of nursery pigs during the early postweaning period [7, 10]. The improvement in growth of nursery pigs fed with dietary lactose could be corresponding to the change of lactase activity by growth. Ekstrom et al. [15] reported that the lactase activity in the small intestine of nursery pigs was linearly decreased by 80% from the birth to 6 weeks of age. Previous studies demonstrated that the reduction of lactase activity could be related to an increased inflammatory response in the intestinal epithelium of the small intestine after weaning [75, 76]. Pié et al. [75] showed that the lactase activity was decreased by 60 to 80% in the entire small intestine in nursery pigs for 8 d after weaning. In particular, proinflammatory cytokines such as interferon-gamma (IFN-γ), interleukin-1beta (IL-1β), and tumor necrosis factor-alpha (TNF-α) produced for immune activation could reduce the production of digestive enzymes in the intestine [77, 78]. According to Jang et al. [7], increasing supplemental levels of whey permeate from 0 to 18.75% as sources of lactose and milk oligosaccharides linearly reduced the lactase activity by 60% compared with 0 whey permeate with increased IL-8 production in the jejunum of nursery pigs during 7 to 11 kg BW. This may be related to immunomodulatory functions of milk oligosaccharides leading to immune activation by binding to the receptors on immune cells or enterocytes in the jejunum. Another possible action could be associated with the lumen pH changes in the small intestine by milk carbohydrates [43] because it has been shown that lactose supplementation induced an increased lactic acid from intestinal fermentation, reducing the luminal pH. This could lead to a favorable condition against pathogenic bacteria [16, 79, 80].
Supplementation of milk oligosaccharides can be one of the ways to improve the utilization of lactose in the intestine of nursery pigs. In the studies using a human model, supplementation of prebiotics could mitigate the adverse effects of lactose intolerance in human adults [17, 81, 82]. When the secretion and activity of lactase in the small intestine are not sufficient and cannot hydrolyze enough lactose, an undigested portion would be fermented by colonic bacteria [83]. However, when the amount of undigested lactose is over the capacity of colonic fermentation, it would increase the occurrence of diarrhea by increased osmotic trapping of water and abnormal intestinal motility, including bloating and flatulence leading to abdominal pain and diarrhea [84, 85]. Milk oligosaccharides may help increase the capacity of intestinal fermentation of lactose by modulating the commensal intestinal bacterial population increasing beneficial microorganisms [68, 86]. Previous studies have shown that supplementation of prebiotics could reduce the symptoms of lactose intolerance by adaptive shifts in intestinal microbiota [17, 82, 87]. Therefore, supplementation of milk coproducts providing both lactose and milk oligosaccharides in nursery feeds would be beneficial for the intestinal maturation and health of nursery pigs compared with the use of crystalline lactose only. According to Jang et al. [7], supplementation of whey permeate in starter feeds improved intestinal development and growth of pigs in the early post-weaning period, indicating that supplementation of whey permeate could have positive effects on preventing jejunal dysfunction in nursery pigs at 7 to 11 kg BW. It was also reported that milk oligosaccharides could prevent pathogenic invasion, facilitate the establishment of intestinal microbiota, improve intestinal development, and stimulate immune activation [74]. These observations have significantly increased research interest in understanding interactive functional roles and nutritional importance of milk carbohydrates in intestinal maturation and health of nursery pigs (Fig. 3).

Overview of the possible functions of milk carbohydrates on the intestine of nursery pigs. Lactose and various type of milk oligosaccharides could improve the intestinal maturation and health of nursery pigs through positively modulating enterocyte proliferation, intestinal immune response, and microbiota. (1) Lactose would be hydrolyzed by lactase producing galactose and lactose, which are utilized to produce energy for the growth and maintenance of nursery pigs. (2) lactose at optimal level could induce a favorable condition against pathogenic bacteria by microbial fermentation of saccharolytic bacterial groups, including Lactobacillus and Bifidobacterium, leading to preventing pathogen colonization in the intestine of nursery pigs. In addition, (3) milk oligosaccharides could modulate the growth of intestinal epithelium through binding to toll-like receptors (TLRs) and epidermal growth factor receptors (EGFR) on the enterocytes. (4) Milk oligosaccharides can increase gene expression of mucus secretion and prevent the pathogen from binding to intestinal immune-related receptors including TLRs and various types of glycan receptors, leading to enhanced mucosal immune homeostasis and tolerance. (5) Milk oligosaccharides can prevent pathogen colonization by increased abundance of Bifidobacterium and production of lactic acid and volatile fatty acids (VFA)
Enterocyte proliferation in nursery pigs
Intestinal enterocytes are intensively proliferated in crypts after birth and during the postweaning period to develop the epithelial structure and recover the epithelial structure from damages by weaning stress. Thus, crypt cell proliferation has been used as a marker showing the status of intestinal maturation, development, or maintenance of pigs. According to Jang et al. [88], the proportion of proliferating cells in a jejunal crypt of neonatal piglets was near 50%, and then the proportion was decreased by 17% on d 18 of lactation. In addition, weaning stress is shown to negatively influence physiological functions and morphology of enterocytes in nursey pigs, causing increased apoptosis, proliferation, and differentiation that are critical in recovery, development, maintenance of intestinal epithelium [89,90,91]. According to Duarte et al. [92], the oral challenge with enterotoxigenic F18+ Escherichia coli to nursery pigs increased the proliferation of enterocytes by about 20% in the jejunum.
Milk oligosaccharides have been known that it can mainly have two important potential functions: 1) enhancement of growth in beneficial bacterial communities and 2) prevention of pathogen attachment to the intestinal epithelial [93]. In addition, recent papers showed that milk oligosaccharides could beneficially modulate the enterocyte proliferation in the immature intestine of the pigs. Wang et al. [94] demonstrated that milk oligosaccharides increased enterocyte proliferation by 60 to 80% in the ileum of mice after the hypoxia exposure for 3 d. Wang et al. [69] further demonstrated that milk oligosaccharides could restore enterocyte proliferation in the ileum of mice challenged with hypoxia and cold stress. Hester et al. [95] also reported that high doses of milk oligosaccharides including fucosylated-oligosaccharides, lacto-N-enotetaose, and sialylated-oligosaccharides reduced the stimulated-cell proliferation by the incubation with nucleotides in the human intestinal epithelial cells (FHs-74 Int). In a normal condition, Holscher et al. [70] showed that incubation with milk oligosaccharides for 72 h reduced the enterocyte proliferation by about 10% in HT-29 culture and by about 25% in Caco-2Bbe cell culture. Kuntz et al. [96] also showed sialylated-oligosaccharides reduced the proliferation in HT-29 and Caco-2 cell cultures. However, there is limited information in porcine studies demonstrating the effects of milk oligosaccharides on intestinal maturation and enterocyte proliferation in the small intestine. Jang et al. [7] recently showed the increasing supplemental whey permeate as sources of lactose and milk oligosaccharides linearly increased the crypt cell proliferation in the jejunum of nursery pigs.
A possible mechanism for milk oligosaccharides improving intestinal health and development would be related to their direct binding to relevant receptors on the enterocytes. Previous studies showed that milk oligosaccharides possess the ability to bind with toll-like receptors family (TLRs) and epidermal growth factor receptors (EGFR) on the cell membrane of the enterocytes, effectively modulating TLR and EGFR signal pathways which are associated with cell proliferation [94, 97, 98]. It has been known that the various TLRs and EGFR could play a role in intestinal maturation and development including enterocyte proliferation, apoptosis, migration, and immune response in the intestine [99,100,101,102]. Following a mucosal injury from viral or bacterial invasions, stress, or inflammatory response, the damage-associated molecular patterns (DAMPs), which are signal molecules released during cell injury, could be recognized by TLRs and the various signaling pathways in the intestine, resulting in an increase of epithelial cell proliferation involved in regeneration [101]. Wang et al. [69] reported that milk oligosaccharides reduced the expression of TLR4 by about 50% and the enterocyte proliferation by about 40% in the ileum of mice challenged with hypoxia and cold stress on d 3 after the challenge. Good et al. [102] also reported that breast milk significantly could reduce the TLR4 signaling with activation of the EGFR in the enterocytes of mice treated with LPS challenge. Thus, milk oligosaccharides may inhibit pathogens from binding to these receptors, and then modulate the TLR and EGFR signals in the immature intestine, leading to an increase in proliferation.
Immune responses in the immature intestine of pigs
Oligosaccharides have been shown to exert various effects on the immune system and potentially modulate immune responses [103]. Recently, functional roles of prebiotic oligosaccharides in the intestine have been increasingly investigated [104, 105]. Among oligosaccharides, milk oligosaccharides have been shown to have functional roles influencing the development of the immune system in the immature intestine of pigs [106]. Several studies have demonstrated that the effects of milk oligosaccharides on innate immunity [107]. In vitro experiments have shown that administration of sialylated-oligosaccharides induced greater bacterial clearance in mice infected with Pseudomonas aeruginosa strain K and promoted receptor-mediated endocytosis and phagocytosis [108]. Furthermore, milk oligosaccharides could easily conjugate with various receptors and oligopeptide carriers in immune cells resulting in decreased virus-receptor interactions [109, 110]. Obelitz-Ryom et al. [111] also reported that the preterm piglets fed milk oligosaccharides had significantly improved the phagocytic activity of neutrophils isolated from blood. These data indicate that milk oligosaccharides could be a strong immunomodulatory agent in nursery pigs.
Newborn piglets would have a relatively low immune capacity with low numbers of leukocytes and immune-modulatory components [112, 113]. Piglets do not have sufficient defense ability due to undeveloped immune cells. In addition, weaning stress could potentially cause the intestinal dysfunction of nursery pigs during post-weaning period [114]. Thus, the neonates would take up to 6 weeks to develop a stable immunity in nursery pigs [114]. Therefore, at the first 2 to 4 weeks of life, pigs are susceptible to various types of stresses and diseases in which the intestinal immunity is not mature. The intestine has an important function as an immune barrier possessing up to 70% of immune cells in an animal body [115, 116]. As the first line of defense, intestinal barrier function is considered innate immunity. Firstly, a layer made of mucin and mucus glycoproteins could be a part of the intestinal barrier. It can act as a physical barrier between the lumen and intestinal epithelium. Milk oligosaccharides may affect the function of goblet cells in the intestine. According to Cheng et al. [117], milk oligosaccharides, especially, 2′-fucosyllactose, 3′-fucosyllactose, and lacto-N-triaose II, could enhance the mucus barrier function through increased the gene expression of mucus secretion including MUC2, TFF3, and RETNLB genes in the enterocytes under inflammatory and stress conditions. It is important to note that the structures of mucin glycans are similar to milk oligosaccharides. It could be further hypothesized that milk oligosaccharides modulate intestinal barrier functions by modulating the composition of intestinal microbiota [118]. Therefore, it could indicate that milk carbohydrates potentially support the intestinal barrier by promoting intestinal mucus secretion.
Secondly, milk oligosaccharides could effectively modulate the immune functions and ligand specialties through direct interaction with the various type of pattern recognition receptors such as multiple classes of lectins and TLRs in the intestinal epithelium or immune cells [33]. Previous studies have shown that milk oligosaccharides could be involved in the expression of various TLRs that could bind to the surface molecules of pathogens in the intestine. He et al. [119] showed that milk oligosaccharides reduced mRNA level of the cluster of differentiation (CD) 14 by 40% in T84 intestinal epithelial cells treated for 48 h and then challenged by LPS compared with the control group. The CD14 is engaged in TLR-4 signaling and is also a receptor enabling the recognition of the gram negative pathogenic bacteria on enterocytes under LPS-induced challenge. Milk oligosaccharides could suppress the expression of TLR4 by mediating the maturation of dendritic cells [72]. It could be supported by Zhang et al. [97], showing that milk oligosaccharides could inhibit the interaction between LPS and TLR4 by reducing pro-inflammatory cytokines including IL-6 and TNF-α in the ileum exposed to hypoxia and cold stress, leading to an increased 25% survival rate of rats.
The family of glycan receptors such as c-type lectins, galectins, selectins, and siglecs could bind to milk oligosaccharides on intestinal epithelium and immune cells [33]. It has been known that lectin receptors are glycan-binding proteins related to immune response and cell recognition [120, 121]. Xiao et al. [72] showed that milk oligosaccharides could mediate the interaction with dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) receptors that are c-type lectins involved in the regulation of the immune response in dendritic cells. In contrast to c-type lectins, milk oligosaccharides can also directly affect the interaction of galectins, which are receptors on T cells or enterocytes, leading to immune regulation of T cell functions [122]. He et al. [123] also reported that milk oligosaccharides increased the T-helper 1 (Th1) cells polarization with improvements in the balance of Th1 / Th2 lymphocytes in the intestinal mucosa, indicating that milk oligosaccharides may induce the maturation of immune response through enhancing the cell-mediated immune response to pathogen infection in the intestine. Nursery pigs are susceptible to intestinal inflammation caused by stress, diseases, or viral infections due to the immature development and maturation in their intestine [124, 125]. According to previous studies, milk oligosaccharides could mediate the immune receptors on immune cells and enterocytes under stressful conditions by reducing the expressions of TLRs and lectin receptors [33, 126]. Therefore, milk carbohydrates could positively support the intestinal immune response of nursery pigs by interacting with immune receptors on the intestinal epithelium.
Microbiota in the immature intestine of nursery pigs
The composition and diversity of intestinal microbiota continue to change as a pig grows [127, 128], which can also be affected by environmental factors and dietary factors [129]. Previous studies showed that Lactobacillus and Bifidobacterium generally dominate the intestine of breast-fed animals [130,131,132]. Interestingly, the microbial fermentation by the saccharolytic bacterial groups, including Lactobacillus and Bifidobacterium, in the intestine would induce the production of short-chain fatty acids, which could affect the intestinal environment favorable to microorganisms reducing luminal pH [80, 133]. According to Gahan et al. [43], increasing the supplemental levels of lactose from 60 to 250 g/kg in feeds linearly decreased the fecal pH of nursery pigs by 11% on d 15 after weaning. The reduced local luminal pH could help inhibit the colonization of pathogens living in the intestinal lumen, feces, or mucosa [134, 135]. Zhao et al. [58] also reviewed that dietary lactose could be fermented by intestinal microbiota resulting in the production of lactic acid and short-chain fatty acids, conferring prebiotic-like effects on intestinal microbiota. Previous studies have also shown that supplementation of dietary lactose resulted in modulation of the abundance of specific microflora, including Lactobacillus [7, 136], E. coli [45], and Bifidobacterium [137].
Frese et al. [138] also showed that milk oligosaccharides could help establish the intestinal microbiota in nursery pigs. Jang et al. [7] reported that increasing the supplemental levels of whey permeate from 0 to 18.7% increased the abundance of Bifidobacteriaceae by 41% and showed the quadratic effects on the abundance Lactobacillaceae showing increasing by 31% at 13% whey permeate in the jejunum of nursery pigs during 7 to 11 kg BW. It could indicate that milk carbohydrates could positively modulate the mucosa-associated microbiota of nursery pigs. In addition, these saccharolytic bacterial groups could effectively utilize milk carbohydrates, resulting in the production of abundant short-chain fatty acids including butyrate and lactate mediated by Bifidobacterium and Bacteroides groups [139, 140]. Short-chain fatty acids have been shown to be essential energy sources for intestinal development and effective immune responses [130, 131]. Specific enzymes such as fucosidase and neuraminidase that hydrolyze milk oligosaccharides have been found in saccharolytic bacteria and these enzymes have specific functions in microbial growth and short-chain fatty acid production [140, 141]. However, so far, information on these potential beneficial impacts of milk oligosaccharides on intestinal fermentation has been elucidated through clinical trials mainly focusing on the colon in humans or in vitro studies for pigs. Pigs could effectively utilize the milk oligosaccharides for intestinal microbial fermentation due to enormous differences in the relative size of the intestine compared with human. Pigs have well-developed cecum where intestinal microbiota would ferment a part of carbohydrates before entering the colon and their cecum is significantly larger than humans [142]. It has also been known that the contribution of short-chain fatty acids to maintenance energy requirement through microbial fermentation is about 10% in humans and 20% in pigs [143, 144]. Therefore, milk oligosaccharides could display great potentials to be effectively for the production of short-chain fatty acids by microbial fermentation in the intestine of nursery pigs.
In addition, milk oligosaccharides could provide antiadhesive effects on intestinal pathogens, which use cell surface glycans to recognize and bind to the target cells, leading to pathogenesis [145, 146]. This may be due to milk oligosaccharides possessing structural similarities to cell surface glycoconjugates utilized by intestinal microbiota [145]. Additionally, it has also been known that oligosaccharides can be involved in cell recognition and signaling, specifically in microbial adhesion and microbial interactions between the intestinal epithelial cells [147]. According to previous studies, milk oligosaccharides could not only inhibit binding of the pathogens such as E. coli [148,149,150], Helicobacter [151], and Salmonella [148], and microbial enterotoxins [152], but also significantly promote the adhesion of Bifidobacteria to intestinal epithelial cells [153, 154]. Walsh et al. [68] demonstrated that milk oligosaccharides-containing products could potentially increase the residence time of probiotic bacteria in the intestine leading to improved or sustained intestinal health.
Conclusion
Milk carbohydrates are essential to support the growth and intestinal health of the pigs, especially during early stage of life when the pigs are vulnerable from enteric diseases or stressful conditions negatively affecting the intestinal health. Although lactase activity is high in their early life, it is important that feeding proper amounts of lactose in early weaner feeds because if feed too high, it can cause abnormal intestinal functions from extensive lactose fermentation, unbalanced osmosis, or abnormal motility. Based on the meta-analysis of published data, it would be recommended that pigs at 5 to 7 kg BW (typical early weaner feeds) need 20% lactose (or 45.6 g/d intake), pigs at 7 to 11 kg BW (typical pre-starter feeds) need 13% lactose (or 57.1 g/d intake). Milk carbohydrates as bioactive compounds play a critical roles in improving intestinal maturation and health of nursery pigs through positively modulating enterocyte proliferation, immune response, and microbiota. Milk oligosaccharides promote the utilization of lactose by positive modulation of immune responses and composition of intestinal microbiota in young pigs. Based on the current knowledge and research, it is warranted to further investigate whether milk carbohydrates can have specificity of prebiotic effects depending on the types, ratio, or combination status.
Availability of data and materials
All data generated or analyzed during this study are available from the corresponding author upon reasonable request.
Abbreviations
- AA:
-
Amino acids
- ATP:
-
Adenosine triphosphate
- BW:
-
Body weight
- CD:
-
Cluster of differentiation
- DAMPs:
-
Damage-associated molecular patterns
- DC-SIGN:
-
Dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin
- DM:
-
Dry matter
- EGFR:
-
Epidermal growth factor receptors
- GIT:
-
Gastrointestinal tract
- GLUT2:
-
Glucose transporter 2
- HMO:
-
Human milk oligosaccharides
- BMO:
-
Bovine milk oligosaccharides
- IFN-γ:
-
Interferon-gamma
- IL:
-
Interleukin
- ISAPP:
-
International scientific association of probiotics and prebiotics
- LPS:
-
Lipopolysaccharides
- SGLT1:
-
Na+- D-glucose cotransporter 1
- Th:
-
T-helper cells
- TLR:
-
Toll-like receptors
- TNF-α:
-
Tumor necrosis factor-alpha
References
Holanda DM, Yiannikouris A, Kim SW. Investigation of the efficacy of a postbiotic yeast cell wall-based blend on newly-weaned pigs under a dietary challenge of multiple mycotoxins with emphasis on deoxynivalenol. Toxins. 2020;12:504.
Liu Y, Espinosa CD, Abelilla JJ, Casas GA, Lagos LV, Lee SA, et al. Non-antibiotic feed additives in diets for pigs: a review. Anim Nutr. 2018;4:113–25.
Celi P, Cowieson AJ, Fru-Nji F, Steinert RE, Kluenter A-M, Verlhac V. Gastrointestinal functionality in animal nutrition and health: new opportunities for sustainable animal production. Anim Feed Sci Technol. 2017;234:88–100.
Taliercio E, Kim SW. Identification of a second major antigenic epitope in the α-subunit of soy β-conglycinin. Food Agric Immunol. 2014;25:311–21.
Taliercio E, Kim SW. Epitopes from two soybean glycinin subunits are antigenic in pigs. J Sci Food Agric. 2013;93(12):2927–32. https://doi.org/10.1002/jsfa.6113.
Tokach MD, Pettigrew JE, Johnston LJ, Overland M, Rust JW, Cornelius SG. Effect of adding fat and (or) milk products to the weanling pig diet on performance in the nursery and subsequent grow-finish stages. J Anim Sci. 1995;73:3358–68.
Jang KB, Purvis JM, Kim SW. Dose–response and functional role of whey permeate as a source of lactose and milk oligosaccharides on intestinal health and growth of nursery pigs. J Anim Sci. 2021;99:1–13.
Mahan DC, Fastinger ND, Peters JC. Effects of diet complexity and dietary lactose levels during three starter phases on postweaning pig performance. J Anim Sci. 2004;82:2790–7.
Kim SW. Chapter 28 - Sow milk. In: Park YW, GFW H, editors. Milk and dairy products in human nutrition production, composition, and health. Chichester, West Sussex, UK: John Wiley and Sons; 2013. p. 614–26.
Cromwell GL, Allee GL, Mahan DC. Assessment of lactose level in the mid- to late-nursery phase on performance of weanling pigs. J Anim Sci. 2008;86:127–33.
Lin C, Mahan DC, Wu GY, Kim SW. Protein digestibility of porcine colostrum by neonatal pigs. Livest Sci. 2009;121:182–6.
Nessmith WB Jr, Nelssen JL, Tokach MD, Goodband RD, Bergström JR, Dritz SS, et al. Evaluation of the interrelationships among lactose and protein sources in diets for segregated early-weaned pigs. J Anim Sci. 1997;75:3214–21.
Pierce KM, Callan JJ, McCarthy P, O’Doherty JV. The interaction between lactose level and crude protein concentration on piglet post-weaning performance, nitrogen metabolism, selected faecal microbial populations and faecal volatile fatty acid concentrations. Anim Feed Sci Technol. 2007;132:267–82.
Xu RJ, Sangild PT, Zhang YQ, Zhang SH. Chapter 5 - Bioactive compounds in porcine colostrum and milk and their effects on intestinal development in neonatal pigs. In: Zabielski R, Gregory PC, Westöm B, editors. Biology of the intestine in growing animals. Amsterdam, NL: Elsevier; 2002. p. 169–92.
Ekstrom KE, Benevenga NJ, Grummer RH. Changes in the intestinal lactase activity in the small intestine of two breeds of swine from birth to 6 weeks of age. J Nutr. 1975;105:1032–8.
Misselwitz B, Butter M, Verbeke K, Fox MR. Update on lactose malabsorption and intolerance: pathogenesis, diagnosis and clinical management. Gut. 2019;68:2080–91.
Savaiano DA, Ritter AJ, Klaenhammer TR, James GM, Longcore AT, Chandler JR, et al. Improving lactose digestion and symptoms of lactose intolerance with a novel galacto-oligosaccharide (RP-G28): a randomized, double-blind clinical trial. Nutr J. 2013;12(1):160. https://doi.org/10.1186/1475-2891-12-160.
Montalto M, Curigliano V, Santoro L, Vastola M, Cammarota G, Manna R, et al. Management and treatment of lactose malabsorption. World J Gastroenterol. 2006;12(2):187–91. https://doi.org/10.3748/wjg.v12.i2.187.
Molino JP, Donzele JL, de Oliveira RFM, Ferreira AS, de Moraes CA, Haese D, et al. Lactose levels in diets for piglets weaned at 21 days of age. Rev Bras Zootec. 2011;40:1233–41.
Bering SB. Human milk oligosaccharides to prevent gut dysfunction and necrotizing enterocolitis in preterm neonates. Nutrients. 2018;10(10):1461.
Bode L. Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology. 2012;22(9):1147–62.
Ramani S, Stewart CJ, Laucirica DR, Ajami NJ, Robertson B, Autran CA, et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nat Commun. 2018;9:5010.
Moukarzel S, Bode L. Human milk oligosaccharides and the preterm infant: a journey in sickness and in health. Clin Perinatol. 2017;44(1):193–207. https://doi.org/10.1016/j.clp.2016.11.014.
Lönnerdal B, Hernell O. An opinion on “staging” of infant formula: a developmental perspective on infant feeding. J Pediatr Gastroenterol Nutr. 2016;62:9–21.
Zuurveld M, van Witzenburg NP, Garssen J, Folkerts G, Stahl B, van’t Land B, et al. Immunomodulation by human milk oligosaccharides: the potential role in prevention of allergic diseases. Front Immunol. 2020;11:801.
Djouzi Z, Andrieux C. Compared effects of three oligosaccharides on metabolism of intestinal microflora in rats inoculated with a human faecal flora. Br J Nutr. 1997;78:313–24.
Moro G, Minoli I, Mosca M, Fanaro S, Jelinek J, Stahl B, et al. Dosage-related bifidogenic effects of galacto- and fructooligosaccharides in formula-fed term infants. J Pediatr Gastroenterol Nutr. 2002;34(3):291–5. https://doi.org/10.1097/00005176-200203000-00014.
Baker JT, Duarte ME, Holanda DM, Kim SW. Friend or foe? Impacts of dietary xylans, xylooligosaccharides, and xylanases on intestinal health and growth performance of monogastric animals. Animals. 2021;11(3):1–28. https://doi.org/10.3390/ani11030609.
Garrido D, Dallas DC, Mills DA. Consumption of human milk glycoconjugates by infant-associated bifidobacteria: mechanisms and implications. Microbiol. 2013;159:649–64.
Yu Z-T, Chen C, Newburg DS. Utilization of major fucosylated and sialylated human milk oligosaccharides by isolated human gut microbes. Glycobiology. 2013;23(11):1281–92. https://doi.org/10.1093/glycob/cwt065.
Yu ZT, Chen C, Kling DE, Liu B, McCoy JM, Merighi M, et al. The principal fucosylated oligosaccharides of human milk exhibit prebiotic properties on cultured infant microbiota. Glycobiology. 2013;23:169–77.
Wiciński M, Sawicka E, Gębalski J, Kubiak K, Malinowski B. Human milk oligosaccharides: Health benefits, potential applications in infant formulas, and pharmacology. Nutrients. 2020;12(1):266.
Triantis V, Bode L, van Neerven JRJ. Immunological effects of human milk oligosaccharides. Front Pediatr. 2018;6:190.
Donovan SM, Comstock SS. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann Nutr Metab. 2016;69:41–51.
Koepsell H. Glucose transporters in the small intestine in health and disease. Pflügers Arch. 2020;472:1207–48.
Leloir LF. The enzymatic transformation of uridine diphosphate glucose into a galactose derivative. Arch Biochem Biophys. 1951;33:186–90.
Jeyapalan AS, Orellana RA, Suryawan A, O’Connor PMJ, Nguyen HV, Escobar J, et al. Glucose stimulates protein synthesis in skeletal muscle of neonatal pigs through an AMPK- and mTOR-independent process. Am J Physiol Metab. 2007;293:E595–603.
Tugnoli B, Giovagnoni G, Piva A, Grilli E. From acidifiers to intestinal health enhancers: how organic acids can improve growth efficiency of pigs. Anim. 2020;10(1):134.
Navarro DMDL, Abelilla JJ, Stein HH. Structures and characteristics of carbohydrates in diets fed to pigs: a review. J Anim Sci Biotechnol. 2019;10:39.
Atkinson RL, Kratzer FH, Stewart GF. Lactose in animal and human feeding: a review. J Dairy Sci. 1957;40:1114–32.
Theil PK, Lauridsen C, Quesnel H. Neonatal piglet survival: impact of sow nutrition around parturition on fetal glycogen deposition and production and composition of colostrum and transient milk. Animal. 2014;8:1021–30.
Redel CA, Shulman RJ, Tivey DR. Determinants of lactose digestion in the miniature pig. Dig Dis Sci. 1997;42:137–44.
Gahan DA, Lynch MB, Callan JJ, O’Sullivan JT, O’Doherty JV. Performance of weanling piglets offered low-, medium- or high-lactose diets supplemented with a seaweed extract from Laminaria spp. Animal. 2009;3:24–31.
Naranjo VD, Bidner TD, Southern LL. Comparison of dried whey permeate and a carbohydrate product in diets for nursery pigs. J Anim Sci. 2010;88(5):1868–79. https://doi.org/10.2527/jas.2009-2438.
O’Doherty JV, Dillon S, Figat S, Callan JJ, Sweeney T. The effects of lactose inclusion and seaweed extract derived from Laminaria spp. on performance, digestibility of diet components and microbial populations in newly weaned pigs. Anim Feed Sci Technol. 2010;157:173–80.
Kim JS, Shinde PL, Yang YX, Yun K, Choi JY, Lohakare JD, et al. Effects of dietary lactose levels during different starter phases on the performance of weaning pigs. Livest Sci. 2010;131:175–82.
Lynch MB, Callan JJ, O’Doherty JV. The interaction between lactose level and enzyme supplementation and form of barley processing on performance, digestibility and faecal volatile fatty acid concentration of weanling pigs fed barley-based diets. Anim Feed Sci Technol. 2008;140:349–64.
Tran H, Moreno R, Hinkle EE, Bundy JW, Walter J, Burkey TE, et al. Effects of lactose and yeast-dried milk on growth performance, fecal microbiota, and immune parameters of nursery pigs. J Anim Sci. 2012;90:3049–59.
Mahan DC. Efficacy of dried whey and its lactalbumin and lactose comnonents at two dietarv lysine levels on postweaning pig performance and nitrogen balance1. J Anim Sci. 1992;70(7):2182–7. https://doi.org/10.2527/1992.7072182x.
Molino JP, Donzele JL, Flávia R, De Oliveira M, Haese D, Fortes EI, et al. L-glutamine and L-glutamate in diets with different lactose levels for piglets weaned at 21 days of age. Rev Bras Zootec. 2012;41(1):98–105.
Jang K, Purvis JM, Kim SW. 365 effects of weaning ages on growth performance of nursery pigs fed diets with increasing lactose levels during at 7 to 11 kg body weight. J Anim Sci. 2019;97:130.
Grinstead GS, Goodband RD, Dritz SS, Tokach MD, Nelssen JL, Woodworth JC, et al. Effects of a whey protein product and spray-dried animal plasma on growth performance of weanling pigs. J Anim Sci. 2000;78(3):647–57. https://doi.org/10.2527/2000.783647x.
Graham PL, Mahan DC, Shields RG. Effect of starter diet and length of feeding regimen on performance and digestive enzyme activity of 2-week old weaned pigs. J Anim Sci. 1981;53:299–307.
Tokach MD, Nelssen JL, Allee GL. Effect of protein and (or) carbohydrate fractions of dried whey on performance and nutrient digestibility of early weaned pigs. J Anim Sci. 1989;67:1307–12.
Mahan DC. Evaluating two sources of dried whey and the effects of replacing the corn and dried whey component with corn gluten meal and lactose in the diets of weanling swine. J Anim Sci. 1993;71:2860–6.
Shen YB, Weaver AC, Kim SW. Effect of feed grade L-methionine on growth performance and gut health in nursery pigs compared with conventional DL-methionine. J Anim Sci. 2014;92:5530–9.
Robbins KR, Saxton AM, Southern LL. Estimation of nutrient requirements using broken-line regression analysis. J Anim Sci. 2006;84(Suppl):E155–65.
Zhao J, Zhang Z, Zhang S, Page G, Jaworski NW. The role of lactose in weanling pig nutrition: a literature and meta-analysis review. J Anim Sci Biotechnol. 2021;12:10.
Urashima T, Taufik E, Fukuda K, Asakuma S. Recent advances in studies on milk oligosaccharides of cows and other domestic farm animals. Biosci Biotechnol Biochem. 2013;77:455–66.
Zivkovic AM, Barile D. Bovine milk as a source of functional oligosaccharides for improving human health. Adv Nutr. 2011;2(3):284–9.
Ayechu-Muruzabal V, van Stigt AH, Mank M, Willemsen LEM, Stahl B, Garssen J, et al. Diversity of human milk oligosaccharides and effects on early life immune development. Front Pediatr. 2018;6:239. https://doi.org/10.3389/fped.2018.00239.
Barile D, Tao N, Lebrilla CB, Coisson J-D, Arlorio M, German JB. Permeate from cheese whey ultrafiltration is a source of milk oligosaccharides. Int Dairy J. 2009;19:524–30.
Mudd AT, Salcedo J, Alexander LS, Johnson SK, Getty CM, Chichlowski M, et al. Porcine milk oligosaccharides and sialic acid concentrations vary throughout lactation. Front Nutr. 2016;3(39):1–10.
Wei J, Wang ZA, Wang B, Jahan M, Wang Z, Wynn PC, et al. Characterization of porcine milk oligosaccharides over lactation between primiparous and multiparous female pigs. Sci Rep. 2018;8:1–16.
Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. 1995;125:1401–12.
Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, et al. Expert consensus document: the international scientific association for probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017;14:491–502.
Marcobal A, Barboza M, Froehlich JW, Block DE, German JB, Lebrilla CB, et al. Consumption of human milk oligosaccharides by gut-related microbes. J Agric Food Chem. 2010;58(9):5334–40. https://doi.org/10.1021/jf9044205.
Walsh C, Lane JA, van Sinderen D, Hickey RM. Human milk oligosaccharides: shaping the infant gut microbiota and supporting health. J Funct Foods. 2020;72:104074.
Wang C, Zhang M, Guo H, Yan J, Liu F, Chen J, et al. Human milk oligosaccharides protect against necrotizing enterocolitis by inhibiting intestinal damage via increasing the proliferation of crypt cells. Mol Nutr Food Res. 2019;63:e1900262.
Holscher HD, Davis SR, Tappenden KA. Human milk oligosaccharides influence maturation of human intestinal Caco-2Bbe and HT-29 cell lines. J Nutr. 2014;144(5):586–91. https://doi.org/10.3945/jn.113.189704.
Rudloff S, Pohlentz G, Borsch C, Lentze MJ, Kunz C. Urinary excretion of in vivo 13C-labelled milk oligosaccharides in breastfed infants. Br J Nutr England. 2012;107:957–63.
Xiao L, van De Worp WRPH, Stassen R, van Maastrigt C, Kettelarij N, Stahl B, et al. Human milk oligosaccharides promote immune tolerance via direct interactions with human dendritic cells. Eur J Immunol. 2019;49:1001–14.
Noll AJ, Yu Y, Lasanajak Y, Duska-McEwen G, Buck RH, Smith DF, et al. Human DC-SIGN binds specific human milk glycans. Biochem J. 2016;473:1343–53.
Donovan SM, Comstock SS. Human milk oligosaccharides influence neonatal mucosal and systemic immunity. Ann Nutr Metab. 2017;69:42–51.
Pié S, Lallès JP, Blazy F, Laffitte J, Sève B, Oswald IP. Weaning is associated with an upregulation of expression of inflammatory cytokines in the intestine of piglets. J Nutr. 2004;134(3):641–7. https://doi.org/10.1093/jn/134.3.641.
Solaymani-Mohammadi S, Singer SM. Host immunity and pathogen strain contribute to intestinal disaccharidase impairment following gut infection. J Immunol. 2011;187(7):3769–75. https://doi.org/10.4049/jimmunol.1100606.
Kany S, Vollrath JT, Relja B. Cytokines in inflammatory disease. Int J Mol Sci. 2019;20(23):6008.
Ziambaras T, Rubin DC, Perlmutter DH. Regulation of sucrase-isomaltase gene expression in human intestinal epithelial cells by inflammatory cytokines. J Biol Chem. 1996;271:1237–42.
Kim CH. Immune regulation by microbiome metabolites. Immunology. 2018;154:220–9.
Fukuda S, Toh H, Hase K, Oshima K, Nakanishi Y, Yoshimura K, et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature. 2011;469:543–7.
Hertzler SR, Savaiano DA. Colonic adaptation to daily lactose feeding in lactose maldigesters reduces lactose intolerance. Am J Clin Nutr. 1996;64:232–6.
Azcarate-Peril MA, Ritter AJ, Savaiano D, Monteagudo-Mera A, Anderson C, Magness ST, et al. Impact of short-chain galactooligosaccharides on the gut microbiome of lactose-intolerant individuals. Proc Natl Acad Sci. 2017;114:E367–75.
Solomons NW. Fermentation, fermented foods and lactose intolerance. Eur J Clin Nutr. 2002;56:50–5.
Venema K. Intestinal fermentation of lactose and prebiotic lactose derivatives, including human milk oligosaccharides. Int Dairy J. 2012;22:123–40.
Deng Y, Misselwitz B, Dai N, Fox M. Lactose intolerance in adults: biological mechanism and dietary management. Nutrients. 2015;7:8020–35.
Marcobal A, Sonnenburg JL. Human milk oligosaccharide consumption by intestinal microbiota. Clin Microbiol Infect. 2012;18:12–5.
Leis R, de Castro M-J, de Lamas C, Picáns R, Couce ML. Effects of prebiotic and probiotic supplementation on lactase deficiency and lactose intolerance: a systematic review of controlled trials. Nutrients. 2020;12:1487.
Jang KB, Kim JH, Purvis JM, Chen J, Ren P, Vazquez-Anon M, et al. Effects of mineral methionine hydroxy analog chelate in sow diets on epigenetic modification and growth of progeny. J Anim Sci. 2020;98(9):1–12. https://doi.org/10.1093/jas/skaa271.
Jang KB, Kim SW. Supplemental effects of dietary nucleotides on intestinal health and growth performance of newly weaned pigs. J Anim Sci. 2019;97:4875–82.
Duarte ME, Zhou FX, Dutra WM, Kim SW. Dietary supplementation of xylanase and protease on growth performance, digesta viscosity, nutrient digestibility, immune and oxidative stress status, and gut health of newly weaned pigs. Anim Nutr. 2019;5(4):351–8.
Wang L, Yan S, Li J, Li Y, Ding X, Yin J, et al. Rapid communication: the relationship of enterocyte proliferation with intestinal morphology and nutrient digestibility in weaning piglets. J Anim Sci. 2019;97:353–8.
Duarte ME, Tyus J, Kim SW. Synbiotic effects of enzyme and probiotics on intestinal health and growth of newly weaned pigs challenged with enterotoxigenic F18+ Escherichia coli. Front Vet Sci. 2020;7:573.
Newburg DS, Ruiz-Palacios GM, Morrow AL. Human milk glycans protect infants against enteric pathogens. Annu Rev Nutr. 2005;25:37–58.
Wang C, Zhang M, Guo H, Yan J, Chen L, Teng W, et al. Human milk oligosaccharides activate epidermal growth factor receptor and protect against hypoxia-induced injuries in the mouse intestinal epithelium and Caco2 cells. J Nutr. 2020;150:756–62.
Hester SN, Donovan SM. Individual and combined effects of nucleotides and human milk oligosaccharides on proliferation, apoptosis and necrosis in a human fetal intestinal cell line. Food Nutr Sci. 2012;03:1567–76.
Kuntz S, Rudloff S, Kunz C. Oligosaccharides from human milk influence growth-related characteristics of intestinally transformed and non-transformed intestinal cells. Br J Nutr. 2008;99:462–71.
Zhang W, He-Yang J, Tu W, Zhou X. Sialylated human milk oligosaccharides prevent intestinal inflammation by inhibiting toll like receptor 4/NLRP3 inflammasome pathway in necrotizing enterocolitis rats. Nutr Metab. 2021;18:5.
Lu P, Sodhi CP, Hackam DJ. Toll-like receptor regulation of intestinal development and inflammation in the pathogenesis of necrotizing enterocolitis. Pathophysiol Off J Int Soc Pathophysiol. 2014;21(1):81–93. https://doi.org/10.1016/j.pathophys.2013.11.007.
Abreu MT. Toll-like receptor signalling in the intestinal epithelium: how bacterial recognition shapes intestinal function. Nat Rev Immunol. 2010;10:131–44.
Artis D. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol. 2008;8(6):411–20. https://doi.org/10.1038/nri2316.
He Y, Lawlor NT, Newburg DS. Human milk components modulate toll-like receptor-mediated inflammation. Adv Nutr. 2016;7:102–11.
Good M, Sodhi CP, Egan CE, Afrazi A, Jia H, Yamaguchi Y, et al. Breast milk protects against the development of necrotizing enterocolitis through inhibition of toll-like receptor 4 in the intestinal epithelium via activation of the epidermal growth factor receptor. Mucosal Immunol. 2015;8:1166–79.
Jeurink PV, Van Esch BCAM, Rijnierse A, Garssen J, Knippels LMJ. Mechanisms underlying immune effects of dietary oligosaccharides. Am J Clin Nutr. 2013;98:572–7.
Azagra-Boronat I, Massot-Cladera M, Mayneris-Perxachs J, Knipping K, van’t Land B, Tims S, et al. Immunomodulatory and prebiotic effects of 2′-Fucosyllactose in suckling rats. Front Immunol. 2019;10:1773.
Tsai C-C, Lin C-R, Tsai H-Y, Chen C-J, Li W-T, Yu H-M, et al. The immunologically active oligosaccharides isolated from wheatgrass modulate monocytes via toll-like receptor-2 signaling. J Biol Chem. 2013;288:17689–97.
Plaza-Díaz J, Fontana L, Gil A. Human milk oligosaccharides and immune system development. Nutrients. 2018;10(8):1038.
Kulinich A, Liu L. Human milk oligosaccharides: the role in the fine-tuning of innate immune responses. Carbohydr Res. 2016;432:62–70. https://doi.org/10.1016/j.carres.2016.07.009.
Kim J, Kim Y-J, Kim JW. Bacterial clearance is enhanced by α2,3- and α2,6-sialyllactose via receptor-mediated endocytosis and phagocytosis. Infect Immun. 2019;87:e00694–18.
Zevgiti S, Zabala JG, Darji A, Dietrich U, Panou-Pomonis E, Sakarellos-Daitsiotis M. Sialic acid and sialyl-lactose glyco-conjugates: design, synthesis and binding assays to lectins and swine influenza H1N1 virus. J Pept Sci. 2012;18:52–8.
Kwon S-J, Na DH, Kwak JH, Douaisi M, Zhang F, Park EJ, et al. Nanostructured glycan architecture is important in the inhibition of influenza a virus infection. Nat Nanotechnol. 2017;12:48–54.
Obelitz-Ryom K, Rendboe AK, Nguyen DN, Rudloff S, Brandt AB, Nielsen DS, et al. Bovine milk oligosaccharides with sialyllactose for preterm piglets. Nutrients. 2018;11(6):1335.
Upham JW, Rate A, Rowe J, Kusel M, Sly PD, Holt PG. Dendritic cell immaturity during infancy restricts the capacity to express vaccine-specific T-cell memory. Infect Immun. 2006;74:1106–12.
Nguyen M, Leuridan E, Zhang T, De Wit D, Willems F, Van Damme P, et al. Acquisition of adult-like TLR4 and TLR9 responses during the first year of life. PLoS ONE. 2010;5:e10407.
Moeser AJ, Pohl CS, Rajput M. Weaning stress and gastrointestinal barrier development: implications for lifelong gut health in pigs. Anim Nutr. 2017;3(4):313–21. https://doi.org/10.1016/j.aninu.2017.06.003.
Castro GA, Arntzen CJ. Immunophysiology of the gut: a research frontier for integrative studies of the common mucosal immune system. Am J Physiol Liver Physiol. 1993;265:599–610.
Furness JB, Kunze WAA, Clerc N II. The intestine as a sensory organ: neural, endocrine, and immune responses. Am J Physiol Liver Physiol. 1999;277(5):G922–8. https://doi.org/10.1152/ajpgi.1999.277.5.G922.
Cheng L, Kong C, Walvoort MTC, Faas MM, de Vos P. Human milk oligosaccharides differently modulate goblet cells under homeostatic, proinflammatory conditions and ER stress. Mol Nutr Food Res. 2020;64:1900976.
Šuligoj T, Vigsnæs LK, P Van den A, Apostolou A, Karalis K, Savva GM, et al. Effects of human milk oligosaccharides on the adult gut microbiota and barrier function. Nutrients. 2020;12:2808.
He YY, Liu SB, Kling DE, Leone S, Lawlor NT, Huang Y, et al. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut. 2016;65:33–46.
Schnaar RL. Glycans and glycan-binding proteins in immune regulation: a concise introduction to glycobiology for the allergist. J Allergy Clin Immunol. 2015;135(3):609–15. https://doi.org/10.1016/j.jaci.2014.10.057.
van Kooyk Y, Rabinovich GA. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat Immunol. 2008;9(6):593–601. https://doi.org/10.1038/ni.f.203.
Noll AJ, Gourdine J-P, Yu Y, Lasanajak Y, Smith DF, Cummings RD. Galectins are human milk glycan receptors. Glycobiology. 2016;26(6):655–69. https://doi.org/10.1093/glycob/cww002.
He Y, Liu S, Leone S, Newburg DS. Human colostrum oligosaccharides modulate major immunologic pathways of immature human intestine. Mucosal Immunol. 2014;7:1326–39.
Zheng L, Duarte ME, Sevarolli Loftus A, Kim SW. Intestinal health of pigs upon weaning: challenges and nutritional intervention. Front Vet Sci. 2021;8:91. https://doi.org/10.3389/fvets.2021.628258.
Xiong X, Tan B, Song M, Ji P, Kim K, Yin Y, et al. Nutritional intervention for the intestinal development and health of weaned pigs. Front Vet Sci. 2019;6:1–14.
Rousseaux A, Brosseau C, Le Gall S, Piloquet H, Barbarot S, Bodinier M. Human milk oligosaccharides: their effects on the host and their potential as therapeutic agents. Front Immunol. 2021;12:680911.
Jiang L, Feng C, Tao S, Li N, Zuo B, Han D, et al. Maternal imprinting of the neonatal microbiota colonization in intrauterine growth restricted piglets: a review. J Anim Sci Biotechnol. 2019;10(1):88. https://doi.org/10.1186/s40104-019-0397-7.
Niu Q, Li P, Hao S, Zhang Y, Kim SW, Li H, et al. Dynamic distribution of the gut microbiota and the relationship with apparent crude fiber digestibility and growth stages in pigs. Sci rep. 2015;5:9938. Available from. https://doi.org/10.1038/srep09938.
Hasan N, Yang H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ. 2019;7:e7502.
Fouhy F, Ross RP, Fitzgerald GF, Stanton C, Cotter PD. Composition of the early intestinal microbiota: knowledge, knowledge gaps and the use of high-throughput sequencing to address these gaps. Gut Microbe. 2012;3:203–20.
Flint HJ, Scott KP, Duncan SH, Louis P, Forano E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes. 2012;3:289–306.
Duarte ME, Kim SW. Intestinal microbiota and its interaction to intestinal health in nursery pigs. Anim Nutr. 2021; in press. J pre-proof.
Thomson P, Medina DA, Garrido D. Human milk oligosaccharides and infant gut bifidobacteria: molecular strategies for their utilization. Food Microbiol. 2018;75:37–46. https://doi.org/10.1016/j.fm.2017.09.001.
Lawley TD, Walker AW. Intestinal colonization resistance. Immunology. 2013;138(1):1–11. https://doi.org/10.1111/j.1365-2567.2012.03616.x.
Matsuki T, Yahagi K, Mori H, Matsumoto H, Hara T, Tajima S, et al. A key genetic factor for fucosyllactose utilization affects infant gut microbiota development. Nat Commun. 2016;7:11939.
Konstantinov SR, Awati A, Smidt H, Williams BA, Akkermans ADL, de Vos WM. Specific response of a novel and abundant Lactobacillus amylovorus-like phylotype to dietary prebiotics in the guts of weaning piglets. Appl Environ Microbiol. 2004;70:3821–30.
Jiang T, Mustapha A, Savaiano DA. Improvement of lactose digestion in humans by ingestion of unfermented milk containing Bifidobacterium longum. J Dairy Sci. 1996;79:750–7.
Frese SA, Parker K, Calvert CC, Mills DA. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome. 2015;3:1–10.
Sela DA, Garrido D, Lerno L, Wu S, Tan K, Eom H-J, et al. Bifidobacterium longum subsp. infantis ATCC 15697 α-Fucosidases are active on fucosylated human milk oligosaccharides. Appl Environ Microbiol. 2012;78:795–803.
Kitaoka M. Bifidobacterial enzymes involved in the metabolism of human milk oligosaccharides. Adv Nutr. 2012;3(3):422S–9S. https://doi.org/10.3945/an.111.001420.
Lawson MAE, O’Neill IJ, Kujawska M, Gowrinadh Javvadi S, Wijeyesekera A, Flegg Z, et al. Breast milk-derived human milk oligosaccharides promote Bifidobacterium interactions within a single ecosystem. ISME J. 2020;14(2):635–48. https://doi.org/10.1038/s41396-019-0553-2.
Kararli TT. Comparison of the gastrointestinal anatomy, physiology, and biochemistry of humans and commonly used laboratory animals. Biopharm Drug Dispos. 1995;16(5):351–80. https://doi.org/10.1002/bdd.2510160502.
Jha R, Fouhse JM, Tiwari UP, Li L, Willing BP. Dietary fiber and intestinal health of monogastric animals. Front Vet Sci. 2019;6:48.
Bergman EN. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol Rev. 1990;70:567–90.
Kunz C, Rudloff S. Biological functions of oligosaccharides in human milk. Acta Paediatr Norway. 1993;82:903–12.
Goldman AS, Goldblum RM. A defense agents in milk. In: Jensen RG, editor. Handbook of milk composition. San Diego: Academic; 1995. p. 727–45.
Zopf D, Roth S. Oligosaccharide anti-infective agents. Lancet. 1996;347:1017–21.
Coppa GV, Zampini L, Galeazzi T, Facinelli B, Ferrante L, Capretti R, et al. Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrheal pathogens: Escherichia coli, vibrio cholerae, and Salmonella fyris. Pediatr Res. 2006;59:377–82.
Martín-Sosa S, Martín M-J, Hueso P. The sialylated fraction of milk oligosaccharides is partially responsible for binding to enterotoxigenic and uropathogenic escherichia coli human strains. J Nutr. 2002;132:3067–72.
Crane JK, Azar SS, Stam A, Newburg DS. Oligosaccharides from human milk block binding and activity of the Escherichia coli heat-stable enterotoxin (STa) in T84 intestinal cells. J Nutr. 1994;124:2358–64.
Xu H-T, Zhao Y-F, Lian Z-X, Fan B-L, Zhao Z-H, Yu S-Y, et al. Effects of fucosylated milk of goat and mouse on Helicobacter pylori binding to Lewis b antigen. World J Gastroenterol. 2004;10:2063–6.
El-Hawiet A, Kitova EN, Klassen JS. Recognition of human milk oligosaccharides by bacterial exotoxins. Glycobiology. 2015;25:845–54.
Chichlowski M, German JB, Lebrilla CB, Mills DA. The influence of Milk oligosaccharides on microbiota of infants: opportunities for formulas. Annu Rev Food Sci Technol. 2011;2:331–51.
Kavanaugh DW, O’Callaghan J, Buttó LF, Slattery H, Lane J, Clyne M, et al. Exposure of Bifidobacterium longum subsp. infantis to milk oligosaccharides increases adhesion to epithelial cells and induces a substantial tanscriptional response. PLoS ONE. 2013;8(6):e67224.
Cravioto A, Tello A, Villafan H, Ruiz J, Del Vedovo S, Neeser JR. Inhibition of localized adhesion of enteropathogenic Escherichia coli to HEp-2 cells by immunoglobulin and oligosaccharide fractions of human colostrum and breast milk. J Infect Dis. 1991;163:1247–55.
Asakuma S, Hatakeyama E, Urashima T, Yoshida E, Katayama T, Yamamoto K, et al. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J Biol Chem. 2011;286:34583–92.
Hakkarainen J, Toivanen M, Leinonen A, Frängsmyr L, Strömberg N, Lapinjoki S, et al. Human and bovine milk oligosaccharides inhibit Neisseria meningitidis pili attachment in vitro. J Nutr. 2005;135:2445–8.
Yu Z-T, Nanthakumar NN, Newburg DS. The human milk oligosaccharide 2′-fucosyllactose quenches campylobacter jejuni–induced inflammation in human epithelial cells HEp-2 and HT-29 and in mouse intestinal mucosa. J Nutr. 2016;146:1980–90.
Perdijk O, van Baarlen P, Fernandez-Gutierrez MM, van den Brink E, Schuren FHJ, Brugman S, et al. Sialyllactose and galactooligosaccharides promote epithelial barrier functioning and distinctly modulate microbiota composition and short chain fatty acid production in vitro. Front Immunol. 2019;10:1–14.
He Y, Liu S, Kling DE, Leone S, Lawlor NT, Huang Y, et al. The human milk oligosaccharide 2′-fucosyllactose modulates CD14 expression in human enterocytes, thereby attenuating LPS-induced inflammation. Gut. 2014;65(1):33–46.
Holscher HD, Bode L, Tappenden KA. Human milk oligosaccharides influence intestinal epithelial cell maturation in vitro. J Pediatr Gastroenterol Nutr. 2017;64:296–301.
Facinelli B, Marini E, Magi G, Zampini L, Santoro L, Catassi C, et al. Breast milk oligosaccharides: effects of 2′-fucosyllactose and 6′-sialyllactose on the adhesion of Escherichia coli and Salmonella fyris to Caco-2 cells. J Matern Neonatal Med. 2019;32(17):2950–2.
Newburg DS, Ruiz-Palacios GM, Altaye M, Chaturvedi P, Meinzen-Derr J, de Lourdes GM, et al. Innate protection conferred by fucosylated oligosaccharides of human milk against diarrhea in breastfed infants. Glycobiology. 2004;14:253–63.
Elison E, Vigsnaes LK, Rindom Krogsgaard L, Rasmussen J, Sorensen N, McConnell B, et al. Oral supplementation of healthy adults with 2′-O-fucosyllactose and lacto-N-neotetraose is well tolerated and shifts the intestinal microbiota. Br J Nutr. 2016;116(8):1356–68. https://doi.org/10.1017/S0007114516003354.
Underwood MA, Gaerlan S, De Leoz MLA, Dimapasoc L, Kalanetra KM, Lemay DG, et al. Human milk oligosaccharides in premature infants: absorption, excretion, and influence on the intestinal microbiota. Pediatr Res. 2015;78(6):670–7. https://doi.org/10.1038/pr.2015.162.
Boudry G, Hamilton MK, Chichlowski M, Wickramasinghe S, Barile D, Kalanetra KM, et al. Bovine milk oligosaccharides decrease gut permeability and improve inflammation and microbial dysbiosis in diet-induced obese mice. J Dairy Sci. 2017;100:2471–81.
Grabinger T, Glaus Garzon JF, Hausmann M, Geirnaert A, Lacroix C, Hennet T. Alleviation of intestinal inflammation by oral supplementation with 2-Fucosyllactose in mice. Front Microbiol. 2019;10:1–14. https://doi.org/10.3389/fmicb.2019.01385.
Monaco MH, Wang M, Pan X, Li Q, Richards JD, Chichlowski M, et al. Evaluation of sialyllactose supplementation of a prebiotic-containing formula on growth, intestinal development, and bacterial colonization in the neonatal piglet. Curr Dev Nutr. 2018;2:1–15.
Li M, Monaco MH, Wang M, Comstock SS, Kuhlenschmidt TB, Fahey GC, et al. Human milk oligosaccharides shorten rotavirus-induced diarrhea and modulate piglet mucosal immunity and colonic microbiota. ISME J. 2014;8(8):1609–20. https://doi.org/10.1038/ismej.2014.10.
Cilieborg MS, Sangild PT, Jensen ML, Østergaard MV, Christensen L, Rasmussen SO, et al. α1,2-Fucosyllactose does not improve intestinal function or prevent Escherichia coli F18 diarrhea in newborn pigs. J Pediatr Gastroenterol Nutr. 2017;64:310–8.
Rasmussen SO, Martin L, Østergaard MV, Rudloff S, Roggenbuck M, Nguyen DN, et al. Human milk oligosaccharide effects on intestinal function and inflammation after preterm birth in pigs. J Nutr Biochem. 2017;40:141–54.
Acknowledgments
The authors acknowledge intellectual support from Dr. Phillip S. Tong of American Dairy Products Institute and also Dr. Marcos Duarte, Ms. Yi-Chi Cheng, Mr. Vitor Hugo Moita, Mr. Zixiao Deng, Mr. Jonathan Baker, Mr. Hyunjun Choi, and Ms. Xiangyi Xu of the Kim Lab at North Carolina State University (Raleigh, NC, USA).
Funding
This study was awarded and funded by American Dairy Products Institute (Elmhurst, IL, USA), USDA-NIFA Hatch Fund (#02636, Washington DC, USA), and North Carolina Agricultural Foundation (#660101, Raleigh, NC, USA).
Author information
Authors and Affiliations
Contributions
KBJ and SWK conceived the purpose and concept of the manuscript. KBJ and SWK critically revised the manuscript. KBJ and SWK collected previous literature and wrote the manuscript. KBJ and SWK was responsible for the meta-analysis of publication. Both authors have read and agreed to the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no real or perceived conflicts of interest.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Jang, K.B., Kim, S.W. Role of milk carbohydrates in intestinal health of nursery pigs: a review. J Animal Sci Biotechnol 13, 6 (2022). https://doi.org/10.1186/s40104-021-00650-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-021-00650-7