
Abstract
Background
Betaine affects fat metabolism in animals, but the specific mechanism is still not clear. The purpose of this study was to investigate possible mechanisms of betaine in altering lipid metabolism in muscle tissue in finishing pigs.
Methods
A total of 120 crossbred gilts (Landrace × Yorkshire × Duroc) with an average initial body weight of 70.1 kg were randomly allotted to three dietary treatments. The treatments included a corn–soybean meal basal diet supplemented with 0, 1250 or 2500 mg/kg betaine. The feeding experiment lasted 42 d.
Results
Betaine addition to the diet significantly increased the concentration of free fatty acids (FFA) in muscle (P < 0.05). Furthermore, the levels of serum cholesterol and high-density lipoprotein cholesterol were decreased (P < 0.05) and total cholesterol content was increased in muscle (P < 0.05) of betaine fed pigs. Experiments on genes involved in fatty acid transport showed that betaine increased expression of lipoprotein lipase(LPL), fatty acid translocase/cluster of differentiation (FAT/CD36), fatty acid binding protein (FABP3) and fatty acid transport protein (FATP1) (P < 0.05). The abundance of fatty acid transport protein and fatty acid binding protein were also increased by betaine (P < 0.05). As for the key factors involved in fatty acid oxidation, although betaine supplementation didn’t affect the level of carnitine and malonyl-CoA, betaine increased mRNA and protein abundance of carnitine palmitransferase-1(CPT1) and phosphorylated-AMPK (P < 0.05).
Conclusions
The results suggested that betaine may promoted muscle fatty acid uptake via up-regulating the genes related to fatty acid transporter including FAT/CD36, FATP1 and FABP3. On the other hand, betaine activated AMPK and up-regulated genes related to fatty acid oxidation including PPARα and CPT1. The underlying mechanism regulating fatty acid metabolism in pigs supplemented with betaine is associated with the up-regulation of genes involved in fatty acid transport and fatty acid oxidation.
Similar content being viewed by others

Background
Betaine is a derivative of the amino acid glycine with three chemically reactive methyl groups. Betaine is distributed widely in animals, plants and microorganisms, and it is also a metabolite of choline oxidation in animals [1]. The principal physiologic role of betaine is as a methyl group donor [2], which means betaine participates in many important biochemical pathways, including methionine-homocysteine cycle and the biosynthesis of many compounds such as carnitine, creatine and phospholipids. Since carnitine is required for transport of long chain fatty acids into mitochondria [3], scientists have paid much attention to effects of betaine on energy metabolism especially lipid metabolism in animals. Studies showed that dietary betaine supplementation affected energy partitioning in pigs [4, 5] and it’s also widely reported that betaine promotes animal growth and decreases carcass fat percentage in finishing pigs [6,7,8,9,10]. Further investigations found that betaine supplementation could decrease hepatic triglyceride accumulation [11, 12] and prevent fatty liver in rats fed high-fat-diets [13, 14]. The intramuscular fat content in the longissimus muscle was increased when pigs were fed betaine [15, 16]. Madeira et al. [17] reported that betaine might be involved in the differential regulation of some key genes of lipid metabolism in muscle and subcutaneous adipose tissue. However, studies on the mechanism of betaine affecting lipid metabolism in muscle are lacking. Therefore, the objective of the present study was to investigate possible mechanisms of betaine in altering lipid metabolism in muscle tissue of finishing pigs.
Methods
Animals and treatments
The experiment protocol used in this study was approved by the Institutional Animal Care and Use Committee of Zhejiang University. A total of 120 crossbred gilts (Landrace × Yorkshire × Duroc) with an average initial body weight of 70.1 kg (SD 0.70 kg) were randomly allotted to three dietary treatments. Each treatment consisted of four pens replicates with 10 gilts per pen. The treatment diets included a corn–soybean meal basal (Table 1) supplemented with 0, 1250 mg/kg (Low Betaine) or 2500 mg/kg (High Betaine) betaine (provided by Healthy Husbandry Sci-tech Co., Ltd. Hangzhou, China) respectively at the expense of corn. The basal diet was formulated to meet or exceed the nutrient requirements of finishing pigs [18]. Chemical analyses of the basal diet were carried out according to the methods of AOAC [19]. The feeding experiment lasted 42 d after a 7-day adaptation period. All pigs were housed in a curtain-sided pig barn with concrete slotted floors. Feed and water were provided for ad libitum consumption throughout the experiment.
Sample collection
At the end of the trial, eighteen pigs (six from each dietary treatment) weighing about 111.8 kg (SD 2.08 kg) were selected to collect tissue samples. Following an overnight fast, pigs were stunned by electrical shock and bleeding. Individual blood samples were collected at slaughter during exsanguinations. After collection of blood, samples were kept at room temperature for 2 h and then centrifuged for 10 min at 3000×g at 4 °C. Serum was collected and frozen at −80 °C until subsequent analyses. Samples of longissimus muscle between the 6th and 7th rib were obtained on the left side of the carcass within 5 min after slaughter, and then snap frozen in liquid nitrogen and stored at −80 °C until subsequent analyses.
Analysis of lipid metabolites in serum
Serum concentration of high-density lipoprotein cholesterol (HDLC), total cholesterol (TC), free fatty acid (FFA) and triglyceride were measured with commercial assay kits (Nanjing Jiancheng Bio-engineering Institute, Code No. A112–2, A111–2, A042–1 and A110–2, respectively, Nanjing, China) following the manufacturer’s instructions.
Muscle lipid metabolites analysis
A 10% muscle homogenate was prepared with a mixture of chloroform and formaldehyde (a volume ratio of 2:1). Then extracted at room temperature for 24 h [20]. The organic solvent layer was taken and the level of triglyceride in muscle was measured with commercial assay kit (Nanjing Jiancheng Bio-engineering Institute, A110–2, Nanjing, China). Before the levels of TC and FFA in muscle were measured by the kits (Nanjing Jiancheng Bio-engineering Institute, Code No. A112–2, A111–2, A042–1 and A110–2, respectively, Nanjing, China), muscle tissue was made homogenate with physiological saline. The concentrations of carnitine and malonyl-CoA were measured using ELISA kits (Biovol Technologies, Code No.50R–E.3088P & 50R–E.3035P, Shanghai, China) for porcine assay according to the instructions.
RT-PCR analysis
Total RNA was extracted from frozen porcine muscle tissue using the Trizol reagent as described by the manufacturer (Invitrogen). The RNA concentration and purity were determined by the NanoDrop ND-2000 spectrophotometer (Thermofisher, USA) and its integrity was confirmed by agarose gel electrophoresis. The cDNA synthesis was performed in a 10-μL reaction volume containing 2 μg total RNA using the SYBR PrimeScript™ RT-PCR kit with gDNA Eraser (Code No. RR047A, TaKaRa, Dalian, China). Genomic DNA is eliminated by treatment for 2 min at 42 °C with gDNA Eraser, which has potent DNA degrading activity. Then a reverse-transcription reaction reagent is added that includes a component that completely inhibits DNA degradation activity, and the reverse-transcription reaction proceeds for 15 min at 37 °C. The abundance of the target genes was measured by quantitative real-time PCR, performed with the ABI Stepone Plus™ RT-PCR system (ABI Biotechnology, USA) using SYBR Premix Ex Taq™ (Tli RNaseH Plus) RT-RCR kit (TaKaRa, Dalian, China). Primers for the selected genes were synthesized commercially by Invitrogen (Shanghai, China), shown in Table 2. The reaction protocol comprised a cycle of 95 °C for 1 min, 40 cycles of 95 °C for 10 s and 64 °C for 25 s. The expression of the target genes were normalized by the endogenous housekeeping gene (β-actin) [21, 22]. Each sample was analyzed in triplicate and the PCR amplification efficiency was close to 100%. The gene expression was calculated by using the comparative (2 − ΔΔCt) method [23].
Western blot analysis
Protein form muscle samples was extracted by T-PER Tissue Protein Extraction Reagent containing protease inhibitor cocktail (Thermo Pierce, Code No.78510, USA), and quantified with BCA protein assay kit (Beyotime, Code No.P0010, Shanghai, China) according to kit instructions. Proteins were separated on SDS - PAGE gels (12%), and then electrophoretically transferred onto immobilon-P polyvinylidene fluoride membranes (PVDF membrane, Millipore, Code No. IPVH00010, America). Membranes were blocked 1 h in Tris-buffered saline containing 5% nonfat-dried milk at room temperature. Membranes were then incubated overnight at 4 °C in blocking buffer containing primary antibodies (as shown in Table 3). A goat anti-rabbit IgG (H + L) Secondary antibody (Thermo Pierce, Code NO.31210, USA) with 1/5000 dilution was used in the detection of specific proteins. For loading control, β-actin antibody was used as control. In addition, the relative expression of p-AMPK was normalized with AMPK. Finally, Super Signal West Dura Extended Duration Substrate (Thermo Pierce, Code No. 34075, USA) was used to visualize the protein bands. Band intensities were determined by using BandScan 5.0 software.
The relative expressions of target proteins = (The optical density of target proteins/The optical density of β-actin).
Statistical analyses
Results were presented as means and standard deviations. Statistical analysis was performed by one-way analysis of variance (ANOVA) and the Duncan method was used to put up multiple comparison with the statistical software SPSS 19.0. In all analyses, the level of significant difference was set at P < 0.05.
Results
Betaine on serum lipid metabolites
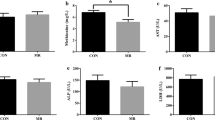
As shown in Fig. 1, there was no significant difference in the levels of serum FFA and triglyceride in the pigs fed betaine compared with control group. Additionally, the concentration of HDLC and TC were significantly lower in the betaine treated pigs (P < 0.05).

Effect of betaine supplementation on serum parameters of lipid metabolism. The levels of serum free fatty acid (FFA, a), triglyceride (b), total cholesterol (c) and high-density lipoprotein cholesterol (HDLC, d). a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively
Betaine on muscle lipid metabolites
The level of FFA and TC were markedly higher in muscle when pigs were fed betaine (P < 0.05, Fig. 2). Compared to the control group, the level of triglyceride in muscle was not affected by betaine addition (P > 0.05).

Effect of betaine supplementation on total cholesterol, FFA and triglyceride in muscle. The levels of total cholesterol (a), free fatty acid (FFA, b) and triglyceride (c) in muscle. a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively
Key factors involved in muscle FFA intake
As shown in Fig. 3, the gene expression of FAT/CD36, FATP1 and PPARγ (P < 0.05) were higher in betaine-fed groups than control group. The addition of 2500 mg/kg betaine markedly up-regulated the gene expression of FABP3 and LPL (P < 0.05). In addition, the abundance of fatty acid transport protein and fatty acid binding protein were significantly increased by betaine supplementation (P < 0.05, Fig. 4).

The relative gene expression of key factors involved fatty acid uptake in muscle. mRNA expression was performed by RT-PCR and β-actin was chosen as reference gene. aThe relative expression of FAT/CD36, FATP1, LPL and PPARγ in muscle, (b) The relative expression of FABP3 in muscle. a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively

The relative protein abundance of FATP1 and FABP3 in muscle. Western blot results were shown in a (The control group: 1–1, 1–2, 1–3; Low betaine group: 2–1, 2–2, 2–3; High betaine group: 3–1, 3–2, 3–3). Data were normalized with β-actin as shown in b, c. a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively
Key factors involved in muscle FA oxidation
Betaine supplementation did not affect carnitine or malonyl-CoA in muscle compared to the control group (P > 0.05, Fig. 5).

Effect of betaine supplementation on the level of carnitine(a) and malonyl-CoA (b) in muscle. Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively
The gene expression of AMPKα2, PPARα and CPT1 were significantly higher in pigs fed with betaine than the control group. (P < 0.05, Fig. 6). Furthermore, betaine supplementation markedly increased the abundance of phosphorylated-AMPK and CPT1 in muscle (P < 0.05, Fig. 7).

The relative mRNA expression of factors involved in fatty acid oxidation in muscle. mRNA expression was performed by RT-PCR and β-actin was chosen as reference gene. a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively

The relative protein abundance of p-AMPK and M-CPT1. The results of western blot were showed a and b (The control group: 1–1, 1–2, 1–3; Low betaine group: 2–1, 2–2, 2–3; High betaine group: 3–1, 3–2, 3–3). p-AMPK (the activated form of AMPK) was normalized with AMPK (shown in c) and MCPT1(the muscle type of CPT1) was normalized with β-actin (shown in d). a,bValues without common superscript letters differ significantly (P < 0.05). Low betaine and high betaine represent 1250 mg/kg and 2500 mg/kg betaine addition, respectively
Discussion
Fatty acid metabolism in muscle includes uptake, synthesis and oxidation [24,25,26], but the synthesis is at a slow rate [27]. The main source of fatty acid in muscle tissue includes transport from plasma and hydrolysis from chylomicron and very-low-density-lipoprotein (VLDL) with LPL. Our study found that the concentration of FFA was significantly increased in muscle when pigs were fed betaine, similarly to the studies carried out by Yang et al. [28] and Fernández-Fígares et al. [29]. We speculated that the transport of FFA and/or the hydrolysis may be enhanced. More experiments were carried out regarding factors involved in fatty acid transport in muscle tissue. It is widely recognized that long chain fatty acid (LCFA) cross the plasma membrane via a protein-mediated mechanism. A number of fatty acid transporters have been identified, including fatty acid translocase/cluster of differentiation (FAT/CD36) and fatty acid transport proteins (FATP1) [30]. We found that betaine supplementation up-regulated gene expression for FATP1 and FAT/CD36. Experiments in vitro have shown that over expression of FATP1 increased the uptake of LCFA in cells [31] and studies in vivo documented that muscle-specific over-expression of FAT/CD36 enhanced cellular fatty acid uptake in mice [32]. FABP3, another important protein in fatty acid transportation, plays a role in transporting fatty acid from the sarcolemma to their intracellular sites of metabolism [33]. In muscle cells, the intracellular transport of LCFAs is facilitated to a great extent by FABP3 [34] Additionally, FABP3 is confirmed to be associated with intramuscular fat in pigs [35]. Our studies showed that feeding betaine up-regulated the protein abundance of FABP3. In addition, the gene expression of FABP3 was enhanced when pigs were fed with 2500 mg/kg betaine but no difference was found with 1250 mg/kg-betaine addition. The possible reason maybe that FABP3 expression is translationally rather than transcriptionally regulated [36]. In summary, betaine may promote the uptake of fatty acids in muscle via regulating the expression of FAT/CD36, FATP1 and FABP3. As mentioned above, LPL is the principal enzyme that hydrolyzes circulating triglycerides and it also can increase lipid uptake [37]. The results showed a significant increase in the gene expression of LPL with the addition of 2500 mg/kg betaine, which indicates betaine might enhance lipid uptake as well as chylomicron hydrolysis. The nuclear receptor PPARγ is a central regulator of adipose tissue development and an important modulator of expression in adipocytes [38]. To date, only a limited number of genes are known to be direct targets of PPARγ in adipose tissue. The majority of these encode proteins with direct links to lipid metabolism including LPL, FATP and FAT/CD36 [39, 40]. In present study, the gene expression of PPARγ was significantly higher in betaine-fed groups than the control group. We found that the effect of betaine on PPARγ was similar to its downstream target genes. All these results were similar to Albuquerque [41] and imply that betaine may facilitate fatty acids uptake in muscle via affecting key factors involved in FFA uptake, and the specific regulation mechanism needs more research.
The concentration of FFA in muscle tissue resultes from the balance of transport and oxidation. As a methyl donor, betaine participates in the biosynthesis of carnitine and because of this, betaine may be related to fatty acid β-oxidation. LCFAs are first transformed into acyl CoA, then transferred into mitochondria after combining with carnitine where it is oxidized. Carnitine palmitoyl transferase I (CPT1) is the rate-limiting enzyme that controls the step of combination and malony-CoA is an allosteric inhibitor of CPT1 [42]. Whereas the synthesis of malonyl-CoA is catalyzed by acetyl-CoA carboxylase (ACC), the activity of the ACC is regulated by phosphorylation of AMPK [43]. Hence, AMPK-ACC-CPT1 is an important signaling pathway to regulate fatty acid β-oxidation in mitochondria. Cai et al. [44] found that gestational dietary betaine supplementation down-regulated expression of ACC in neonatal piglets and Pekkinen et al. [11] found betaine supplementation had an impact on carnitine metabolism in high-fat-fed mice. Our experiment didn’t find significant changes in muscle concentrations of malonyl-CoA or carnitine. The different results might be related to the different experiment condition and the mechanism needs to be further investigated. Increased gene expression and protein expression of CPT1 were up-regulated with betaine addition, which implied betaine may enhance fatty acid β-oxidation in muscle tissue. However, others have shown betaine supplementation reduced the activity of CPT1 and mRNA abundance, and further increased IMF in finishing pigs [Duroc × (Seghers × Seghers)] [15]. We speculate that the effect of betaine addition on CPT1 might be influenced by breed and muscle type. In order to get a better understanding, we further analyzed effects of betaine on AMP-activated protein kinase (AMPK) and PPARα, which are both upstream regulatory factors of CPT1. AMPK is a crucial energy sensor for cells, which can promote the catabolism of fatty acids by enhancing their uptake into mitochondria and their consequent breakdown by beta-oxidation [45]. It was reported that activated AMPK in muscle enhances the gene expression of PPARα and CPT1 [46], and CPT1 also seems to be a target of PPARα [47]. In the current experiment, the gene expression of both PPARα and AMPK were higher in betaine-fed groups as well as protein expression of p-AMPK (the activated form of AMPK). Similar to our previous report in rat liver [12], it can be inferred that betaine affected fatty acid oxidation in muscle via activating AMPK and up-regulated PPARα and CPT1 gene expression.
The effect of betaine supplementation on cholesterol metabolism was of interest. The present study showed that betaine supplementation decreased the concentration of serum cholesterol and HDLC and increased cholesterol level in muscle, which was consistent with the studies by Albuquerque et al. [41] and Yang et al. [20]. However, Matthews et al. [48] and Martins et al. [49] reported that betaine supplemented pigs presented higher serum cholesterol. The efficacy of betaine in regulating the concentration of cholesterol in pigs shows variable results and seems to depend on both animal and dietary factors. Although the results were inconsistent, it seems to indicate that betaine might affect cholesterol partitioning or maybe enhances the transport of cholesterol in pigs, and more research is needed to clarify the specific mechanism.
Conclusions
In present study, betaine supplementation increased the level of free fatty acids in muscle, which may have resulted due to a change in the balance of fatty acid uptake and oxidation. Betaine may promote fatty acid uptake via increasing the expression of fatty acid transporters including FAT/CD36, FATP1 and FABP3 in muscle. Additionally, betaine activated AMPK and up-regulated PPARα and CPT1, and may enhance fatty acid oxidation in muscle. Fatty acid accretion in muscle represents a balance between uptake and oxidation, and it seems that the effect of betaine on uptake was stronger than oxidation.
Abbreviations
- CPT1:
-
Carnitine palmitoyl transferase 1
- FABP3:
-
Fatty acid binding protein
- FAT/CD36:
-
Fatty acid translocase/cluster of differentiation
- FATP1:
-
Fatty acid transporter protein1
- FFA:
-
Free fatty acid
- HDLC:
-
High-density lipoprotein cholesterol
- LPL:
-
Lipoprotein lipase
- TC:
-
Total cholesterol
References
Eklund M, Bauer E, Wamatu J, Mosenthin R. Potential nutritional and physiological functions of betaine in livestock. Nutr Res Rev. 2005;18(01):31–48.
Craig SAS. Betaine in human nutrition. Am J Clin Nutr. 2004;80(3):539–49.
Wray-Cahen D, Fernández-Fígares I, Virtanen E, Steele NC, Caperna TJ. Betaine improves growth, but does not induce whole body or hepatic palmitate oxidation in swine (Sus Scrofa Domestica). Comp Biochem Physiol A Mol Integr Physiol. 2004;137(1):131–40.
Fernández-Fígares I, Wray-Cahen D, Steele NC, Campbell RG, Hall DD, Virtanen E, et al. Effect of dietary betaine on nutrient utilization and partitioning in the young growing feed-restricted pig. J Anim Sci. 2002;80(2):421–8.
Schrama JW, Heetkamp MJW, Simmins PH, Gerrits WJJ. Dietary betaine supplementation affects energy metabolism of pigs. J Anim Sci. 2003;81(5):1202–9.
Wang YZ, Xu ZR, Feng J. The effect of betaine and DL-methionine on growth performance and carcass characteristics in meat ducks. Anim Feed Sci Technol. 2004;116(1):151–9.
Feng J, Liu X, Wang YZ, Xu ZR. Effects of betaine on performance, carcass characteristics and hepatic betaine-homocysteine methyltransferase activity in finishing barrows. Asian-Australas J Anim Sci. 2006;19(3):402.
Huang QC, Xu ZR, Han XY, Li WF. Changes in hormones, growth factor and lipid metabolism in finishing pigs fed betaine. Livest Sci. 2006;105(1):78–85.
Huang QC, Xu ZR, Han XY, Li WF. Effect of betaine on growth hormone pulsatile secretion and serum metabolites in finishing pigs. J Anim Physiol Anim Nutr (Berl). 2007;91(3–4):85–90.
Nakev J, Popova T, Vasileva V. Influence of dietary betaine supplementation on the growth performance and carcass characteristics in male and female growing-finishing pigs. Bulg J Agric Sci. 2009;15(3):263–8.
Pekkinen J, Olli K, Huotari A, Tiihonen K, Keski-Rahkonen P, Lehtonen M, et al. Betaine supplementation causes increase in carnitine metabolites in the muscle and liver of mice fed a high-fat diet as studied by nontargeted LC-MS metabolomics approach. Mol Nutr Food Res. 2013;57(11):1959–68.
Xu L, Huang D, Hu Q, Wu J, Wang Y, Feng J. Betaine alleviates hepatic lipid accumulation via enhancing hepatic lipid export and fatty acid oxidation in rats fed with a high-fat diet. Br J Nutr. 2015;113(12):1835–43.
Zhang W, Wang L, Wang L, Li X, Zhang H, Luo L, et al. Betaine protects against high-fat-diet-induced liver injury by inhibition of high-mobility group box 1 and toll-like receptor 4 expression in rats. Dig Dis Sci. 2013;58(11):3198–206.
Deminice R, Da Silva RP, Lamarre SG, Kelly KB, Jacobs RL, Brosnan ME, et al. Betaine supplementation prevents fatty liver induced by a high-fat diet: effects on one-carbon metabolism. Amino Acids. 2015;47(4):839–46.
Huang QC, Han XY, Xu ZR, Yang XY, Chen T, Zheng XT. Betaine suppresses carnitine palmitoyltransferase I in skeletal muscle but not in liver of finishing pigs. Livest Sci. 2009;126(1):130–5.
Martins JM, Neves JA, Freitas A, Tirapicos JL. Effect of long-term betaine supplementation on chemical and physical characteristics of three muscles from the Alentejano pig. J Sci Food Agric. 2012;92(10):2122–7.
Madeira MS, Rolo EA, Alfaia CM, Pires VR, Luxton R, Doran O, et al. Influence of betaine and arginine supplementation of reduced protein diets on fatty acid composition and gene expression in the muscle and subcutaneous adipose tissue of cross-bred pigs. Br J Nutr. 2016;115(06):937–50.
National Research Council. Nutrient requirements of swine: eleventh revised edition. Washington, D.C.: The National Academies Press; 2012.
Association of Official Analytical Chemists (AOAC). Van Nostrand's Encyclopedia of Chemistry. Hoboken: John Wiley & Sons, Inc. 2005.
Zeng T, Xie K, Zhang C, Yu L, Zhu Z. Determination the level of triglycerides in liver with chloroform/ methanol homogenate. J Hyg Res. 2008;37(5):550–1. (in Chinese)
Nygard A-B, Jørgensen CB, Cirera S, Fredholm M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol Biol. 2007;8(1):67.
Erkens T, Van Poucke M, Vandesompele J, Goossens K, Van Zeveren A, Peelman LJ. Development of a new set of reference genes for normalization of real-time PT-PCR data of porcine backfat and longissimus dorsi muscle, and evaluation with PPARGC1A. BMC Biotechnol. 2006;6:41.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods. 2001;25(4):402–8.
Lopaschuk GD, Ussher JR, Folmes CDL, Jaswal JS, Stanley WC. Myocardial fatty acid metabolism in health and disease. Physiol Rev. 2010;90(1):207–58.
Kelley DE, Goodpaster B, Wing RR, Simoneau J-A. Skeletal muscle fatty acid metabolism in association with insulin resistance, obesity, and weight loss. Am J Physiol-Endoc M. 1999;277(6):E1130–41.
Clarke SD. Regulation of fatty acid synthase gene expression: an approach for reducing fat accumulation. J Anim Sci. 1993;71(7):1957.
Christensen K. In vitro studies on the synthesis of intramuscular fat in the longissimus dorsh muscle of pigs. Livest Prod Sci. 1975;2(1):59–68.
Yang HS, Lee JI, Joo ST, Park GB. Effects of dietary glycine betaine on growth and pork quality of finishing pigs. Asian-australas J Anim Sci. 2009;22(5):706–11.
Fernández-Fígares I, Lachica M, Martín A, Nieto R, Gonzalez-valero L, Rodriguez-Lopez JM, et al. Impact of dietary betaine and conjugated linoleic acid on insulin sensitivity, protein and fat metabolism of obese pigs. Animal. 2012;6(07):1058–67.
Nickerson JG, Alkhateeb H, Benton CR, Lally J, Nickerson J, Han XX, et al. Greater transport efficiencies of the membrane fatty acid transporters FAT/CD36 and FATP4 compared with FABPpm and FATP1 and differential effects on fatty acid esterification and oxidation in rat skeletal muscle. J Biol Chem. 2009;284(24):16522–30.
Sebastián D, Guitart M, García-Martínez C, Mauvazin C, Orellana-Gavalda JM, Serra D, et al. Novel role of FATP1 in mitochondrial fatty acid oxidation in skeletal muscle cells. J Lipid Res. 2009;50(9):1789–99.
Ibrahimi A, Bonen A, Blinn WD, Hajri T, Li X, Zhong K, et al. Muscle-specific overexpression of FAT/CD36 enhances fatty acid oxidation by contracting muscle, reduces plasma triglycerides and fatty acids, and increases plasma glucose and insulin. J Biol Chem. 1999;274(38):26761–6.
Hertzel AV, Bernlohr DA. The mammalian fatty acid-binding protein multigene family: molecular and genetic insights into function. Trends Endocrinol Metab. 2000;11(5):175–80.
Glatz JFC, Schaap FG, Binas B, Bonen A, Van Der Vusse GJ, Luiken JJFP. Cytoplasmic fatty acid-binding protein facilitates fatty acid utilization by skeletal muscle. Acta Physiol Scand. 2003;178(4):367–71.
Cho KH, Kim MJ, Jeon GJ, Chung HY. Association of genetic variants for FABP3 gene with back fat thickness and intramuscular fat content in pig. Mol Biol Rep. 2011;38(3):2161–6.
Gerbens F, Verburg FJ, Van Moerkerk HTB, Engel B, Buist W, Veerkamp JH, et al. Associations of heart and adipocyte fatty acid-binding protein gene expression with intramuscular fat content in pigs. J Anim Sci. 2001;79(2):347–54.
Yagyu H, Chen G, Yokoyama M, Hirata K, Augustus A, Kako Y, et al. Lipoprotein lipase (LpL) on the surface of cardiomyocytes increases lipid uptake and produces a cardiomyopathy. J Clin Invest. 2003;111(3):419–26.
Walczak R, Tontonoz P. PPARadigms and PPARadoxes: expanding roles for PPARγ in the control of lipid metabolism. J Lipid Res. 2002;43(2):177–86.
Rosen ED, Walkey CJ, Puigserver P, Spiegelman BM. Transcriptional regulation of adipogenesis. Genes Dev. 2000;14(11):1293–307.
Ahmadian M, Suh JM, Hah N, Liddle C, Atkins AR, Downes M, et al. PPAR [gamma] signaling and metabolism: the good, the bad and the future. Nat Med. 2013;99(5):557–66.
Albuquerque A, Neves JA, Redondeiro M, Laranjo M, Felix MR, Freitas A, et al. Long term betaine supplementation regulates genes involved in lipid and cholesterol metabolism of two muscles from an obese pig breed. Meat Sci. 2017;124:25–33.
Saha AK, Ruderman NB. Malonyl-CoA and AMP-activated protein kinase: an expanding partnership. Mol Cell Biochem. 2003;253(1):65–70.
Xue B, Kahn BB. AMPK integrates nutrient and hormonal signals to regulate food intake and energy balance through effects in the hypothalamus and peripheral tissues. J Physiol Lond. 2006;574(1):73–83.
Cai D, Wang J, Jia Y, Liu H, Yuan M, Dong H, et al. Gestational dietary betaine supplementation suppresses hepatic expression of lipogenic genes in neonatal piglets through epigenetic and glucocorticoid receptor-dependent mechanisms. Biochim Biophys Acta. 2016;1861(1):41–50.
Hardie DG, Ross FA, Hawley SA. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol. 2012;13(4):251–62.
Lee WJ, Kim M, Park HS, Kim HS, Jeon MJ, Oh KS, et al. AMPK activation increases fatty acid oxidation in skeletal muscle by activating PPARα and PGC-1. Biochem Biophys Res Commun. 2006;340(1):291–5.
Ferré P. The biology of peroxisome proliferator-activated receptors. Diabetes. 2004;53(suppl 1):S43–50.
Matthews JO, Southern LL, Higbie AD, Persica MA, Bidner TD. Effects of betaine on growth, carcass characteristics, pork quality, and plasma metabolites of finishing pigs. J Anim Sci. 2001;79(3):722–8.
Martins JM, Neves JA, Freitas A, Tirapicos JL. Research article Betaine supplementation affects the cholesterol but not the lipid profile of pigs. Eur J Lipid Sci Technol. 2010;112:295–303.
Acknowledgements
We would like to thank Healthy Husbandry Sci-tech Co., Ltd. (Hangzhou, China) for their offer with betaine.
Availability of data and materials
The raw data for the current study are available from the corresponding author on reasonable request.
Funding
This work was funded and supported by National Basic Research Program of China (No.2012CB124705), Zhejiang Provincial Key Research and Development Program (2015C03006) and Provincial Key S&T Special Projects (2015C02022).
Author information
Authors and Affiliations
Contributions
YW and JF designed the study. SL and HW were involved in performing the experiment and data interpretation. SL drafted the manuscript and XW revised the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval
The experiment protocols used in this study was approved by the Institutional Animal Care and Use Committee of Zhejiang University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no completing interests.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, S., Wang, H., Wang, X. et al. Betaine affects muscle lipid metabolism via regulating the fatty acid uptake and oxidation in finishing pig. J Animal Sci Biotechnol 8, 72 (2017). https://doi.org/10.1186/s40104-017-0200-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-017-0200-6