
Abstract
In humans, an adaptable internal biological system generates circadian rhythms that maintain synchronicity of behavior and physiology with the changing demands of the 24-h environment. Development of the circadian system begins in utero and continues throughout the first few years of life. Maturation of the clock can be measured through sleep/wake patterns and hormone secretion. Circadian rhythms, by definition, can persist in the absence of environmental input; however, their ability to adjust to external time cues is vital for adaptation and entrainment to the environment. The significance of these external factors that influence the emergence of a stable circadian clock in the first years of life remain poorly understood. Infants raised in our post-modern world face adverse external circadian signals, such as artificial light and mistimed hormonal cues via breast milk, which may increase interference with the physiological mechanisms that promote circadian synchronization. This review describes the very early developmental stages of the clock and common circadian misalignment scenarios that make the developing circadian system more susceptible to conflicting time cues and temporal disorder between the maternal, fetal, infant, and peripheral clocks.
Similar content being viewed by others


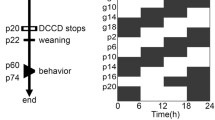
Introduction
The internal circadian clock of humans governs daily physiological functions, including the sleep/wake cycle and rhythms of the hormones melatonin and cortisol [1, 2]. In healthy adults and children, the central biological clock adheres to a 24-h cycle by entraining to environmental time cues, the strongest being light. Circadian disruption and dysregulation in adults are associated with a number of physical and mental health conditions, including insomnia, neurodegenerative disorders, metabolic disease, and psychiatric disorders [3,4,5,6,7]. Development of the circadian system begins in utero and continues throughout early life. Data from rodent studies indicate that circadian disruption during fetal development is associated with adult-onset cardiovascular disease, reduced bone mass and strength, glucose intolerance, insulin resistance, and mood disorder-related behaviors including anhedonia- and depression-like behaviors [8,9,10,11,12]. In human fetuses and infants, little is known about the links between circadian system maturation and health, as well as the factors that influence the development of an adaptive biological clock.
Sleep disruption during infancy is prevalent and persistent. Cross-sectional data show that 10–46% of infants ages 6–18 months exhibit some type of sleep disturbance (e.g., bedtime fussiness, difficulty falling asleep, frequent, and prolonged night awakenings) [13,14,15,16,17]. Persistence of infant sleep problems through the preschool years is common and can track into late childhood and adolescence [15, 17]. Results from previous studies indicate a relationship between parent-reported sleep difficulties and child psychosocial issues, including aggressive behavior, attention problems, anxiety, hyperactivity, and mood problems [17,18,19,20,21,22]. Additionally, lower cognitive performance and poor school functioning are associated with childhood sleep problems, suggesting that sleep issues may play a role in diminishing cognitive processes [23,24,25]. On the familial level, infant sleep problems are associated with higher maternal depression and poorer global health in both parents [14, 17, 26]. Identifying modifiable risk factors at the earliest stage of development provides an important opportunity to understand how to prevent and remedy infant sleep problems and related health conditions, both concurrent and those that emerge in the future.
The prevalence of infant sleep problems may be due to misalignment between environmental inputs and endogenous biological rhythms across the prenatal and post-natal periods that influence development of the circadian system. Human fetuses and infants mature in the context of an artificial light-saturated environment, and the acute and chronic effects of this circadian misalignment on development remain largely unknown. This comprehensive review will describe what is known about human circadian rhythms in fetal and infant development, as well as factors that influence sleep and the circadian system in early life. It will (1) discuss the two-process model of sleep regulation, a foundational framework for understanding the interaction between sleep homeostasis and the circadian rhythm; (2) review the fundaments of circadian physiology; and (3) present different proposed mechanisms underlying development of the circadian system (Fig. 1).

Model for circadian system development
Light is the strongest zeitgeber to the maternal suprachiasmatic nucleus (SCN) and regulates the diurnal fluctuation of maternal hormonal time cues directly to the fetus through the placenta and indirectly to the infant through breast milk. Additionally, light entrains the infant SCN via the developing retinohypothalamic tract (RHT). These environmental and maternal signals interact with the fetus and infant to influence the development of the circadian system.
Two-process model of sleep regulation
As first proposed by Borbély in 1982 [27], sleep regulation is governed by the interaction between a clock-dependent circadian process, Process C, and a sleep/wake dependent homeostatic process, Process S. Process C oscillates as an approximate 24-h rhythm, with the drive for sleep beginning in the evening and reaching its peak in the middle of the night, followed by a drive for wakefulness that rises in the early morning [28]. Process S is determined by the amount of prior wakefulness, such that the longer an individual is awake, the greater the “pressure” to sleep. Sleep pressure builds in an exponential fashion across the day and dissipates throughout time spent sleeping [29]. These two processes are independent but interact to control levels of alertness and sleep drive. As sleep pressure rises throughout the day, it is opposed by a stronger circadian drive for wakefulness. At bedtime, sleep propensity is promoted by the coordination of melatonin secretion and a high level of sleep pressure [30]. Melatonin is synthesized and released by the pineal gland and binds to MT1 and MT2 receptors in the suprachiasmatic nucleus (SCN) to help synchronize the circadian rhythm [31]. As sleep unfolds across the night, sleep pressure dissipates; however, the increase in the circadian promotion of sleep results in consolidation until waking in the morning. The synergy of these two systems is essential to promote optimal sleep at night and wakefulness during the day, and misalignment results in reduced sleep quality and duration [32,33,34,35].
Infant sleep
Newborns spend about 70% of their first few weeks after birth sleeping, and the timing of their sleep episodes is distributed equally throughout the 24-h day with no clear rhythm [36, 37]. At 2 weeks of age, infants sleep in approximately 4-h intervals [38]. As measured by actigraphy, Jenni and colleagues [39] observed day-night differences in rest-activity patterns within the first weeks of infancy (Fig. 2). By 5 weeks of age, there was emergence of an infradian rhythm, with a 25-h period as circadian rhythmicity began to appear [36]. At about 15 weeks, more consolidated wake and sleep episodes were apparent, and by 6 to 9 months of age most infants were able to sleep through the night, displaying at least 6-h consolidated sleep episodes [40,41,42]. Average total sleep duration across the first year of life remained about 14 h per day. However, it was the lengthening and timing of sleep and wake episodes, an increase in sleep during the night and an increase in wakefulness during the day, that matured with development. In other words, older infants, with more mature circadian regulation, were more likely to sleep during the night for longer durations than newborns. It should be noted, however, that alongside circadian system development, maturation of the sleep homeostatic process also plays a role in the emergence of this observed 24-h pattern in the first few months of life [40, 43]. Additionally, individual variability of sleep timing and duration is large [42], representing differences in the developmental manifestation of sleep-wake and circadian synchronicity.

Development of rest-activity patterns in infants
Double plotted rest-activity patterns of three infants. The horizontal axis represents 48 h, black areas represent activity, and white areas depict rest episodes. Data were continuously collected in 1-min epochs via actigraphy over 6 to 12 months. Initially, post-birth, infants had weak day-night asymmetry, with sleep and wake occurring at all hours of the 24-h day. Infants 1 and 5 developed an ultradian pattern of rest and activity that emerged in the first few weeks of life, followed by more nighttime sleep consolidation and a regular daytime nap schedule. Infant 2 displayed a less robust ultradian rhythm in the first few weeks and more irregular daytime naps. These three infants demonstrate the interindividual variability in rest-activity patterns observed in early development. Note: Reprinted from “Development of the 24-h rest-activity pattern in human infants,” by O. G. Jenni, T. Doboer, and P. Achermann, 2006, Infant Behavior & Development, 29, p. 143–152. Copyright 2005 by Elsevier Inc. Adapted with permission.
Circadian fundaments
Circadian (Latin for “circa diem,” meaning “around a day”) rhythms are endogenous biological rhythms within organisms that oscillate in the absence of environmental time cues, entrain to environmental signals, and remain stable in environments with fluctuating temperatures [44]. For biological processes to occur at the appropriate time in the proper sequence, organisms must be able to synchronize to and predict their external environment. This ‘temporal framework’ allows organisms to anticipate daily changes and optimize their behavior and physiology in advance of the daily demands of the light-dark cycle [1].
At the molecular level, the SCN maintains 24-h rhythmicity through positive and negative transcriptional-translational feedback loops of clock genes [45]. In mammals, the positive feedback loop is initiated during the biological day through positive regulating genes Bmal1 (Brain and muscle Arnt-like factor) and Clock (Circadian locomotor output cycle kaput) and their protein products [1, 46]. Within the nucleus, proteins BMAL1 and CLOCK heterodimerize and bind to E-box response elements within promoter regions of negative regulating genes Per (Per1 and Per2) and Cry (Cry1 and Cry2), inducing their transcription. Per and Cry mRNA diffuse into the cytoplasm where they are translated into PER and CRY proteins by ribosomes. At night, PER and CRY proteins in association with casein kinase 1 and other proteins translocate back into the nucleus with the help of GAPVD1, a cytoplasmic trafficking factor, where they inhibit CLOCK:BMAL1 transcriptional activity and subsequently close the negative feedback loop [47]. Casein kinase 1 also mediates proteasome degradation of PER through phosphorylation, thus lifting the inhibition of BMAL1 and CLOCK and allowing a new cycle of positive transcriptional activity to commence [48]. The CLOCK:BMAL1 complex also drives the transcription of Rorα (retinoic acid receptor-related orphan receptor α) and Rev-Erbα (reverse erythroblastosis virus α) genes, which serve as activators and inhibitors of Bmal1 transcription respectively [45]. This second feedback loop in conjunction with the previously described loop is responsible for a robust 24-h oscillatory molecular circadian cycle.
Although the mammalian molecular clock was first identified in the SCN in the late 1990s, subsequently the presence of an operational molecular clock has been observed in many tissues (“peripheral oscillators”) throughout the body [1]. Importantly, however, the circadian system is organized in a hierarchal fashion, with the SCN of the anterior hypothalamus serving as the master clock that works to synchronize peripheral oscillators [49]. SCN lesioned rodents lack circadian activity rhythms, and implantation of new SCN neural grafts is sufficient to restore circadian expression with a period length relative to the new SCN [50]. Thus, the SCN is the central driver of overt circadian rhythms.
In mammals, the SCN is comprised of a dorsal shell of neurons that express the neuropeptide arginine vasopressin (AVP) surrounding a ventral core composed of vasoactive intestinal polypeptide (VIP) expressing neurons [51]. When placed in culture, in vitro SCN neurons maintained a robust 24-h rhythm of Per1 clock gene expression for more than 30 days [52]. Conversely, peripheral tissues that underwent the same treatment only retained Per1 rhythmicity for less than a week, suggesting a loss of synchronization in the absence of rhythmic signals from the SCN. Furthermore, when rats experienced a 6-h delay or advance in their light:dark (LD) 12 h:12 h cycle, their SCN neurons in vitro shifted rapidly in response to environmental light, while peripheral cells had a less adaptive response, indicating that these damped oscillators required an external circadian signal for both rhythmicity and adjustment [52]. These peripheral oscillators are subordinate to the SCN and have been demonstrated in vivo in many tissues including various brain regions [53,54,55]. It is noteworthy that the phase of oscillatory clock gene expression can vary between different peripheral oscillators and the SCN [56,57,58]. Therefore, one role of peripheral oscillators is likely to transduce a daily environmentally entrained timing signal from the SCN into the appropriate molecular oscillatory phase for optimal daily function of each tissue [59].
Light is the strongest zeitgeber (“time giver”) of the circadian system. Because the average SCN oscillation in humans is greater than 24 h (approximately 24.15 h), a daily reset is necessary to prevent the endogenous oscillation from drifting too far from the environmental light:dark cycle [60]. Through the retinohypothalamic tract (RHT), a neuronal pathway from the retina to the SCN, the SCN receives photic information that enables it to entrain to the light-dark cycle. This neuronal pathway consists of a special subset of retinal ganglion cells (intrinsic photosensitive retinal ganglion cells) that are exquisitely sensitive to low light levels, especially in the blue portion of the visible spectrum [61]. The SCN transmits this perceived time-of-day information via neural and hormonal signals to peripheral clocks located in brain areas outside of the SCN, as well as to most organs including the pancreas, liver, kidney, and heart [1, 56, 62,63,64]. The coordination of the SCN with its peripheral oscillators results in necessary and time-of-day appropriate physiological and behavioral outcomes [65]. It should be noted, however, that RHT stimulation of the SCN in response to light at night triggers a phase shift in SCN oscillation as the SCN adjusts its timing to an unexpected environmental light:dark cycle stimulus [66]. This inappropriate phase adjustment provides a major source of circadian disruption to the body.
Circadian phase markers
The physiology of the SCN cannot be directly assessed in humans, and therefore studying output rhythms of the clock is necessary to determine the timing of the circadian rhythm. Measurable outputs, known as circadian phase markers, include melatonin, cortisol, and core body temperature. Timing of the internal clock is established through measurements of its period, the time necessary to complete a full oscillation, and phase, the state of an oscillation at a specific point in time [67].
Melatonin secretion is regulated by the SCN via a multi-synaptic pathway: from the SCN, neurons project to the superior cervical ganglion (SCG) and then to the pineal gland, which synthesizes and secretes melatonin [2]. Light exposure during the biological night results in rapid suppression of melatonin through this multi-synaptic pathway [68]. In adult humans, melatonin levels increase in the evening, peak during the middle of the night, and return to low levels in the morning where they remain throughout the day.
Cortisol is secreted from the adrenal glands in response to the pituitary gland releasing adrenocorticotropic hormone (ACTH). The circadian sensitivity of the adrenal gland to ACTH is regulated via a multi-synaptic neural output pathway from the SCN [2]. Cortisol secretion is lowest during the early night, begins to increase several hours before awakening, and peaks in the morning within 30–45 min after awakening. Of note, the cortisol awakening response (CAR) is a separate process that is superimposed on the underlying circadian peak of cortisol secretion [69]. It has been hypothesized that the magnitude of the morning CAR is associated with anticipation of stressors for the upcoming day [70].
Core body temperature peaks during the day and reaches its nadir in the middle of the night, which typically aligns with the sleep midpoint [71, 72]. Data from well-controlled studies indicate that the evening rise in melatonin prompts the nocturnal decrease in core body temperature through distal vasodilation and subsequent heat loss [73, 74].
There is growing support for the feasibility of measuring clock gene expression in human peripheral tissues as a useful circadian phase biomarker. Samples studied for these assessments include white blood cells, skin, oral mucosa, colon cells, adipose tissue, and post-mortem brain tissue [58]. Determining clock gene expression in peripheral tissues provides a method to monitor both central and peripheral clock regulation. Researchers are currently working towards validating gene expression from noninvasive skin samples as a biomarker of molecular clock phase to utilize in the rising field of circadian medicine [75].
Factors influencing circadian development in utero
Maternal SCN
The developing fetal circadian system relies solely on the maternal circadian system to set the environment context and relay time-of-day information. During pregnancy, maternal physiology changes to meet the needs of the developing fetus, and some of these adaptations are under circadian control. Pregnant women experience changes in circadian regulated plasma cortisol and melatonin concentration, core body temperature, and metabolism [76]. Additionally, women undergo an earlier shift in their chronotype, or their diurnal preference for sleep time, in the first and second trimesters as exhibited through an earlier sleep onset time [77].
Fetal SCN and peripheral oscillators
The fetal SCN and organs have been proposed to function as peripheral maternal circadian oscillators that are entrained through signals from the maternal SCN [78]. Similar to an orchestra, the maternal SCN serves as a conductor of the fetal clocks to maintain synchronization to the environment. This hierarchal arrangement produces temporal order similar to that found in the peripheral clocks of the adult human.
Circadian rhythmicity may depend on the maturity of the circadian system. Although clock gene expression is present in the SCN in embryonic mice, it appears that circadian system development is contingent on both the maturation of the central clock as well as the signaling pathways to its peripheral oscillators [79]. Dolatshad and colleagues [80] found that embryonic mouse tissues express circadian regulatory genes that appear rhythmic in vitro but lack the tissue level function to oscillate synchronously in vivo. In other words, the cells can synchronize and produce a tissue level rhythm, but for some reason do not. Animals born more developed may have more functional circadian system pathways, as presence of entrained 24-h rhythms in heart rate, respiratory movements, and hormones have been discovered in sheep, monkey, and human fetuses [81].
The human SCN begins developing in utero, and can be detected through histology by 40% gestation [82]. Additionally, the fetal SCN in late term squirrel monkeys exhibits oscillations of glucose utilization; suggesting it may be functional [83]. Because of limitations in human research, utilizing primate models is a powerful approach for determining the function and independence of the fetal SCN. In the SCN of capuchin monkeys at 90% gestation, robust oscillation of clock genes Bmal1 and Per2, and the MT1 melatonin receptor are apparent; however, the amplitude of these gene expression and receptor oscillations are smaller in the fetal primate SCN compared to the adult primate SCN. Additionally, fetal Bmal1 and Per2 oscillate in an antiphase pattern relative to the adult SCN similar to that found in the peripheral clocks of adult capuchin monkeys [84, 85]. It should be noted that in baboons, peripheral clock gene expression is not antiphasic to SCN clock gene expression; thus, this pattern may not be universal in primates [55]. Nonetheless, these blunted, antiphase oscillations in fetal capuchin monkeys suggest that the fetal SCN may be controlled by the maternal SCN. This was further demonstrated through suppression of primate maternal melatonin which yielded a change in the expression of clock genes in the fetal SCN that was restored through maternal melatonin replacement [84]. Taken together, existing research indicates that the maternal primate SCN communicates with the fetal primate SCN via zeitgebers, such as maternal melatonin, to produce rhythmicity.
Although the fetal SCN does not function independently from the maternal SCN, it may be important for receiving maternal signals and integrating them into physiological outputs. Lunshof and colleagues [86] tested this hypothesis by recording 24-h rhythms in fetal heart rate in a discordant anencephalic twin pregnancy, a disorder that results in an absent cerebellum and SCN. Results showed a significant diurnal rhythm in the mother as well as the intact twin fetus but not in the anencephalic fetus. These findings suggest that the fetal brain, and most likely the fetal SCN, are vital for the generation of circadian rhythms within the developing fetus.
In adult humans, rhesus monkeys, and mice, the adrenal gland serves as a peripheral clock that receives input from the SCN and in turn generates circadian rhythmicity of glucocorticoids [87,88,89]. Conversely, the fetal adrenal gland likely receives input from the maternal SCN. Torres-Farfan [84] and colleagues found that capuchin monkeys at 90% gestation exhibit circadian gene expression of Bmal1 and Per2, and the MT1 receptor. This temporal pattern of gene oscillation was identical to that of the fetal SCN, indicating that both the fetal adrenal gland and SCN are under similar control. In adult capuchin monkeys, there is a phase delay between peripheral oscillators and the SCN [85], and the absence of this delay between the fetal SCN and adrenal gland suggests that the maternal SCN, not the fetal SCN, regulates the fetal adrenal. However, there may be another integrator necessary to maintain rhythmicity of the fetal adrenal gland. In capuchin monkeys, suppression of the maternal SCN resulted in shifted fetal clock gene expression, but the fetal adrenal gland remained unaffected [84]. The maternal adrenal gland may account for lack of rhythmic change. In humans, suppression of maternal adrenal function with glucocorticoid treatment (triamcinolone) resulted in the disappearance of rhythms in fetal heart rate and limb movements. Rhythms were restored post-treatment, indicating fetal rhythm dependence on the maternal adrenal [90]. These data suggest that the fetal adrenal gland relies on input from both the maternal SCN and adrenal gland and remains independent from the fetal SCN. Further research is needed to examine the hierarchy of maternal signals to which the fetal adrenal entrains to.
Fetal circadian entrainment through melatonin
In utero, the fetus is reliant on time-of-day signals from maternal circadian cues (e.g., melatonin, cortisol, temperature), which serve as entrainers to the fetal circadian system and peripheral clocks. One zeitgeber necessary to the fetus is maternal melatonin, which crosses the placenta rapidly and unaltered to provide photoperiod information [91]. Melatonin receptors are present at 18 weeks of gestation in the fetal SCN and are widely distributed in fetal tissue [92]. The human fetus and neonate do not secrete melatonin, as synthesis begins post-natally [93] and are therefore dependent on maternal melatonin for circadian rhythmicity and development in the absence of external light-dark information. Results from Bellavia and colleagues [94] indicate that post-natal litter synchronization was not present among rat pups born to mothers that had undergone a ganglionectomy and pinealectomy, resulting in a nonfunctional SCN. Synchronization was restored when pregnant rats received melatonin in a diurnal fashion during the late period of gestation, indicating the importance of maternal melatonin in post-natal rhythms. Additionally, under constant maternal light conditions in utero and subsequent constant maternal melatonin suppression, newborn capuchin monkeys had twice as much plasma cortisol compared to those developed under normal light:dark conditions [95]. Those born to mothers who were exposed to chronic light but received daily melatonin replacement displayed normal cortisol levels, suggesting that maternal melatonin serves as an aid in the development of cortisol regulation in fetuses. Collectively, these studies indicate that maternal melatonin serves as a cue for developing fetal circadian rhythms and is important for healthy circadian outcomes.
A modern example of chronodisruption that pregnant women may face is shift work, which disrupts melatonin rhythms, such circadian misalignment also likely impacts fetal circadian development. Epidemiological data indicate that shift work during pregnancy is associated with increased spontaneous abortions, premature delivery, and low birth weight [96,97,98]. Data from one study in pregnant rats that simulated shift work found long terms effects on dams, including disrupted timing of corticosterone, leptin, glucose, insulin, free fatty acids, triglycerides, and cholesterol concentrations, as well as arrhythmic expression of circadian clock genes in the liver [99]. Further research on the longitudinal effects of gestational chronodisruption is necessary to understand how desynchronized signals during pregnancy impact fetal circadian system development.
Fetal circadian entrainment through time restricted feeding
Time restricted feeding during pregnancy can serve as a circadian cue to the developing fetus. SCN-lesioned rats produce offspring that display asynchronous litter drinking rhythms; however, restricting the pregnant rat’s feeding schedule to a 4-h window each day resulted in offspring that exhibited synchronization both within and between litters, indicating that prenatal restricted feeding can entrain fetuses [100]. Additionally, when offspring of SCN-lesioned, restricted rats were raised by lesioned, unrestricted mothers, similar synchronicity was observed, supporting the circadian effect of prenatal restricted feeding in utero. Similarly, Nováková et al. [101] found that maternal exposure to constant light during pregnancy, a condition which inhibits time-of-day cues, resulted in flattened circadian rhythmicity of Avp and c-fos gene expression in the SCN of rat pups. However, these rhythms developed normally when pregnant rats were restricted to a 6-h feeding window during their normal resting time. Surprisingly, pregnant rats that were exposed to a LD cycle and restricted feeding during their normal resting time reared pups that were not impacted despite only being able to eat during their rest time. This may be explained by findings from another study that observed maternal feeding during the light portion of a LD 12:12 light cycle entrained the fetal SCN to the maternal feeding time rather than the light schedule that the mother was entrained to, suggesting that the fetal SCN may not be exclusively controlled by the maternal SCN [102]. These studies indicate that rhythmic signals from the mother are necessary for fetal entrainment and there may be a hierarchy of redundant maternal mechanisms (e.g., melatonin, cortisol, food timing) that can serve as time-of-day cues to the developing fetus. However, the interaction and importance of these different maternal signals is not well understood and should be further investigated.
Factors influencing circadian development in infancy
Neonatal SCN
At birth, neonates lose direct communication with their mother’s SCN and must develop their own independent oscillating master circadian clock, undergoing a substantial change in function and reorganization of their circadian system. Infants are born with an immature circadian system that does not produce overt rhythms as evident by their absence of significant circadian rhythmicity in melatonin and cortisol rhythms and a stable sleep-wake cycle [39, 93, 103]. The neonatal SCN is not fully developed and contains only 13% of the adult number of AVP-expressing neurons and few VIP-expressing neurons—adult levels are attained by the first 2–3 years of life [82, 104]. During this sensitive developmental window, the SCN may be vulnerable to maternal and environmental influences that impact maturation of the circadian system.
Light
Light is the strongest environmental time cue to our internal clocks and exposing the developing clock to non-optimal lighting may have disruptive and long-lasting effects. In mice specifically, Ohta and colleagues [105] found that raising pups in constant light for their first 7 weeks of life resulted in disrupted circadian locomotor activities and cellular desynchrony exhibited through weak SCN clock gene oscillations. Conversely, the same researchers observed that mice raised in a LD 12:12 light cycle for their first 3 weeks of life and then exposed to continuous light conditions displayed orderly circadian rhythmicity for several weeks and were less vulnerable to disrupted circadian locomotor patterns [106]. Similarly, another study assigned mice to two different photoperiods (short-day, LD 8:16 or long-day, LD 16:8) during their first 3 weeks of life and then subjected each group to subsequent altered photoperiods for the next 4 weeks. Results indicated that compared to those raised under the short-day condition in very early life, those raised under the long-day condition exhibited a more stable molecular rhythm that tracked closely with the timing of dusk during later exposure to both long-day and short-day cycles [107]. Thus, the first few weeks of life may represent a sensitive window of circadian programming and organization where developing clocks are more susceptible to light exposure, and development during this time may set the stage for responsiveness to altered light cycles later in life.
Little is known about the impact of light on circadian entrainment in primate newborns. In one study, researchers exposed infant baboons to light at 22:30 for 50 min and observed increases in SCN metabolic activity and gene expression [108]. Data from this study also found that these newborns were able to entrain to a low intensity (200 lux) light:dark cycle. These findings indicate that the biological clock of premature infants is responsive to light via the RHT [109]. In humans, the RHT can be detected at 36-week gestation [110], indicating one developed pathway to the SCN by birth. In human neonates, constant dark conditions during the first days of life significantly increased melatonin plasma levels relative to a normal light:dark cycle [111], suggesting that the neonate is sensitive to light exposure post-birth and has the ability to establish connectivity between the SCN and the retina.
The preterm newborn represents a unique clinical context in which direct circadian signals are prematurely lost between the pregnant woman and fetus and replaced with environmental signals through the light-dark cycle. These newborns remain in the hospital for weeks or months where the light schedule is determined by medical personnel. Data from clinical studies indicate that exposing preterm infants to a light:dark cycle rather than constant dim light is associated with distinct patterns of rest and activity that appear 1 week after discharge, better growth rate, reduced fussing and crying, and a shorter hospital stay [112,113,114,115,116]. Additionally, a 24-h sleep-wake rhythm appeared earlier in preterm infants compared to term infants kept under the same cycled light, indicating that external time cues may play a vital role in sleep-wake rhythm development [117]. These studies provide evidence of light detection and entrainment in human preterm infants and highlight the importance of environmental signals for healthy development of the circadian system and sleep.
Circadian signals in breast milk
In addition to direct environmental circadian signals, infants maintain their connection to the maternal SCN by receiving maternal time cues through breast milk. Data from animal and human studies suggest that the diurnal fluctuations in breast milk evolved to send vital time-of-day information—a form of chrononutrition—to promote the development of an entrained circadian clock [118]. Human milk expressed during the day contains higher levels of cortisol [119], tyrosine [120], and immune factors [121], whereas night milk is comprised of increased levels of leptin [122], melatonin [123], and tryptophan [120]. Lodemore and colleagues [124] found that circadian rhythmicity in body temperature appeared earlier in breastfed infants compared to bottle fed infants, suggesting that breast milk may facilitate maturation of the clock. It is likely that infants reap the benefits when they drink circadian matched breast milk as these hormonal time signals may have evolved to support circadian system development.
Melatonin
Neonates rely on exogenous melatonin found in breast milk for their nightly hormonal circadian time cue. In humans, breast milk exhibits a congruent circadian melatonin pattern to that detectable in blood and saliva, and the concentration is approximately 35% of that found in the mother’s blood serum [123, 125]. From the maternal pineal, melatonin is secreted into the bloodstream and diffused into the mother’s breast milk [123, 125], where it crosses the infant’s intestinal barrier and circulates into tissues [126]. Here, it serves as a regulator of the infant circadian system. Compared to formula fed infants, breastfed infants show more regular nocturnal increases in 6-sulfatoxymelatonin, a metabolite of melatonin excreted in urine [127], higher sleep efficiency, less fragmented sleep [118], longer nocturnal sleep duration, and lower incidence of infant colic [125]. In the absence of breast milk, supplementation with exogenous melatonin may promote circadian clock development; however, this research remains unexplored. Infants given tryptophan-enriched formula, a melatonin metabolite precursor, displayed improvements in sleep and increases in urinary metabolites of serotonin from internal serotonin to melatonin synthesis [128]. Injection of melatonin in neonatal hamsters born to SCN-lesioned mothers resulted in entrained circadian rhythms of wheel-running activity on the day of weaning, PND21 [129]. Cumulatively, these data suggest that exogenous forms of melatonin serve as early circadian signals to the developing clock and are a necessary component to facilitate entrainment to the day/night cycle.
Cortisol
The circadian rhythm of cortisol in infants has been observed from as early as 2 weeks up to 9 months of age and is linked with the emergence of the sleep-wake cycle [103, 130,131,132]. The wide range in the cortisol rhythm appearance may be due to varying exposures of exogenous cortisol through breast milk. In humans, higher milk cortisol levels are associated with developmental outcomes including lower BMI and higher fear reactivity [133, 134]. Similar to maternal melatonin, cortisol in breast milk circulates from the maternal plasma and follows the same diurnal variation found in plasma and saliva [119, 135]. Human milk cortisol levels vary based on season and psychological distress [136, 137]. Data from rodent studies have established that ingested glucocorticoids diffuse from the neonatal intestines into the plasma and brain [138]. In adults, cortisol has been established as a major entrainment factor for peripheral clocks, indicating a pathway by which milk cortisol may regulate the infant peripheral clocks [59].
Summary and future directions
In the context of post-modern electric lighting environments, pregnant women and infants face daily challenges of circadian disruption, and the associated early developmental outcomes remain poorly understood. In a case study of an infant who was breastfed and was only exposed to natural light for the first 6 months of life, measurable outputs of circadian rhythmicity appeared more rapidly than previously reported—the temperature rhythm was observed at 1 week of age, rhythms of wake and melatonin secretion at sunset emerged at day 45, and nighttime sleep onset aligned with sunset by day 60 [139]. This finding suggests that infants growing in the natural world with no electric lighting results in more favorable circadian-related developmental outcomes. Longitudinal studies to determine how maternal and environmental signals interact with circadian system development, such as breast milk and the light/dark cycle (Fig. 1) are necessary to establish what circadian cues to prioritize in prenatal and post-natal maturation.
Research in the field of circadian rhythm development has centered on two developmental timepoints, the prenatal and post-natal periods. As previously discussed, rhythms in the prenatal stage are influenced by maternal signals while rhythms in the post-natal stage are guided by environmental signals that the infant is exposed to as well as a maternal connection via breast milk. Although these two periods are distinct, they should not be treated as unconnected—prenatal development likely sets the stage for healthy post-natal development. Few studies have followed women and fetuses from pregnancy through early development in post-natal life. Capitalizing on such longitudinal data would allow examination of how factors during pregnancy may impact later development.
Although the multi-dimensional construct of “sleep health” as measured by self-reports and actigraphy has received considerable attention in the past decade [140,141,142], “circadian health” is a construct that is presently not well-defined, especially in the context of development. In part, this may be due to the demands of reliable measurement of circadian outputs, such as cortisol, melatonin, and core body temperature, but also reflects less developed conceptualization of a full distribution of core behavioral and physiological circadian features from good to poor. The SCN is complex with multiple neuronal and glial cell types, as well as different subregions and outputs. Therefore, within the SCN there is likely not a single biomarker that would reflect good circadian health. Understanding of how the SCN communicates with peripheral clocks is rapidly advancing based upon animal and adult human data; however, a similar appreciation is limited across development. Indeed, a significant component of circadian health may be at the level of communication between the SCN and peripheral clocks through entrainment signals. Additionally, most circadian research centers on how circadian systems respond to perturbation rather than how to create optimal environments for good circadian health. Also, current studies focus on the time course of clock outputs to quantify circadian clock development; however, with few longitudinal studies available, it is difficult to determine if early rhythms are sensitive predictors of an adaptive system later in life. Other features indicating optimal circadian system maturation may include robustness of rhythms, adaptability of the clock to changing environments, and consistency of rhythms. Future developmental research should consider not only clock outputs but also such metrics in early life that may predict circadian health across the lifespan.
Pregnant women, mothers, and other infant caregivers serve as gatekeepers to external factors that influence circadian system development. Conventional behaviors in our post-modern world can result in conflicting temporal time cues for developing fetuses and infants. During pregnancy, a woman’s physiological signals set time-of-day cues for the fetus, and exposure to light at night results in a desynchronized circadian system that may disorganize the fetus’ circadian development and adaptation to environmental stimuli. Environmental lighting may also impact the developing infant through direct and indirect signals. For example, infants sleeping in a darkened room during daytime naps or exposed to light at night during feedings may experience a misperception of the time-of-day. Also, mothers breastfeeding at night may turn on an electric light or use a blue light-enriched device, which likely suppresses melatonin and other nighttime hormones found in breast milk, thus dampening maternal time cues to the infant’s circadian clock. Further, data suggest that infants who are fed formula do not benefit from the diurnal fluctuation of hormones present in breast milk. Additionally, women who pump may record the date but not the time-of-day that milk was expressed, which can result in infants receiving milk at a non-optimal circadian time. That is, if a woman feeds her infant milk in the evening that she previously pumped during the day, the infant will not receive nighttime hormonal time cues such as melatonin. The outcome of these mismatched signals and how they interact in early development and beyond remain unknown. One important question illustrating this type of interaction is if an infant is fed milk containing melatonin during the night but is exposed to artificial light. Which time cue is dominant? Or what if milk for a nighttime feeding originates from daytime expressed milk and the infant is also exposed to light—how do these factors synergize? Understanding such interactions will inform early prevention and intervention programs on modifiable factors that influence circadian system maturation, which could be cost effective and easily implemented. Based upon the above reviewed literature and the importance of the circadian system for health and development, future work should focus on the extent to which conflicting time cues influence early clock development and propose evidence-based parental care habits that will strengthen the daily environmental synchronization of the developing human.
Availability of data and materials
Not applicable
Abbreviations
- ACTH:
-
Adrenocorticotropic hormone
- AVP:
-
Arginine vasopressin
- Bmal1:
-
Brain and muscle Arnt-like factor
- CAR:
-
Cortisol awakening response
- Clock:
-
Circadian locomotor output cycle kaput
- GAPVD1:
-
GTPase activating protein and VPS9 domains 1
- LD:
-
Light:dark
- PND:
-
Post-natal day
- Rev-Erbα:
-
Reverse erythroblastosis virus α
- RHT:
-
Retinohypothalamic tract
- Rorα:
-
Retinoic acid receptor-related orphan receptor α
- SCG:
-
Superior cervical ganglion
- SCN:
-
Suprachiasmatic nucleus
- VIP:
-
Vasoactive intestinal polypeptide
References
Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418(6901):935–41.
Kalsbeek A, Palm IF, La Fleur SE, Scheer FAJL, Perreau-Lenz S, Ruiter M, et al. SCN outputs and the hypothalamic balance of life. J Biol Rhythm. 2006;21(6):458–69.
Hou Y, Liu L, Chen X, Li Q, Li J. Association between circadian disruption and diseases: a narrative review. Life Sci. 2020;262:118512.
Logan RW, McClung CA. Rhythms of life: circadian disruption and brain disorders across the lifespan. Nat Rev Neurosci. 2019;20(1):49–65.
Abbott SM, Malkani RG, Zee PC. Circadian disruption and human health: A bidirectional relationship. Eur J Neurosci. 2020;51(1):567–83.
Depner CM, Stothard ER, Wright KP. Metabolic consequences of sleep and circadian disorders. Curr Diabetes Rep. 2014;14(7):507.
Bedrosian TA, Fonken LK, Nelson RJ. Endocrine effects of circadian disruption. Annu Rev Physiol. 2016;78:109–31.
Chaves I, van der Eerden B, Boers R, Boers J, Streng AA, Ridwan Y, et al. Gestational jet lag predisposes to later-life skeletal and cardiac disease. Chronobiol Int. 2019.
Galdames HA, Torres-Farfan C, Spichiger C, Mendez N, Abarzua-Catalan L, Alonso-Vazquez P, et al. Impact of gestational chronodisruption on fetal cardiac genomics. J Mol Cell Cardiol. 2014;66:1–11.
Zhang P, Li G, Li H, Tan X, Cheng H-YM. Environmental perturbation of the circadian clock during pregnancy leads to transgenerational mood disorder-like behaviors in mice. Sci Rep. 2017;7(1):1–14.
Varcoe TJ, Wight N, Voultsios A, Salkeld MD, Kennaway DJ. Chronic phase shifts of the photoperiod throughout pregnancy programs glucose intolerance and insulin resistance in the rat. PLoS One. 2011;6(4):e18504.
Mendez N, Halabi D, Spichiger C, Salazar ER, Vergara K, Alonso-Vasquez P, et al. Gestational chronodisruption impairs circadian physiology in rat male offspring, increasing the risk of chronic disease. Endocrinology. 2016;157(12):4654–68.
Hiscock H, Wake M. Infant sleep problems and postnatal depression: a community-based study. Pediatrics. 2001;107(6):1317–22.
Martin J, Hiscock H, Hardy P, Davey B, Wake M. Adverse associations of infant and child sleep problems and parent health: an Australian population study. Pediatrics. 2007;119(5):947–55.
Byars KC, Yolton K, Rausch J, Lanphear B, Beebe DW. Prevalence, patterns, and persistence of sleep problems in the first 3 years of life. Pediatrics. 2012;129(2):e276–e84.
Thunström M. Severe sleep problems among infants in a normal population in Sweden: prevalence, severity and correlates. Acta Paediatr. 1999;88(12):1356–63.
Lam P, Hiscock H, Wake M. Outcomes of infant sleep problems: a longitudinal study of sleep, behavior, and maternal well-being. Pediatrics. 2003;111(3):e203–e7.
Lavigne JV, Arend R, Rosenbaum D, Smith A, Weissbluth M, Binns HJ, et al. Sleep and behavior problems among preschoolers. J Dev Behav Pediatr. 1999;20(3):164–9.
Yokomaku A, Misao K, Omoto F, Yamagishi R, Tanaka K, Takada K, et al. A study of the association between sleep habits and problematic behaviors in preschool children. Chronobiol Int. 2008;25(4):549–64.
Bruni O, Reto FL, Miano S, Ottaviano S. Daytime behavioral correlates of awakenings and bedtime resistance in preschool children. Suppl Clin Neurophysiol. 2000;53:358–61 Elsevier.
Goodnight JA, Bates JE, Staples AD, Pettit GS, Dodge KA. Temperamental resistance to control increases the association between sleep problems and externalizing behavior development. J Fam Psychol. 2007;21(1):39.
Hiscock H, Canterford L, Ukoumunne OC, Wake M. Adverse associations of sleep problems in Australian preschoolers: national population study. Pediatrics. 2007;119(1):86–93.
Ravid S, Afek I, Suraiya S, Shahar E, Pillar G. Kindergarten children's failure to qualify for first grade could result from sleep disturbances. J Child Neurol. 2009;24(7):816–22.
Touchette É, Petit D, Séguin JR, Boivin M, Tremblay RE, Montplaisir JY. Associations between sleep duration patterns and behavioral/cognitive functioning at school entry. Sleep. 2007;30(9):1213–9.
Meijer AM. Chronic sleep reduction, functioning at school and school achievement in preadolescents. J Sleep Res. 2008;17(4):395–405.
Hall WA, Moynihan M, Bhagat R, Wooldridge J. Relationships between parental sleep quality, fatigue, cognitions about infant sleep, and parental depression pre and post-intervention for infant behavioral sleep problems. BMC Pregnancy Childbirth. 2017;17(1).
Borbély AA. A two process model of sleep regulation. Hum Neurobiol. 1982;1(3):195–204.
Dijk D-J, Czeisler CA. Paradoxical timing of the circadian rhythm of sleep propensity serves to consolidate sleep and wakefulness in humans. Neurosci Lett. 1994;166(1):63–8.
Achermann P, Borbély AA. Mathematical models of sleep regulation. Front Biosci. 2003;8(6):1064.
Borb AA, Achermann P. Sleep homeostasis and models of sleep regulation. J Biol Rhythm. 1999;14(6):559–70.
Dubocovich ML. Melatonin receptors: role on sleep and circadian rhythm regulation. Sleep Med. 2007;8:34–42.
Torsvall L, Akerstedt T, Gillander K, Knutsson A. Sleep on the night shift: 24-hour EEG monitoring of spontaneous sleep/wake behavior. Psychophysiology. 1989;26(3):352–8.
Åkerstedt T. Shift work and disturbed sleep/wakefulness. Occup Med. 2003;53(2):89–94.
Åkerstedt T, Wright KP. Sleep loss and fatigue in shift work and shift work disorder. Sleep Med Clin. 2009;4(2):257–71.
Baron KG, Reid KJ. Circadian misalignment and health. Int Rev Psychiatry. 2014;26(2):139–54.
Kleitman N, Engelmann TG. Sleep characteristics of infants. J Appl Physiol. 1953;6(5):269–82.
Sheldon SH. Development of sleep in infants and children. Principles and Practice of Pediatric Sleep Medicine. 2nd ed. Philadelphia: Elsevier Inc; 2012. p. 17–23.
Anders TF, Keener M. Developmental course of nighttime sleep-wake patterns in full-term and premature infants during the first year of life. I. Sleep. 1985;8(3):173–92.
Jenni OG, Deboer T, Achermann P. Development of the 24-h rest-activity pattern in human infants. Infant Behav Dev. 2006;29(2):143–52.
Jenni OG, Borbély AA, Achermann P. Development of the nocturnal sleep electroencephalogram in human infants. Am J Phys Regul Integr Comp Phys. 2004;286(3):R528–R38.
Parmelee AH. Sleep patterns in infancy a study of one infant from birth to eight months of age. Acta Paediatr. 1959;48(2):160–70.
Iglowstein I, Jenni OG, Molinari L, Largo RH. Sleep duration from infancy to adolescence: reference values and generational trends. Pediatrics. 2003;111(2):302–7.
Jenni OG, LeBourgeois MK. Understanding sleep–wake behavior and sleep disorders in children: the value of a model. Curr Opin Psychiatry. 2006;19(3):282.
Edery I. Circadian rhythms in a nutshell. Physiol Genomics. 2000;3(2):59–74.
Buhr ED, Takahashi JS. Molecular Components of the Mammalian Circadian Clock. Circadian Clocks. Berlin Heidelberg: Springer; 2013. p. 3–27.
Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum Mol Genet. 2006;15(suppl_2):R271–R7.
Aryal RP, Kwak PB, Tamayo AG, Gebert M, Chiu P-L, Walz T, et al. Macromolecular assemblies of the mammalian circadian clock. Mol Cell. 2017;67(5):770–82 e6.
Meng Q-J, Logunova L, Maywood ES, Gallego M, Lebiecki J, Brown TM, et al. Setting clock speed in mammals: the CK1ɛ tau mutation in mice accelerates circadian pacemakers by selectively destabilizing PERIOD proteins. Neuron. 2008;58(1):78–88.
Inouye S-I, Kawamura H. Persistence of circadian rhythmicity in a mammalian hypothalamic" island" containing the suprachiasmatic nucleus. Proc Natl Acad Sci. 1979;76(11):5962–6.
Ralph MR, Foster RG, Davis FC, Menaker M. Transplanted suprachiasmatic nucleus determines circadian period. Science. 1990;247(4945):975–8.
Abrahamson EE, Moore RY. Suprachiasmatic nucleus in the mouse: retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001;916(1-2):172–91.
Yamazaki S, Numano R, Abe M, Hida A, Takahashi R-i, Ueda M, et al. Resetting central and peripheral circadian oscillators in transgenic rats. Science. 2000;288(5466):682–5.
Balsalobre A, Brown SA, Marcacci L, Tronche F, Kellendonk C, Reichardt HM, et al. Resetting of circadian time in peripheral tissues by glucocorticoid signaling. Science. 2000;289(5488):2344–7.
Stokkan K-A, Yamazaki S, Tei H, Sakaki Y, Menaker M. Entrainment of the circadian clock in the liver by feeding. Science. 2001;291(5503):490–3.
Mure LS, Le HD, Benegiamo G, Chang MW, Rios L, Jillani N, et al. Diurnal transcriptome atlas of a primate across major neural and peripheral tissues. Science. 2018;359(6381):eaao0318.
Pezük P, Mohawk JA, Wang LA, Menaker M. Glucocorticoids as entraining signals for peripheral circadian oscillators. Endocrinology. 2012;153(10):4775–83.
Chun LE, Woodruff ER, Morton S, Hinds LR, Spencer RL. Variations in phase and amplitude of rhythmic clock gene expression across prefrontal cortex, hippocampus, amygdala, and hypothalamic paraventricular and suprachiasmatic nuclei of male and female rats. J Biol Rhythm. 2015;30(5):417–36.
Cermakian N, Boivin D. The regulation of central and peripheral circadian clocks in humans. Obes Rev. 2009;10:25–36.
Spencer RL, Chun LE, Hartsock MJ, Woodruff ER. Glucocorticoid hormones are both a major circadian signal and major stress signal: How this shared signal contributes to a dynamic relationship between the circadian and stress systems. Front Neuroendocrinol. 2018;49:52–71.
Duffy JF, Cain SW, Chang A-M, Phillips AJ, Münch MY, Gronfier C, et al. Sex difference in the near-24-hour intrinsic period of the human circadian timing system. Proc Natl Acad Sci. 2011;108(Supplement 3):15602–8.
Hattar S, Liao H-W, Takao M, Berson DM, Yau K-W. Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science. 2002;295(5557):1065–70.
Oishi K, Sakamoto K, Okada T, Nagase T, Ishida N. Antiphase circadian expression between BMAL1 and period homologue mRNA in the suprachiasmatic nucleus and peripheral tissues of rats. Biochem Biophys Res Commun. 1998;253(2):199–203.
Oishi K, Sakamoto K, Okada T, Nagase T, Ishida N. Humoral signals mediate the circadian expression of rat period homologue (rPer2) mRNA in peripheral tissues. Neurosci Lett. 1998;256(2):117–9.
Yoo S-H, Yamazaki S, Lowrey PL, Shimomura K, Ko CH, Buhr ED, et al. PERIOD2: LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc Natl Acad Sci. 2004;101(15):5339–46.
Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010;330(6009):1349–54.
Boivin DB, Duffy JF, Kronauer RE, Czeisler CA. Dose-response relationships for resetting of human circadian clock by light. Nature. 1996;379(6565):540–2.
Duffy JF, Wright KP Jr. Entrainment of the human circadian system by light. J Biol Rhythm. 2005;20(4):326–38.
Macchi MM, Bruce JN. Human pineal physiology and functional significance of melatonin. Front Neuroendocrinol. 2004;25(3-4):177–95.
Wilhelm I, Born J, Kudielka BM, Schlotz W, Wüst S. Is the cortisol awakening rise a response to awakening? Psychoneuroendocrinology. 2007;32(4):358–66.
Fries E, Dettenborn L, Kirschbaum C. The cortisol awakening response (CAR): facts and future directions. Int J Psychophysiol. 2009;72(1):67–73.
Roenneberg T. What is chronotype? Sleep Biol Rhythms. 2012;10(2):75–6.
Refinetti R, Menaker M. The circadian rhythm of body temperature. Physiol Behav. 1992;51(3):613–37.
Cagnacci A, Kräuchi K, Wirz-Justice A, Volpe A. Homeostatic versus circadian effects of melatonin on core body temperature in humans. J Biol Rhythm. 1997;12(6):509–17.
Kräuchi K. How is the circadian rhythm of core body temperature regulated? Steinkopff Verlag; 2002.
Wu G, Ruben MD, Francey LJ, Smith DF, Sherrill JD, Oblong JE, et al. A population-based gene expression signature of molecular clock phase from a single epidermal sample. Genome Med. 2020;12(1):1–12.
McCarthy R, Jungheim ES, Fay JC, Bates K, Herzog ED, England SK. Riding the rhythm of melatonin through pregnancy to deliver on time. Front Endocrinol. 2019;10:616.
Martin-Fairey CA, Zhao P, Wan L, Roenneberg T, Fay J, Ma X, et al. Pregnancy induces an earlier chronotype in both mice and women. J Biol Rhythm. 2019;34(3):323–31.
Serón-Ferré M, Mendez N, Abarzua-Catalan L, Vilches N, Valenzuela FJ, Reynolds HE, et al. Circadian rhythms in the fetus. Mol Cell Endocrinol. 2012;349(1):68–75.
Honma S. Development of the mammalian circadian clock. Eur J Neurosci. 2020;51(1):182–93.
Dolatshad H, Cary AJ, Davis FC. Differential expression of the circadian clock in maternal and embryonic tissues of mice. PLoS One. 2010;5(3):e9855.
Seron-Ferre M, Valenzuela GJ, Torres-Farfan C. Circadian clocks during embryonic and fetal development. Birth Defects Res C Embryo Today. 2007;81(3):204–14.
Swaab D, Hofman M, Honnebier M. Development of vasopressin neurons in the human suprachiasmatic nucleus in relation to birth. Dev Brain Res. 1990;52(1-2):289–93.
Reppert SM, Schwartz WJ. Functional activity of the suprachiasmatic nuclei in the fetal primate. Neurosci Lett. 1984;46(2):145–9.
Torres-Farfan C, Rocco V, Monso C, Valenzuela F, Campino C, Germain A, et al. Maternal melatonin effects on clock gene expression in a nonhuman primate fetus. Endocrinology. 2006;147(10):4618–26.
Valenzuela F, Torres-Farfan C, Richter HG, Mendez N, Campino C, Torrealba F, et al. Clock gene expression in adult primate suprachiasmatic nuclei and adrenal: is the adrenal a peripheral clock responsive to melatonin? Endocrinology. 2008;149(4):1454–61.
Lunshof S, Boer K, van Hoffen G, Wolf H, Mirmiran M. The diurnal rhythm in fetal heart rate in a twin pregnancy with discordant anencephaly: comparison with three normal twin pregnancies. Early Hum Dev. 1997;48(1-2):47–57.
Engeland WC, Arnhold MM. Neural circuitry in the regulation of adrenal corticosterone rhythmicity. Endocrine. 2005;28(3):325–31.
Lemos DR, Downs JL, Urbanski HF. Twenty-four-hour rhythmic gene expression in the rhesus macaque adrenal gland. Mol Endocrinol. 2006;20(5):1164–76.
Oster H, Damerow S, Kiessling S, Jakubcakova V, Abraham D, Tian J, et al. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006;4(2):163–73.
Arduini D, Rizzo G, Parlati E, Giorlandino C, Valensise H, Del'Acqua S, et al. Modifications of ultradian and circadian rhythms of fetal heart rate after fetal-maternal adrenal gland suppression: A double blind study. Prenat Diagn. 1986;6(6):409–17.
Okatani Y, Okamoto K, Hayashi K, Wakatsuki A, Tamura S, Sagara Y. Maternal-fetal transfer of melatonin in pregnant women near term. J Pineal Res. 1998;25(3):129–34.
Reppert S, Weaver D, Rivkees S, Stopa E. Putative melatonin receptors in a human biological clock. Science. 1988;242(4875):78–81.
Kennaway DJ, Stamp GE, Goble FC. Development of melatonin production in infants and the impact of prematurity. J Clin Endocrinol Metab. 1992;75(2):367–9.
Bellavia S, Carpentieri A, Vaque A, Macchione A, Vermouth N. Pup circadian rhythm entrainment—effect of maternal ganglionectomy or pinealectomy. Physiol Behav. 2006;89(3):342–9.
Torres-Farfan C, Richter HG, Germain AM, Valenzuela GJ, Campino C, Rojas-García P, et al. Maternal melatonin selectively inhibits cortisol production in the primate fetal adrenal gland. J Physiol. 2004;554(3):841–56.
Zhu JL, Hjollund NH, Andersen A-MN, Olsen J. Shift work, job stress, and late fetal loss: The National Birth Cohort in Denmark. J Occup Environ Med. 2004;46(11):1144–9.
Aspholm R, Lindbohm M-L, Paakkulainen H, Taskinen H, Nurminen T, Tiitinen A. Spontaneous abortions among Finnish flight attendants. J Occup Environ Med. 1999;41(6):486–91.
Abeysena C, Jayawardana P, de A Seneviratne R. Maternal sleep deprivation is a risk factor for small for gestational age: a cohort study. Aust N Z J Obstet Gynaecol. 2009;49(4):382–7.
Varcoe TJ, Boden MJ, Voultsios A, Salkeld MD, Rattanatray L, Kennaway DJ. Characterisation of the maternal response to chronic phase shifts during gestation in the rat: implications for fetal metabolic programming. PLoS One. 2013;8(1):e53800.
Weaver DR, Reppert SM. Periodic feeding of SCN-lesioned pregnant rats entrains the fetal biological clock. Dev Brain Res. 1989;46(2):291–5.
Nováková M, Sládek M, Sumová A. Exposure of pregnant rats to restricted feeding schedule synchronizes the SCN clocks of their fetuses under constant light but not under a light-dark regime. J Biol Rhythm. 2010;25(5):350–60.
Ohta H, Xu S, Moriya T, Iigo M, Watanabe T, Nakahata N, et al. Maternal feeding controls fetal biological clock. PLoS One. 2008;3(7):e2601.
Ivars K, Nelson N, Theodorsson A, Theodorsson E, Ström JO, Mörelius E. Development of salivary cortisol circadian rhythm and reference intervals in full-term infants. PLoS One. 2015;10(6):e0129502.
Swaab D, Zhou J, Ehlhart T, Hofman M. Development of vasoactive intestinal polypeptide neurons in the human suprachiasmatic nucleus in relation to birth and sex. Dev Brain Res. 1994;79(2):249–59.
Ohta H, Mitchell AC, McMahon DG. Constant light disrupts the developing mouse biological clock. Pediatr Res. 2006;60(3):304–8.
Ohta H, Yamazaki S, McMahon DG. Constant light desynchronizes mammalian clock neurons. Nat Neurosci. 2005;8(3):267–9.
Ciarleglio CM, Axley JC, Strauss BR, Gamble KL, McMahon DG. Perinatal photoperiod imprints the circadian clock. Nat Neurosci. 2011;14(1):25–7.
Rivkees SA, Hofman PL, Fortman J. Newborn primate infants are entrained by low intensity lighting. Proc Natl Acad Sci. 1997;94(1):292–7.
Hao H, Rivkees SA. The biological clock of very premature primate infants is responsive to light. Proc Natl Acad Sci. 1999;96(5):2426–9.
Glotzbach S, Sollars P, Pagano M. Development of the human retinohypothalamic tract. Soc Neurosci. 1992.
Jaldo-Alba F, Munoz-Hoyos A, Molina-Carballo A, Molina-Font J, Acuna-Castroviejo D. Light deprivation increases plasma levels of melatonin during the first 72 h of life in human infants. Eur J Endocrinol. 1993;129(5):442–5.
Brandon DH, Holditch-Davis D, Belyea M. Preterm infants born at less than 31 weeks’ gestation have improved growth in cycled light compared with continuous near darkness. J Pediatr. 2002;140(2):192–9.
Rivkees SA, Mayes L, Jacobs H, Gross I. Rest-activity patterns of premature infants are regulated by cycled lighting. Pediatrics. 2004;113(4):833–9.
Watanabe S, Akiyama S, Hanita T, Li H, Nakagawa M, Kaneshi Y, et al. Designing artificial environments for preterm infants based on circadian studies on pregnant uterus. Front Endocrinol. 2013;4:113.
Vásquez-Ruiz S, Maya-Barrios JA, Torres-Narváez P, Vega-Martínez BR, Rojas-Granados A, Escobar C, et al. A light/dark cycle in the NICU accelerates body weight gain and shortens time to discharge in preterm infants. Early Hum Dev. 2014;90(9):535–40.
Guyer C, Huber R, Fontijn J, Bucher HU, Nicolai H, Werner H, et al. Cycled light exposure reduces fussing and crying in very preterm infants. Pediatrics. 2012;130(1):e145–e51.
Guyer C, Huber R, Fontijn J, Bucher HU, Nicolai H, Werner H, et al. Very preterm infants show earlier emergence of 24-hour sleep–wake rhythms compared to term infants. Early Hum Dev. 2015;91(1):37–42.
Cubero J, Narciso D, Terron P, Rial R, Esteban S, Rivero M, et al. Chrononutrition applied to formula milks to consolidate infants' sleep/wake cycle. Neuroendocrinol Lett. 2007;28(4):360–6.
Pundir S, Wall CR, Mitchell CJ, Thorstensen EB, Lai CT, Geddes DT, et al. Variation of human milk glucocorticoids over 24 hour period. J Mammary Gland Biol Neoplasia. 2017;22(1):85–92.
Sánchez CL, Cubero J, Sánchez J, Franco L, Rodríguez AB, Rivero M, et al. Evolution of the circadian profile of human milk amino acids during breastfeeding. J Appl Biomed. 2013;11(2):59–70.
Franca EL, Nicomedes TR, Calderon IMP, França ACH. Time-dependent alterations of soluble and cellular components in human milk. Biol Rhythm Res. 2010;41(5):333–47.
Cannon AM, Kakulas F, Hepworth AR, Lai CT, Hartmann PE, Geddes DT. The effects of leptin on breastfeeding behaviour. Int J Environ Res Public Health. 2015;12(10):12340–55.
Illnerova H, Buresova M, Presl J. Melatonin rhythm in human milk. J Clin Endocrinol Metab. 1993;77(3):838–41.
Lodemore M, Petersen S, Wailoo M. Factors affecting the development of night time temperature rhythms. Arch Dis Child. 1992;67(10):1259–61.
Cohen Engler A, Hadash A, Shehadeh N, Pillar G. Breastfeeding may improve nocturnal sleep and reduce infantile colic: potential role of breast milk melatonin. Eur J Pediatr. 2012;171(4):729–32.
Reppert SM, Klein DC. Transport of maternal [3H] melatonin to suckling rats and the fate of [3H] melatonin in the neonatal rat. Endocrinology. 1978;102(2):582–8.
Cubero J, Valero V, Sánchez J, Rivero M, Parvez H, Rodríguez A, et al. The circadian rhythm of tryptophan in breast milk affects the rhythms of 6-sulfatoxymelatonin and sleep in newborn. Neuroendocrinol Lett. 2005;26(6):657–62.
Aparicio S, Garau C, Esteban S, Nicolau MC, Rivero M, Rial RV. Chrononutrition: use of dissociated day/night infant milk formulas to improve the development of the wake–sleep rhythms. Effects Tryptophan. Nutr Neurosci. 2007;10(3-4):137–43.
Grosse J, Velickovic A, Davis FC. Entrainment of Syrian hamster circadian activity rhythms by neonatal melatonin injections. Am J Phys Regul Integr Comp Phys. 1996;270(3):R533–R40.
De Weerth C, Zijl RH, Buitelaar JK. Development of cortisol circadian rhythm in infancy. Early Hum Dev. 2003;73(1-2):39–52.
Santiago LB, Jorge SM, Moreira AC. Longitudinal evaluation of the development of salivary cortisol circadian rhythm in infancy. Clin Endocrinol. 1996;44(2):157–61.
Kiess W, Meidert A, Dressendörfer R, Schriever K, Kessler U, Köunig A, et al. Salivary cortisol levels throughout childhood and adolescence: relation with age, pubertal stage, and weight. Pediatr Res. 1995;37(4):502–6.
Nolvi S, Uusitupa HM, Bridgett DJ, Pesonen H, Aatsinki AK, Kataja EL, et al. Human milk cortisol concentration predicts experimentally induced infant fear reactivity: moderation by infant sex. Dev Sci. 2018;21(4):e12625.
Hahn-Holbrook J, Le TB, Chung A, Davis EP, Glynn LM. Cortisol in human milk predicts child BMI. Obesity. 2016;24(12):2471–4.
Hamosh M. Bioactive factors in human milk. Pediatr Clin N Am. 2001;48(1):69–86.
Lindberg M, Nolvi S, Härkönen J, Aatsinki AK, Karlsson L, Karlsson H, et al. Associations between maternal socioeconomic, psychosocial and seasonal factors, infant characteristics and human milk cortisol concentrations. Am J Hum Biol. 2021;33:e23561.
Kortesniemi M, Slupsky CM, Aatsinki A-K, Sinkkonen J, Karlsson L, Linderborg KM, et al. Human milk metabolome is associated with symptoms of maternal psychological distress and milk cortisol. Food Chem. 2021;356:129628.
Angelucci L. A model for later-life effects of perinatal drug exposure: maternal hormone mediation. Neurobehav Toxicol Teratol. 1985;7:511–7.
Mcgraw K, Hoffmann R, Harker C, Herman JH. The Development of Circadian Rhythms in a Human Infant. Sleep. 1999;22(3):303–10.
Chung J, Goodman M, Huang T, Bertisch S, Redline S. Multidimensional sleep health in a diverse, aging adult cohort: concepts, advances, and implications for research and intervention. Sleep Health. 2021;7:699–707.
Meltzer LJ, Williamson AA, Mindell JA. Pediatric sleep health: it matters, and so does how we define it. Sleep Med Rev. 2021;57:101425.
Buysse DJ. Sleep health: can we define it? Does it matter? Sleep. 2014;37(1):9–17.
Acknowledgements
Not applicable
Funding
Not applicable
Author information
Authors and Affiliations
Contributions
SDW contributed to the research, writing, and editing of this manuscript. MKL, RLS, KPW, CV, and LMH contributed to the editing of this manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Consent obtained for data collected is found in Fig. 2.
Competing interests
MKL reports receiving travel funds from the Australian Research Council and research support from the National Institutes of Health, beyond the submitted work. KPW reports research support/donated materials: DuPont Nutrition & Biosciences, Grain Processing Corporation, and Friesland Campina Innovation Centre, and being a consultant to and/or receiving personal fees from Circadian Therapeutics, Inc., Circadian Biotherapies, Inc., Philips, Inc, and U.S. Army Medical Research and Materiel Command-Walter Reed Army Institute of Research, outside the submitted work. CV reports being a consultant NIHM and having received research support from the National Institutes of Health outside the submitted work. RLS, SDW, and LMH declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wong, S.D., Wright, K.P., Spencer, R.L. et al. Development of the circadian system in early life: maternal and environmental factors. J Physiol Anthropol 41, 22 (2022). https://doi.org/10.1186/s40101-022-00294-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40101-022-00294-0