
Abstract
Although onion, Allium cepa L., production has been enhanced by the addition of organic matter to soil, other reports indicated no effect. In this study, onion plots (var. Sweet Sunrise) were treated with conventional fertilizer, composted sugar mill ash and vegetative yard waste, or fertilizer + compost. Measurements during two consecutive growing seasons in the Lower Rio Grande Valley of Texas were recorded on soil nutrients, nutrients in onion leaf tissue, leaf lengths, bulb yields in terms of numbers and weights in different size classes, and biochemical qualities. Our study demonstrates that the compost enriched soil fertility, particularly P, K, and Zn; percentage N in the onion leaf tissue was increased to the same extent as plants that received fertilizer; and leaves grew as long as in plots with fertilizer. At harvest, small cull bulbs were consistently the most prevalent in the control, and although compost had a positive influence on numbers and weights of marketable bulbs, fertilizer provided the highest yields during the first growing season. During the second season, however, marketable bulbs in composted plots were as abundant as in the plots receiving fertilizer. Treatment effects on thrips-induced leaf scarring were negligible. We conclude that the compost was useful for increasing marketable onion yield, and that effects improved as the compost decomposed over the two consecutive growing seasons.
Similar content being viewed by others

Introduction
Worldwide, onion, Allium cepa L., production occurs in at least 175 countries, with China, India, the United States, Turkey, and Pakistan having the most, yielding 97 billion kg/yr (FAOstat 20,019). In the United States, onion plantings comprise 52,400 ha yielding about 3.2 billion kg/yr valued at ≈US$1 billion (FAOstat 2019; NASS 2019). The onion crop in the United States is considered to be of high value and it is one of the most pesticide-intensive vegetable crops (NASS 2019) due to weed competition sensitivity (Bleasdale 1959; Wicks et al. 1973; Khokhar et al. 2010; Hatterman-Valenti 2012) and injury from diseases and arthropods (Adams et al. 2009; Culpepper 2009; Langstron 2009; Sell and Aakre 2009).
Onions are generally considered to be cool season plants (Boyhan et al. 2009; Sell and Aakre 2009; Voss and Mayberry 2009), and although onions require two seasons to complete the cycle from planted seed to seed production, they are cultivated for consumption as an annual (Voss and Mayberry 2009). Onions are planted as seed or transplanted as seedlings (Boyhan et al. 2009; Sell and Aakre 2009; Voss and Mayberry 2009). Transplants take 8–10 wk for bulbs to become harvestable (Boyhan et al. 2009), but harvest is often triggered when the necks weaken and the tops fall over (Sell and Aakre 2009; Voss and Mayberry 2009).
Conventional chemical fertilizers directly enhance crop yield because plants can assimilate the supplied nutrients directly or indirectly (Erana et al. 2019). Chemical fertilizers, however, can also have negative effects on agricultural ecosystems, such as degradation of soil (Jayathilake et al. 2006; Naeem et al. 2006), reduced soil microbiotic diversity, contamination of ground water, and pollution of the atmosphere (Aisha et al. 2007; Hernandez et al. 2010; Erana et al. 2019). Chemical fertilizers usually require replenishment every cultivation season because N, P, and K are rapidly lost through evaporation and leaching (Aisha et al. 2007), and continuous use of inorganic fertilizers often results in micronutirient deficiencies, imbalances of soil physiochemical properties, and unsustainable onion production (Jayathilake et al. 2006).
Composted materials are changed in the soil to organic fertilizer through biological processes, producing a stable source of nutrients from biological oxidative transformation (Ahmed and Varshney 2011; Deepesh et al. 2016). In many instances, composts are sources of stable, humus-like organic matter that can improve soil structure, increase soil moisture retention, reduce erosion, modify and stabilize soil pH, heighten cation exchange capacity, provision nutrients (especially N, P, and K), and stimulate microbial growth and activity in the soil (de Brito et al. 1995; Tejada and Gonzalez 2007; Hepperly et al. 2009; Mahmound et al. 2009; Diacono and Montemurro 2010; Xiao et al. 2010; Ahmed et al. 2013; Islam et al. 2017; Lee et al. 2018; Erana et al. 2019). Some composts can also suppress soil borne plant pathogens, including Allium white rot, Sclerotium cepivorum and Fusarium oxysporum f. sp. lycopersici, most likely by introducing or augmenting antagonistic microbes (Smolinska and Horbowicz 1999; Smolinska 2000; Coventry et al. 2002, 2005, 2006; Erana et al. 2019). While some studies did not find that compost amendments improved onion yield (Warman 2005; Rathod et al. 2009), others showed increases, especially when the compost was augmented with nitrogen (Maynard and Hill 2000a,b; Krishnamurthy 2005).
Onion thrips, Thrips tabaci Lindeman, cause serious yield losses to onions worldwide (Lewis 1997) by stunting the plant and reducing bulb weight from leaf feeding, predisposing plant sto bacterial and fungal pathogens, and transmitting iris yellow spot virus (Gill et al. 2015). Thrips pressure often occurs as outbreaks which are primarily controlled using insecticides (Morse and Hoddle 2006; Nault and Shelton 2010), leading to insecticide resistance in the pest (Shelton et al. 2003, 2006; Herron et al. 2008). The purpose of this study was to examine the effects of compost, with and without conventional fertilizer application, on plant nutrient levels in the soil, selected concentrations in the leaf tissue, infestations by onion thrips, Thrips tabaci Lindeman, and bulb quality.
Materials and methods
This study was conducted on 0.11 ha of a larger experimental field located at the USDA-ARS Kika de la Garza Subtropical Agricultural Research Center at Weslaco, Hidalgo County, Texas, for two growing seasons between Oct 2009 and Apr 2011. We used a randomized complete block experimental design. The experimental area was divided into six blocks, each of which was subdivided into four plots that were six rows (on 1 m row spacing) wide × 7.6-m long. The four plots within each block were randomly assigned to be a control without any compost or fertilizer, and two plots in each block received ≈605 kg of composted sugar mill mud, fly ash, and bottom ash from the mill fire box, and yard waste (Natural Solutions, Mission, Texas) rototilled into the top 30 cm of soil on 4 Mar 2009 (≈8 mo before the onions were planted). One of the two composted plots was, after planting, also treated with a conventional 28–0–5 (N-P-K) fertilizer application. The fourth plot received fertilizer only (fertilizer was applied by injection fertigation). One kg of the compost was sent to the Texas Plant and Soil Lab (Edinburg, Texas) for analysis of nutrients (N, P, K, Na, Mg, Zn, Fe, Mn, Cu, B, and S) and organic content, and the same quantity of the compost was sent to Soil Food Web (Corvallis, Oregon) for quantification of selected microbiota.
Onions (var. Sweet Sunrise) were double-row planted on top of the beds, each bed row 25 cm apart from adjacent bed rows, on 19 Oct 2009 and 20 Oct 2010, and fertilizer was applied on 14 Jan 2009 and 25 Jan 2011. Flood irrigation to furrows was provided on 20 and 30 Oct, 13 Nov, 10 Dec 2009, 8 and 25 Mar, 20 Oct, 1 and 22 Nov, 9 Dec 2010, 4, 12, and 31 Jan, 18 Feb, 4 Mar, 7 and 21 Apr, and 5 May 2011. Weeds were manually removed as needed, and no pesticides were applied for control of insects and pathogens. Because much of the previous year’s compost remained in the soil at the start of the second growing season, compost was not added, representing what would most likely occur in a compost-augmented system.
Six soil probe samples, each 15 cm deep, were collected early and later during each growing season at random spots from the rhizosphere area of rows and mixed from within each plot on 26 Oct 2009, 10 Mar, and 27 Nov 2010, and 15 Mar 2011. The samples were sent to the Texas A&M AgriLife Extension Service Soil, Water and Forage Testing Laboratory (College Station, Texas) for determining pH, conductivity, and amounts of nitrate, P, K, Ca, Mg, Na, S, Zn, Fe, Cu, Mn, and percentage organic matter. Six leaves from the uppermost three full expanded leaves, each leaf from a different onion plant, were collected from every plot on 25 Feb 2010 and 3 Mar 2011, oven dried at 38 °C for 48 h, then sent to the Texas A&M AgriLife Extension Service Soil, Water and Forage Testing Laboratory for quantifying percentage N, and amounts of P, K, Ca, Mg, Na, Zn, Fe, Cu, and Mn.
On 29 Dec 2009 and 18 Jan 2011, numbers of onion plants were counted on two 1-m-long sections of bed (included both rows of onion plants on top of each bed). Leaf lengths from five randomly selected onion plants per plot were recorded on 25 and 18 Jan 2010 and 2011, respectively. Percentages of leaves that had been injured by onion thrips was determined by examining all the green leaves on three onion plants per plot for feeding scars. Numbers of onion thrips (nymphs and adults) were counted using a hand lens on eight fully expanded leaves from each plot on 8 Dec 2009, 2 Apr and 11 Dec 2010, and 10 Apr 2010.
Harvest occurred on 6 May 2010and 19 May 2011. Onion bulbs were hand-pulled from two 7.6-m-long beds of the four inner beds in each plot, the leaves clipped off, and the bulbs were separated into diameter classes: small culls were < 5.1 cm, “mediums” were 5.1–7.6 cm, “large” bulbs were 7.7–10.2 cm, and “colossals” were > 10.2 cm. Five “large” onions from each plot, where possible, were analyzed for percentage moisture and brix, and pungency, measured as concentration of pyruvic acid, and soluble solids concentration were determined using the method of Randle and Bussard (1993). Twenty-five bulbs per plot were examined externally, and by slicing them in half, for signs of rot.
Treatment differences for all data sets were detected using both two-tailed analysis of variance (ANOVA) as a randomized block design, and as a factorial with compost and fertilizer used as treatment factors (Analytical Software 2008). Because insufficient numbers of large onions were produced in the control, factorial analysis was not conducted for bulb quality parameters. Normality and homogeneity of variance assumptions were not violated; hence, data were not transformed. Percentage data was arcsine-square root-transformed before analysis to conform to ANOVA assumptions. Differences between growing seasons were not analyzed because other, nontreatment, variables might have influenced production.
Results
Soil percentage organic matter, pH, and macronutrients
The compost was comprised of 26.3% organic matter, had a C:N ratio of 11.8:1, and quantities of each quantified element (Table 1). Microbial analysis of the same compost revealed that there were 53.7 and 5.01 μg/g of active bacteria and fungi, respectively. Soil analysis of samples collected on 26 Oct 2009, 8 mo after the compost was incorporated, showed that the compost-amended plots had greater amounts of organic matter (Table 2), and factorial analysis detected a positive compost effect on every sampling date (F ≥ 18.66, df = 1, 23, P ≤ 0.0076). All of the treatments, however, were less alkaline, with a mean pH of ≈7.9 ± 0.1, than the control at pH 8.4 ± 0.02 (F = 25.88, df = 3, 23, P < 0.0001), and factorial analysis indicated that pH was decreased by compost (F = 35.40, df = 1, 23 P = 0.0019) and by the fertilizer (F = 27.77, df = 1, 23, P = 0.0083), but a compost × fertilizer interaction was detected (F = 18.05, df = 1, 23, P = 0.0081). By March, pH in the control remained the same as in the previous autumn, but the pH in soil augmented with compost had risen to 8.2 ± 0.02; only the fertilizer and compost + fertilizer plots were still more acidic at 8.0 ± 0.1 and 7.8 ± 0.06, respectively (F = 16.21, df = 3, 23, P = 0.0001), and factorial analysis detected a negative fertilizer effect (F = 23.69, df = 1, 23, P = 0.0046). At the start of the second growing season, pH had declined in the control to 7.6 ± 0.07, but only the soil in the compost + fertilizer treatment was lower, at 7.3 ± 0.09 (F = 4.26, df = 3, 23, P = 0.0231) and factorial analysis did not indicate compost and fertilizer effects. By 15 Mar, pH had become greater in the control, 8.0 ± 0.04, than in the fertilizer and compost + fertilizer treatments, 7.6 ± 0.09 and 7.5 ± 0.03, respectively (F = 14.63, df = 3, 23, P = 0.0001). Factorial analysis showed that a fertilizer effect was linked to relatively low pH levels (F = 41.11, df = 1, 23, P = 0.0014).
Early during the first growing season, soil nitrate concentrations were ≥ 3.3-fold higher in the treatments than in the control (Table 2), and factorial analysis detected a positive compost effect (F = 45.80, df = 1, 23, P = 0.0011) and a compost × fertilizer interaction (F = 23.57, df = 1, 23, P = 0.0047). Late in the same growing season, however, the nitrate level was ≥ 3.2-fold greater in the fertilizer treatment than in the compost treatment and the control, but the compost + fertilizer treatment soil was 2.1-fold higher in nitrate than the fertilizer treatment (Table 2). Factorial analysis indicated that fertilizer was responsible for the heightened availability of nitrate (F = 20.94, df = 1, 23, P = 0.0061). At the start of the following growing season, nitrate concentrations were not affected by the treatments, but in March, nitrate levels were ≥ 9.5-fold higher in the two treatments where fertilizer had been applied (Table 2), confirmed by factorial analysis (F = 18.96, df = 1, 23, P = 0.0073).
Levels of P in the soil were ≥ 3.1-fold greater in the two treatments with compost than in the fertilizer treatment and the control on each of the sampling dates (Table 2), and factorial analysis detected a positive compost effect (F ≥ 31.88, df = 1, 23, P ≤ 0.0024). During the first growing season, K was ≥ 1.2-fold more abundant in the compost treatment than in any of the other treatments and the control, but during the second growing season the compost treatment had 1.2-fold more K than the fertilizer treatment only, and no differences were detected in Mar 2011 (Table 2). Factorial analysis, however, found a positive compost effect on all four sampling dates (F ≥ 14.91, df = 1, 23, P = 0.0120) and a negative fertilizer effect in Oct 2009 (F = 10.58, df = 1, 23, P = 0.0226).
Soil micronutrients
Treatment effects were not detected for Fe, but Zn was ≥ 1.5-fold more concentrated in the compost augmented plots than in the other plots (Table 2), confirmed by factorial analyses for each sampling date (F ≥ 25.68, df = 1, 23, P ≤ 0.0039). On the last sampling date, a positive fertilizer effect was also detected (F = 115.74, df = 1, 23, P = 0.0001). Treatment differences in abundances of Mg, Mn, and Cu were inconsistent over the sampling dates (Tables 2 and 3). Each of the treatments had ≥ 1.9-fold greater soil concentrations of S on the first sampling date, and, later in that growing season, S was 2.4-fold greater in the compost + fertilizer treatment than in the fertilizer treatment and the control; factorial analyses showed that compost increased levels of S (F ≥ 10.91, df = 1, 23, P ≤ 0.0214). Treatment effects were not detected during the second growing season (Table 2). Amounts of Na during the second growing season were ≥ 1.4-fold greater in plots without compost (Table 3), and factorial analysis detected a negative fertilizer effect on both sampling dates (F ≥ 41.39, df = 1, 23, P ≤ 0.0011).
Onion leaf growth and macronutrient concentrations
Plant density was the same (≈32/m row) among the control and treatments during the early part of each growing season. Leaf lengths during the first and second growing seasons were ≥ 1.4- and ≥ 1.2-fold longer in the treatment plots than in the control (F ≥ 25.67, df = 3, 23, P < 0.0001), respectively (Table 4), and factorial analysis detected a positive compost effect for both growing seasons (F ≥ 83.55, df = 1, 23, P = 0.0003). During each of the two growing seasons, percentage N in onion leaf tissue was ≥ 1.2-fold greater in the treatments compared with the control (Table 5), and factorial analysis consistently detected positive compost (F ≥ 30.19, df = 1, 23, P ≤ 0.0029) and fertilizer (F ≥ 37.52, df = 1, 23, P ≤ 0.0017) effects. P concentrations were ≥ 1.4-fold higher in the control during the first growing season than in the fertilizer and compost + fertilizer plots, but during the second growing season, P was ≥ 1.1-fold more concentrated in the fertilizer and the compost treatments than in the control (Table 5). Factorial analysis reflected the difference by detecting a compost × fertilizer interaction (F = 6.92, df = 1, 23, P = 0.0465) in the first growing season and a positive compost effect (F = 29.85, df = 1, 23, P = 0.0028) in the second growing season. Differences in amounts of K in onion leaf tissue were only found during the second growing season when treatments with compost had ≥ 1.3-fold more than in the control (Table 5), and factorial analysis showed a positive compost effect (F = 8.51, df = 1, 23, P = 0.0331). Ca was ≥ 1.2-fold more concentrated in onion leaf tissue collected from the control than in the treatments during the first growing season, and ≥ 1.1-fold more in the control and the compost + fertilizer treatment than in the other two treatments (Table 5). Factorial analysis of Ca concentrations in the first growing season showed a negative compost effect (F = 11.45, df = 1, 23, P = 0.0196) and a positive fertilizer effect (F = 9.03, df = 1, 23, P = 0.0171) with a compost × fertilizer interaction (F = 11.25, df = 1, 23, P = 0.0202). During the second growing season, factorial analysis again indicated a negative compost effect for Ca (F = 56.74, df = 1, 23, P = 0.0007). Treatment effects were not detected for Mg concentrations in onion leaf tissue during the first growing season, but Mg was ≥ 1.2-fold more abundant in the compost + fertilizer treatment than in the control during the second growing season (Table 5), and factorial analysis indicated a positive compost effect (F = 22.58, df = 1, 23, P = 0.0051).
Onion leaf micronutrient concentrations
Although Na was ≥ 1.3-fold more concentrated in the control than in the treatments during the first growing season, the compost + fertilizer treatment had 1.5-fold more than the compost treatment in the second growing season, and the compost treatment had ≥ 1.3-fold more than the fertilizer treatment and the control (Table 6). Factorial analysis detected a negative compost effect (F ≥ 8.05, df = 1, 23, P ≤ 0.0364) and a positive fertilizer effect (F ≥ 10.42, df = 1, 23, P ≤ 0.0232) during the two growing seasons, and during the second growing season a compost × fertilizer interaction (F = 14.14, df = 1, 23, P = 0.0132) was also found. While Zn was not affected by treatments in the first growing season, 1.2-fold more was found in leaves from the fertilizer treatment than from the other treatments and the control (Table 6), and factorial analysis showed a positive compost effect (F = 53.33, df = 1, 23, P = 0.0008) and a compost × fertilizer interaction (F = 20.83, df = 1, 23, P = 0.0060). Fe was 2.5-fold more concentrated in leaf tissues of the control than of the fertilizer treatment, which had ≥ 1.5-fold more than in the compost and the compost-fertilizer treatments during the first growing season (Table 6). During the second season, concentration of Fe was ≥ twofold higher in the control than in any of the treatments (Table 6). Factorial analysis detected a negative compost effect (F ≥ 6.90, df = 1, 23, P ≤ 0.0467) and a compost × fertilizer interaction (F ≥ 9.54, df = 1, 23, P ≤ 0.0272) during each growing season. No treatment differences in levels of Cu were found during the first growing season, despite a negative factorial compost effect (F = 8.27, df = 1, 23, P = 0.0348), but in the second season, Cu was ≥ 1.3-fold more concentrated in the leaf tissue in the compost treatment than in the other treatments and the control (Table 6), reflected by a negative fertilizer effect (F = 31.95, df = 1, 23, P = 0.0024) and a compost × fertilizer interaction (F = 7.06, df = 1, 23, P = 0.0450). Mn was 1.5-fold more concentrated in leaf tissue from the control than in the two compost-amended treatments during the first growing season, and in the second season Mn was ≥ 1.2-fold more abundant in the compost + fertilizer treatment than in any of the other treatments and the control (Table 6); factorial analysis showed a negative compost effect for both seasons (F ≥ 15.21, df = 1, 23, P ≤ 0.0114).
Thrips infestations
Early in each growing season (December), no onion thrips and thrips feeding scars were found on leaves, but nearing harvest (April), all of the leaves were scarred and thrips populations were not affected by treatments (the pooled average over both growing seasons was 10.4 ± 1.6 thrips per leaf). Late during the first growing season, ≥ 1.4-fold more green leaves per plant were found in the treatment plots (9.3 ± 0.1) than in the control (6.7 ± 0.1) (F = 220.08, df = 3, 23, P < 0.0001), but no treatment differences were detected in the second growing season (8.9 ± 0.6). Numbers of dry leaves were not affected by treatments in either year (200, 2.7 ± 0.1; 2011, 4.0 ± 2.5). Incidence of bulbs with rot was low (< 0.3%) regardless of the treatment.
Onion bulb harvest measurements
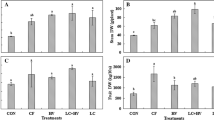
During the first growing season, numbers of small (cull) onions at harvest were ≥ 2.8-fold more abundant in the control than in any of the treatments (F = 66.50, df = 3, 23, P < 0.0001) (Fig. 1A), and factorial analysis showed that numbers of culls were negatively affected by compost (F = 81.12, df = 1, 23, P = 0.0003) and fertilizer (F = 23.69, df = 1, 23, P = 0.0046), and a compost × fertilizer interaction was detected (F = 32.47, df = 1, 23, P = 0.0023). Culls were ≥ twofold more numerous in the fertilizer treatment and in the control during the second growing season than in treatments that received compost (F = 23.30, df = 3, 23, P < 0.0001) (Fig. 1B), and factorial analysis detected a compost effect (F = 134.70, df = 1, 23, P < 0.0001). Medium-diameter onions in the first growing season were 1.6-fold more abundant in the fertilizer treatment than in the two compost-amended treatments, which were ≥2.2-fold more abundant than in the control (F = 19.05, df = 3, 23, P < 0.0001) (Fig. 1A), and during the second season the fertilizer treatment had ≥1.7-fold more than in the other treatments and the control (F = 22.02, df = 3, 23, P < 0.0001) (Fig. 1B). Although factorial analysis detected a positive compost effect in the first growing season (F = 18.49, df = 1, 23, P = 0.0077), compost had a negative effect on medium-diameter onions in the second season (F = 38.57, df = 1, 23, P = 0.0016). A positive fertilizer effect was detected for numbers of medium-diameter onions in both growing seasons (2010, F = 19.67, df = 1, 23, P = 0.0068; 2011, F = 68.67, df = 1, 23, P = 0.0004), and a compost × fertilizer interaction was detected in the second growing season (F = 15.56, df = 1, 23, P = 0.0109). Large-diameter onions were 1.9-fold more numerous in the two treatments with fertilizer than in the compost treatment, which had 43.3-fold more large onions than the control during the first growing season (Fig. 1A), and factorial analysis showed that there were positive compost (F = 206.02, df = 1, 23, P < 0.0001) and fertilizer (F = 8.26, df = 1, 23, P = 0.0342) effects and a compost × fertilizer interaction (F = 35.77, df = 1, 23, P = 0.0019). During the second growing season, there were ≥3.4-fold more large onions in the two treatments that had been augmented with compost than in the fertilizer treatment and the control (F = 9.70, df = 1, 23, P = 0.0008), confirmed by detection of a factorial analysis compost effect (F = 31.76, df = 1, 23, P = 0.0024) (Fig. 1B). Colossal onions were ≥2.5-fold more abundant during the first growing season in plots where fertilizer had been applied compared with the compost treatment, and the control produced no colossal onions (F = 42.01, df = 3, 23, P < 0.0001) (Fig. 1A), and factorial analysis detected compost (F = 126.53, df = 1, 23, P = 0.0001) and a compost × fertilizer interaction (F = 18.01, df = 1, 23, P = 0.0081). Numbers of marketable (medium, large, and colossal bulbs) during the first growing season were ≥ 1.7-fold more abundant in the treatments that received fertilizer than in the compost treatment, and the compost treatment produced 3.9-fold more marketable bulbs than the control (F = 100.09, df = 3, 23, P < 0.0001) (Fig. 1A), and factorial analysis detected compost (F = 47.63, df = 3, 23, P < 0.0001) and fertilizer (F = 153.29, df = 1, 23, P < 0.0001) effects. In 2011, marketable onions were ≥ 1.9-fold more numerous in the treatments than in the control (F = 7.79, df = 3, 23, P = 0.0023) (Fig. 1B).

Mean (± SE) numbers of small culled (< 5.1 cm diam), medium (5.1–7.6 cm diam), large (7.61–10.2 diam), colossal (> 10.2 cm diam), and total marketable (medium + large + colossal) onion harvested in plots receiving conventional fertilizer, compost, compost + conventional fertilizer, and no fertilizer or compost (control) at harvest in A) 2010 and B) 2011; lower-case letters indicate treatment differences (P < 0.05) within each size category (n = 6 replicates, two-way ANOVA, Tukey’s HSD)
Weight of discarded cull bulbs during the first growing season was highest in the control, moderate in the compost treatment, and lowest in the two treatments with fertilizer (F = 43,60, df = 3, 23, P < 0.0001) (Fig. 2A), but during the second growing season loss in kg to culls was highest in the fertilizer treatment and the control and lowest in plots amended with compost (F = 13.62, df = 3, 23, P = 0.0001) (Fig. 2B). Factorial analysis detected a negative compost effect on kg culls during both growing seasons (F ≥ 43.60, df = 1, 23, P < 0.0001). The fertilizer treatment producede ≥ than the control during each both seasons (Fig. 2A, B). Factorial analysis showed a positive compost effect on weight of medium bulbs in the first growing season (F = 57.42, df = 1, 23, P = 0.0006), but the effect was negative in the following season (F = 14.64, df = 1, 23, P = 0.0123). A positive fertilizer effect was detected for weights of medium-diameter bulbs in both years (F ≥ 39.85, df = 1, 23, P ≤ 0.0015). Large bulb production by weight was ≥ twofold greater in the two treatments that received fertilizer than in the compost treatment, and that treatment was 56.5-fold greater than the control (F = 78.76, df = 3, 23, P < 0.0001) (Fig. 2A) during the first growing season, and during the second, total large bulbs were 4.1-fold heavier in the compost + fertilizer treatment than in the fertilizer treatment, the compost treatment was intermediate between the two but 7.9-fold greater than the control (F = 6.53, df = 3, 23, P = 0.0048) (Fig. 2B). Factorial analysis indicated a positive compost effect on weights of large bulbs in both growing seasons (F ≥ 13.48, df = 1, 23, P = 0.0144) and a compost × fertilizer interaction in the first season (F = 28.81, df = 1, 23, P = 0.0030). The first-season fertilizer treatment yielded a 1.4-fold greater weight of colossal onions than the compost + fertilizer treatment, which a produced a 2.5-fold heavier crop of colossal inions than the compost treatment (F = 49.94, df = 3, 23, P < 0.0001), and no colossal onions were produced in the control (Fig. 2A). Weight of colossal onions during the second season, however, was ≥ 15.2-fold greater in the treatments with compost amendment than in the fertilizer treatment and the control (F = 4.13, df = 3, 23, P = 0.0326) (Fig. 2B). Factorial analysis detected a positive compost effect on colossal onions in both growing seasons (F ≥ 7.03, df = 1, 23, P ≤ 0.0461) and a positive fertilizer effect in the first season (F = 29.89, df = 1, 23, P = 0.0028). The weight of marketable onions in the treatments with fertilizer from the first growing season was twice than that of the compost treatment, and the compost treatment was 8.9-fold greater than the control (F = 151.49, df = 3, 23, P < 0.0001) (Fig. 2A). In the second growing season, the treatments produced ≥ 2.7-fold heavier yields of marketable onions than the control (F = 4.57, df = 3, 23, P = 0.0182) (Fig. 2B). Differences in percentage moisture and pyruvic acid concentration of first- and second-season bulbs were not detected between the treatments (Table 7).

Mean (± SE) weights of small culled (< 5.1 cm diam), medium (5.1–7.6 cm diam), large (7.61–10.2 diam), colossal (> 10.2 cm diam), and total marketable (medium + large + colossal) onion harvested in plots receiving conventional fertilizer, compost, compost + conventional fertilizer, and no fertilizer or compost (control) at harvest in A) 2010 and B) 2011; lower-case letters indicate treatment differences (P < 0.05) within each size category (n = 6 replicates, two-way ANOVA, Tukey’s HSD)
Discussion
Compost amended soils can have improved organic matter content, pH, and exchangeable K, Mg, Ca, Na, Fe, Zn, Cu, and other plant nutrients (Ouédraogo et al. 2001; Lee et al. 2012, 2018; Muktamar et al. 2018; Erana et al. 2019; Setyowati et al. 2021). Much of the nutritional enhancement of soil is attributable to increased microbial biomass and its action on releasing nutrients from the organic amendments (Vidali 2001; Suresh et al. 2004; Shaheen et al. 2007; Lee et al. 2018). The shallow (top 30 cm of soil) and sparsely branched onion root system, which makes the crop particularly susceptible to nutrient deficiencies, requires good fertilizer placement in the soil to produce a marketable crop (Greenwood et al. 1982; Brewster 1994, 2008). Heightened microbial activity in the onion rhizosphere, which facilitates nutrient release and root uptake, can help to mitigate problems with nutrient deficiencies (Vidali 2001).
The observed reduction of soil pH in treatments amended with compost was consistent with reports on the pH-stabilizing effects of other composts (Stamatiadis et al. 1999; Goyal et al. 2005; Erana et al. 2019), and it is likely that chemical changes in the soil brought about by plant and microbial activity in the onion rhizosphere affected pH. The decline in pH that accompanied soil amendment using compost likely resulted from the addition of hydrogen that is commonly associated with decomposition of organic matter and cation exchange (Baath et al. 1995; Bots and Benites 2005). Soil nitrate concentration was the most affected by nitrogen fertilizer application, although compost also caused changes, albeit to a lesser extent. Slow release of N from organic sources can result in N deficiencies in crops (Pang and Letey 2000), which might explain the lower levels of some nutrients in the leaves in our compost plots. Gaskell and Smith (2007) found that N availability from compost to the succeeding crop was variable. Rizk et al. (2014) suggested that the relatively gradual release of nutrients can be advantageous because they are less likely to be wasted through leaching, while ameliorating soil structure. Some researchers have also reported that compost augments N (Tejada and Gonzalez 2007; Hepperly et al. 2009; Diacono and Montemurro, 2010). Babajide et al. (2008) indicated that N, P, and K uptake was better where organic fertilizers were used compared to nonamended soils. Organic matter is associatated with release of mineralized nutrients for plant uptake (Shaheen et al. 2007; Paulin and O'Malley 2008; Guimaraes et al. 2013; Shedeed et al. 2014). Our compost increased levels of P, K, Zn, and occasionally other elements, while fertilizer had a minor influence on those elements in comparison. The generally heightened soil concentrations of macronutrients P and K in the compost treatment were not associated with heavier onion bulbs particularly during each growing season. This suggests that the nitrates supplied by the conventional fertilizer, which in the first growing season was associated with greater weights and numbers of marketable bulbs, were more available for plant uptake and utilization than in the compost treated plots. By the second growing season, numbers and weights of onion bulbs were more uniform than in the first season, a possible result of the compost applied in the first growing season releasing nitrates more slowly than conventional fertilizer, affecting the crop by the second growing season. Treatments did consistently affect soil concentrations of Ca, Mg, and S, but those three nutrients were not associated with differences in onion yield parameters. None of the soil micronutrients measured were associated with differences in marketable onion bulb abundances and weights. While compost does not necessarily affect soil electrical conductivity and concentrations of P and Mn (Erana et al. 2019), organic fertilizers release N at different rates; for example, 40% of total N in soil composted with chicken manure was available in the first year, with the remainder available in subsequent years at 6–12% per year (Whitmore 2007). For onion growing, more organic fertilizer is likely required compared to conventional fertilizers presumably because nutrients from organic fertilizers are less available due to relatively slow mineralization rates (Boyhan and Hill 2008).
Despite lower concentrations of soil N in the compost-only treatment than where fertilizer was applied, the onion plants accumulated N to the same extent as in the treatments that included fertilizer, permitting the same degree of leaf growth as plots enriched with fertilizer. Concentrations of other elements in leaf tissue did not consistently reflect soil concentrations, suggesting that biological processes, possibly stresses from differences in soil nutrition and soil water retention, resulted in changes to plant chemistry (Showler 2002; Reay-Jones et al. 2005; Showler et al. 2007; Showler and Castro 2010). Soil-incorporated composts are known to improve water retention of soils (Liebig and Doran 1999; McLaughlin and Kszos 2005; Triberti et al. 2008; Rizk et al. 2014; Lee et al. 2018; Erana et al. 2019), which could have had a role in accumulations of nutrients in the leaves.
Leaf nutritional quality, particularly regarding N, in some host crops can affect oviposition preference and injury by insect pests (Mattson and Haack 1987; Reay-Jones et al. 2005; Showler and Castro 2010; Showler 2012). This relationship, however, was not observed for onion thrips populations and the amount of associated leaf scarring. Our results indicate that the differences in soil and onion leaf nutrients did not influence thrips populations and the amount of leaf scarring, nor did they influence percentage bulb moisture and concentrations of pyruvic acid.
Despite composting onions with a variety of organic materials (i.e., animal manure, food waste, straw, and yard waste), some researchers reported that bulb production did not increase (Warman 2005; Rathod et al. 2009). Sheep and poultry dung incorporated into field plots was also ineffective, but covering the composted plots with black plastic was comparable to conventional fertilizer for increasing plant height, leaf number, pseudo-stem length and weight, and bulb diameter and pungency (Assi and Abu-Rayyan 2007). On the other hand, 48-d-old onion seedlings transplanted into plots where soil was amended with up to 160 kg/ha N or amended with up to 20 t/ha of composted livestock manure yielded ≈43% more than nonfertilized, or noncompost-augmented, control plots after 5 mo (Dixit 1997).
Although onion densities at the seedling stage in our study were not different between treatments in both growing seasons, treatment effects detected later in each of the two growing seasons were related to the size the bulbs attained. Numbers of culls in both growing seasons, and medium bulbs in the first season, in each treatment were inversely associated with the numbers of larger-diameter onions; hence, the control yielded relatively few marketable bulbs, their weight and economic value were lower than in any of the treatments. Although the fertilizer treatment produced more medium-diameter bulbs than the other treatments, large and colossal sizes were harvested in greater abundances in the compost-amended treatments during the second growing season relative to the first growing season. This change in terms of treatment effects on bulb diameter from the first to the second growing season might have been a result of organic matter’s amelioration of soil compaction, allowing onion bulbs to increase in diameter with less resistance from the enveloping soil. Zaharah et al. (1994) reported that increased onion yield in soils with increasing amounts of organic fertilizer was mainly due to bulb size. Soil fertility is not necessarily the main factor for optimum marketable onion yield (Mogren et al. 2009; Lee 2010; Yoldas et al. 2011; Saviello et al. 2013), suggesting that soil structure amenable to water retention and sufficient porosity to allow for bulb expansion play important roles. Soil-incorporated compost is known to improve soil structure (Ouédraogo et al. 2001; McLaughlin and Kszos 2005; Dauda et al. 2008; Triberti et al. 2008; Ahmed et al. 2013; Rizk et al. 2014; Islam et al. 2017; Lee et al. 2018).
Our findings show that, in terms of numbers and weights of marketable bulbs, fertilizer was superior to compost owing to greater nutrient availability in the rhizosphere (although the compost strongly increased concentrations of some nutrients), but in the second growing season compost was at least as effective as fertilizer. Because onions are sold by weight, it appears that fertilizer was essential to heightening yield in the first growing season where compost was applied, but with more growing seasons, we suggest that continued decomposition of the organic matter releases nutrients and reduces soil compaction, consequently improving onion yield. Another study demonstrated higher onion yields (bulb weight and percentage of marketable bulbs) after 3 yr where leaf litter was incorporated into the soil and augmented with conventional 10–10-10 N-P-K fertilizer, indicating that the leaf litter had to first compost and break down in the soil before improving production (Maynard and Hill 2000a, b). Because the treatments did not affect biochemical quality factors, the chief advantage of amending the soil with fertilizer and compost was to decrease culls while increasing marketable bulbs. It is likely that the degree of benefit obtained from our compost depends on the application rate and the constitution and condition of the soil. Fertilizer, on the other hand, was associated with high marketable yield regardless of whether the soil was made less compacted by the addition of organic matter. While compost had a positive influence on marketable onion bulb yield, it did not improve upon the fertilizer. Another study showed that composted Glyricidia green biomass and poultry manure (1:1 w/w) resulted in high levels of N, P, K, and S that, when augmented with 125 kg/ha N equivalent in urea, produced elevated yields of large and medium grade bulbs compared with conventionally fertilized soil (Krishnamurthy 2005). In our study, we determined that the compost enriched the soil with nutrients and reduced compaction, influencing plant growth and bulb yield across two growing seasons.
Compost and other sources of organic matter added to soil has been reported to increase onion bulb shoot weights, leaf numbers and lengths, and bulb weights and girths (Akoun 2004; Shaheen et al. 2007; Dina et al. 2010; Yoldas et al. 2011; Lee 2012; Lee et al. 2012, 2018; Assefa et al. 2015; Lema and Debrassa 2013; Yohannes et al. 2013; Rizk et al. 2014; Bua et al. 2017; Abou-El-Hassan et al. 2018; Ali et al. 2018; Erana et al. 2019; Petrovic et al. 2019). Some researchers found that organic manure improved onion nutrient uptake and and yields compared to conventional chemical fertilizers (Sharma et al. 2003; Shaheen et al. 2007; Yoldas et al. 2011). Asgele et al. (2018) indicated that vermicompost not only improved onion yield, it also speeded maturation.
Despite the positive effects of our compost on onions, different organic fertilizers can have variable quality and have different effects on crop productivity (Roe 1998; Gaskell and Smith 2007; Lee et al. 2018). Lee et al. (2018), for example, reported that composted cattle manure-fertilized onion bulb yield was 21.8% lower than in conventional systems but this result was not consistent. The array of different kinds of composted organic matter can supply widely differing amounts of nutrients and have varying rates of mineralization (Lee et al. 2012). In some instances, composted materials had no effect on pH, electrical conductivity, organic matter, and exchangeable cations (Lee et al. 2012), and some organic amendments, such as chicken litter, did not improve onion production (Boyhan et al. 2010). Further, excessive compost augmentation can negatively affect onion bulb yield (Boyhan et al. 2010; Vidigal et al. 2010; Lee 2012), and long-term manure application sometimes results in excessive P and K accumulations (Bary et al. 2000). Lee (2012), however, found that excessive applications of composted cattle manure did not detract from onion yields, and excessive amounts of pig manure resulted in onion yields that leveled off but were not detrimentally affected (Lee et al. 2012).
Use of organic manures can reduce the amount of conventional N-P-K fertilizers needed for onions (Selvakumari et al. (2001). Jayathilake et al. (2002) reported that integration of organic amendments with inorganic fertilizers resulted in better onion yields than inorganic fertilizers alone. Some researchers have suggested that onion production might be optimized by using selective combinations of organic and inorganic fertilizers (Seran et al. 2010). Because of the wide range of composted materials and other factors that can affect onion production, further research should be conducted to determine the best kinds of composts, as well as optimal combinations of organic and inorganic fertilizers.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
References
Abou-El-Hassan S, Elmehrat HG, Ragab AA, Megiud A, Abo-Dahab MS (2018) Growth, yield, quality and storability of some onion cultivars response to compost, vermicompost and plant growth promoting rhizobacteria. Middle East J Agric 7:292–306
Adams D, Riley D, Boyhan G (2009) Onion insects and their control, In: Onion production guide Coop Extension Service, University of Georgia Athens Georgia USA.
Ahmed JK, Varshney S (2011) Prolyactides chemistry, properties and green packaging technology. Int J Food Prop 14:37–58
Ahmed KA, Sharief AE, Fathalla HF (2013) Effect of organic and mineral fertilizers on vegetative growth, bulb yield and quality of onion cultivars. Crop Prod 2:91–100
Aisha AH, Rizk FA, Shaheen AM, Abdel-Mouty MM (2007) Onion plant growth, bulbs yield and its physical and chemical properties as affected by organic and natural fertilization. Res J Agric Biol Sci 3:380–388
Akoun J (2004) Effect of plant density and manure on the yield and yield components of the common onion (Allium cepa L.) var. nsukka red. Nigerian J Hort Sci 9:43–48
Ali M, Khan N, Khan A, Ullah R, Naeem A, Khan MW, Khan K, Farooq S, Rauf K (2018) Organic manures effect on the bulb production of onion cultivars under semiarid condition. Pure Appl Biol 7:1161–1170
Asgele K, Woldetsadik K, Gedamu F (2018) Effect of inorganic NP fertilizers and vermicompost on growth, seed yield and yield components of onion (Allium cepa L.) at Maitsebri, northern Ethiopia. J Hort Forest 10:89–96
Assefa AG, Mesgina SH, Abrha YW (2015) Response of onion (Allium cepa L.) growth and yield to different combinations of N, P, S, Zn fertilizers and compost in northern Ethiopia. Int J Sci Res 4:985–989
Assi N, Abu-Rayyan A (2007) Yield and quality of onion bulbs as affected by manure applications. Acta Hort 741:265–271
Baath E, Frostgard A, Pennanen T, Fritze H (1995) Microbial community structure and pH response in relation to soil organic matter quality in wood-ash fertilized, clear-cut or burned coniferous forest soils. Soil Biol Biochem 27:229–240
Babajide PA, Olabode OS, Akanbi WB, Olatunji OO, Ewetola EA (2008) Influence of composted Tithonia-biomass and N-mineral fertilizer on soil physicochemical properties and performance of tomato (Lycopersicon lycopersicum). Res J Agron 2:101–106
Bary AI, Cogger CG, Sullivan DM (2000) Fertilizing with manure. Washington State Univ Coop Ext, Pacific Northwest Ext Publ 533.
Bleasdale JKA (1959) The yield of onions and red beet as affected by weeds. J Hort Sci 34:7–13
Bots A, Benites J (2005) The importance of soil organic matter. Food Agric Organiz Soils Bull 80, Food and Agriculture Organization of the United Nations, Rome, Italy
Boyhan GE, Hill CR (2008) Organic fertility sources for the production of short-day organic onion transplants. Hort Technol 18:227–231
Boyhan GE, Hicks RJ, Torrance RL, Riner CM, Hill R (2010) Evaluation of poultry litter and organic fertilizer rate and source for production of organic short-day onions. HortTechnol 20:304–307
Boyhan GE, Granberry D, Kelley WT (2009) Transplant production. In: Boyhan GE, Kelley WT (eds) Onion production guide Coop Extension Service, University of Georgia, Athens, Georgia, USA, pp 4–5
Brewster JL (1994) Onions and other vegetable Allium, 1st edn. CABI, Wallingford
Brewster JL (2008) Onions and other vegetable alliums, 2nd edn. CABI, Wallingford
Bua B, Owiny R, Ocwa A (2017) Response of onion to different organic amendments in Central Uganda. J Agric Sci Technol 7:79–85
Coventry E, Noble R, Mead A, Whipps JM (2002) Control of Allium white rot (Sclerotium cepivorum) with composted onion waste. Soil Biol Biochem 34:1037–1045
Coventry E, Noble R, Mead A, Whipps JM (2005) Suppression of Allium white rot (Sclerotium cepivorum) in different soils using vegetable wastes. Eur J Plant Pathol 111:101–112
Coventry E, Noble R, Mead A, Marin FR, Perez JA, Whipps JM (2006) Allium white rot suppression with composts and Trichoderma viride in relation to sclerotia viability. Phytopathol 96:1009–1020
Culpepper AS (2009) Weed management in onion. In: Boyhan GE, Kelley WT (eds) Onion production guide. Coop Extension Service, University of Georgia, Athens, Georgia, USA, pp 32–38
Dauda SN, Ajayi FA, Ndor E (2008) Growth and yield of water melon (Citrullus lanatus) as affected by poultry manure application. J Agric Soc Sci 4:121–140
De Brito AMA, Gagne S, Antoun H (1995) Effect of compost on rhizosphere microflora of the tomato and on the incidence of plant growth-promoting rhizobacteria. Appl Environ Microbiol 61:194–199
Deepesh V, Verma VK, Suma K, Ajay S, Gnanavelu A, Madhusudanan M (2016) Evaluation of an organic soil amendment generated from municipal solid waste seeded with activated sewage sludge. J Mater Cycles Waste Manag 18:273–286
Diacono M, Montemurro F (2010) Long-term effects of organic amendments on soil fertility. Agron Sustain Devel 30:401–422
Dina MS, Shafeek MR, Abdallah MMF (2010) Effect of different nitrogen sources and soil solarization on green onion productivity for exportation. Ann Agric Sci Cairo Univ 55:97–106
Dixit SP (1997) Response of onion (Allium cepa) to nitrogen and farmyard manure in dry temperate high hills of Himachal Pradesh. Indian Agric Sci 67:222–223
Erana FG, Tenkegna TA, Asfaw SL (2019) Effect of agro industrial wastes compost on soil health and onion yields improvements: study at field condition. Int J Org Waste Agric 8:S5161-5171
FAOstat, Food and Agriculture Organization of the United Nations—agricultural data (2019) http://faostat.fao.org/site/408/DesktopDefalut.asp?PageID=408.20019
Gaskell M, Smith R (2007) Nitrogen sources for organic vegetable crops. HortTechnol 17:431–441
Gill HK, Garg H, Gill AK, Gillett-Kaufman JL, Nault BA (2015) Onion thrips (Thripidae: Thysanoptera) biology, ecology, and management in onion production systems. J Integr Pest Manag. https://doi.org/10.1093/jipm/pmv006
Goyal S, Dhull SK, Kapoor KK (2005) Chemical and biological changes during composting of different organic wastes and assessment of compost maturity. Bioresource Technol 96:1584–1591
Greenwood DJ, Gerwitz A, Stone DA, Barnes A (1982) Root development of vegetable crops. Plant Soil 68:75–96
Guimaraes DV, Gonzaga MIS, Silva TO, Silva TL, Dias NS, Matias MIS (2013) Soil organic matter pools and carbon fractions in soil under different land uses. Soil Tillage Res 126:177–182
Hatterman-Valenti H (2012) Managing weeds with reduced herbicide inputs: developing a novel system for onion. In: Alvarez-Fernandez R (ed) Herbicides—environmental impact studies and management approaches. In Techopen, London
Hepperly P, Lotter D, Ulsh CZ, Seidel R, Reider C (2009) Compost, manure and synthetic fertilizer influences crop yields, soil properties, nitrate leaching and crop nutrient content. Compost Sci Utiliz 17:117–126
Hernandez A, Catillo H, Ojeda D, Arras A, Lopez J, Sanchez E (2010) Effect of vermicompost and compost on lettuce production. Chilean J Agric Res 70:583–589
Herron G, James TM, Rophail J, Mo J (2008) Australian population of onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), are resistant to some insecticides used for their control. Austr J Entomol 47:361–364
Islam A, Islam S, Ayasha A, Rahman H, Nandwani D (2017) Effect of organic and inorganic fertilizers on soil properties and the growth, yield and quality of tomato in Mymensingh. Bangladesh Agric 7:18
Jayathilake PKS, Reddy IP, Srihari D, Neeraja G, Ravinder R (2002) Effect of nutrient management on growth, yield and yield attributes of rabi onion (Allium cepa L.). Veg Sci 29:184–185
Jayathilake PKS, Reddy LP, Srihari D, Reddy KR (2006) Productivity and soil fertility status as influenced by integrated use of N-fixing biofertilizers, organic manures and inorganic fertilizers in onion. J Agric Sci 2:46–58
Khokhar KM, Mahmood T, Hussain SI (2010) Weed management in onion using chemical, nonchemical, and manual methods. J Hort Sci Technol 85:473–476
Krishnamurthy DS (2005) Effect of sole and integrated use of improved composts and NPKfertilizers on the quality, productivity and shelflife of Bangalore rose red onion (Allium depa L). Mysore J Agric Sci 39:47–53
Langston DB (2009) Diseases of Vidalia onion, In: Boyhan GE, Kelley WT (eds) Onion production guide. Coop Extension Service, University of Georgia, Athens, Georgia, pp 18–27
Lee J (2010) Effect of application methods of organic fertilizer on growth, soil chemical properties and microbial densities in organic bulb onion production. SciHortic 124:299–305
Lee J (2012) Evaluation of composted cattle manure rate on bulb onion grown with reduced rates of chemical fertilizer. HortTechnol 22:798–803
Lee J, Kim H, Lee S, Ro C (2012) Evaluation of composted pig manure and organic fertilizer for organic onion production in paddy soil. Kr J Hort Sci Technol 30:123–128
Lee J, Hwang S, Min B, Kim H, Kim J, Hong K, Lee S, Shim S, Boyhan GE (2018) Effect of compost and mixed oilseed cake application rates on soil chemical properties, plant growth, and yield of organic bulb onions. Hort Sci Technol 36:666–680
Lema A, Degebassa A (2013) Comparison of chemical fertilizer, fish offal’s fertilizer and manure applied to tomato and onion. Afr J Agric Res 8:274–278
Lewis T (1997) Thrips as crop pests. CAB International, New York
Liebig MA, Doran JW (1999) Impact of organic production practices on soil quality indicators. J Environ Qual 28:1601–1609
Mahmound E, Ibrahim M, Robin P, Akkal-Corfini N, El-Saka M (2009) Rice straw composting and its effect on soil properties. Compost Sci Utiliz 17:146–150
Mattson WJ, Haack RA (1987) The role of drought in outbreaks of plant-eating insects. BioSci 37:110–118
Maynard AA, Hill DE (2000a) Cumulative effect of leaf compost on yield and size distribution of onions. Compost Sci Utiliz 8:12–18
Maynard AA, Hill DE (2000b) Leaf compost suppresses disease, improves onion yields. Biocycle 41:69–71
McLaughlin SB, Kszos LA (2005) Development of switchgrass (Panicum virgatum) as a bioenergy feedstock in the United States. Biomass Bioenergy 28:515–535
Mogren LM, Caspersen S, Olsson ME, Gertsson UE (2009) Organically fertilized onions (Allium cepa L.): effects of the fertilizer placement method on quercetin content and soil nitrogen dynamics. J Agric Food Chem 56:361–367
Morse JG, Hoddle MS (2006) Invasion biology of thrips. Ann Rev Entomol 51:67–89
Muktamar Z, Adiprasetyo T, Sari Y, Suprato S, Setyowati N, Fahrurrozi F (2018) Residual effect of vermicompost on sweet corn growth and selected chemical properties of soil from different organic farming practices. Int J Agric Technol 14:1471–1482
Naeem M, Iqbal J, Baksh MAA (2006) Comparative study of inorganic fertilizers and organic manures on yield components of mungbean (Vigna radiate L.). J Agric Soc Sci 2:227–229
NASS (National Agricultural Statistics Service) (2019) Vegetables: 2019 annual summary. http://usda.mannlib.conrell.edu/usda/nass/VegeSumm-01-26-2019.pdf
Nault BA, Shelton AM (2010) Impact of insecticide efficacy on developing action thresholds for pest management: a case study of onion thrips (Thysanoptera: Thripidae) on onion. J Econ Entomol 103:1315–1326
Ouédraogo E, Mando A, Zombré N (2001) Use of compost to improve soil properties and crop productivity under low input agricultural system in West Africa. Agric Ecosyst Environ 84:259–266
Pang XP, Letey J (2000) Organic farming: challenge of timing nitrogen availability to crop nitrogen requirements. Soil Sci Soc Am J 64:247–253
Paulin B, O'Malley P (2008) Compost production and use in horticulture. West Austral Agri Authority, Western Australia, Perth, Bull. 4746
Petrovic B, Kopta T, Pokluda R (2019) Effect of biofertilizers on yield and morphological parameters of onion cultivars. Folia Hort 31:51–59
Randle WM, Bussard ML (1993) Streamlining Onion Pungency Analyses. Hortsci 28:60
Rathod PH, Patel JC, Shah MR, Jhala AJ (2009) Recycling gamma irradiated sewage sludge as fertilizer: a case study using onion (Allium cepa). Appl Soil Ecol 41:223–233
Reay-Jones FPF, Showler AT, Reagan TE, Legendre BL, Way MO (2005) Integrated tactics for managing the Mexican rice borer (Lepidoptera: Crambidae) in sugarcane. Environ Entomol 34:1558–1565
Rizk FA, Shaheen AM, Abd El-Samad EH, El-Labban TT (2014) Response of onion plants to organic fertilizer and foliar spraying of some micronutrients under sandy soil conditons. J Appl Sci Res 10:383–392
Roe NE (1998) Compost utilization for vegetable and fruit crops. HortSci 33:934–937
Saviello G, Iovieno P, Morelli R, Baldantoni D, Morra L, Alfani A (2013) Long-term compost amendment of a Mediterranean horticultural soil. Part 1: Relationships among biological and chemical soil quality and the yield of two crops. Proc RAMIRAN 2013, 15th Int Conf. Rural-Urban Symbiosis, Verseilles, France, 3–5 June 2013, S1.43 ref 11
Sell R, Aakre D (2009) Onions. http://www.ag.ndsu.edu/pubs/alt-ag/onions.html.
Selvakumari G, Santhi R, Natesan R, Sathiyabama K (2001) Soil test and vegetable crop response under integrated plant nutrition system for optimization of fertilizer doses. S Ind Hort 49:130–136
Seran TH, Srikrishnah S, Ahamd MMZ (2010) Effect of different levels of inorganic fertilizers and compost as basal application on the growth and yield of onion (Allium cepa L.). J Agric Sci 5:64–70
Setyowati M, Aryamo DN, Wilman B, Muktamar Z (2021) Growth and yield of onion as affected by mulch types and vermicompost dose. Adv Biol Sci Res 14:51–58
Shaheen A, Fatma M, Rizk A, Singer SM (2007) Growing onion plants without chemical fertilization. Res J Agr Biol Sci 3:95–104
Sharma RP, Datt N, Sharma PK (2003) Combined application of nitrogen, phosphorus, potassium and farmyard manure in onion under high hills, dry temperate conditions of North-Western Himalayas. Ind J Agric Sci 73:225–227
Shedeed SI, El-Sayed SAA, Bash DMA (2014) Effectiveness of bio-fertilizers with organic matter on the growth, yield and nutrient content of onion (Allium cepa L.) plants. Eur Int J Sci Technol 3:115–122
Shelton AM, Nault BA, Plate J, Zhao JZ (2003) Regional and temporal variation in susceptibility to λ-cyhalothrin in onion thrips, Thrips tabaci (Thysanoptera: Thripidae), in onion fields in New York. J Econ Entomol 96:1843–1848
Shelton AM, Zhao JZ, Nault BA, Plate J, Musser FR, Larentzaki E (2006) Patterns of insecticide resistance in onion thrips (Thysanoptera: Thripidae) in onion fields in New York. J Econ Entomol 99:1798–1804
Showler AT (2002) Effects of water deficit stress, shade, weed competition, and kaolin particle film on selected foliar free amino acid accumulations in cotton, Gossypium hirsutum L. J Chem Ecol 28:631–651
Showler AT (2012) Drought and arthropod pests of crops. In: Neves DF, Sanz JD (eds) Invited book chapter for droughts. Nova Science Publishers, New York
Showler AT, Castro BA (2010) Influence of drought stress on Mexican rice borer (Lepidoptera: Crambidae) oviposition preference in sugarcane. Crop Prot 29:415–421
Showler AT, Cavazos JO, Moran PJ (2007) Dynamics of free amino acids accumulations in cotton leaves measured on different time scales after irrigation. Subtrop Plant Sci 59:38–55
Smolinska U (2000) Survival of Sclerotium cepivorum sclerotia and Fusarium oxysporum chlamydospores in soil amended with cruciferous residues. J Phytopathol 148:343–349
Smolinska U, Horbowicz M (1999) Fungicidal activity of volatiles from selected cruciferous plants against resting propagules of soil-borne fungal pathogens. Phytopath z 147:119–124
Analytical Software (2008) Statistix for Windows, Analytical Software, Tallahassee, Florida, USA.
Stamatiadis S, Werner M, Buchanan M (1999) Field assessment of soil quality as affected by compost and fertilizer application in a broccoli field (San Benito County, California). Appl Soil Ecol 12:217–225
Suresh KD, Sneh G, Krishn KK, Mool CM (2004) Microbial biomass carbon and microbial activities of soils receiving chemical fertilizers and organic amendments. Arch Agron Soil Sci 50:641–647
Tejada M, Gonzalez JL (2007) Application of different organic wastes on soil properties and what yield. Agron J 99:1597–1606
Triberti L, Nastri A, Giordani G, Comellini F, Baldoni G, Toderi G (2008) Can mineral and organic fertilization help sequestrate carbon dioxide in cropland? Eur J Agr 29:13–20
Vidali M (2001) Bioremediation. an overview. Pure Appl Chem 73:1163–1172
Vidigal SM, Sediyama MAN, Pedrosa MW, Santos MR (2010) Onion yield in organic system using compost of swine manure. Hort Brasil 28:168–173
Voss RE, Mayberry KS (2009) Fresh-market bulb onion production in California University of California, Div Agric & Natural Resources, Davis, California, Publ 7242:4
Warman PR (2005) Soil fertility, yield and nutrient contents of vegetable crops after 12 years of compost or fertilizer amendments. Biol Agric Hort 23:85–96
Whitmore AP (2007) Determination of the mineralization of nitrogen from composted chicken manure as affected by temperature. Nutr Cycl Agroecosyst 77:225–232
Wicks GA, Johnson DN, Nuland DS, Kimbacher EJ (1973) Competition between annual weeds and sweet Spanish onions. Weed Sci 21:436–439
Xiao M, Reddi LN, Howard J (2010) Erosion control on roadside embankment using compost. Appl Eng Agric 26:97–106
Yohannes KW, Belew D, Debela A (2013) Effect of farmyard manure and nitrogen fertilizer rates on growth, yield and yield components of onion (Allium cepa L.) at Jimma Southwest Ethiopia. Asian J Plant Sci 12:228–234
Yoldas F, Ceylan S, Mordogan N, Esetlili BC (2011) Effect of organic and inorganic fertilizers on yield and mineral content of onion (Allium cepa L.). Afr J Biotechnol 10:11488–11492
Zaharah A, Vimala P, Zainab RS, Salbiah H (1994) Response of onion and shallot to organic fertilizer on Bris (Rudua Series) soil in Malaysia. Acta Hart 385:429–432
Acknowledgements
Thanks to Jaime O. Cavazos, Veronica Abrigo, José Morales, Andy Cruz, Jaime Luna, Martín Galvan, and Tony Bautista for technical assistance.
Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Funding
No funding was obtained for this study.
Author information
Authors and Affiliations
Contributions
Allan T. Showler planned, conducted, and analyzed the study, interpreted the results, and wrote the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
No conflicting interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Showler, A.T. Effects of compost on onion quality, yield, and thrips infestation. Environ Syst Res 11, 22 (2022). https://doi.org/10.1186/s40068-022-00268-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40068-022-00268-2