
Abstract
Background
Sepsis is one of the most common clinical diseases, which is characterized by a serious and uncontrollable inflammatory response. LPS-induced inflammation is a critical pathological event in sepsis, but the underlying mechanism has not yet been fully elucidated.
Methods
The animal model was established for two batches. In the first batch of experiments, Adult C57BL/6J mice were randomly divided into control group and LPS (5 mg/kg, i.p.)group . In the second batch of experiments, mice were randomly divided into control group, LPS group, and LPS+VX765(10 mg/kg, i.p., an inhibitor of NLRP3 inflammasome) group. After 24 hours, mice were anesthetized with isoflurane, blood and intestinal tissue were collected for tissue immunohistochemistry, Western blot analysis and ELISA assays.
Results
The C57BL/6J mice injected with LPS for twenty-four hours could exhibit severe inflammatory reaction including an increased IL-1β, IL-18 in serum and activation of NLRP3 inflammasome in intestine. The injection of VX765 could reverse these effects induced by LPS. These results indicated that the increased level of IL-1β and IL-18 in serum induced by LPS is related to the increased intestinal permeability and activation of NLRP3 inflammasome. In the second batch of experiments, results of western blot and immunohistochemistry showed that Slit2 and Robo4 were significant decreased in intestine of LPS group, while the expression of VEGF was significant increased. Meanwhile, the protein level of tight junction protein ZO-1, occludin, and claudin-5 were significantly lower than in control group, which could also be reversed by VX765 injection.
Conclusions
In this study, we revealed that Slit2-Robo4 signaling pathway and tight junction in intestine may be involved in LPS-induced inflammation in mice, which may account for the molecular mechanism of sepsis.
Similar content being viewed by others

Introduction
Sepsis is one of the most common clinical diseases in the emergency department and ICU, which is characterized by the high expression of various inflammatory mediators and cytokines caused by the invasion of pathogenic microorganisms into the human body [1,2,3]. Epidemiologic studies and clinical investigations have shown that lipopolysaccharide (LPS), the main component of cell wall of Gram-negative bacteria, is an important cause of sepsis induced by bacterial infection [4, 5]. Many medical documents showed that LPS can induce the synthesis of a large number of pro-inflammatory cytokines such as interleukin-1β (IL-1β), interleukin-18 (IL-18), tumor necrosis factor-α (TNF-α), and interferon-γ (IFN-γ). The excessive secretion of these cytokines, in turn, can further induce more serious and unregulated inflammatory reactions [6,7,8]. Although there are more and more experimental and clinical studies on sepsis, the pathogenesis of sepsis and its underlying molecular mechanism have not yet been fully elucidated up to now.
Studies have shown that inflammasomes are involved in the pathophysiological process of various inflammatory diseases [9,10,11]. At present, the most clearly studied inflammasome is nucleotide-binding oligomerization domain (NOD)‑like receptor protein 3 (NLRP3) inflammasome, which is composed of NLRP3, apoptosis‑associated speck‑like protein (ASC) and aspartate proteolysis. Aspartate proteolysis is a protein complex composed of cysteinyl aspartate‑specific proteases‑1(caspase‑1) protein which can cut pro‑interleukin1β (pro-IL-1β) and pro‑interleukin 18 (pro-IL-1β) into active IL‑1β and IL‑18 [12]. NLRP3 inflammasome complex can be activated by a broad spectrum of stimuli, including bacteria, virus, fungi, and components of dying cells [13,14,15]. The activation of NLRP3 inflammasome can produce a large number of inflammatory factors, which can influence the permeability of endothelial cells [16].
More and more evidence has indicated that the dynamic balance of tight junction proteins in endothelial cells is related to a variety of inflammatory mediators and innate immunity, even can coordinate the host response in sepsis [17, 18]. The barrier properties of vascular and intestinal endothelial cells are essential for maintaining homeostasis of the internal environment, impairment of which is closely related to the occurrence and/or progression of various diseases, including enteritis, ulcerative colitis, stroke, cancer, sepsis, and neurodegenerative diseases [19, 20]. It is well known that these barrier properties are related to the aggregation of various proteins on endothelial cells to form specific membrane domains, such as adhesion junctions, gap junctions, and tight junctions [21]. Tight junctions are composed of different proteins such as occludin, claudin family members and zonula occludins (ZO) family members, which form a selective molecular channel [22]. This channel usually only allows small ions and uncharged molecules to pass through, and the change of its permeability is often related to the occurrence of diseases.
Slit Homolog (Slit) and Roundabout Homolog (Robo) are evolutionarily conserved proteins, which are widely expressed in different tissues such as heart, lung, and intestine [23, 24]. Slit can be divided into three subtypes, Slit1–3, and which has four membrane receptors, Robo1–4. Recently, more and more literature indicated that secretory protein Slit2 and its receptor Robo4 play an important role in regulating the fluidity and permeability of endothelial cells [25, 26]. For instance, Slit2 bind to the receptors Robo1 and Robo4, and then induces a series of intracellular signaling events. Slit2 regulates angiogenesis and protects endothelial integrity during sepsis and HIV infection [27]. However, the regulation of Slit2–Robo4 signal pathway in inflammatory process is still unclear. Therefore, it is urgent to understand the role of LPS in regulating Slit2 and Robo4 expression and disease progression.
The aim of this study is to determine the underlying mechanism of LPS-induced inflammation in sepsis. This study was designed to test the hypothesis that Slit2–Robo4 signaling pathway and tight junction in intestine may be involved in LPS-induced inflammation in mice model of sepsis.
Material and methods
Animals
Adult C57BL/6 J male mice (weighing 18 ± 1 g) were purchased from Shanghai Laboratory Animal Co., Ltd (Shanghai, China) and housed in a standard SPF rodent laboratory with a 12:12 h light:dark cycle. The environmental parameters of all animals were as follows: controlled temperature (22 ± 2 °C), humidity (50 ± 10%), standard diet and water ad libitum.
Animal model of LPS-induced sepsis
The animal model was established for two batches. In the first batch of experiments, 16 mice were randomly divided into control (Con) group and LPS group (n = 8 in each group). The LPS group was intraperitoneally injected with 0.1 ml LPS solution (solvent: PBS, 5 mg/kg, i.p.), and the control (Con) group was injected with the same amount of 0.9% saline. After 24 h, mice were anesthetized with isoflurane, blood and intestinal tissue were collected for further study. In the second batch of experiments, 24 mice were randomly divided into control (Con) group, LPS group, and LPS + VX765 group (n = 8 in each group). The LPS group was intraperitoneally injected with 0.1 ml LPS solution (solvent: PBS, 5 mg/kg, i.p.), LPS + VX765 group were injected with 0.1 ml VX765 (solvent: DMSO, 10 mg/kg, i.p.) at the same time of LPS injection, and the control (Con) group was injected with the same amount of saline. All drug concentrations used in this study were confirmed based on literature and preliminary trials [28, 29]. After 24 h, mice were anesthetized with isoflurane, blood and intestinal tissue were collected for further study.
Tissue immunohistochemistry
The intestinal tissues in each group were cut and washed by cold PBS solution (4 ℃). All intestine first fixed with 10% formalin and then embedded in paraffin wax, for cutting into 3-mm sections. Deparaffinized tissue sections were stained with Slit2, Robo4, and VEGF antibodies (Abcam, CA, USA) at 1:200 and then observed under a light microscope.
Western blot analysis
Total protein was isolated from the intestine by homogenization in cell lysis buffer (Sangon Biotech Co., Ltd., Shanghai, China), and then centrifuged at 12,000 × g for 30 min at 4 ℃. The proteins (5 μg/μL) were separated by 10% SDS-PAGE in Tris–glycine–SDS buffer. Separated proteins were transferred to NC membranes which were blocked with 5% skim milk (Bright Dairy Co., Ltd, China) for one hour at room temperature and then incubated with a 1:500 dilution of the individual primary antibodies anti-Slit2 (ab134166), anti-Robo4 (ab180824), anti-VEGF (ab150375), anti-NLRP3 (ab263899), anti-ASC (ab2236), anti-Caspase1 (ab207802), anti-ZO1 (ab61357), anti-Claudin5 (ab131259), anti-Occludin (ab167161), and anti-β-actin (ab6276) (Abcam, CA, USA). The antibodies were diluted in 3% BSA in Tris-buffered saline containing 0.1% Tween 20 (TBST), and applied overnight at 4℃. Membranes were washed and incubated with HRP-conjugated secondary antibodies (Proteintech, USA). Protein bands were detected using the Western Blot Luminol Reagent (Bio-rad, USA).
ELISA assays of inflammatory cytokines
Firstly, blood samples were centrifuged at 1000 × g for 5 min, and then serum was collected for further study. Next, IL-1β and IL-18 were determined by ELISA kits according to the instructions of the product (R&D, USA).
Statistical analysis
The data were analyzed using one-way ANOVA followed by the Dunnett's multiple range test using SPSS 21.0. All data are expressed as means ± SEM, and in all group, differences were considered statistically significant at p < 0.05.
Results
LPS can induce inflammatory response mediated by NLRP3 inflammasome
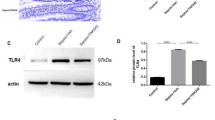
Firstly, in order to investigate the effects of LPS on C57BL/6 J mice, the LPS group mice were injected with LPS solution (5 mg/kg, i.p.), and the control (Con) group mice were injected with the same amount of saline. After 24 h, serum and intestinal tissue were collected for evaluating the expression of IL-1β, IL-18 and NLRP3 inflammasome by ELISA and western blot. ELISA results showed that LPS exposure for 24 h can induce significant high levels of IL-1β and IL-18 in serum when compared to Con group (Fig. 1A, B). Western blot results showed that the expression of NLRP3 and Caspase-1 protein increased significantly (Fig. 1C–F).

LPS-induced inflammation in serum and activation of NLRP3 inflammasome in intestine. IL-1β and IL-18 were detected by ELISA, NLRP3 inflammasome were detected by WB. All experiments were repeated twice. A IL-1β, B IL-18, C representative protein bands, D protein expression levels of NLRP3; E protein expression levels of ASC, F protein expression levels of Caspase-1. Data are presented as the mean ± SEM, n = 8 in each group. **P < 0.01 vs. control. Con control
LPS can increase the permeability of intestine and activation of Slit2–Robo4 signal pathway
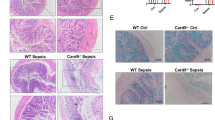
Many literatures show that the occurrence of sepsis is related to the change of intestinal permeability [30, 31]. Therefore, we wonder to explore whether LPS induces inflammation in C57BL/6 J mice is associated with intestinal permeability or not. The immunohistochemistry and western blot results in Figs. 2 and 3 show that the intestine of LPS groups displayed decreased expression of Slit2, Robo4 and higher expression of VEGF when compared with Con group. Western blot results showed that LPS can induce decreased expression of tight junction protein ZO-1, Occludin, and Claudin-5.

LPS-induced inhibition of Slit2–Robo4 signal pathway and activation of VEGF. Slit2, Robo4 and VEGF were detected by immunohistochemistry. All experiments were repeated twice

LPS-induced activation of Slit2–Robo4 signal pathway and disruption of tight junction. Slit2, Robo4, VEGF, ZO-1, Occludin, and Claudin-5 were detected by WB. All experiments were repeated twice. A, B Representative protein bands, C-H protein expression levels of Slit2, Robo4, VEGF, ZO-1, Occludin, and Claudin-5. Data are presented as the mean ± SEM, n = 8 in each group. **P < 0.01 vs. control. Con control
VX765 reversed LPS-induced inflammatory response mediated by NLRP3 inflammasome
In order to determine whether the increased IL-1β and IL-18 in serum induced by LPS injection is related to the activation of NLRP3 inflammasome, the second batch of experiments as described in Material and methods was carried out in this study. ELISA results showed that LPS-induced significant high level of IL-1β and IL-18 in serum can be reversed by VX765 when compared to LPS group (Fig. 4A, B). Western blot results showed that the expression of NLRP3 and Caspase-1 protein increased significantly after LPS injection for 24 h, which can also be reversed by VX765 (Fig. 4C–F).

VX765 reversed LPS-induced inflammation and activation of NLRP3 inflammasome. IL-1β and IL-18 were detected by ELISA, NLRP3 inflammasome were detected by WB. All experiments were repeated twice. A IL-1β, B IL-18, C representative protein bands, D protein expression levels of NLRP3, E protein expression levels of ASC, F protein expression levels of Caspase-1. Data are presented as the mean ± SEM, n = 8 in each group. **P < 0.01 vs. control; #P < 0.05, ##P < 0.01 vs. LPS. Con control
VX765 reversed the higher permeability of intestine and activation of Slit2–Robo4 signal pathway induced by LPS
In order to determine whether VX765 can reverse the permeability of intestine, western blot was used to examine the expression of Slit2, Robo4, VEGF, tight junction protein ZO-1, Occludin, and Claudin-5. The results in Fig. 5 showed that the intestine of LPS groups displayed decreased expression of Slit2, Robo4, tight junction protein ZO-1, Occludin, and Claudin-5 and higher expression of VEGF, all of which can be reversed by VX765 when compared to LPS group (Fig. 5).

VX765 reversed LPS-induced activation of Slit2–Robo4 signal pathway and disruption of tight junction. Slit2, Robo4, VEGF, ZO-1, Occludin, and Claudin-5 were detected by WB. All experiments were repeated twice. A, B Representative protein bands, C-H protein expression levels of Slit2, Robo4, VEGF, ZO-1, Occludin, and Claudin-5. Data are presented as the mean ± SEM, n = 8 in each group. **P < 0.01 vs. control; #P < 0.05, ##P < 0.01 vs. LPS. Con control
Discussion
Sepsis is a common, but poorly understood clinical syndrome with high motality in the world [32]. Although with the continuous progress of medical technology and updating of international guidelines, the death caused by sepsis is still increasing rapidly, which is because the pathogenesis has not been fully elucidated. Therefore, it is the common wish of researchers to elucidate the molecular mechanism of sepsis all over the world. In the present study, we showed that mice injected with LPS (5 mg/kg, i.p.) for 24 h could induce severe inflammatory reactions including an increased IL-1β, IL-18 in serum, activation of NLRP3 inflammasome, Slit2–Robo4 signaling pathway and disruption of tight junction in intestine. The injection of VX765 (10 mg/kg, i.p.), an inhibitor of NLRP3 inflammasome, could reverse these effects induced by LPS. Altogether, our findings revealed that Slit2–Robo4 signaling pathway and tight junction in intestinal probably involved in LPS-induced inflammation in mice, which may account for the molecular mechanism of sepsis.
A large number of researches have shown that barrier dysfunction and microvascular leakage of intestinal endothelial cells are the key factors of organ failure in sepsis and sepsis-related complications [33, 34]. The intestinal barrier consists of endothelial cells, adherens junctions, tight junctions, and extracellular components. Normally, the endothelial barrier is semi-permeable, which enables the transport of fluids and solutes from the blood to the tissues [35]. However, in patients with sepsis, abnormal barrier function leads to enhanced protein and solute transport. Breed et al. and other researchers have reported that the intestinal permeability of young C57BL/6 J female mice in cecal ligation and puncture group was higher than that of in the control group, which was related to the changes of claudins 1, 2, 3, 4, 5 and 8 [36, 37]. Wang et al. reported that the activation of autophagy and NLRP3 inflammasome by LPS + ATP can result in paracellular permeability increase and morphological disruption of both ZO-1 and Occludin [38]. In this study, tight junction protein ZO-1, Occludin, and Claudin-5 in the intestine of LPS group and Con group were examined by western blot. The results showed that LPS can induce decreased expression of tight junction protein ZO-1, Occludin, and Claudin-5. These effects can be reversed by VX765. All of these results demonstrated that inflammation induced by LPS is related to the increased permeability of intestine. However, this conclusion is preliminary, which also needs further experiments such as researches in gene knockdown animal model to prove it in the future.
Many studies have shown that endothelial cells specifically express receptor Robo4 which binding to its ligand Slit can inhibit inflammation and endothelial permeability by enhancing adhesion and adjusting cytoskeleton dynamics [39, 40]. Slit2 binds to the receptors Robo1 and Robo4, and then induces a series of intracellular signaling events. Slit2 regulates angiogenesis and protects endothelial integrity during sepsis and HIV infection [27]. In this study, Slit2 and Robo4 were examined by western blot in LPS group and Con group. The results showed that LPS can decrease the expression of Slit2 and Robo4, which is relative to the inflammation in sepsis. These effects also can be reversed by VX765. The mechanisms by which Slit2 regulates endothelial permeability are complex. The research results of Vincent et al. showed that Slit2N was the active fragment of Slit2, which could promote and accelerate the localization of VE cadherin and p120-catenin on the surface of endothelial cells [26, 41]. Therefore, Slit2N can reduce the permeability changes of endothelial cells induced by inflammatory mediators such as VEGF, LPS, TNF-α, and IL-1β [42, 43]. It was found that Slit2N reduced the accumulation of neutrophils and protein exudates in the alveolar cavity of LPS treated mice. But this effect is not present in Robo4 knockout mice, which proves the importance of this receptor [44]. In human umbilical vein endothelial cells (HUVECs), LPS downregulated Slit2, Robo4 and VE-cadherin protein expression and increased endothelial cell permeability in vitro during inflammation. Chen et, al. reported that hPMSC-derived Slit2 may inhibit LPS-induced CD11b, CD18 expression to decrease cell migration and increase adhesion through modulating the activity and motility of inflammatory macrophages in placenta [45, 46]. Vascular endothelial growth factor (VEGF), a kind of glycoprotein produced by endothelial cells, regulates vascular permeability through binding to VEGF receptors and activating the corresponding signal pathway [47]. Recently, some studies have focused on these effects of sepsis [48]. In this study, VEGF was examined, and the results showed that LPS can induce higher expression of VEGF which can be reversed by VX765. These results indicated that the increased permeability of the endothelium is associated with high expression of VEGF, but the underlying mechanisms need to be elucidated.
Although the experimental results of this paper are very interesting, we recognize the study still has some limitations. Firstly, this study did not carry out functional experiments to verify the established sepsis animal model, such as measuring heart rate, blood pressure and statistical mortality. Secondly, in the experimental design, the detection indexes that measured only after LPS injection for 24 h. The time point was single, and it would be more meaningful to examine the relevant detection indexes of animals after LPS injection at different time points, which can reflect the whole dynamic process of inflammatory factors with time. Third, there was no corresponding inhibitor and/or agonist group or used gene knockout technology to verify the Slit2–Robo4 signaling pathway. These deficiencies are interesting and worth for studying in the future by setting more experimental groups, time points and inflammatory factors and so on. What is more, Slit2 knockout mouse model could also be established, which can be used for further observing tight junction and other indicators by immunofluorescence or transmission electron microscope.
In conclusion, our study indicated that LPS could induce severe inflammatory reactions including an increased IL-1β, IL-18 in serum, activation of NLRP3 inflammasome, Slit2–Robo4 signal pathway and disruption of tight junction in intestine. VX765, an inhibitor of NLRP3 inflammasome, could reverse these effects induced by LPS. Altogether, our novel findings revealed that Slit2–Robo4 signaling pathway and tight junction in intestinal may be involved in LPS-induced inflammation in mice, which may account for the molecular mechanism of sepsis.
Availability of data and materials
All data generated or analyzed during this study are included in this published.
Abbreviations
- LPS:
-
Lipopolysaccharide
- IL-1β:
-
Interleukin-1β
- IL-18:
-
Interleukin-18
- TNF-α:
-
Tumor necrosis factor-α
- IFN-γ:
-
Interferon-γ
- NLRP3:
-
Nucleotide-binding oligomerization domain‑like receptor protein 3
- ASC:
-
Apoptosis‑associated speck‑like protein
- VEGF:
-
Vascular endothelial growth factor
References
Gunne S, Heinicke U, Parnham MJ, Laux V, Zacharowski K, von Knethen A. Nrf2-A molecular target for sepsis patients in critical care. Biomolecules. 2020;10(12):E1688.
Laura E, Andrew R, Waleed A, et al. Surviving sepsis campaign: international guidelines for management of sepsis and septic shock 2021. Crit Care Med. 2021;49(11):e1063–143.
Li J, Hongliang W. Autophagy-dependent ferroptosis in infectious disease. J Transl Int Med. 2023;11(4):355–62.
Rosadini CV, Kagan JC. Early innate immune responses to bacterial LPS. Curr Opin Immunol. 2017;44:14–9.
Cavaillon JM. Exotoxins and endotoxins: inducers of inflammatory cytokines. Toxicon. 2018;149:45–53.
Li Q, Wu C, Liu Z, Zhang H, Du Y, Liu Y, et al. Increased TLR4 expression aggravates sepsis by promoting IFN-gamma expression in CD38(-/-) mice. J Immunol Res. 2019;2019:3737890.
Liu L, Sun B. Neutrophil pyroptosis: new perspectives on sepsis. Cell Mol Life Sci. 2019;76(11):2031–42.
Kunze FA, Bauer M, Komuczki J, Lanzinger M, Gunasekera K, Hopp AK, et al. ARTD1 in myeloid cells controls the IL-12/18-IFN-gamma axis in a model of sterile sepsis, chronic bacterial infection, and cancer. J Immunol. 2019;202(5):1406–16.
Fan J, Xie K, Wang L, Zheng N, Yu X. Roles of inflammasomes in inflammatory kidney diseases. Mediators Inflamm. 2019;21(2019):2923072.
Zhong Z, Sanchez-Lopez E, Karin M. Autophagy, NLRP3 inflammasome and auto-inflammatory/immune diseases. Clin Exp Rheumatol. 2016;34(4 Suppl 98):12–6.
Guo H, Callaway JB, Ting JP. Inflammasomes: mechanism of action, role in disease, and therapeutics. Nat Med. 2015;21(7):677–87.
Jo EK, Kim JK, Shin DM, Sasakawa C. Molecular mechanisms regulating NLRP3 inflammasome activation. Cell Mol Immunol. 2016;13(2):148–59.
Guo HC, Jin Y, Zhi XY, Yan D, Sun SQ. NLRP3 inflammasome activation by viroporins of animal viruses. Viruses. 2015;7(7):3380–91.
Hayward JA, Mathur A, Ngo C, Man SM. Cytosolic recognition of microbes and pathogens: inflammasomes in action. Microbiol Mol Biol Rev. 2018;82(4):e00015-18.
Inserra A, Rogers GB, Licinio J, Wong ML. The microbiota-inflammasome hypothesis of major depression. BioEssays. 2018;40(9): e1800027.
Malavige GN, Ogg GS. Pathogenesis of vascular leak in dengue virus infection. Immunology. 2017;151(3):261–9.
Wang C, Li Q, Ren J. Microbiota-immune interaction in the pathogenesis of gut-derived infection. Front Immunol. 2019;7(10):1873.
Schnoor M, García Ponce A, Vadillo E, Pelayo R, Rossaint J, Zarbock A. Actin dynamics in the regulation of endothelial barrier functions and neutrophil recruitment during endotoxemia and sepsis. Cell Mol Life Sci. 2017;74(11):1985–97.
Haohao Z, Xueying W, Jing Z, Yixuan H, Xiumin Y, et al. Crosstalk between gut microbiota and gut resident macrophages in inflammatory bowel disease. J Transl Int Med. 2023;11(4):382–92.
Kim MH, Kim H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int J Mol Sci. 2017;18(5):1051.
Adu-Gyamfi EA, Czika A, Gorleku PN, Ullah A, Panhwar Z, Ruan LL, et al. The involvement of cell adhesion molecules, tight junctions, and gap junctions in human placentation. Reprod Sci. 2020 Nov 4.
Cai H, Liu W, Xue Y, Shang X, Liu J, Li Z, et al. Roundabout 4 regulates blood-tumor barrier permeability through the modulation of ZO-1, occludin, and claudin-5 expression. J Neuropathol Exp Neurol. 2015;74(1):25–37.
London NR, Zhu W, Bozza FA, Smith MC, Greif DM, Sorensen LK, et al. Targeting Robo4-dependent slit signaling to survive the cytokine storm in sepsis and influenza. Sci Transl Med. 2010;2(23):23ra19.
Jaouadi H, Jopling C, Bajolle F, et al. Expanding the phenome and variome of the ROBO-SLIT pathway in congenital heart defects: toward improving the genetic testing yield of CHD. J Transl Med. 2023;21(1):160–160160.
Bekes I, Haunerdinger V, Sauter R, Holzheu I, Janni W, Wöckel A, et al. Slit2/Robo4 signaling: potential role of a vegf-antagonist pathway to regulate luteal permeability. Geburtshilfe Frauenheilkd. 2017;77(1):73–80.
Weng J, Zhou X, Xie H, Gao Y, Wang Z, Gong Y. Slit2/Robo4 signaling pathway modulates endothelial hyper-permeability in a two-event in vitro model of transfusion-related acute lung injury. Blood Cells Mol Dis. 2019;76:7–12.
Anand AR, Nagaraja T, Ganju RK. A novel role for Slit2/Robo1 axis in modulating HIV-1 replication in T cells. AIDS. 2011;25(17):2105–11.
Chung K, Kim K, Choi Y, An H, Lee B, Kim D, et al. The critical role played by endotoxin-induced liver autophagy in the maintenance of lipid metabolism during sepsis. Autophagy. 2017;13(7):1113–29.
Yang X, Downey J, Cohen M, Housley N, Alvarez D, Audia J. The highly selective caspase-1 inhibitor VX-765 provides additive protection against myocardial infarction in rat hearts when combined with a platelet inhibitor. J Cardiovasc Pharmacol Ther. 2017;22(6):574–8.
Yoseph BP, Klingensmith NJ, Liang Z, Breed ER, Burd EM, Mittal R, et al. Mechanisms of intestinal barrier dysfunction in sepsis. Shock. 2016;46(1):52–9.
Kumar M, Leon Coria A, Cornick S, Petri B, Mayengbam S, Jijon HB, et al. Increased intestinal permeability exacerbates sepsis through reduced hepatic SCD-1 activity and dysregulated iron recycling. Nat Commun. 2020;11(1):483.
Cohen J, Vincent JL, Adhikari NK, Machado FR, Angus DC, Calandra T, et al. Sepsis: a roadmap for future research. Lancet Infect Dis. 2015;15(5):581–614.
Sharawy N, Pavlovic D, Wendt M, Cerny V, Lehmann C. Evaluation of the effects of gender and estradiol treatment on the intestinal microcirculation during experimental sepsis. Microvasc Res. 2011;82(3):397–403.
Li Y, Zhou J, Qiu J, Huang Z, Wang W, Wu P, et al. Berberine reduces gut-vascular barrier permeability via modulation of ApoM/S1P pathway in a model of polymicrobial sepsis. Life Sci. 2020;15(261): 118460.
Liu H, Yu X, Yu S, Kou J. Molecular mechanisms in lipopolysaccharide-induced pulmonary endothelial barrier dysfunction. Int Immunopharmacol. 2015;29(2):937–46.
Cao Y, Chen Q, Wang Z, Yu T, Wu J, Jiang X, et al. PLK1 protects against sepsis-induced intestinal barrier dysfunction. Sci Rep. 2018;8(1):1055.
Lorentz CA, Liang Z, Meng M, Chen CW, Yoseph BP, Breed ER, et al. Myosin light chain kinase knockout improves gut barrier function and confers a survival advantage in polymicrobial sepsis. Mol Med. 2017;23:155–65.
Feng Y, Wang Y, Wang P, Huang Y, Wang F. Short-chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell Physiol Biochem. 2018;49(1):190–205.
Enomoto S, Mitsui K, Kawamura T, Iwanari H, Daigo K, Horiuchi K, et al. Suppression of Slit2/Robo1 mediated HUVEC migration by Robo4. Biochem Biophys Res Commun. 2016;469(4):797–802.
Gorbunova EE, Gavrilovskaya IN, Mackow ER. Slit2-Robo4 receptor responses inhibit ANDV directed permeability of human lung microvascular endothelial cells. Antiviral Res. 2013;99(2):108–12.
Garrett JP, Lowery AM, Adam AP, Kowalczyk AP, Vincent PA. Regulation of endothelial barrier function by p120-cateninVE-cadherin interaction. Mol Biol Cell. 2017;28(1):85–97.
Yu J, Zhang X, Kuzontkoski PM, Jiang S, Zhu W, Li DY, et al. Slit2N and Robo4 regulate lymphangiogenesis through the VEGF-C/VEGFR-3 pathway. Cell Commun Signal. 2014;7(12):25.
Jiang S, Du Y, Liu D, He J, Huang Y, Qin K, et al. Inhibitory effect of Slit2-N on VEGF165-induced proliferation of vascular endothelia via Slit2-N-Robo4-Akt pathway in choroidal neovascularization. Cell Cycle. 2019;18(11):1241–53.
Zhao H, Anand AR, Ganju RK. Slit2-Robo4 pathway modulates lipopolysaccharide-induced endothelial inflammation and its expression is dysregulated during endotoxemia. J Immunol. 2014;192(1):385–93.
Chen Q, Zhou X, Hou R, Zhou Z, Wang Z, Chen Y, Weng J, Xu J. Aminophylline modulates the permeability of endothelial cells via the Slit2-Robo4 pathway in lipopolysaccharide-induced inflammation. Exp Ther Med. 2021;22(3):1042.
Chen CP, Wang LK, Chen CY, Chen CY, Wu YH. Placental multipotent mesenchymal stromal cell-derived Slit2 may regulate macrophage motility during placental infection. Mol Hum Reprod. 2021;27(2):gaaa076.
Yu Y, Chen R, Sun Y, Pan Y, Tang W, Zhang S, et al. Manipulation of VEGF-induced angiogenesis by 2-N, 6-O-sulfated chitosan. Acta Biomater. 2018;15(71):510–21.
Besnier E, Brakenhielm E, Richard V, Tamion F. Does anti-VEGF bevacizumab improve survival in experimental sepsis? Crit Care. 2017;21(1):163.
Acknowledgements
No.
Funding
This work was supported by grants from the Medical New Star Culture Plan of Baoshan Branch of Renji Hospital (rbxxrc-2019–007).
Author information
Authors and Affiliations
Contributions
L. Wang and Y-T Chen carried out most of the experiments and analyzed the data. L-H Ma conceived and supervised the project, provided suggestion to the experiments, discussed the data and wrote the manuscript with contributions from H-H Yu. All authors contributed to editing the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal procedures were carried out in accordance with the guidelines for the use of laboratory animals published by the People's Republic of China Ministry of Health, with the approval of the Ethical Committee of Experimental Animals of Naval Medical University and Changzheng Hospital.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, L., Chen, Y., Wu, H. et al. Slit2–Robo4 signal pathway and tight junction in intestine mediate LPS-induced inflammation in mice. Eur J Med Res 29, 349 (2024). https://doi.org/10.1186/s40001-024-01894-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40001-024-01894-5