
Abstract
Aim
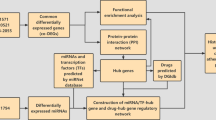
The study aimed to identify the underlying differentially expressed genes (DEGs) and mechanism of unstable atherosclerotic plaque using bioinformatics methods.
Methods
GSE120521, which includes four unstable samples and four stable atherosclerotic samples, was downloaded from the GEO database. DEGs were identified using LIMMA. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analyses of DEGs were performed using the Database for metascape Visualization online tool. Based on the STRING database, protein–protein interactions (PPIs) network among DEGs were constructed. Regulatory networks were visualized using Cytoscape. We use the xCell to analyze the different immune cell subtypes.
Results
A total of 1626 DEGs (1034 up-regulated and 592 down-regulated DEGs) were identified between unstable and stable samples. I pulled 62 transcription factors (34 up-regulated TFs and 28 down-regulated TFs) from the Trust database. The up-regulated TFs were mainly enrichment in positive regulation of myeloid leukocyte differentiation, and the down-regulated TFs were mainly enrichment in connective tissue development. In the PPI network, RB1, CEBPA, PPARG, BATF was the most significantly up-regulated gene in ruptured atherosclerotic samples. The immune cell composition enriched in CD cells and macrophages in the unstable carotid plaque.
Conclusions
Upregulated RB1, CEBPA, PPARG, BATF and down-regulated SRF, MYOCD, HEY2, GATA6 might perform critical promotional roles in atherosclerotic plaque rupture, furthermore, number and polarization of macrophages may play an important role in vulnerable plaques.
Similar content being viewed by others


Introduction
Cardiovascular and cerebrovascular diseases are the main causes of human death, in which AS is the culprit. Atherosclerosis (AS) is a chronic inflammatory disease which is characterized by the formation of medium- and large-sized atherosclerotic plaques on the vascular walls [1]. Moreover, of all atherosclerotic, carotid AS accounts for about 25%, easier to occur at the bifurcation of the carotid artery and is strongly associated with stroke [2]. Vulnerable plaque rupture, secondary thrombosis or embolism are major causes of stroke [3]. Pathology studies showed that vulnerable atherosclerotic plaques have a obvious morphology of thin-cap fibroatheromas, covering a large lipidic and necrotic core [4].
Activation of the immune system promotes the biological processes of vulnerable plaques [5]. Differences in cellular composition between vulnerable and stable plaques have been well established [6]. Macrophages are key players in cardiovascular disease (CVD) and have an important influence on the stability of atherosclerotic plaques.
In recent decades, the study found that transcription factors are key cellular components that control gene expression, many of them as “master regulators” exerting control over processes that specify cell types and developmental patterning and controlling specific pathways such as immune response [7]. For example, IRF5 is a major regulator of macrophage activation, regulates macrophage phenotypes and co-locates with CD11c + macrophages on the shoulder of plaques. It is associated with plaque vulnerability and symptoms of carotid endarterectomy in humans. In a mouse model of induced carotid artery plaque rupture, IRF5 drives plaque rupture [8]. In addition, researchers found that macrophage NCOR1 could block the pro-atherogenic effect of PPARc and suggest that stabilizing the NCOR1–PPARc binding could be a promising strategy to block the pro-atherogenic functions of plaque macrophages and lesion progression in atherosclerotic patients. Therefore, NCOR1 plays an important role in the regulation of atherosclerosis [9]. However, specific regulation mechanisms of transcription factors (TFs) and miRNAs involved in the progression of plaque stability have not been revealed.
In our study, we analyzed the regulatory role of a group of TFs in atherosclerotic plaques. The database obtained from GEO, we further explored the potential biological process of the dysregulated TFs and mRNAs relying on a variety of biological tools. Finally, we constructed the TFs–mRNA network and screened for potential hub genes to provide an important target for preventing the progression of atherosclerotic plaque.
Materials and methods
Subjects
Bioinformatics datasets
Expression profiling by high-throughput sequencing of mRNA GSE120521 was obtained from the platform Illumina HiSeq 2500 (Homo sapiens) of Gene Expression Omnibus database (GEO, https://www.ncbi.nlm.nih.gov/geo/).
Data preprocessing and identification of DEGs
The expression profile was preprocessed by Oligo package. After processed, DEGs between groups were screened out using the limma R package; p-value between gene expressions was determined using the T test and adjusted using Benjamini–Hochberg (BH) method. Those genes with the cut-off criteria of p-value < 0.05 and |log2fold change (FC)|> 0.58 were considered as DEGs.
Construction of TFs–mRNA network
We use TRRUST database-identified TFs, we find the TFs–mRNA functional relation by Spearman correlation, the screen condition is p value < 0.01, cor > 0.9. The visualized network was generated by Cytoscape software.
GO and KEGG enrichment analysis
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis of DEGs was performed with metascape (https://metascape.org/). GO analysis mainly reflects three aspects of gene information: biological process (BP), molecular function (MF), and cellular component (CC).
PPI network construction
The protein–protein interaction (PPI) network of DEGs were constructed using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING, https://string-db.org/),demonstrate the interaction relationships between DEGs. The whole network was further imported into Cytoscape software (https://cytoscape.org/), an open-source platform for visualizing complex networks and analyzed with its MCODE.
Immune cell composition
Cell-type enrichment scores were computed by running xCell on the full TPM gene expression matrix. In this study, we use the xCell to analyze the different immune cell subtypes between unstable plaque and stable plaque.
Results
Identification of differentially expressed mRNA in unstable carotid plaque (identification of DEGs)
Raw data of the high-throughput sequencing datasets were downloaded from GEO database and then further processed with Limmaffy in R package. As a result, compared with stable plaque, there have differentially expressed genes (DEGs), there are 1034 up-regulated and 592 down-regulated genes in unstable plaque of atherosclerosis (Supplementary Excel1).According to adjust p-value, TM4SF19, DCSTAMP, MDFIC, LIPA, SVEP1, CCDC88A, DBI, VNN1, MMP12, ATP6V0D2 were ranked among the top 10 up-regulated DEGs (Table1, Fig. 1A); and viceSGCA, FBLN5, SCHIP1, NTF3, HABP4, ERBB2, CSDC2, CLEC18A, MFGE8, ARHGEF25-1 had the most significant changes in down-regulated DEGs (Table1, Fig. 1B).

Identification of DEGs in controls and unstable carotid plaque. A Volcano plots showing the top 10 DEGs, the red dots represent up-regulated and the blue dots represent down-regulated mRNAs. B Heatmap illustrating the top differentially expressed genes (DEGs), red represents relatively high and blue represents relatively low
GO and KEGG analysis of DEGs
GO and KEGG pathway analyses were also performed on all the DEGs to fully explore the biological role of all the DEGs, which include all 1034 and 592 target genes. Among the up-regulated DEGs, the GO biological processes were involved in regulation of cell activation, immune effector process, leukocyte migration, regulation of cytokine production, myeloid cell activation involved in immune response. The KEGG pathway were involved the ‘lysosome’ pathway, chemokine signaling pathway, leukocyte trans-endothelial migration, Fc gamma R-mediated phagocytosis, platelet activation. Among the down-regulated genes, the GO biological processes were related to actin filament-based process, muscle structure development, muscle system process, cell-substrate adhesion; the KEGG pathway were related to vascular smooth muscle contraction and TGF-beta signaling pathway, PI3K–Akt signaling pathway, MAPK signaling pathway, adherens junction (Fig. 2A–D).

GO and KEGG pathway analysis of DEGs between carotid unstable plaque and stable plaque. a, c Show the major GO Biological process analysis of all up-regulated and down-regulated mRNAs. b, d The most KEGG signaling pathway enrichment of all up-regulated and down-regulated mRNAs
Prediction of upstream TFs and downstream TFs target genes of DEGs
I pulled 62 transcription factors from the Trust database. Indeed, 34 up-regulated and 28 down-regulated TFs were identified (Fig. 3A, B), and is presented as Heatmap (Fig. 3C). 13,532 TF–mRNA pairs were found in 34 up-regulated TFs, 8748 TF–mRNA pairs were found in 27 down-regulated TFs. Transcriptional regulatory networks indicated HMGA1 as the top-ranked regulator of up-regulated TFs, which promotes epithelial-to-mesenchymal transition in cancer and associated with PAH [10]; NFIX is the top-ranked regulator of down-regulated TFs(Fig. 3A). In order to display the regulatory network directly, we continued to construct the complex TFs–mRNA network by TFsNET, We show the top 50 TFs–mRNA pairs network (Fig. 3D, E).

Analysis upstream and downstream TFs of DEGs between unstable carotid plaque and stable plaque. A Transcription factors of all up-regulated mRNAs. B Transcription factors of the all down-regulated mRNAs. C The heatmap of upstream and downstream TFs. TFs regulates the network diagram of mRNA. The red diamond in the center represents the up-regulated TFs–mRNA, the surrounding red ellipses represent the up-regulated mRNA, and the green ellipses represent the down-regulated mRNA D; the green diamond in the center represents the down-regulated TFs–mRNA, the surrounding red ellipses represent the up-regulated mRNA, and the green ellipses represent the down-regulated E, which show top 50 TFs–mRNAs pairs
GO analysis of TFs
The up-regulated TFs were mainly enriched in positive regulation of myeloid leukocyte differentiation, and the down-regulated TFs were mainly enriched in connective tissue development (Fig. 4A, B). TF protein interaction network could also be obtained based on metascape database, in which macrophage differentiation and myeloid cell differentiation were identified, according to this, wo select 4 TFs(RB1, CEBPA, PPARG, BATF)for further investigated from all up-regulated TFs (Fig. 4C); TFs protein interaction network could also be obtained based on metascape database, in which cardiac vascular smooth muscle cell differentiation was identified, 4 down-regulated TFs (SRF, MYOCD, HEY2, GATA6) were selected from all TFs (Fig. 4D).

GO analysis of TFs. a The major GO enrichment of up-regulated TFs, b the major GO enrichment of down-regulated TFs. c Upregulated TFs of PPI network; d down-regulated TFs of PPI network.TF, transcription factor
Immune cell composition in the carotid unstable plaque and stable plaque
The result of immune cell composition of DEGs showed that the up-regulated DEGs were activated in the DC cell and macrophages, which enrichment score were significant differences in the unstable carotid plaque (p < 0.05), down-regulated DEGs were activated in the fibroblasts, myocytes, smooth muscle, which enrichment score were significant differences (p < 0.05) in the stable carotid plaque (Fig. 5).

Immune cell composition generated by xCell-inferred enrichment score of cell types across unstable plaque and stable plaque. *p < 0.05; **p < 0.01; ***p < 0.001 by Kruskal–Wallis test
Discussion
The role of inflammation in the formation, vulnerability and rupture of atherosclerotic plaque has received increasing attention. Macrophages are key instigators of atherosclerotic inflammation, but are also critical for the regression phase of the inflammatory process, and their contribution depends on their activation state. Rewiring the transcriptional state of macrophages is an attractive therapeutic strategy for CVD to prevent excessive inflammation [11]. Through bioinformatics analysis, we screened out hub genes that affect the stability of atherosclerotic plaques, as well as related biological processes and immune cells.
After reanalyzing mRNA high-throughput dataset of carotid plaque, compare to the stable plaque, we found that these upregulated mRNAs were enrichment in biological process including immune effector process and myeloid effector activation. This result is consistent with some studies, their result suggest that innate and adaptive immune dysregulation play an important role at the atherosclerotic site in patients with recent CV events [6, 12]. However, in a recent work of screening key TFs in regulating plaque stability with mRNA expression profiles, According to the up-regulated mRNA and down-regulated mRNA to predict upstream TFs and downstream TFs targets.
We select 4 up-regulated hub genes (RB1/CEBPA/PPARG/BATF) by gene expression signature in unstable carotid plaque. Among them, the retinoblastoma susceptibility gene (RB1) was the first tumor suppressor gene to be molecularly defined. Product RB1(pRB) is a chromatin-associated protein that limits the transcription of cell cycle genes, primarily via regulation of the E2F transcription factor [13]. It is involved in the development of many tumor diseases, for example, human glioblastoma [14], Human Colon Cancer [15], small cell lung cancer [16], and so on. However, it is not clear in the mechanism of cardiovascular disease. In 2019, Prestel, M et al. founding that allele-specific transcriptional regulation of HDAC9 via E2F3 and RB1 as a major mechanism mediating vascular risk at rs2107595 [17]; but Yu Cao et al. another findings reveals that endothelial RB positively impacts arterial function by supporting vasoprotective endothelial DHFR/NO pathway activity, leading to reduced abdominal aortic aneurysm (AAA) development [18], their results are inconsistent. In the results of our analysis, RB1 expression was increased in unstable plaques, Therefore, further animal experiments are needed to confirm this.
CEBPA as a myeloid transcription factor, its mutations play an important role in the pathogenesis of hematological tumors [19, 20]. But it is less well studied in cardiovascular disease and in promoting atherosclerosis. A research showed that C/EBP transcription factors mediated epicardial activation during heart development and injury, disruption of C/EBP signaling in the adult epicardium reduced injury-induced neutrophil infiltration and improved cardiac function [21]. However, no studies have shown the mechanism of its role in the progression of atherosclerotic plaque. Our analysis of the results showed that he was up-regulated in unstable plaques, At the same time, a single-cell sequencing database analysis showed that (https://panglaodb.se/index.html) CEBPA was highly expressed in macrophages(Additional file 1: figure S1). Therefore, we infer that it plays an important role in the development of atherosclerosis.
Another transcription factor, The peroxisome proliferator-activated receptor gamma gene (PPARG), encodes a member of the peroxisome-activated receptor subfamily of nuclear receptors. PPARG-gamma has been implicated in the pathology of numerous diseases including obesity, diabetes, atherosclerosis and cancer [22], and also plays a key role in subcellular metabolism of arterial wall macrophage foam cells [23]. Genetic variation in PPARG has been associated with metabolic and cardiovascular end points. Khalid Z Al-Shali et al. results suggested that PPARG c.1431 T allele had greater total carotid plaque volume (TPV) than others, with no between-genotype difference in IMT. The findings show an association between PPARG genotypes and carotid arterial phenotypes [24]. Our bioinformatics analysis suggest that it was up-regulated in unstable plaques, Therefore, the mechanism of its effect on vascular plaque stability needs to be explored in further experiments.
Transcription factor BATF as a key transcription factor, regulation of Treg differentiation in non-lymphoid cells [25], which most are used to regulate T cells in viral and bacterial infections and in tumor immunity disease [26, 27], very little research has been done on cardiovascular disease. One studies have shown that after in vitro and in vivo injection of endotoxin and LPS, six candidate genes (BATF, BID, C3aR1, IL1RN, SEC61B and SLC43A3) were identified to be associated with inflammation and atherosclerosis [28]. Another study demonstrate that the BATF and BATF3 are critical regulators of T effector functions, thus making them attractive targets for therapeutic interventions in heart allograft [29]. Our xCell results showed that the number of macrophages in unstable plates increased significantly, indicating that the number and function of macrophages play a very important role in the progression of plaques, above studies indicate that BATF is mainly involved in the progression of cardiovascular disease as a regulatory factor of T cells, a single-cell sequencing database analysis showed that (https://panglaodb.se/index.html) BATF was also highly expressed in macrophages, so, it may be involved in regulating plaque instability or rupture.
SRF, MYCOD, HEY2, GATA6 are as transcription factors, our results showed that they were down-regulated in unstable plaques, whereas they were up-regulated in stable plaques. Its xCell results turned out to be fibroblasts and muscle cells (p < 0.001). These transcription factors are involved in the molecular regulation of heart development and disease. For example, SRF、MYOCD are related to the development of cardiac vascular smooth muscle and molecular regulation of diseases [30,31,32], Mutations in HEY2 cause Brugada syndrome, a rare heart disease [33, 34]. Fibroblast GATA6 promotes myocardial adaptation to pressure overload by enhancing cardiac angiogenesis [35].
TFs protein interaction network could also be obtained based on metascape database, in which macrophage differentiation and myeloid cell differentiation were identified. This is consistent with immune cell composition result. Single-cell sequencing of some atherosclerotic plaques found that T cells and macrophages dominate the atherosclerotic plaque immune landscape [6].
Conclusion
In conclusion, findings in the current study demonstrated that the vulnerability or unstable atherosclerotic plaques might be the result of imbalance between macrophages and fibrosis. Crosstalk between macrophages and SMCs may be involved in this process. Specifically, up-regulated RB1, CEBPA, PPARG, BATF, and down-regulated SRF, MYCOD, HEY2, GATA6 performed important promotional effects for the rupture of atherosclerotic plaques. These findings provide a theoretical basis for the future study of specific mechanisms of atherosclerotic rupture.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
References
Kobiyama K, Ley K. Atherosclerosis. Circ Res. 2018;123(10):1118–20.
Jiang P, Chen Z, Hippe D, Watase H, Sun B, Lin R, et al. Association between carotid bifurcation geometry and atherosclerotic plaque vulnerability: a Chinese atherosclerosis risk evaluation study. Arterioscler Thromb Vasc Biol. 2020;40(5):1383–91.
Bentzon J, Otsuka F, Virmani R, Falk E. Mechanisms of plaque formation and rupture. Circ Res. 2014;114(12):1852–66.
Gonçalves I, den Ruijter H, Nahrendorf M, Pasterkamp G. Detecting the vulnerable plaque in patients. J Intern Med. 2015;278(5):520–30.
Gisterå A, Hansson GK. The immunology of atherosclerosis. Nat Rev Nephrol. 2017;13(6):368–80.
Fernandez D, Rahman A, Fernandez N, Chudnovskiy A, Amir E, Amadori L, et al. Single-cell immune landscape of human atherosclerotic plaques. Nat Med. 2019;25(10):1576–88.
Lambert S, Jolma A, Campitelli L, Das P, Yin Y, Albu M, et al. The human transcription factors. Cell. 2018;172(4):650–65.
Edsfeldt A, Swart M, Singh P, Dib L, Sun J, Cole J, et al. Interferon regulatory factor-5-dependent CD11c+ macrophages contribute to the formation of rupture-prone atherosclerotic plaques. Eur Heart J. 2022;43(19):1864.
Oppi S, Nusser-Stein S, Blyszczuk P, Wang X, Jomard A, Marzolla V, et al. Macrophage NCOR1 protects from atherosclerosis by repressing a pro-atherogenic PPARγ signature. Eur Heart J. 2020;41(9):995–1005.
Hopper R, Moonen J, Diebold I, Cao A, Rhodes C, Tojais N, et al. In pulmonary arterial hypertension, reduced bmpr2 promotes endothelial-to-mesenchymal transition via HMGA1 and Its target slug. Circulation. 2016;133(18):1783–94.
Kuznetsova T, Prange K, Glass C, de Winther M. Transcriptional and epigenetic regulation of macrophages in atherosclerosis. Nat Rev Cardiol. 2020;17(4):216–28.
Depuydt MAC, Prange KHM, Slenders L, Örd T, Elbersen D, Boltjes A, et al. Microanatomy of the human atherosclerotic plaque by single-cell transcriptomics. Circ Res. 2020;127(11):1437–55.
Dyson N. RB1: a prototype tumor suppressor and an enigma. Genes Dev. 2016;30(13):1492–502.
Wang L, Karpova A, Gritsenko M, Kyle J, Cao S, Li Y, et al. Proteogenomic and metabolomic characterization of human glioblastoma. Cancer Cell. 2021;39(4):509-28.e20.
Vasaikar S, Huang C, Wang X, Petyuk V, Savage S, Wen B, et al. Proteogenomic analysis of human colon cancer reveals new therapeutic opportunities. Cell. 2019;177(4):1035-49.e19.
George J, Lim J, Jang S, Cun Y, Ozretić L, Kong G, et al. Comprehensive genomic profiles of small cell lung cancer. Nature. 2015;524(7563):47–53.
Prestel M, Prell-Schicker C, Webb T, Malik R, Lindner B, Ziesch N, et al. HDAC9The atherosclerosis risk variant rs2107595 mediates allele-specific transcriptional regulation of via E2F3 and Rb1. Stroke. 2019;50(10):2651–60.
Cao Y, Xiong D, Kong R, Dai G, Zhong M, Li L, et al. Endothelial retinoblastoma protein reduces abdominal aortic aneurysm development via promoting DHFR/NO pathway-mediated vasoprotection. Mol Cell Biochem. 2019;460:29–36.
Taube F, Georgi J, Kramer M, Stasik S, Middeke J, Röllig C, et al. CEBPA mutations in 4708 patients with acute myeloid leukemia: differential impact of bZIP and TAD mutations on outcome. Blood. 2022;139(1):87–103.
Brown A, Hahn C, Scott H. Secondary leukemia in patients with germline transcription factor mutations (RUNX1, GATA2, CEBPA). Blood. 2020;136(1):24–35.
Huang G, Thatcher J, McAnally J, Kong Y, Qi X, Tan W, et al. C/EBP transcription factors mediate epicardial activation during heart development and injury. Science. 2012;338(6114):1599–603.
Mustafa H, Albkrye A, AbdAlla B, Khair M, Abdelwahid N, Elnasri H. Computational determination of human PPARG gene: SNPs and prediction of their effect on protein functions of diabetic patients. Clin Transl Med. 2020;9(1):7.
Li A, Brown K, Silvestre M, Willson T, Palinski W, Glass C. Peroxisome proliferator-activated receptor gamma ligands inhibit development of atherosclerosis in LDL receptor-deficient mice. J Clin Investig. 2000;106(4):523–31.
Al-Shali K, House A, Hanley A, Khan H, Harris S, Zinman B, et al. Genetic variation in PPARG encoding peroxisome proliferator-activated receptor gamma associated with carotid atherosclerosis. Stroke. 2004;35(9):2036–40.
Delacher M, Imbusch C, Hotz-Wagenblatt A, Mallm J, Bauer K, Simon M, et al. Precursors for nonlymphoid-tissue Treg cells reside in secondary lymphoid organs and are programmed by the transcription factor BATF. Immunity. 2020;52(2):295-312.e11.
Wu X, Kasmani M, Zheng S, Khatun A, Chen Y, Winkler W, et al. BATF promotes group 2 innate lymphoid cell-mediated lung tissue protection during acute respiratory virus infection. Sci immunol. 2022;7(67):eabc9934.
Wei J, Long L, Zheng W, Dhungana Y, Lim S, Guy C, et al. Targeting regnase-1 programs long-lived effector T cells for cancer therapy. Nature. 2019;576(7787):471–6.
Sivapalaratnam S, Farrugia R, Nieuwdorp M, Langford C, van Beem R, Maiwald S, et al. Identification of candidate genes linking systemic inflammation to atherosclerosis; results of a human in vivo LPS infusion study. BMC Med Genom. 2011;4:64.
Wang Y, Xiao X, Kong G, Wen M, Wang G, Ghobrial R, et al. Genetically targeting the BATF family transcription factors BATF and BATF3 in the mouse abrogates effector T cell activities and enables long-term heart allograft survival. Am J Transpl Off J Am Soc Transpl Am Soc Transpl Surg. 2021;22(2):414–26.
Owens G, Kumar M, Wamhoff B. Molecular regulation of vascular smooth muscle cell differentiation in development and disease. Physiol Rev. 2004;84(3):767–801.
Huang J, Elicker J, Bowens N, Liu X, Cheng L, Cappola T, et al. Myocardin regulates BMP10 expression and is required for heart development. J Clin Investig. 2012;122(10):3678–91.
Huang J, Wang T, Wright A, Yang J, Zhou S, Li L, et al. Myocardin is required for maintenance of vascular and visceral smooth muscle homeostasis during postnatal development. Proc Natl Acad Sci USA. 2015;112(14):4447–52.
Bezzina C, Barc J, Mizusawa Y, Remme C, Gourraud J, Simonet F, et al. Common variants at SCN5A-SCN10A and HEY2 are associated with Brugada syndrome, a rare disease with high risk of sudden cardiac death. Nat Genet. 2013;45(9):1044–9.
Veerman C, Podliesna S, Tadros R, Lodder E, Mengarelli I, de Jonge B, et al. HEY2The brugada syndrome susceptibility gene modulates cardiac transmural ion channel patterning and electrical heterogeneity. Circ Res. 2017;121(5):537–48.
Dittrich G, Froese N, Wang X, Kroeger H, Wang H, Szaroszyk M, et al. Fibroblast GATA-4 and GATA-6 promote myocardial adaptation to pressure overload by enhancing cardiac angiogenesis. Basic Res Cardiol. 2021;116(1):26.
Acknowledgements
My deepest gratitude goes first and foremost to professor xiuyun Ren, my supervisor, for her constant encouragement and guidance. She has walked me through all the stages of the writing of this thesis. Without her consistent and illuminating instruction, this thesis could not have reached its present form.
Funding
The study was supported by grants from the National Natural Science Foundations of China (No. 81271144), Excellent Achievements (Science and Technology) Cultivation Project of Shanxi Colleges and Universities (2020KJ014); Local Science and Technology Development Fund Project guided by the Central Government of Shanxi Provincial Department of Science and Technology (YDZX20201400001131).
Author information
Authors and Affiliations
Contributions
Study conception or design: XYR, FYY, JHG; data analyzing and draft manuscript preparation: RC, XJX, critical revision of the paper: SRY, ZQM, supervision of the research: YZ, final approval of the version to be published (the names of all authors must be listed): RC, XJX, SRY, ZQM, YZ, JHG, FYY, XYR. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Competing interests
The authors declare no competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1. CEBPA
was highly expressed in macrophages. This analysis comes from https://panglaodb.se/index.html.
Additional file 2: Excel S1.
All the differentially expressed genes between unstable and stable plaque of atherosclerosis.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Cheng, R., Xu, X., Yang, S. et al. The underlying molecular mechanisms and biomarkers of plaque vulnerability based on bioinformatics analysis. Eur J Med Res 27, 212 (2022). https://doi.org/10.1186/s40001-022-00840-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40001-022-00840-7