
Abstract
Background
Soil respiration (SR) is a critical process for understanding the impact of climatic conditions and land degradation on the carbon cycle in terrestrial ecosystems. We measured the SR and soil environmental factors over 1 year in four land uses with varying levels of disturbance and different vegetation types viz., mixed forest cover (MFC), Prosopis juliflora (Sw.) forest cover (PFC), agricultural field (AF), and vegetable field (VF), in a semi-arid area of Delhi, India. Our primary aim was to assess the effects of soil moisture (SM), soil temperature (ST), and soil microbial activity (SMA) on the SR.
Methods
The SR was measured monthly using an LI-6400 with an infrared gas analyser and a soil chamber. The SM was measured using the gravimetric method. The ST (10 cm) was measured with a probe attached to the LI-6400. The SMA was determined by fluorescein diacetate hydrolysis.
Results
The SR showed seasonal variations, with the mean annual SR ranging from 3.22 to 5.78 μmol m−2 s−1 and higher SR rates of ~ 15–55% in the cultivated fields (AF, VF) than in the forest sites (MFC, PFC). The VF had significantly higher SR (P < 0.05) than the other land uses (AF, PFC, MFC), which did not vary significantly from one another in SR (P < 0.05). The repeated measures ANOVA evaluated the significant differences (P < 0.05) in the SR for high precipitation months (July, August, September, February). The SM as a single factor showed a strong significant relationship in all the land uses (R2 = 0.67–0.91, P < 0.001). The effect of the ST on the SR was found to be weak and non-significant in the PFC, MFC, and AF (R2 = 0.14–0.31; P > 0.05). Contrasting results were observed in the VF, which showed high SR during summer (May; 11.21 μmol m−2 s−1) and a significant exponential relationship with the ST (R2 = 0.52; P < 0.05). The SR was positively related to the SMA (R2 = 0.44–0.5; P < 0.001). The interactive equations based on the independent variables SM, ST, and SMA explained 91–95% of the seasonal variation in SR with better model performance in the cultivated land use sites (AF, VF).
Conclusion
SM was the key determining factor of the SR in semi-arid ecosystems and explained ~ 90% of the variation. Precipitation increased SR by optimizing the SM and microbial activity. The SMA, along with the other soil factors SM and ST, improved the correlation with SR. Furthermore, the degraded land uses will be more susceptible to temporal variations in SR under changing climatic scenarios, which may influence the carbon balance of these ecosystems.
Similar content being viewed by others

Background
Soil respiration (SR) is the second largest flux of carbon (C) between terrestrial ecosystems and the atmosphere (Hanson et al. 2000). SR is considered a key process in the terrestrial C cycle, and it releases 98 Pg C per year into the atmosphere (Bilandžija et al. 2016; Zhao et al. 2017). Any small variation in SR has a significant impact on the carbon dioxide (CO2) concentration in the atmosphere which in turn affects the global C cycle (Black et al. 2017). Therefore, understanding the dynamics of the SR in any ecosystem is critical for combatting climate change. SR has two components: auto- (roots and rhizosphere) and heterotrophic respiration (soil microbes and soil fauna) (Chen et al. 2017). Several abiotic and biotic factors, including soil temperature (ST), soil moisture (SM) (Bao et al. 2016), availability of C substrates for microorganisms, soil microbial activity (SMA) (Tang et al. 2018), soil fertility (Butnor et al. 2003), plant photosynthetic activity (Zhang et al. 2013a, 2013b), and soil organisms (Rai and Srivastava 1981), influence the rate of soil CO2 efflux. SR is also affected by various management activities, including land use change, which contributes 12.5% of the global CO2 emissions to the atmosphere, mainly as a result of deforestation (IPCC 2013). Several studies have reported the potential impacts of cultivation and deforestation activities on the soil C storage and efflux of CO2 (Lou et al. 2004; Rey et al. 2011; Peri et al. 2015).
Climate change has a strong impact on precipitation patterns across the globe (Arredondo et al. 2018; Darrouzet-Nardi et al. 2018). Changes in precipitation patterns in any terrestrial ecosystem will ultimately affect the SM, which in turn influence SMA, soil organic matter (SOM) decomposition pattern, and SR (Bao et al. 2019). Arid and semi-arid ecosystems cover approximately 41% of the terrestrial land surface of the Earth (Wang et al. 2014). The unpredictable and random precipitation events in semi-arid ecosystems interact with the seasons and the functioning of auto- and heterotrophic ecosystem processes (Miao et al. 2017). Such events are vulnerable to climate change and can have crucial impacts on SR, often causing a pulse of CO2 to be emitted into the atmosphere (Shen et al. 2016; Gu et al. 2018). There have been studies on the SR in arid and semi-arid ecosystems with respect to annual flux measurements (Subke et al. 2006; Sawada et al., 2016b). The SM is considered an important environmental determinant controlling the rate of SR in these ecosystems (Rey et al. 2002; Conant et al. 2004; Jarvis et al. 2007; Miao et al. 2017). The SM can limit the widely accepted positive linear and exponential relationship between the SR and ST by limiting the soil microbial activity (SMA) under low moisture conditions (Wang et al. 2014). Therefore, in such an ecosystem, both the ST and SM strongly influence respiration rates, and their relative importances can vary seasonally and spatially (Reichstein et al. 2002; Tang and Baldocchi 2005; Sun et al. 2018). Hence, we assume that the SR in semi-arid ecosystems is limited by the SM and SMA in the different land use/land cover systems.
Delhi has a unique forest ecosystem located on ridges that are extensions of the Aravalli hills; these ridges are 32 km long and serve various ecological, environmental, and social functions. The Delhi ridge has been designated as a reserved forest and is managed mainly with the objectives of increasing forest cover, biodiversity, and conservation through public participation and reduction in monoculture plantations and encroachments (Sinha 2014). Delhi is also considered one of the most polluted cities in the world. The land use pattern of Delhi showed that 15.00% (221.4 km2) of the total geographical area comprised the net sown area, 8.07% (119 km2) was the current fallow area, 6.71% (98.9 km2) was culturable wasteland, and 1.00% (14.8 km2) was forest cover (FSI 2017). Previous studies on SR have been conducted in temperate and riparian subtropical regions of India (Jha and Mohapatra 2011), and the data from the semi-arid ecosystems of India are very limited. Our study will provide relevant data on SR for future research in the semi-arid ecosystems of India. The objectives of this study were (1) to measure and compare the spatio-temporal variations in SR in the different types of land use systems in a semi-arid area of Delhi, India, and (2) to understand the impacts of SM, ST, and SMA and/or the interactive effect of these factors on SR.
Methods
Study area
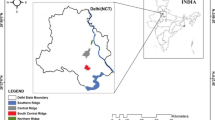
The study was carried out in a semi-arid area of Delhi, which is part of the National Capital Territory (NCT) of India (28.40° N to 28.41° N, 76.84° E to 77.40° E) and covering an area of 1483 km2 (Fig. 1). The area is bounded by the Indo-Gangetic alluvial plains in the north and east, the Thar Desert in the west, and the Aravalli Range and the hill ranges in the south. Delhi lies within the interior of the northern plains of the Indian subcontinent. The climate of Delhi is greatly influenced by the Himalayas and the Thar Desert due to its proximity. The climate of the area is semi-arid and dry except during the monsoon season and is characterized by hot summers (April–June), monsoons (July–September), cool and dry winters (November–December), and two periods of pleasant transitional weather, i.e., autumn (October) and spring (February to March). The climate is influenced by two weather events, i.e., the western disturbances and south-westerly winds. The study area receives most of the annual rainfall during the monsoon season. The vegetation of the study area is ravine thorn forest, which belongs to the ecosystem type of tropical thorn forest (6B/C) (Champion and Seth 1968) and covers 33% of the total forest area and 67% of plantation and tree outside forest (TOF) areas. The vegetation is mainly dominated by middle-story thorny trees, which are interspersed with open patches due to their scattered distribution (Sinha 2014). The soil type on the ridge has been reported as sandy loam to loam (Chibbar 1985). Prosopis juliflora (Sw.) DC, which is an exotic species, is the dominant tree in the forests. Acacia nilotica (L.) Delile, Acacia leucophloea (Roxb.) Willd., Salvadora oleoides Decne, and Cassia fistula L. are among the commonly found native trees (Sinha 2014; Meena et al. 2016). The naturally growing shrubs in the forests are Justicia adhatoda L., Capparis sepiaria L., Carissa spinarum L., Jatropha gossypifolia L., and Opuntia dillenii L.

Map of the study area (Delhi) showing the land use sites. PFC Prosopis juliflora forest cover, MFC mixed forest cover, AF agriculture field and VF vegetable field
Land use site description
To study the spatio-temporal variation in SR, we chose four different land uses based on the levels of human disturbance (mainly cultivation) and plant species cover (Fig. 1). The land uses were (1) mixed forest cover (MFC) as the native vegetation cover (28.61° N; 77.17° E); (2) P. juliflora dominated forest cover (PFC) as the exotic tree cover (28.69° N; 77.22° E); (3) an agricultural field (AF), which was located near the Nazafgarh drain (28.54° N; 76.87° E); and (4) a vegetable field (VF) located along the Yamuna flood plains (28.52° N; 77.34° E). The vegetation of the PFC stand was characterized by a total tree density (TD) of 350 individuals ha−1 and a mean basal area (MBA) of 25.05 m2 ha−1. Under the MFC, TD and MBA were higher than under the PFC, at 400 individuals ha−1 and 117.26 m2 ha−1, respectively. However, under the MFC, P. juliflora was found to be the most dominant tree species with the highest TD (200 individuals ha−1), but other associated tree species viz., Pongamia pinnata L. (50 individuals ha−1), Azadirachta indica Juss. (50 individuals ha−1), A. nilotica (75 individuals ha−1), and C. fistula (25 individuals ha−1) were also observed (Meena et al. 2019). The maximum basal area (BA) values were estimated for A. nilotica (160.52 m2 ha−1) and C. fistula (147.37 m2 ha−1), whereas the BA was comparatively low for P. juliflora (95.74 m2 ha−1). The AF was mainly cropped with Triticum aestivum L. during winter (October–May) and Phaseolus vulgaris L. (September–October). The field was irrigated by a tube well during the growing season. The VF field was mainly cultivated with Capsicum annum L. throughout the year except between September and November, during which Brassica oleraceae L. was grown. The VF was regularly irrigated by water pumped from the Yamuna River. The soil type in PFC, MFC, and VF was sandy loam, whereas in AF, it was loamy sand.
Soil respiration measurements
The SR was measured in the selected land use systems from April 2012 to March 2013 with a LI-6400 (LI-COR Inc., Lincoln, NE, USA), which consisted of an infrared gas analyser (IRGA) and a soil chamber (LI-6400-09) of 962 cm3 in volume and 72 cm2 area. The SR was measured with a stratified random sampling design in each land use type to account for the spatial variability in soil properties and vegetation cover. Each measurement was the mean value of three observations at each sampling site. Collars made from polyvinyl chloride (PVC) pipes (10 cm diameter and 6 cm height) were gently inserted 2 cm into the soil, leaving approximately 4 cm of the collar above the soil surface at each point for 24 h prior to the SR measurement to minimize any disturbance during the measurement. Before taking the measurement, the soil surface within the collar was kept free of any live vegetation and residues by removing the seedlings and their roots to avoid autotrophic respiration. All measurements were conducted in the morning between 09:00 and 11:00 a.m. to avoid the high midday temperatures in the study region (Lou et al. 2004). The soil chamber was placed on the PVC collar and fixed to the ring to record the SR inside the collar. Before the SR measurement, the concentrations of CO2 within the soil chamber were lowered to below the ambient CO2 concentration, and then the increase in CO2 was logged until it stabilized.
Soil sampling and analysis
Soil samples were collected from five different points at 0–10 cm depth and pooled together to obtain a composite sample for each land use type. The visible root mass was removed from the soil samples by hand. The SM content was measured with the gravimetric method by oven drying approximately 50 g of fresh soil at 105 °C until it reached a constant weight and then weighing it to note the dry weight. The ST, up to 10 cm, was measured with a probe attached to the LI-6400. The ST readings were recorded at the same time as the SR readings.
The soil samples were passed through a 2-mm sieve, ground in a mortar with a pestle, and stored at room temperature for further analysis. The soil carbon (SC) and soil nitrogen (SN) concentrations were measured with an Elementar CHNS analyser. The SMA was determined by fluorescein diacetate (FDA) hydrolysis according to the method of Adam and Duncan (2001). A 2-g moist and sieved soil sample was taken in a conical flask and mixed with 15 ml potassium phosphate buffer (60 mM) with a pH of 7.6. To the soil, 0.2 ml of FDA stock solution (1000 μg FDA ml−1) was added to start the reaction. The blanks were prepared without the addition of FDA. The samples were shaken at 100 rev min−1 in an orbital incubator shaker at 30 °C for 20 min. After incubation, a 15 ml chloroform:methanol (2:1) solution was added to the soil samples to terminate the reaction. The contents were then centrifuged at 2000 rev min−1 for 3 min. Supernatants from each sample were filtered through Whatman filter paper No. 2. Standards were made by using a fluorescent stock solution (2000 μg ml−1). The absorbance of the standards and samples was measured at 490 nm using a spectrophotometer (RIGOL, USA).
Data analysis
One-way analysis of variance (ANOVA) was used to evaluate the variations in the SR, ST, SM, SMA, SC, and SN among the different land use types using Tukey’s test at P < 0.05. A repeated measures ANOVA was performed on the SR data using the measurement months and land uses (MFC, PFC, AF, and VF) as factors. Pearson analysis was performed to investigate the correlation of the environmental factors with SR in all land uses. The linear regression was used to study the relationship of the SR with the SM and SMA. For the relationship of the SR with the ST, exponential and nonlinear regression analysis was done.
The interactive effect of the ST and SM on the SR was determined by using two independent variable regression equations as described by Li et al. (2018) as follows:
The interactive effect of the ST, SM and SMA on the SR was evaluated as follows:
where a, b, c, and d are coefficients.
The criteria used for model selection were Akaike’s information criteria (AIC) and the coefficient of determination (R2). The AIC was calculated as follows:
where RSS is the residual sum of squares, N is the sample size, and p is the number of independent variables. The model with the lowest AIC and the highest R2 value was selected as the best-fitting model.
All statistical analyses were performed using SPSS version 16.0.
Results
The monthly climatic variables of the study area during the SR measurement period are shown in Fig. 2. The air temperature during winter reached a minimum of 6 °C (January) and began rising in March, peaked in summer (May and June) to a maximum of 41 °C, and then declined in the monsoon season. The total precipitation received during the study period was 719.98 mm, of which 73% was received during the monsoon season (July–September). High precipitation was also recorded during spring (February), which contributed 15% of the total rainfall.

Monthly average maximum and minimum air temperature (°C) and total precipitation (mm) for the period 2012–2013 in study area Delhi, India
Seasonal variation in the soil environmental factors and SR
The mean monthly ST (°C) was recorded to be high in summer (May: 37.69 ± 1.94, 36.04 ± 0.99, 46.63 ± 1.8, and 34.5 ± 0.09 in MFC, PFC, AF, and VF, respectively), to gradually decline in the monsoon season (July–September), and to be low in winter (November: 19.65 ± 0.11, 16.87 ± 0.11, and 18 ± 0.07 in MFC, PFC, and AF, respectively; December: 19.5 ± 0.18 in AF). The mean annual ST (°C) values were 26.33 ± 2.98, 25.42 ± 2.99, 29.36 ± 5.19, and 26.36 ± 3.06 for MFC, PFC, AF, and VF, respectively. There were no significant differences in the ST values between any land use sites (P > 0.05) (Fig. 3b). The ST was significantly correlated with the SR only in the VF (R = 0.7; Table 1), whereas in the other land uses, the relationship was found to be non-significant.

Annual spatial variations of soil respiration (SR) and environmental factors among land uses. Mean annual SR soil respiration (a), ST soil temperature (at 10 cm depth) (b), SM soil moisture (c), SMA soil microbial activity (d), SC soil carbon (e), and SN soil nitrogen (f) of PFC Prosopis juliflora forest cover, MFC mixed forest cover, AF agriculture field and VF vegetable field in a semi-arid area. Standard error of each mean is represented over each bar. Different letters denote significant (P < 0.05) differences among the four land uses sites (Tukey test after one-way ANOVA)
For the SM content, no significant difference was found among the PFC, MFC, and AF (P > 0.05), but a significant different was found for the VF (P < 0.05). The mean annual SM (%) values were 1.99 ± 0.92, 1.49 ± 0.94, 2.08 ± 0.9, and 3.97 ± 0.43 for MFC, PFC, AF, and VF, respectively (Fig. 3c). The seasonal SM pattern was influenced by the monthly rainfall (mm), which had high values (%) in the monsoonal month of September (3.96 ± 0.02 and 5.23 ± 0.02 in MFC and PFC, respectively) and in February (5.9 ± 0.14 in AF). In the VF, a consistently high SM was recorded throughout the year, with maximum values in summer (May: 5.59 ± 0.14) (Fig. 4b).

Seasonal variation of soil respiration (SR) and environmental factors for the four land uses. Mean monthly ST soil temperature (at 10 cm depth) (a), SM soil moisture (b), SMA soil microbial activity (c), and SR soil respiration (d) of PFC Prosopis juliflora forest cover, MFC mixed forest cover, AF agriculture field, and VF vegetable field in a semi-arid area of Delhi. Standard error of each mean is represented over each line
The mean monthly SMA was significantly higher in the forests (PFC and MFC) than in the cultivated sites (AF and VF) (P < 0.05). The annual mean SMA (μg g−1 min−1) was 7.95 ± 0.42, 7.55 ± 0.53, 2.98 ± 0.3, and 3.08 ± 0.33 in the MFC, PFC, AF, and VF, respectively (Fig. 3d). Similarly, the mean annual SC and SN (g kg−1) were also significantly higher in the forests, i.e., 38.95 ± 0.21 and 3.85 ± 0.25 in the MFC, respectively, and 29.31 ± 2.07 and 3.13 ± 0.25 in the PFC, respectively, compared to those in the arable land uses, i.e., 6.88 ± 0.32 and 0.75 ± 0.04 in the AF, respectively, and 14.03 ± 0.43 and 1.31 ± 0.08 in the VF, respectively (Fig. 3e, f). However, no significant correlation of the SC or SN with the SMA was found at any of the sites (Table 1). Similar to the SM pattern, the SMA was also high in the monsoon season (July: 9.76 ± 0.24 in the MFC; August: 4.02 ± 0.02 in the AF) and in February (9.01 ± 0.01 and 5.34 ± 0.42 in the PFC and VF, respectively) (Fig. 4c). Furthermore, the positive correlation of the SMA with the SM with a significant correlation (R = 0.62, 0.64 in MFC and VF, respectively) suggests that the SM influences the seasonal SMA along with SC and SN (Table 1).
SR showed a seasonal pattern with a peak in the monsoon season and a sharp decline in summer for the PFC, MFC, and AF (Fig. 4d). The SR (μmol m−2 s−1) was the lowest in summer (May), at 0.97 ± 0.27, 0.65 ± 0.14, and 0.53 ± 0.1 in the MFC, PFC, and AF, respectively, and the highest in the monsoon season (September: 8.12 ± 0.31 and 7.9 ± 0.39 in the MFC and PFC, respectively) and in February (9.42 ± 0.09 in AF). In contrast, in the VF, the SR was high in summer (May; 11.21 ± 0.08) and low in winter (November: 2.33±0.02). The SR in the VF was significantly different from those of the other land uses (P < 0.05). The mean annual SR was 3.22 ± 1.24, 2.57 ± 1.28, 3.75 ± 1.47 and 5.78 ± 1.39 μmol m−2 s−1 in the MFC, PFC, AF, and VF, respectively (Fig. 3a). The repeated measures ANOVA of the monthly SR evaluated significant differences for the monsoonal months (July, August, September) and February from rest of the year (P < 0.05) and a significant interaction between the monthly SR and the land use sites (F = 219.14, P < 0.05, df = 6.38). A strong and significant correlation between the SR and the SM (R = 0.82–0.95, P = 0.01) at all land use sites suggested that the SM was an important controlling factor of the SR. Furthermore, a significant correlation of the SR with the SMA (R = 0.67–0.71, P = 0.05) in all land uses except in the PFC (Table 1) further supported the influence of microbial activity on the SR.
Soil environmental factors controlling SR
The linear regression function effectively represented the influence of the SM on the SR and showed a strong significant positive interaction (R2 = 0.67–0.91, P < 0.001) in all land uses (Fig. 5b). SM as a single factor explained 67–92% of the total variation in SR. However, the exponential and nonlinear functions, considering the ST alone, determined only 12–50% of the changes in SR. The effect of the ST was found to be significantly positive only in the VF (R2 = 0.5, P < 0.05) and non-significant at other sites (P > 0.05) (Fig. 5a). The effect of the SMA alone was significant in the cultivated land uses (R2 = 0.5 and 0.67 in the AF and VF, respectively, P < 0.05) and explained 15–67% of the total variation in SR (Fig. 5c).

Relationship between SR soil respiration and environmental factors among four land uses. ST Soil temperature (at 10 cm depth) (a), SM soil moisture (b), and SMA soil microbial activity (c) (showing the best fit) of PFC Prosopis juliflora forest cover, MFC mixed forest cover, AF agriculture field, and VF vegetable field
The model that used the interactive effects (equations 1 and 2) and considered the ST and SM as independent variables showed an improved relationship in the VF only (R2 = 0.87; Table 2). However, the model with SMA along with SM and ST (equations 3, 4, 5) improved the model parameters with comparatively low AIC values and higher R2 values of 0.84, 0.95, 0.85, and 0.91 in the MFC, PFC, AF and VF, respectively. However, a better fit was obtained in the cultivated (AF, VF) sites than in the forest land use (MFC, PFC) sites (Table 2). This suggests that the SR was controlled by the interaction of the ST, SM, and SMA rather than by one factor.
Discussion
The main aim of our study was to understand the soil factors (SM, ST, and SMA) that influence the rate of SR in the different land use systems of the semi-arid area of Delhi by using various regression equation models. The obtained results clearly demonstrated that the SM alone (90%) controlled the SR rates of the studied region. In addition, the interactive models that consisted of the SM and other factors, such as the ST and SMA, effectively explained the variations in SR. Compared to the reported values for other semi-arid ecosystems, the annual mean SR rates of the studied region (2.55–5.78 μmol m−2 s−1) were higher than those of steppe ecosystems of Spain (0.72–1.24 μmol m−2 s−1, Rey et al. 2011) and North China (1.37–1.91 μmol m−2 s−1, Zeng et al. 2018) and were comparable with those of the Loess Plateau in China (2.03 to 3.23 μmol m−2 s−1, Shi et al. 2014).
Seasonal dynamics of SR
The variations in the rainfall pattern, intensity, and frequency have significant impacts on the SM, causing variation in SOM decomposition, SC mineralization, microbial activity, plant growth and species composition, above- and belowground biomass production, and plant phenological traits (Bao et al. 2019; Zhang et al. 2019). We observed a strong seasonal variation in the SR across all land uses, with higher SR rates during the rainy season, i.e., the hot-humid climate (July–September) when SM was not limiting (Fig. 4d). This suggests that the SM and ST would increase the production of aboveground biomass as a result of the high availability of resources for photosynthesis and the activity of the microorganisms in the monsoon season compared to those in other seasons (Zhou et al. 2014; Zhang et al. 2019). These results are in accordance with previous studies showing seasonal SR in riparian and subtropical semi-arid regions of India, where the highest CO2 effluxes were recorded in monsoons (Jha and Mohapatra 2011; Arora and Chaudhry, 2017). However, in dry seasons, soil water stress conditions could have reduced microbial activity, thereby decreasing SR (Li et al. 2018). This could also explain the contrasting rise of SR in VF in summers compared to other land use sites, as here, the SM content is consistent because of regular irrigation due to its proximity to the Yamuna River (Fig. 4b, d).
In this study, high rainfall increased the SM content and the SR during the monsoon season by ~ 80–120% in forest land uses compared to ~ 17–46% at the cultivated sites. Furthermore, the effect of the sudden precipitation events after the drought periods was evident in this study, with an evident peak in the SR across all land uses in February and June (Fig. 4d). Similar findings have also been reported across various ecosystems (Smith and Johnson 2004; Almagro et al. 2009, Rey et al. 2011; Matteucci et al. 2015), suggesting that the rainfall after long drought periods caused physical disruption of the soil aggregates and increased the decomposition of the OM, hence releasing more microbially derived soil CO2 (Li et al. 2018). Furthermore, rewetting of the soil releases the microbial biomass C derived from microbial death during the dry season (Emmerich 2003; Sawada et al., 2016, b; Li et al. 2018). In our study, among the land uses, the responses to these sudden precipitation events appeared to be lower in the forested sites (MFC, PFC) compared to in the cultivated or arable land uses (AF, VF); a CO2 increase of 16–21% was seen in the VF and AF compared to 9–14% in the MFC and PFC. Rey et al. (2011) also observed similar results, where the CO2 efflux was higher in degraded sites than in non-degraded sites. This buffered response of SR rates at the forest sites (PFC, MFC) could be explained by hydraulic lift (the passive movement of the water present in the lower soil layer), which maintains the fine root activity and the other soil microorganisms and microbial activity during prolonged dry conditions (Querejeta et al. 2007; Bauerle et al. 2008; Almagro et al. 2009).
Factors controlling the variation in SR
The ST and SM are usually taken as the most important factors controlling SR and can explain most of the variation. It has been well documented that more than 50% of the spatio-temporal variation in SR is governed by fluctuations in the ST and the SM content (Lloyd and Taylor 1994; Davidson et al. 2000; Zhang et al., 2013a, b; Bao et al. 2016). In our study, the regression function considering only the SM was positively correlated with the SR, accounting for approximately 90% of the seasonal variability in the SR, and appeared to be more important than the ST (Fig. 5b). Non-significant and weak nonlinear and negative relationships were found between the ST and SR in the forest sites and the AF, respectively (Fig. 5a), which explained 15–30% of the variation in the SR. This is in contrast with the well-documented strong positive exponential relationship that has been reported in previous studies, which have considered ST as the best predictor of SR (Fang and Moncrieff 2001; Cao et al. 2004; Peri et al. 2015; Rubio and Detto 2017). Rey et al. (2011), in a semi-arid steppe ecosystem, observed that the ST controlled the SR during the winter season only, and the effect disappeared at higher values, i.e., 0.5 over 20 °C. Furthermore, the SR decreased below 12–15% of the SM with greater impact at the degraded sites. Similarly, this study also observed the lowest SR during the summer (May), with high ST coinciding with low SM in the MFC, PFC, and AF. In contrast, in the VF, the increase in the SM along with the ST enhanced the SR (Fig. 4a, b). A nonlinear bell-shaped relationship between the SR and ST in the forest land uses (MFC, PFC) (Fig. 5a) suggests that the SR would have increased up to an optimal temperature (~ 28 °C in our study) when the microbial activity was high, while at a still higher ST value could have decreased the activity, hence reducing the SR (Conant et al. 2004). However, the optimal temperature for the SR varies depending upon the substrate availability and can reach up to 35 °C (Richardson et al. 2012; Liu et al. 2018). Therefore, in this ecosystem, the control of the ST on the SR would only occur for short durations, i.e., in the winter season (November–January), which suggests that the SM was the single best predictive variable for most parts of the year. However, in the VF, high SM and ST values would have favored the SMA, hence enhancing the SR rates. This is evident from the significant positive exponential relationship between the ST and SR. As concluded by the previous studies among various semi-arid ecosystems, we can also emphasize that the SM has the potential to modulate the relationship of the ST with the SR and hence is considered the single best predictive variable of SR (Conant et al. 2004; Rey et al. 2005; Jia et al. 2006; Almagro et al. 2009; Jha and Mohapatra 2011; Tucker and Reed 2016). However, with the poor correlation between the SM and ST (Table 1), it is assumed that stimulation of SR was controlled not only by the optimal SM and ST values but also by the seasonal variation in fine root growth, microbial activity, and respiration (Adachi et al. 2006; Makita et al. 2018; Wang et al. 2019). A significant positive relationship was found between the SR and SMA in the MFC, AF, and VF (Fig. 5c). This was evident in the results of the interactive models where the inclusion of the SMA along with the ST and SM improved the variation from 73 to 85% in the AF and 87 to 91% in the VF. However, in the forest land uses, the improvement in the relationship was very small, with only 83–84% and 94–95% in MFC and PFC, respectively (Table 2).
Soil respiration in different land uses
Vegetation cover and/or types have a strong influence on the belowground processes in terrestrial ecosystems (Han et al. 2014). We observed higher SR in the cultivated land uses, with ~ 14 to 32% higher values in the AF and ~ 44 to 56% higher values in the VF than in the PFC and MFC (Fig. 3a). An earlier study (Xue and Tang 2018) reported an increase of 29% in the SR during the conversion of free grazing grassland into cropland in a semi-arid agropastoral ecotone in North China and suggested that soil management activities, mainly tillage and fertilizer input, decrease the level and storage of SC and that the soil aeration enhances SMA and SOM decomposition in cropland. In this study, the SC and SN varied significantly among all land use sites and decreased by 76–82% in the AF and 52–64% in the VF compared with the PFC and MFC (Fig. 3e, f). However, no correlation was observed between the SC and the SN with the SR for any land use (Table 1). In contrast, there have been studies that reported decreases in SR with SOC content during the conversion of forest to cropland (Wang et al. 2007). These studies suggested that the intensive management activities in cultivated land uses would influence the soil structure (soil aeration and soil aggregation), microbial functions, and decomposition of SOM, which controls the soil C dynamics and alters the SR processes (Smith et al. 2008; Kravchenko et al. 2011; Fan et al. 2015). The significantly higher SMA in forest land uses (PFC, MFC) compared to that in cultivated sites (AF, VF) align with the results of other studies in different ecosystems, such as semiarid steppe, tropical water shed, forests, plantations, and degraded lands (Acosta-Martínez et al. 2007; da Silva et al. 2012; Araujo et al. 2013; Zhao et al. 2016). The low SC and SN content in the cultivated sites (Fig. 3e, f) could have limited the SOM decomposition by reducing the enzyme activity and microbial biomass, which would significantly decrease the SMA (Son et al. 2003; Allison et al., 2005). In contrast, the availability of high biomass, detritus (Nsambimana et al. 2004), and fresh OM for microbiota (Chen et al. 2005) increased the SMA in the forest soils (Araujo et al. 2013).
Furthermore, the influence of plant photosynthesis should also be considered when explaining the spatial and temporal variation in the SR in different land uses. The aboveground plant photosynthesis and the time required to transport the photosynthetic substrates from the roots to the leaves and then to the soil regulate the heterotrophic and autotrophic SR (Tang et al. 2005). Zhang et al. (2018) reported that the inclusion of recently added photosynthetic substrates and SM in SR models explained the seasonal variation in the SR-ST hysteresis relationship. In this study, the four land uses experienced similar climatic conditions (air temperature and precipitation); hence, the variation in the SR could also be explained on the basis of the differences in the vegetation types and growing seasons. Between the arable land uses, in the VF, the significant increase in the SR throughout the year that peaked in May could be related to the increased plant biomass due to the growth of C. annum. Similarly, in the AF, the early and peak growing seasons for T. aestivum (October–April) and P. vulgare (August–October) could also have contributed to the high aboveground and belowground biomass and the increased SR rates during the monsoon season and from February–March (Fig. 4d). On the other hand, during the summers (non-growing season), the bare soil in the AF with no root or shoot biomass had reduced SR in May. In the forest land uses (MFC, PFC), the high herbaceous growth during the monsoon season (growing season) could have also been attributed to the enhanced SR. This was supported by the findings suggesting that in addition to the SM and ST, the changes in plant biomass also influenced the spatial and temporal variation in the SR (Nakano and Shinoda 2010; Geng et al. 2012; Han et al. 2014).
Conclusion
Our results suggest that variations in precipitation events affect the SM levels and in turn control the SR rates in the semi-arid ecosystems of Delhi. Increased numbers of precipitation events drastically altered the SM levels and consequently resulted in higher SR rates during the monsoon season in the studied ecosystem. Furthermore, sudden rainfall events after a long drought period release C from the soil and result in an ~ 20% increase in the SR, with greater impact on the arable land uses (AF, VF). Our findings emphasize that the seasonal dynamics of the SR in semi-arid ecosystems are mainly controlled by the SM patterns and can alone explain ~ 90% of the variability. A strong positive linear fit between the SM and SR suggested that SM was the best predictor of the SR in the semi-arid ecosystems. Our study also highlighted the relevance of the SMA in SR studies, as the correlation improved from 73 to 85% in the AF and 87 to 91% in the VF when the SMA was combined with the SM and ST. Furthermore, it was inferred that intensive management activities in cultivated land use reduce the SOM content and vegetation cover and may alter the soil C balance in these ecosystems in the future.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AF:
-
Agriculture field
- BA:
-
Basal area
- C:
-
Carbon
- CO2 :
-
Carbon dioxide
- FDA:
-
Fluorescein diacetate
- MBA:
-
Mean basal area
- MFC:
-
Mixed forest cover
- PFC:
-
Prosopis juliflora forest cover
- SC :
-
Soil carbon
- SM :
-
Soil moisture
- SMA :
-
Soil microbial activity
- SN :
-
Soil nitrogen
- SR :
-
Soil respiration
- ST :
-
Soil temperature
- SOM:
-
Soil organic matter
- TD:
-
Tree density
- VF:
-
Vegetable field
References
Acosta-Martínez V, Mikha MM, Vigil MF (2007) Microbial communities and enzyme activities in soils under alternative crop rotations compared to wheat-fallow for the Central Great Plains. Appl Soil Ecol 37(1-2):41–52
Adachi M, Bekku YS, Rashidah W, Okuda T, Koizumi H (2006) Differences in soil respiration between different tropical ecosystems. Appl Soil Ecol 34(2-3):258–265
Adam G, Duncan H (2001) Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol Biochem 33(7-8):943–951
Allison VJ, Miller RM, Jastrow JD, Matamala R, Zak DR (2005) Changes in soil microbial community structure in a tallgrass prairie chronosequence. Soil Sci Soc Am J 69(5):1412–1421
Almagro M, López J, Querejeta JI, Martinez-Mena M (2009) Temperature dependence of soil respiration is strongly modulated by seasonal patterns of moisture availability in a Mediterranean ecosystem. Soil Biol Biochem 41(3):594–605
Araujo ASF, Cesarz S, Leite LFC, Borges CD, Tsai SM, Eisenhauer N (2013) Soil microbial properties and temporal stability in degraded and restored lands of Northeast Brazil. Soil Biol Biochem 66:175–181
Arora P, Chaudhry S (2017) Dependency of rate of soil respiration on soil parameters and climatic factors in different tree plantations at Kurukshetra, India. Trop Ecol 58(3):573–581
Arredondo T, Delgado-Balbuena J, Huber-Sannwalda E, García-Moya E, Loescher HW, Aguirre-Gutiérrez C, Rodriguez-Robles U (2018) Does precipitation affects soil respiration of tropical semiarid grasslands with different plant cover types? Agr Ecosyst Environ 251:218–225
Bao K, Tian H, Su M, Qiu L, Wei X, Zhang Y, Liu J, Gao H, Cheng J (2019) Stability of ecosystem CO2 flux in response to changes in precipitation in a semiarid grassland. Sustainability 11(9):2597
Bao X, Zhu X, Chang X, Wang S, Xu B, Luo C, Zhang Z, Wang Q, Rui Y, Cui X (2016) Effects of soil temperature and moisture on soil respiration on the Tibetan plateau. PLoS One 11(10):e0165212
Bauerle TL, Richards JH, Smart DR, Eissenstat DM (2008) Importance of internal hydraulic redistribution for prolonging the lifespan of roots in dry soil. Plant Cell Environ 31(2):177–186
Bilandžija D, Zgorelec Ž, Kisić I (2016) Influence of tillage practices and crop type on soil CO2 emissions. Sustainability 8(1):1–10
Black CK, Davis SC, Hudiburg TW, Bernacchi CJ, DeLucia EH (2017) Elevated CO2 and temperature increase soil C losses from a soybean-maize ecosystem. Glob Chang Biol 23(1):435–445
Butnor JR, Johnsen KH, Oren R, Katul GG (2003) Reduction of forest floor respiration by fertilization on both carbon dioxide enriched and reference 17-year-old loblolly pine stands. Glob Chang Biol 9(6):849–861
Cao GM, Tang YH, Mo WH, Wang Y, Li Y, Zhao X (2004) Grazing intensity alters soil respiration in an alpine meadow on the Tibetan plateau. Soil Biol Biochem 36(2):237–243
Champion HG, Seth SK (1968) A revised survey of the forest types of India. Govt. of India Publications, New Delhi
Chen CL, Liao M, Huang CY (2005) Effect of combined pollution by heavy metals on soil enzymatic activities in areas polluted by tailings from Pb, Zn, Ag mine. J Environ Sci 17:637–640
Chen Z, Xu Y, Fan J, Yu H, Ding W (2017) Soil autotrophic and heterotrophic respiration in response to different N fertilization and environmental conditions from a cropland in Northeast China. Soil Biol Biochem 110:113–115
Chibbar RK (1985) Soils of Delhi and their management. In: Biswas BC, Yadav DS, Maheshwari S (eds) Soils of India and their management. Fertiliser Association of India, New Delhi, pp 72–86
Conant RT, Dalla-Betta P, Klopatek CC, Klopatek JM (2004) Controls on soil respiration in semiarid soils. Soil Biol Biochem 36(6):945–951
da Silva DKA, Freitas NO, Souza RG, da Silva FSB, de Araujo ASF, Maia LC (2012) Soil microbial biomass and activity under natural and regenerated forests and conventional sugarcane plantations in Brazil. Geoderma 189-190:257–266
Darrouzet-Nardi A, Reed SC, Grote EE, Belnap J (2018) Patterns of longer-term climate change effects on CO2 efflux from bio crusted soils differ from those observed in the short term. Biogeosciences 15:4561–4573
Davidson EA, Verchot LV, Cattanio JH, Ackerman IL, Carvalho JEM (2000) Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry 48(1):53–69
Emmerich WE (2003) Carbon dioxide fluxes in a semi-arid environment with high carbonate soils. Agric For Meteorol 116:91–102
Fan LC, Yang MZ, Han WY (2015) Soil respiration under different land uses in eastern China. PLoS One 10(4):e0124198
Fang C, Moncrieff JB (2001) The dependence of soil CO2 efflux on temperature. Soil Biol Biochem 33:155–165
FSI (2017) Forest and Tree Resources in States and Union Territories, State of forest report - Delhi. Forest Survey of India, Ministry of Environment and Forests, Government of India, India
Geng Y, Wang YH, Yang K, Wang SP, Zeng H, Baumann F, Kuehn P, Scholten T, He J (2012) Soil respiration in Tibetan alpine grasslands: belowground biomass and soil moisture, but not soil temperature, best explain the large-scale patterns. PLoS One 7(4):e34968
Gu Q, Wei J, Luo S, Ma M, Tang X (2018) Potential and environmental control of carbon sequestration in major ecosystems across arid and semi-arid regions in China. Sci Total Environ 645(2):796–805
Han G, Xing Q, Luo Y, Rafique R, Yu J, Mikle N (2014) Vegetation types alter soil respiration and its temperature sensitivity at the field scale in an estuary wetland. PLoS One 9(3):e91182
Hanson PJ, Edwards NT, Garten CT, Andrews JA (2000) Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry 48(1):115–146
IPCC (2013) Climate Change 2013: The physical science basis. Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Jarvis PG, Rey A, Petsikos C, Wingate L, Rayment M, Pereira J, Banza J, David J, Miglietta F, Borghetti M, Manca G, Valentini R (2007) Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the “Birch effect”. Tree Physiol 27(7):929–940
Jha P, Mohapatra KP (2011) Soil respiration under different forest species in the riparian buffer of the semi-arid region of northwest India. Curr Sci 100(9):1412–1420
Jia B, Zhou G, Wang Y, Wang F, Wang X (2006) Effects of temperature and soil water content on soil respiration of grazed and ungrazed Leymus chinensis steppes, Inner Mongolia. J Arid Environ 67(1):60–76
Kravchenko AN, Wang ANW, Smucker AJM, Rivers ML (2011) Long-term differences in tillage and land use affect intra-aggregate pore heterogeneity. Soil Sci Soc Am J 75(5):1658–1666
Li JT, Wang JJ, Zeng DH, Zhao S, Huang W, Sun X, Hua Y (2018) The influence of drought intensity on soil respiration during and after multiple drying-rewetting cycles. Soil Biol Biochem 127:82–89
Liu Y, He N, Wen X, Xu L, Sun X, Yu G, Liang L, Schipper LA (2018) The optimum temperature of soil microbial respiration: patterns and controls. Soil Biol Biochem 121:35–42
Lloyd J, Taylor JA (1994) On the temperature dependence of soil respiration. Funct Ecol 8(3):315–323
Lou Y, Li Z, Zhang T, Liang Y (2004) CO2 emissions from subtropical arable soils of China. Soil Biol Biochem 36(11):1835–1842
Makita N, Kosugi Y, Sakabe A, Kanazawa A, Ohkubo S, Tani M (2018) Seasonal and diurnal patterns of soil respiration in an evergreen coniferous forest: evidence from six years of observation with automatic chambers. PLoS One 13(2):e0192622
Matteucci M, Gruening C, Ballarin IG, Seufert G, Cescatti A (2015) Components, drivers and temporal dynamics of ecosystem respiration in a Mediterranean pine forest. Soil Biol Biochem 88:224–235
Meena A, Bidalia A, Hanief M, Dinakaran J, Rao KS (2019) Assessment of above- and belowground carbon pools in a semi-arid forest ecosystem of Delhi, India. Ecol Process 8:8
Meena A, Hanief M, Bidalia A, Dinakaran J, Rao KS (2016) Structure, composition and diversity of tree strata of semi-arid forest community in Delhi, India. Phytomorphology 66(3-4):95–102
Miao Y, Han H, Du Y, Zhang Q, Jiang L, Hui D, Wan S (2017) Nonlinear responses of soil respiration to precipitation changes in a semiarid temperate steppe. Sci Rep 7:45782
Nakano T, Shinoda M (2010) Response of ecosystem respiration to soil water and plant biomass in a semiarid grassland. Soil Sci Plant Nutr 56:773–781
Nsambimana D, Haynes RJ, Wallis FM (2004) Size, activity and catabolic diversity of the soil microbial biomass as affected by land use. Appl Soil Ecol 26(2):81–92
Peri PL, Bahamonde H, Christiansen R (2015) Soil respiration in Patagonian semiarid grasslands under contrasting environmental and use conditions. J Arid Environ 119:1–8
Querejeta JI, Egerton-Warburton LM, Allen MF (2007) Hydraulic lift may buffer rhizosphere hyphae against the negative effects of severe soil drying in a California oak savanna. Soil Biol Biochem 39(2):409–417
Rai B, Srivastava AK (1981) Studies on microbial population of a tropical dry deciduous forest soil in relation to soil respiration. Pedobiologia 22(3):185–190
Reichstein M, Tenhunen JD, Roupsar O, Ourcival J, Rambal S, Miglietta F, Peressotti A, Pecchiari M, Tirone G, Valentini R (2002) Severe drought effects on ecosystem CO2 and H2O fluxes at three Mediterranean evergreen sites: revision of current hypotheses? Glob Chang Biol 8(10):999–1017
Rey A, Pegoraro E, Oyonarte C, Were A, Escribano P, Raimundo J (2011) Impact of land degradation on soil respiration in a steppe (Stipa tenacissima L.) semiarid ecosystem in the SE of Spain. Soil Biol Biochem 43(2):393–403
Rey A, Pegoraro E, Tedeschi V, Parri ID, Jarvis PG, Valentini R (2002) Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Glob Chang Biol 8(9):851–866
Rey A, Pepsikos C, Jarvis PG, Grace J (2005) The effect of soil temperature and soil moisture on carbon mineralization rates in a Mediterranean forest soil. Eur J Soil Sci 56(5):589–599
Richardson J, Chatterjee A, Jenerette GD (2012) Optimum temperatures for soil respiration along a semi-arid elevation gradient in southern California. Soil Biol Biochem 46:89–95
Rubio VE, Detto M (2017) Spatiotemporal variability of soil respiration in a seasonal tropical forest. Ecol Evol 7(17):7104–7116
Sawada K, Funakawa S, Kosaki T (2016) Short-term respiration responses to drying-rewetting in soils from different climatic and land use conditions. Appl Soil Ecol 103:13–21
Shen W, Jenerette GD, Hui D, Scott RL (2016) Precipitation legacy effects on dryland ecosystem carbon fluxes: direction, magnitude and biogeochemical carryovers. Biogeosciences 13:425–439
Shi W, Yan M, Zhang J, Guan J, Du S (2014) Soil CO2 emissions from five different types of land use on the semiarid Loess Plateau of China, with emphasis on the contribution of winter soil respiration. Atmos Environ 88:74–82
Sinha GN (2014) An Introduction to the Delhi Ridge. Department of Forests and Wildlife. Govt. of NCT of Delhi, New Delhi, pp Xxiv–154
Smith DL, Johnson L (2004) Vegetation mediated changes in microclimate reduce soil respiration as woodlands expand into grasslands. Ecology 85(12):3348–3361
Smith P, Martino D, Cai Z, Gwary D, Janzen H, Kumar P, McCarl B, Ogle S, O'Mara F, Rice C, Scholes B, Sirotenko O, Howden M, McAllister T, Pan G, Romanenkov V, Schneider U, Towprayoon S, Wattenbach M, Smith J (2008) Greenhouse gas mitigation in agriculture. Philos Trans R Soc B 363(1492):789–813
Son Y, Yang SY, Jun YC, Kim RH, Lee YY, Hwang JO, Kim JS (2003) Soil carbon and nitrogen dynamics during conversion of agricultural lands to natural vegetation in central Korea. Commun Soil Sci Plan 34(11-12):1511–1527
Subke JA, Inglima I, Cotrufo MF (2006) Trends and methodological impacts in soil CO2 efflux partitioning: a meta-analytical review. Glob Chang Biol 12(6):921–943
Sun Q, Wang R, Hu Y, Yao L, Guo S (2018) Spatial variations of soil respiration and temperature sensitivity along a steep slope of the semiarid Loess Plateau. PLoS One 13(4):e0195400
Tang J, Baldocchi DD (2005) Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry 73(1):183–207
Tang J, Baldocchi DD, Xu L (2005) Tree photosynthesis modulates soil respiration on a diurnal time scale. Glob. Change Biol, 11(8):1298–1304.
Tang Z, Sun X, Luo Z, He N, Sun OJ (2018) Effects of temperature, soil substrate, and microbial community on carbon mineralization across three climatically contrasting forest sites. Ecol Evol 8(2):879–881
Tucker CL, Reed SC (2016) Low soil moisture during hot periods drives apparent negative temperature sensitivity of soil respiration in a dryland ecosystem: a multi-model comparison. Biogeochemistry 128(1-2):155–169
Wang C, Wang XB, Liu DW, Wu H, Lü X, Fang Y, Cheng W, Luo W, Jiang P, Shi J, Yin H, Zhou J, Han X, Bai E (2014) Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands. Nat Commun 5:4799
Wang G, Huang W, Mayes MA, Liu X, Zhang D, Zhang Q, Han T, Zhou G (2019) Soil moisture drives microbial controls on carbon decomposition in two subtropical forests. Soil Biol Biochem 130:185–194
Wang XG, Zhu B, Wang YQ, Zheng XH (2007) Soil respiration and its sensitivity to temperature under different land use conditions. Acta Ecol Sin 27:1960–1968
Xue H, Tang H (2018) Responses of soil respiration to soil management changes in an agropastoral ecotone in Inner Mongolia, China. Ecol Evol 8(1):220–230
Zeng X, Song Y, Zhang W, He S (2018) Spatio-temporal variation of soil respiration and its driving factors in semi-arid regions of North China. Chin Geogr Sci 28(1):12–24
Zhang Q, Phillips RP, Manzoni S, Scott RL, Oishi AC, Finzi A, Daly E, Vargas R, Novick KA (2018) Changes in photosynthesis and soil moisture drive the seasonal soil respiration-temperature hysteresis relationship. Agric For Meterol 259:184–195
Zhang R, Zhao X, Zuo X, Degend AA, Shang Z, Luo Y, Zhang Y, Chen J (2019) Effect of manipulated precipitation during the growing season on soil respiration in the desert-grasslands in Inner Mongolia. Catena 176:73–80
Zhang XB, Xu MG, Sun N, Wang XJ, Wu L, Wang BR, Li D (2013a) How do environmental factors and different fertilizer strategies affect soil CO2 emission and carbon sequestration in the upland soils of southern China? Appl Soil Ecol 72(2):109–118
Zhang Y, Gu F, Liu S, Liu Y, Li C (2013b) Variations of carbon stock with forest types in subalpine region of southwestern China. For Ecol Manage 300:88–95
Zhao C, Miao Y, Yu C, Zhu L, Wang F, Jiang L, Hui D, Wan S (2016) Soil microbial community composition and respiration along an experimental precipitation gradient in a semiarid steppe. Sci Rep 6:24317
Zhao J, Li R, Li X, Tian L (2017) Environmental controls on soil respiration in alpine meadow along a large altitudinal gradient on the central Tibetan Plateau. Catena 159:84–92
Zhou L, Zhou X, Zhang B, Lu M, Luo Y, Liu L, Li B (2014) Different responses of soil respiration and its components to nitrogen addition among biomes: a meta-analysis. Glob Chang Biol 20(7):2332–2343
Acknowledgments
Authors thankfully acknowledge the two reviewers for their constructive suggestions and comments on earlier version of the manuscript.
Funding
We thank Council of Scientific and Industrial Research (CSIR; Ref No. 20-12/2009(ii) EU-IV), University Grants Commission (UGC; Ref No. 20-6/2009(ii) EU-IV) and Science and Engineering Research Board (SERB), Department of Science and Technology (DST; SR/FT/LS-59/2012), India for financial support. We also thank University of Delhi for providing Research and Development for providing grant for doctoral research program.
Author information
Authors and Affiliations
Contributions
AM proposed the idea and conducted the field sampling, data collection, laboratory analysis, data interpretation, and manuscript writing. MH carried out field sampling and data collection. DJ helped in analysis of data and edited the manuscript. KSR guided the study, interpreted the results and critically reviewed the idea. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
No existing ethics and consent of interests.
Consent for publication
NA
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Meena, A., Hanief, M., Dinakaran, J. et al. Soil moisture controls the spatio-temporal pattern of soil respiration under different land use systems in a semi-arid ecosystem of Delhi, India. Ecol Process 9, 15 (2020). https://doi.org/10.1186/s13717-020-0218-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-020-0218-0