
Abstract
Shea (Vitellaria paradoxa C.F. Gaertn.) is a multipurpose tree species indigenous to the Sudano Sahelian zone of Africa and occurs as the most abundant economic tree species in northern Ghana. The edible oil (shea butter) extracted from shea kernel is ranked as the most economic product of the species. Although fruit set and yield of shea are known to be influenced by insect pollination, the actual contribution of insect pollinators to its reproductive success has rarely been studied. This study estimated the percentage insect pollinator dependence and monetary value of insect pollination per bag of shea kernel (85 kg) in the Guinea savanna zone. Open pollination and insect exclusion treatments were applied to the flowers of 18 randomly selected matured shea trees and observed from the onset of flowering to fruit maturity. Proportion of total production value attributed to insect pollination approach was used in estimating the monetary value of pollination per bag of shea kernel with the average market price as proxy. The study revealed an insect pollinator dependence of 77% and 73% for fruit set and dry kernel yield, respectively. Mean number of fruit set per inflorescence varied significantly between insect-excluded and open-pollinated inflorescences (p < 0.05). The monetary value of insect pollination was estimated at GH₵ 73.21 (US$ 18.67) per bag of kernel as of August 2016. Shea is a high insect pollinator–dependent species and the conservation of insect pollinators would be critical to the sustainability of yield.
Similar content being viewed by others


Introduction
Insect pollinator dependence (IPD) is a theoretical metric that depicts the percentage or proportion of total crop output loss in the absence of pollination services (Breeze et al. 2016). The ratios are relevant in estimating the degree to which various plant species rely on insect pollinators for fruit/seed yield (Klein et al. 2007). Insect pollinator dependence of a crop species can differ from one region to another due to variation in soil nutrients, macroclimate as well as pest and disease status (Klein et al. 2007). It can also differ between varieties of plant species (Breeze et al. 2016). This was evident in studies that assessed the pollinator dependence of oilseed rape, where Stanley et al. (2013) recorded a pollinator dependence of 30%, but Bartomeus et al. (2014) recorded 20% for two varieties of the same crop.
The positive effect of pollination on quantity and quality of plant yield also translates into higher economic/monetary returns (Zebrowska 1998). Insects are known to contribute to an estimated 10% of the economic value of the world’s food produce (Chaplin-Kramer et al. 2014) through the enhancement of commercial value. Like pollinator dependence, economic value of pollination could vary from one region to another due to differences in food prices, production, and labor cost (Breeze et al. 2016). However, most variations in economic value estimates of pollination are attributed to differences in methodological approaches (Burgett et al. 2004).
Some of the common approaches used in economic/monetary value estimates of pollination include total production value, proportion of total production value attributable to insect pollinators (Morse & Calderone 2000; Losey & Vaughan 2006), cost of replacement (Allsopp et al. 2008), and direct-managed pollination value approach (Burgett et al. 2004). Variations in economic value estimates of insect pollination do not only reflect differences in methods but also reflects a paucity of accurate information on pollination. This highlights the need for extensive research to provide better estimates of the value of pollination (Klein et al. 2007).
Monetary estimates of pollination as an ecosystem service could help in justifying resource allocation for conservation purposes since major human decisions are largely driven by financial implications (Curtis 2004). These estimates could inform trade-offs between the conservational value of insect habitats and the other alternative land uses. Pollinator conservation is critical to the sustainability of food production and biodiversity owing to the high dependence of tropical plants on animal-mediated pollination services. According to Ollerton et al. (2011), about 94% of tropical plants depend on animal pollination for fruit/seed set.
Shea (Vitellaria paradoxa) is an insect pollinator–dependent tropical plant (Hall et al. 1996; Okullo 2004) which occurs naturally in Sub-Saharan Africa (SSA) especially arid and semi-arid zones (Naughton et al. 2015). It is often retained on farmlands for economic, medicinal, cultural, nutritional, and ecological purposes (Yaro 2008; Bayala et al. 2013). According to Glew and Lovett (2014), it contributes to the livelihoods of an estimated 16.2 million shea nut collectors in SSA. In some seasons, income from shea nut could constitute about 12% of rural household income (Pouliot & Treue 2012). According to Elias et al. (2006), shea nut processing and commercialization are two of the few plant industries primarily controlled by women in SSA which contribute to the economic empowerment of women. Aside local uses, there is an increasing demand for shea butter in confectionary, cosmetic, and pharmaceutical industries at the international level (Alander 2004; Teklehaimanot 2004; Rousseau et al. 2015).
However, the sustainability of the economic and ecological benefits of V. paradoxa could be affected by the availability of pollination services. It is an insect-pollinated plant (Hall et al. 1996) in which floral structures and sequence of events in anthesis are known to favor pollen out-crossing (Okullo 2004). Stingless and honeybees have been identified as the primary pollinators of V. paradoxa in most countries including Ghana, Burkina Faso, and Uganda (Okullo 2004; Kwapong 2014; Lassen et al. 2018). The studies of Okullo (2004) and Lassen et al. (2018) reported reduced fruit set in shea when insect pollination and bagged inflorescences were compared. However, information on insect-pollinated plants will remain incomplete until we estimate the quantity of shea yield dependent on insect pollination as well as the monetary value of insect pollination services. This study aimed to estimate the percentage of shea yield dependence on insect pollination and monetary value of insect pollination services.
Materials and methods
Study area
The study was conducted in cultivated fields of the Zini community in the Sissala West District of the Upper West Region, Ghana. The District lies approximately between longitude 2°13´ W to 2° 36´ W and latitude 10° 00´ N to 11° 00´ N (Sissala West District Assembly, 2010) with experimental site 1 located at 10° 50′ 00.0″ N and 2° 22′ 57.2″ W, whilst site 2 was located at 10° 52′ 14.1″ N and 2° 24′ 48.8″ W (Fig. 1).

Map of Sissala West district showing the location of the experimental sites (source: adapted from GSS, 2014)
The area records a unimodal rainfall pattern with an average annual rainfall of 1127 mm (SARI 2016). Mean monthly minimum and maximum temperatures are 22 °C and 35 °C, respectively, whilst the mean annual temperature is 28 °C (GSS, 2014). Relative humidity fluctuates between 70% and 90% in the rainy season but can drop to as low as 20% in the dry season (SWDA 2010).
The vegetation is generally grassland interspersed with drought-resistant perennial woody species. The common woody species include shea (Vitellaria paradoxa), dawadawa (Parkia biglobosa), baobab (Adansonia digitata), neem (Azardirachta indica), and accasia (Faidherbia albida). Some common herbaceous plant species in Guinea savanna include tridax (Tridax procumbens), thatch grass (Andropogon pseudapricus), Elephant grass (Pennisetum purpureum), and pig weed (Boerhavia diffusa) (Ziblim et al. 2015).
Selection of experimental sites and trees
A reconnaissance survey was conducted on farmlands of Zini to identify suitable sites for the experiment considering accessibility to site, shea tree density, duration of continuous cultivation, and the cooperation of the land owner. Five medium-cultivated parklands (lands that were cultivated continuously for 6–10 years) were identified by asking farmers on the duration of continuous cultivation to ensure uniformity of landuse on the experimental sites. A simple random sampling procedure was then used in randomly selecting two sites out of five potential sites for the experiment. Again, to avoid potential bias due to specific farmers’ land management practices on pollinators and also to increase independence of sampling sites, the study considered a minimum distance of 1 km between experimental sites as prescribed by Stanley et al. (2013).
In selecting experimental trees, three 50 × 50 m plots were systematically laid in each site by laying the first plot at a random location, whilst subsequent plots were laid at a regular distance of 100 m apart. Three flowering matured (DBH > 30 cm) shea trees were selected from each plot using a simple random sampling procedure, thus, a total of nine (9) trees per site. Matured trees were used to avoid potential bias associated with inconsistent fruiting (fruit size and quantity) of juvenile (10–15 years) shea trees (Sanou et al. 2004).
Research design
Experimental treatments were arranged in a randomized complete block design with selected branches representing blocks. Three branches (blocks) were randomly selected from each tree and experimental treatments applied to one inflorescence in a block. The experimental treatments were as follows:
Open pollination treatment (control)—tagged inflorescences that were un-manipulated and left exposed to pollination by natural agents in the environment throughout the flowering phase (Fig. 2).
Insect exclusion treatment—inflorescences bagged with tulle netting (1.2 mm diameter mesh) to restrict insect pollinators from accessing the flowers throughout the flowering season but allowing wind pollination as described in Jacobs et al. (2009). Inflorescences were bagged as soon as flower buds were observed (Fig. 3). The bags were however removed immediately after the end of the flowering phase to enable continued plant growth. All experimental trees and inflorescences were given unique labels to enable continuous data collection on data variables.

Open pollination

Insect exclusion
Data collection
The number of flowers/flower buds in each experimental inflorescence was counted at the onset of flowering. Also, the number of fruit set per inflorescence was recorded weekly from the onset of fruiting to fruit maturity (first ripe fruit fall from a tree). Matured fruits were harvested by hand plucking and fruit weight per inflorescence was recorded with an electronic scale. Fruits were then depulped and fresh nut weight per inflorescence was recorded. The fresh nuts were sun dried for 5 weeks before dry nut weight per inflorescence was taken. Nut shells were cracked open to remove kernels, after which kernel weight per inflorescence was recorded.
The purchasing price per bag of shea kernel in August 2016 was sought from five shea marketing companies as well as the Zini community market. These prices were used in calculating the average price per bag of shea kernel as a proxy for the monetary value of shea kernel as of August 2016. The purchasing prices of August were used because most women sell out their shea kernels around this period. The five shea marketing companies contacted were Star Shea Company Limited, Savannah Fruit Company Limited, Mother’s Shea Company Limited, OLAM Company Limited, and Kasajan Company Limited.
Data analysis
A simple t test was used in comparing average fruit/kernel yield per inflorescence between open-pollinated and insect-excluded inflorescences with a p value threshold set at 0.05. Genstat software version 17 and Microsoft Excel were used for data analysis.
Insect pollinator dependence (IPD) was calculated following the approach of Klein et al. (2007) by estimating the magnitude of fruit set or kernel yield (weight) difference comparing experimental treatments with and without insect pollinators. Based on this approach, insect pollinator contribution was calculated as the fruit set/kernel yield attributable to insect pollinator interaction with flowers expressed in percentage. Insect pollinator dependence (IPD) was estimated separately for fruit set and kernel yield with the following formulae:
where FSOP is the total number of matured fruits produced from the open pollination treatment in a site, FSIE is the total number of matured fruits produced from the insect exclusion treatment in a site, KWOP is the total dry weight of kernels produced from open pollination treatment in a site, and KWIE is the total dry weight of kernels produced from insect exclusion treatment in a site.
The proportion of total production value attributable to insect pollinators approach was used to estimate monetary value of insect pollination because hand/mechanized and managed pollination services were not practiced in the area. Therefore, monetary value of insect pollination was estimated per bag of shea kernel (85 kg) as the monetary value of kernel yield (kg) attributable to insect pollination.
where QAI is the quantity (kg) of kernel attributable to insect pollination, MVI is the monetary value of insect pollination (GH₵), and UP is the unit price of 1 kg of shea kernel.
Results and discussion
Insect pollinator dependence of V. paradoxa
The insect pollinator dependence (IPD) of shea was estimated at 77% for fruit set and 73% for dry kernel yield (weight). Thus, the exclusion of insect pollinators from accessing shea flowers resulted in a 77% decrease in the number of fruit set and 73% decrease in dry kernel yield (weight) (Tables 1 and 2). IPD also varied slightly between the two sites, with site 1 recording less fruit set and kernel yield dependence on insect pollination.
The percentage pollinator dependence recorded in this study suggests that V. paradoxa is a high pollinator-dependent species based on Klein et al. (2007) categorization. About 18.7% of crops grown in developing countries fall within this category of high animal pollinator dependence (Alizen et al. 2009). Variation in insect pollinator dependence between experimental sites is not unique to this study. Carr & Davidar (2015) also reported variations in pollinator dependencies between different study sites. This could be attributed to differences in microclimatic conditions between two sites especially soil conditions. Moreover, variation in genotypic characteristics between trees could have equally contributed to these differences between sites since trees in a particular site are most likely to share similar traits. Breeze et al. (2016) indicated that pollinator dependency could vary between crop varieties of the same species (Breeze et al. 2016). Similar influence of crop variety on insect pollinator dependence occurred in oil seed rape comparing the findings of Stanley et al. (2013) and Bartomeus et al. (2014).
The influence of these extraneous variables on plant yield could limit the generalization of estimated insect pollinator dependency of a plant. This would therefore require an extensive study across a larger shea-growing area to provide wholistic estimates of insect pollinator dependency of shea. There is currently a paucity of information on insect pollinator dependence of shea. This stresses the recommendations of Cook et al. (2007) and Klein et al. (2007) for an intensive review of insect pollinator dependence of commercially important crops of the world.
The assertion that insect pollinator-dependent plants represent a relatively small volume of world staple food consumption seems to be one of the reasons for this information paucity. However, Richards (2001) and Ghazoul (2005) argued that high pollinator-dependent species provide disproportionately large economic returns to the local and international market. For instance, although shea does not contribute directly to global staple food consumption, it contributes to the livelihoods of an estimated 16.2 million people in shea-growing areas of Sub-Saharan Africa. This assertion tends to agree with Alizen et al. (2009) that valuing an insect-pollinated plant solely on the bases of its contribution to global staple food consumption could obscure local relevance and tend to be deceptive. Some insect pollinator-dependent species are valued five times higher than non-pollinator-dependent species (Gallai et al. 2009).
Influence of insect pollination on fruit production and kernel yield
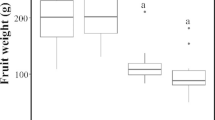
The mean number of matured fruits produced per inflorescence was significantly higher in open-pollinated inflorescences as compared with insect-excluded inflorescences (p < 0.05) in both sites (Fig. 4). Mean number of matured fruit set per inflorescence also varied significantly between experimental sites in open-pollinated inflorescences (p < 0.05) with site 1 producing a mean of 2 fruits per inflorescence, whilst site 2 recorded 0.85 per inflorescence. However, the number of fruit set per inflorescence did not vary significantly between sites in the insect exclusion treatment (p > 0.05).

Mean number of matured fruits produced per inflorescence in the Guinea savanna zone of Ghana
The significant difference in matured fruit set per inflorescence between open-pollinated and insect-excluded flowers agrees with Kwapong (2014) who reported that V. paradoxa relies primarily on insects especially bees for pollination services. The yield of V. paradoxa can therefore be influenced significantly by the availability of insect pollinators (Okullo 2004). In the study of Lassen et al. (2018), the exclusion of larger bees (honeybee) from accessing shea flowers resulted in 35% reduced fruit yield which indicates high dependence on honeybees for pollination.
The internal colors of the inflorescence bract and the odoriferous yellow flowers of V. paradoxa were all noted as major qualities of insect pollinated plants (Okullo 2004). Moreover, V. paradoxa pollen is known to be copious and sticky in nature (Yidana 1994; Hall et al. 1996) and therefore adheres to insect visitors of shea flowers. The fruit set recorded in the insect-excluded inflorescences could be attributed to wind pollination (self- and cross-pollination). Shea produces hermaphrodite flowers (Okullo 2004) which could have facilitated self-pollination
Owing to the high insect pollinator dependence of V. paradoxa, the occurrence of self-pollination could be an adaptation response of shea to the limited pollination services in shea parklands. This can be supported by the findings of Yidana (2004) who reported pollen limitation in V. paradoxa. However, fruit production in insect-excluded treatment is not limited to shea; flowers of oil seed rape still produced some few seeds when insects were excluded from pollination (Bommarco et al. 2012; Stanley et al. 2013). The significantly low fruit set recorded under insect exclusion agrees with Free (1993) who reported that the contribution of wind pollination is insignificant in deciduous fruit crops that require insect vectors for pollination.
The mean weight of dry kernels produced per inflorescence showed a significant difference between treatments (p < 0.05). Open-pollinated inflorescences produced kernels with a significantly higher weight as compared with the insect-excluded inflorescences in both experimental sites (Fig. 5). This significant difference in mean weight agrees with Lassen et al. (2018) and Stout et al. (2018) who recorded heavier nuts under open pollination as compared with bagged inflorescences. This could be due to the ability of pollination to mediate the production of growth hormones in plants (Klatt et al. 2013). For instance, in strawberry, fertilized achenes mediated the production of auxine which intend to induce the accumulation of gibberellic acid (Csukasi et al. 2011).

Mean weight of dry shea kernels produced per inflorescence in the Guinea savanna zone of Ghana
These hormones facilitate fruit growth by enhancing cell progeny and size which results in an increased fruit/seed weight (Roussos et al. 2009). Some of the fruits produced from open pollination had two solitary seeds. The presence of two seeds in some fruits could have also contributed to the significantly higher weight under open pollination.
The positive effect of pollination on yield (weight) is not limited to shea; this phenomenon was reported in other plants. For instance, insect pollinated strawberry fruits were found to be 11% heavier than wind-pollinated fruits and 30% heavier than self-pollinated fruits (Klatt et al. 2013). Stanley et al. (2013) reported oil seed rape produced from open pollination as having significantly higher seed weight per pod compared with insect-excluded pods. Similarly, coffee and blueberry recorded an increased fruit set and size under insect pollination services as compared with insect exclusion (Klein et al. 2003; Isaacs and Kirk 2010; Klatt et al. 2013).
Monetary value of insect pollination per bag of shea kernel
The purchasing price per bag of dry shea kernel varied from one company to another (Table 3). The price of a bag of shea kernel ranged from GH₵ 90.00 (US$ 22.95) to GH₵ 105.00 (US$ 26.78) with an average price of GH₵ 100.00 (US$ 25.51). This was used as a proxy for the total monetary value of a bag of kernel as of August 2016.
The study estimated the quantity of shea kernel yield attributable to insect pollination at 62.05 kg per bag (85 kg) with a unit price of a kilogram at GH₵ 1.18. Therefore, the monetary value of shea kernel weight (62.05 kg) attributable to insect pollination was GH₵ 73.21 (US$ 18.67) which was used as a proxy for the monetary value of insect pollination (Table 4).
The monetary value of insect pollination in site 2 was higher than that of site 1 because of the variation in insect pollinator dependence. This finding confirms the assertion of Zebrowska (1998) that an increase in yield does not only reflect productivity but translates into substantial amount of income for crop farmers. Estimates of economic value of pollination can vary from one region to another in response to produce prices, production cost, labor, and other factors of production (Breeze et al. 2016).
Although Allsopp et al. (2008) suggests a more accurate estimation of economic value of pollination, services should consider the cost of replacement of wild pollinators with managed ones or cost of replacement with mechanical pollination services. There currently exist neither managed bee keeping nor mechanized pollination in shea parklands. Therefore, all estimates were based on the economic value of the quantity of the response variable (kernel yield) attributed to insect pollination.
Unfortunately, quantity of kernel yield might not represent monetary value completely because economic value also depends on the quality of yield (Ferguson & Watkins 1992). For instance, the contribution of insect pollination to shelf life alone added US$ 0.32 billion to the commercial value of strawberry. Yield quality was however not considered in this study, but studies on other insect-pollinated plants showed a positive effect of pollination on quality of fruit/seed yield. For instance, Bartomeus et al. (2014) reported higher oil content with less chlorophyll in oil seed rape exposed to open pollination as compared with seeds that were produced from insect-excluded flowers. Strawberry that was adequately pollinated by honeybees produced heavier and firmer fruits with less sugar-acid ratio and longer commercial shelf life as compared with wind-pollinated plants (Klatt et al. 2013). Therefore, subsequent studies on monetary value of insect pollination in shea should consider butter quality owing to the influence of pollination on yield quality in other plants.
Conclusion
Insect pollination significantly influenced fruit set, accounting for 73% of fruit set in shea. This high insect pollinator contribution to shea yield translates into higher monetary value for shea nut collectors. Therefore, a decline in insect pollinators will not only affect the yield of V. paradoxa but will also translate into reduced income for the rural population who depends on shea for livelihood. Subsequent estimates of the monetary value of pollination should consider the influence of pollination on the quality of fruit and shea kernel as well as shea butter properties.
Availability of data and materials
The datasets generated and analyzed during the current study are not publicly available but from the corresponding author on reasonable request
Change history
09 January 2020
In the original publication of this article (Nasare et al. 2019), the ‘₵’ in Ghanaian currency symbol (GH₵) was omitted in below content in the pdf version of the article. It should be ‘GH₵’.
Abbreviations
- MVB:
-
Monetary value of a bag of shea kernel
- UP:
-
Unit price of a kilogram of shea kernel
- QAI:
-
Quantity (kg) of kernel attributable to insect pollination
- MVI:
-
Monetary value of insect pollination (GH₵/US$)
- IPD:
-
Insect pollinator dependence
- FSOP:
-
The total number of matured fruits produced from the open pollination treatment
- FSIE:
-
The total number of matured fruits produced from the insect exclusion treatment
- KWOP:
-
The total dry weight of kernels produced from open pollination treatment
- KWIE:
-
The total dry weight of kernels produced from insect exclusion treatment
References
Alander J (2004) Shea butter - a multifunctional ingredient for food and cosmetics. Lipid Technol 16:202–216
Alizen MA, Garibaldi LA, Cunningham SA, Klein AM (2009) How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann Bot 103:1579–1588
Allsopp MH, de Lange WJ, Veldtman R (2008) Valuing insect pollination services with cost of replacement. PLoS ONE 3(9):e3128
Bartomeus I, Potts SG, Steffan-Dewenter I, Vaissière BE, Woyciechowski M, Krewenka KM, Tscheulin T, Roberts SPM, Szentgyörgyi H, Westphal C, Bommarco R (2014) Contribution of insect pollinators to crop yield and quality varies with agricultural intensification. PeerJ 2:328.
Bayala J, Sanou J, Teklehaimanot Z, Kalinganire A, Ouedraogo SJ (2013) Parklands for buffering climate risk and sustaining agricultural production in the Sahel of West Africa. Curr Opin Environ Sustain 6:28–34
Bommarco R, Marini L, Vaissi’ere BE (2012) Insect pollination enhances seed yield, quality, and market value in oilseed rape. Oecologia 169(4):1025–1032
Breeze TD, Gallai N, Garibaldi LA, Li XS (2016) Economic measures of pollination services: shortcomings and future directions. Trends Ecol Evol 31:927–939
Burgett M, Rucker RR, Thurman W (2004) Economics and honeybee pollination markets. Am Bee J 144:269–271
Carr SA, Davidar P (2015) Pollinator dependency, pollen limitation and pollinator visitation rates to six vegetable crops in southern. Indian J Pollination Ecol 16(8):51–57
Chaplin-Kramer R, Dombeck E, Gerber J, Knuth KA, Mueller ND, Mueller M, Ziv G, Klein AM (2014) Global malnutrition overlaps with pollinator dependent micronutrient production. Proc R Soc B Biol Sci 281:20141799
Cook DC, Thomas MB, Cunningham SA, Anderson DL, De Barro PJ (2007) Predicting the economic impact of an invasive species on an ecosystem service. Ecol Appl 17:1832–1840
Csukasi F, Osorio S, Gutierrez JR, Kitamura J, Giavalisco P, Nakajima G, Fernie AR, Rathjen JP, Botella MA, Valpuesta V, Medina-Escobar N (2011) Gibberellin biosynthesis and signalling during development of the strawberry receptacle. New Phytol 191:376–390
Curtis IA (2004) Valuing ecosystem goods and services: a new approach using a surrogate market and the combination of a multiple criteria analysis and a Delphi panel to assign weights to the attributes. Ecol Econ 50:163–194
Elias M, Bayala J, Dianda M (2006) Impediments and innovations in knowledge sharing: the case of the African Vitellaria paradoxa sector. Knowl Manag Develop J 2(1):52–67
Ferguson LB, Watkins CB (1992) Crop load affects mineral concentrations and incidence of bitter pit in ‘Cox’s Orange Pippin’ apple fruit. J Am Soc Hortic Sci 117:373–376
Free JB (1993) Insect pollination of crops. Academic Press, London
Galla N, Salles JM, Settele J, Vaissière BE (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol Econ 68:810–821
Ghana Statistical Service (GSS) (2014) Population and housing census. district analytical report, Sissala West District
Ghazoul J (2005) Buzziness as usual? Questioning the global pollination crisis. Trends Ecol Evol 20:367–373
Glew D, Lovett PN (2014) Life cycle analysis of shea butter use in cosmetics: from parklands to product, low carbon opportunities. J Cleaner Produc 68:73–80
Hall JB, Aebischer PO, Tomlinson HF, Osei-Amaning E, Hindle JR (1996) Vitellaria paradoxa: a monograph. School of Agricultural and Forest Sciences, University of Wales, Bangor, UK.105 pp.
Isaacs R, Kirk AK (2010) Pollination services provided to small and large high bush blueberry fields by managed bees. J App Ecol 47:841–849
Jacobs JH, Clark SJ, Denholm I, Goulson D, Stoate C, Osborne JL (2009) Pollination biology of fruit-bearing hedgerow plants and the role of flower-visiting insects in fruit-set. Ann Bot 104:1397–1404
Klatt BK, Holzschuh A, Westphal C, Clough Y, Smite I, Pawelzik E, Tscharntke T (2013) Bee pollination improves crop quality, shelf life and commercial value. Proc R Soc B Biol Sci 281:20132440
Klein AM, Steffan-Dewenter I, Tscharntke T (2003) Fruit set of highland coffee increases with the diversity of pollinating bees. Proc R Soc B Biol Sci 270:955–961
Klein AM, Vaissière BE, Cane JH, Steffan-Dewenter I, Cunningham SA (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc B Biol Sci 274:303–313
Kwapong P (2014). Pollination requirement of shea butter tree—Vitellaria paradoxa in West Africa. University of Cape Coast, Ghana. http://www.aratishea.com/Publica tion/Global-Shea-Ghana-2011-en/Kwapong-Pollinationreq.-of-Shea.pdf. Accessed 30 December, 2016.
Lassen KM, Nielsen LR, Lompo D, Dupont YL, Kjær ED (2018) Honey bees are essential for pollination of Vitellaria paradoxa subsp. paradoxa (Sapotaceae) in Burkina Faso. Agroforest Syst 92:23–34
Losey JE, Vaughan M (2006) The economic value of ecological services provided by insects. BioScience 56:311–323
Morse RA, Calderone NW (2000) The value of honeybees as pollinators of U.S. crops in 2000. Bee Cult 128:1–15
Naughton CC, Lovett PN, Mihelcic JR (2015) Land suitability modelling of shea (Vitellaria paradoxa) distribution across sub-Saharan Africa. Appl Geogr 58:217–227
Okullo JBL (2004) Vitellaria paradoxa in Uganda: population structures and reproductive characteristics. Dissertation, University of Wales. Bangor, UK.
Ollerton J, Winfree R, Tarrant S (2011) How many flowering plants are pollinated by animals? Oikos 120:321–326
Pouliot M, Treue T (2012) Rural people’s reliance on forests and the non-forest environment in West Africa: evidence from Ghana and Burkina Faso. World Development 43:180–193
Richards AJ (2001) Does low biodiversity resulting from modern agricultural practices affect crop pollination and yield? Ann Bot 88:165–172
Rousseau K, Gautier D, Wardell DA (2015) Coping with the upheavals of globalization in the shea value chain: the maintenance and relevance of upstream shea nut supply chain organization in western Burkina Faso. World Development 66:413–427
Roussos PA, Denaxa NK, Damvakaris T (2009) Strawberry fruit quality attributes after application of plant growth stimulating compounds. Sci Hortic 119:138–146
Savannah Agricultural Research Institute (SARI) (2016). Annual Report for the year 2016. Nyankpala.
Sanou H, Kambou S, Teklehaimanot Z, Dembélé M, Yossi H, Sibidou S, Djingdia L, Bouvet JM (2004) Vegetative propagation of Vitellaria paradoxa by grafting. Agroforest Syst 60:93–99
Sissala West District Assembly (SWDA) (2010) District medium term development plan, unpublished.
Stanley DA, Gunning D, Stout JC (2013) Pollinators and pollination of oilseed rape crops (Brassica napus L.) in Ireland: ecological and economic incentives for pollinator conservation. J Insect Conserv 17:1181–1189
Stout JC, Bruijin D, Delaney A, Doke DA, Gyimah T, Kamano F, Kelly R, Lovett P, Marshall E, Nana A, Nasare LI, Nombre I, Roberts J, Tankoano P, Tayleur C, Thomas D, Vickey J, Kwapong P (2018) Insect pollination improves yield of shea (Vitellaria paradoxa subsp. paradoxa) in the agroforestry parklands of West Africa. J Pollination Ecol 22(2):11–20
Teklehaimanot Z (2004) Exploiting the potential of indigenous agro-forestry trees: Parkia biglobosa and Vitellaria paradoxa in sub-Saharan Africa. Agroforest Syst 61:207–220
Yaro JN (2008) An examination of theories on savannasation and the peasant-environment debate. West Afr J Appl Ecol 13:3–16
Yidana JA (1994) Pollination of shea trees. Cocoa Research Institute, Ghana, Annual Report 1992/1993
Yidana JA (2004) Progress in developing technologies to domesticate the cultivation of shea tree (Vitellaria paradoxa L.) in Ghana. Agric Food Sci J Ghana 3:249–267
Zebrowska J (1998) Influence of pollination modes on yield components in strawberry (Fragariax ananassa Duch). Plant Breeding 117:255–260
Ziblim AI, Abdul-Rasheed S, Aikins TK (2015) Forage species used by livestock in the Kumbungu district of the northern region, Ghana. UDS Int J Development 1(1):18–29
Acknowledgements
This work was partly funded by BirdLife International and the Ghana Wild Society. We wish to express our gratitude to Mr. Yussif Jibril for the assistance provided during field data collection. We also wish to acknowledge the farmers for allowing us to use their farmlands for the experiment.
Funding
The data collection was partly funded by Birdlife International.
Author information
Authors and Affiliations
Contributions
LIN did the data collection, entry, analysis and wrote the first draft; PKK designed the experiment; and DAD wrote the introduction. All authors helped in writing and editing the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interest
The authors declare that they have no competing interests.
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Nasare, L.I., Kwapong, P.K. & Doke, D.A. Insect pollinator dependence of shea (Vitellaria paradoxa C.F. Gaertn.) in the Guinea Savanna zone of Ghana. Ecol Process 8, 48 (2019). https://doi.org/10.1186/s13717-019-0202-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-019-0202-8