
Abstract
Key message
Bird assemblages in planted spruce (Picea abies L.) forests were compositionally distinct from those in naturally regenerated mixed stands. Despite rapid stand development on productive soils, even-aged silviculture cannot host most old-forest species. However, their habitat quality could be supported in landscape mosaics with retention forestry and set-asides.
Context
The most productive forest lands have naturally the richest bird assemblages but tend to be also most intensively managed. Sustainable solutions to this conflict are unclear.
Aim
To assess bird assemblages and their successional dynamics in planted Norway spruce (Picea abies L.) stands compared to naturally developing stands.
Methods
We mapped breeding bird assemblages in forty 5-ha plots on highly productive soils in Estonia. The plots included sets of naturally regenerated and planted stands, and (as successional endpoints) clear-cuts and old stands.
Results
Planted stands had fewer bird species and pairs than naturally regenerated stands; the latter having a species composition resembling late-successional deciduous-dominated stands. Importantly, the species composition in mature spruce plantations converged toward the composition observed in late-successional conifer-dominated stands. Downed dead wood, stand age, deciduous trees, and stock density were the most significant stand characteristics shaping bird assemblages.
Conclusion
The habitat value of established spruce plantations can be primarily improved by allowing for some deciduous trees and gaps with deciduous undergrowth. At the landscape scale, the bird diversity of even-aged systems would be enhanced by multi-scale applications of retention forestry—from retention trees to old-growth set-asides.
Similar content being viewed by others


1 Introduction
Bird populations are becoming widely used indicators for assessing the state of forest biodiversity (Gregory and van Strien 2010; Voříšek et al. 2020; Oettel and Lapin 2021). At the assemblage scale, forest birds can reflect alterations of stand structure (LaManna and Martin 2016), critical loss of old-forest habitats in landscapes (Schmiegelow and Mönkkönen 2002; Roberge et al. 2018), and broad-scale habitat turnover related to intensive production forestry (Lamanna and Martin 2016, Roberge et al. 2018). While the general development of bird assemblages after clear-cutting or salvage logging is well understood (Roberge et al. 2018; Georgiev et al. 2020; Thorn et al. 2020), its alternative successional trajectories and their underlying forest management decisions have been seldom explicitly studied.
In Europe, late-successional hemiboreal (boreo-nemoral) mixed forests constitute a hotspot of bird diversity where the species pools of boreal and temperate forests meet (Nilsson 1997; Roberge et al. 2018). Notably on fertile soils, the mixtures of coniferous and deciduous trees provide structurally heterogeneous conditions there (Lõhmus and Kraut 2010; Põldveer et al. 2020), supporting more diverse bird assemblages than pure deciduous or conifer stands (Nilsson 1997; Lõhmus et al. 2016). In old-growth stands, a large dead-wood supply provides additional resources for many species (Nilsson 1979a; Rosenvald et al. 2011; Bergner et al. 2015). Although most European hemiboreal forests have a centuries-long silvicultural history, intensification of timber production continues to raise the issues of ecologically sustainable management regimes (Felton et al. 2020), including a response to regional declines in forest bird abundance (Nellis and Volke 2019). This raises the issue of ecologically sustainable management regimes in these forests, specifically the consequences of further intensification of the even-aged systems (Felton et al. 2020).
In the even-aged systems, a major approach is to plant and manage clearcuts as economically beneficial monocultures; in Northern Europe mostly as planted Scots pine (Pinus sylvestris L.) and Norway spruce (P. abies) stands (Spiecker 2003; Roberge et al. 2018). Compared with natural forests, spruce plantations have a particularly homogenous and simplified stand structure (Nilsson 1979b; Swanson et al. 2011; Bergner et al. 2015), causing local reductions in bird species richness (Gjerde and Saetersdal 1997, Felton et al. 2011) and a loss of some specialist species across landscapes (MacKay et al. 2014; Lindbladh et al. 2019). The bird diversity in spruce plantations can be increased by promoting the abundance of deciduous trees (Lindbladh et al. 2017; Felton et al. 2021; Kebrle et al. 2021; Vélová et al. 2021), dead wood (Lindbladh et al. 2019), and large trees (Kebrle et al. 2021). However, the plantation impacts may be context dependent. For example, a small share of spruce plantations may add diversity in homogeneous Scots pine (P. sylvestris) dominated landscapes (Gjerde and Saetersdal 1997), while adjacent deciduous stands can reduce bird richness in the plantations (Vélová et al. 2021).
The current study investigates post-clearcut bird assemblages in the most productive hemiboreal sites, where natural succession would involve a mixture of deciduous tree species, a later development of the shade-tolerant Norway spruce and, possibly, another late-successional shift to shade-tolerant deciduous stands (Liira and Sepp 2009; Lõhmus and Kraut 2010). Such compositional changes are eliminated in spruce plantations, together with an impoverished layer structure and dead wood abundance. Despite such an obvious divergence in forest successional pathways, there is a lack of comparative studies on the successional development of bird assemblages depending on management scenarios.
In this study, our main focus is the parallel development of bird assemblages in planted versus naturally regenerated stands, between the reference conditions in clearcuts and late-successional assemblages. We ask three questions: (I) how does the origin (spruce plantation vs naturally regenerated stand) affect bird assemblages along with the succession? In general, maturing plantations may retain a species-poorer and sparser bird assemblage (Sweeney et al. 2010; MacKay et al. 2014), but another question is whether, and at which rate, they compositionally converge toward natural stand conditions. This could imply cost-effective restoration for biodiversity either through setting aside plantations or harvesting and replacing these with early successional natural regeneration. (II) Which forest structural elements affect the post-clearcut bird assemblage development? It is well known that several key structural characteristics develop along with the stand age, e.g., dead wood, large trees and vertical stratification (Fuller 2003; Roberge et al. 2018; Klein et al. 2020). An advanced question for management is, however, their contribution independent of age, and depending on the regeneration pathway (e.g., given that plantations lack certain tree species). (III) How are the characteristic bird species of old forests represented across management types, and are resident species more likely to belong to this group? Previous studies have indicated that high site productivity accelerates also the development of old-forest species assemblages (e.g., Nilsson 1997; Lõhmus et al. 2020; 2021). We thus expect their presence already prior to the old-growth stage.
2 Materials and methods
2.1 Study area and sampling design
The study was conducted in the mainland of Estonia, between 57°32′–59°12′ and 23°50′–27°5′. The region belongs to the non-oceanic section of the hemiboreal vegetation zone (Ahti et al. 1968); the mean air temperature is 16.5 °C in July and 6.5 °C in February; the average annual precipitation is 650–750 mm. Forests cover about half of the Estonian area; 46% of the forests belong to the State Forest Management Centre; the average stand age is 55 years. (Raudsaar and Valgepea 2020). Clear-cutting is the dominant approach to timber production in the country.
Our study plots (N = 40) were located in a recently established forest reserve network (see Lõhmus et al. 2020). Before protection, the areas were under intensive management, so that only 7% of the network area was covered with stands > 100 years old. The rest of the area comprised open clear-cuts (10%), and regenerating stands of different age. Importantly for this study, 45% of the network area were artificially regenerated stands, mostly of Norway spruce (Lõhmus et al. 2020).
We sampled five ‘stand types’ that were distinguished according to stand age, origin, and management history: (1) old deciduous-dominated stands and (2) old conifer-dominated stands (> 90 years age), with no thinning or sanitary cutting in the last 20 years (6 and 4 stands, respectively); (3) 10 mixed natural stands of age 30 to 70 years; (4) 10 artificially cultivated spruce stands of age 30 to 70 years; and (5) 10 recent clear-cuts either without or < 20 years old tree layer (Pass et al. 2022). Stand selection was based using the state forest survey and cuttings databases (provided by the Estonian Environmental Agency and the Estonian Environmental Board).
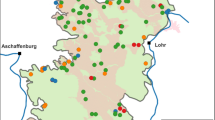
The size of each individual plot was 5 ha and if possible, a cluster of plots consisting each type was located in the same reserve (Fig. 1). The distances between nearest plots within a cluster varied between 100 m and 5 km, depending on the landscape structure. The plots were near-equally divided between eutrophic (22 stands; Aegopodium site type) and meso-eutrophic forests (18 stands, mostly Hepatica site type; sensu Lõhmus 1984). The most frequent dominant tree species in the stands was Norway spruce (21 stands), followed by birch Betula sp. (12), Scots pine (4), and European aspen Populus tremula L. (3).

Locations of the 40 study sites in Estonia. The zoom-in exemplifies a site cluster of four 5-ha sites.
2.2 Bird counts
In each of the 40 plots, breeding birds were surveyed in May–June 2020 by one of two observers; each observer studied a similar mixed set of stand types. We used a standard two-visit survey: one visit between 10 and 20 May and the second between 25 May and 10 June (Rosenvald et al. 2011). Each visit comprised two mapping censuses: a standard morning census between the sunrise and 10 a.m., supplemented by an evening census between 6 p.m. and the sunset (see Tomiałojć and Lontkowski 1989 for the rationale). In each of the four censuses, the observer mapped all birds and their behaviour within the 5-ha plot and around its borders by walking slowly at approximately 50–70 m distances (ca. 10–30 min spent per hectare, depending on visibility).
The field maps were later interpreted by the author E.P. for nesting territories (‘pairs’) using conventional criteria and a common procedure (e.g., Wechsler 2018). The interpretation first distinguished the most obvious territories based on nests or behaviour directly related to nesting (e.g., nest building), or singing or alarmed adults or pairs at the same spot in subsequent censuses. If two or more males of the same species were singing simultaneously on the same visit, these were counted as separate territories. The following observations were counted as 0.5 territories: (i) territorial birds moving across plot borders; (ii) species with large home range if the observations indicated a nesting territory but no nest was found (e.g., woodpeckers and raptors); (iii) multiple observations of a species that only forms unstable pairs (e.g., Scolopax rusticola L.).
The tests of these field and interpretation methods in Estonian forests have shown that, under bird-rich conditions (such as in this study), the method provides slightly (ca. 20%) lower abundance estimates than a standard multiple-visit mapping (Lõhmus and Rosenvald 2005; Lõhmus 2020). However, since the surveys were performed in the same year and by the same observers, the relative assemblage characteristics should be well comparable among sites.
2.3 Stand structure measurements
Each stand was described for the structure of the woody vegetation and dead wood, following the procedures described by Lõhmus and Kraut (2010). In a central 2-ha area in each plot, we pre-planned, in a geographical information system, four straight 50-m spaced-out transect lines. Along each 50-m transect, all living trees (diameter at breast height, DBH ≥ 10 cm), snags (DBH ≥ 1.0 m; including whole standing dead trees) and windthrow mounds (≥ 30 cm in height) were described on strip transects at both sides of the line; coarse downed dead wood items (diameter ≥ 10 cm) were registered at crossings with the transect lines; and fine woody items (> 0.3 cm in diameter) were censused in 1 × 1 m subplots. Tree species, diameter, and condition parameters (such as decay stage of dead wood) of each item were described.
For this paper, we used the plot-scale estimates (the four transects pooled) of the volumes of live trees and of coarse downed dead wood, and the densities of snags, windthrow mounds, and saplings per hectare. Forest stand age (mean age of overstorey trees), overstorey tree species per plot, and proportion of overstorey deciduous tree species (hereafter: deciduous component) were calculated using the data from the national forest survey database of the Estonian Environment Agency.
2.4 Linear modelling of the assemblage characteristics
We used general linear models (GLM) to analyze three bird assemblage characteristics as dependent variables: density (no. of nesting territories ha−1); resident bird density (a subset of the former, including partial short-distance migrants that are abundant year-long (see suppl. data) and species richness (total number of species having nesting territories in the plot). Prior to a multi-factor approach, we explored one-way ANOVA models for the stand type effects (Tukey HSD post hoc comparison for pairwise differences). All the models were run with STATISTICA 7.
To explain the relationship between these assemblage characteristics and forest structure (study question II), we used type III sums of squares GLM analyses. Due to fundamental structural differences, clear-cuts were not included in these models. The assumptions of normal distribution of the errors were checked, as well as correlations between the independent factors (forest structure). The continuous factor variables included were: (i) stand age (years); (ii) downed dead wood (m3 ha−1); (iii) tree stock (volume m3 of live trees ha−1); (iv) deciduous component (% deciduous trees in the overstorey); (v) number of tree species, and (vi) snag density (no. ha−1).
For each bird assemblage characteristic, we performed two analyses. First, as a basic model, we analyzed the set of independent variables, except “stand age” and “tree stock” that were closely (p < 0.001) correlated with each other (r = 0.61) and with the downed dead wood (stand age: r = 0.68; tree stock: r = 0.58). Then, we explored alternative models where the stand age or tree stock were added to the basic model. We report the alternative models only if the addition was either significant or exposed an effect of another, previously non-significant variable.
2.5 Species composition analyses
We explored differences in the bird species composition among stand types and their specific indicator species using PC-ORD 6.0 program (McKinney and Lockwood 2011). Separate analyses were carried out with and without clear-cuts, which had distinctly lower bird densities and species richness than stocked stands. The data matrix included the number of territories of each species by plot, excluding 13 species that were observed in only one plot.
We tested the compositional differences between stand types using multi-response permutation procedures (MRPP) with Sørensen (Bray-Curtis) distance measure. The differences were illustrated using non-metric multi-dimensional scaling (NMS). The number of axes was chosen with medium autopilot mode. Three sets of NMSs with real (and randomized) data with 250 runs each were performed manually. Linear correlations between the ordination scores and forest structural variables were calculated; the variables considered included in the GLM analyses (Section 5.4), as well as shrub cover (%), canopy cover (%), sapling density (no. ha−1), and windthrow density (no. ha−1).
To distinguish stand type-specific (characteristic) species, we calculated the indicator values, which combines the abundance and frequency of each species by pre-defined classification of sites (Dufrêne and Legendre 1997). Their p-levels in PC-ORD are estimated for the maximum observed value by comparing these with the mean and variance of 1000 Monte Carlo permutations. The values calculated are not originally corrected for multiple tests, and should be interpreted together with the MRPP tests to reduce the type I error probabilities.
3 Results
In the 40 5-ha plots, a total of 1386 nesting territories of 64 species were distinguished (6.9 territories ha−1). These numbers included 460.5 territories (33%) of 26 species of resident birds. Red crossbill (Loxia curvirostra L.) was also a possible breeder (< 5 pairs), but since its fledglings were already dispersed by the survey time, we omitted this species from the analysis.
The stand types differed significantly in terms of the overall breeding density (ANOVA: F4, 35 = 25.2; p < 0.001), species richness (F4, 35 = 26; p < 0.001), and resident bird density (F4, 35 = 39.6; p < 0.001) (Fig. 2). Clear-cuts had significantly smaller values of all these plot-scale characteristics (Tukey HSD post hoc tests p < 0.05); for example, the mean total densities between clear-cuts and late-successional forests differed on average by 7.3 nesting territories ha−1. In contrast, no significant contrasts were detected between planted spruce stands and successional naturally regenerated stands. Late-successional conifer stands were represented with the smallest sample and the confidence intervals of the estimates were wide; nevertheless, their differences from planted spruce stands also appeared relatively small (no significant post hoc contrasts).

Mean (± 95% CI) density (a) and species richness (b) of breeding birds in 5-ha plots by management type: CC, clear-cut; SP, planted spruce stands; NR, successional naturally regenerated stands; LSC, late-successional conifer; LSD, late-successional deciduous
The basic models explaining those differences in 30 forest plots (clear-cuts omitted) showed that all the three bird assemblage characteristics increased along with the downed dead wood volume; the species richness also responded to the “deciduous component” (Table 1). Adding to the basic model “stand age” and, in the case of resident bird density, also “tree stock”, further explained the variation, but (i) the downed dead wood variable lost its significance, and (ii) previously hidden positive influences were exposed in the breeding density model (“deciduous component”) and in the species richness model (“tree species/ha”) (Table 1). Notably, “stand age” was not significant for the resident bird density, even if “tree stock” was removed from the model.
The MRPP test confirmed that bird species compositions differed among the stand types (p < 0.01), except between the pairs of naturally regenerated successional vs. late-successional deciduous stands (p = 0.59), and planted spruce vs. late-successional conifer stands (p = 0.32). In terms of the ordination axes, the bird species composition varied along with the stand age, tree stock, downed dead wood, and the deciduous component (Fig. 3).

NMS ordination graphs of bird communities in all 40 sites (a), and for the subset of 30 sites with tree cover (b). Each symbol refers to one 5-ha site. Two most informative axes (% variance explained indicated) of three-dimensional solutions are shown. Environmental gradients are depicted as arrows proportional in length to their co-variation with the axes (those with r2 > 0.1 shown). DDW, volume of downed dead wood
Out of 52 bird species included in the indicator species analysis, 23 species showed significant stand-type associations. Fifteen species were primarily associated with late-successional deciduous stands, three species with late-successional conifer stands, and five species with clear-cuts. Of the 18 species associated with late-successional stands, ten species were resident birds, while no resident species was associated with clearcuts (Table 2). No significant indicator values were detected for either planted spruce or naturally regenerated stands. However, both types of successional stands pooled, common chiffchaff (Phylloscopus collybita Vieill.), and icterine warbler (Hippolais icterina Vieill.) reached significant indicator values, and all 2.5 pairs of the hazel grouse (Tetrastes bonasia L.) were found there.
4 Discussion
4.1 The structure of forest bird assemblages
A major result was that the forest bird densities, species richness and resident bird densities were roughly ordered at the scale of naturalness (Fig. 2) and stand age (Fig. 2; Tables 1 and 2). The average values indicated that planted spruce stands had the most depauperate bird assemblages among stocked stands, and the values grew with the stand age (study question I). Surprisingly, the MRPP test did not support species composition differences between the planted and late-successional conifer stands, which can be partly due to our small sample of the latter (i.e., low test power). Nevertheless, all conifer stands obviously provide some distinct conditions, such as conifer seeds, year-round shade and, in older stands, bark beetle infestation sites (Hughes and Hudson 1997; Williams et al. 2017; Vélová et al. 2021). Considering also the influences of stand structural factors, we conclude that bird assemblages in ageing, structurally rich spruce plantations can develop toward native late-successional conifer stands (Steverding and Leuschner 2002; Yamaura et al. 2019).
Thus, spruce planting after clear-cutting transforms bird assemblages fundamentally for at least one tree generation. This is distinct from a site-type specific development in naturally regenerated stands where the general bird composition resembled (impoverished) late-successional deciduous stands. There can be species-level exceptions, however. Notably, in line with Swenson and Angelstam (1993), we found that the hazel grouse (Tetrastes bonasia) inhabited successional stands of both regeneration types.
For interpreting the absolute densities recorded, one must acknowledge possible census bias. Our estimates seem consistent with the previous research in Estonia, although they can still be best comparable with the studies using the same methodology. Of these, Rosenvald et al. (2011) found slightly higher bird densities in boreo-nemoral old-growth stands (12 pairs ha−1) than we had in our late-successional stands (10.2 pairs ha−1). This may indicate that the densities still increase between 100 and 200 years of stand age (e.g., Zawadzka et al. 2018). The fact that siskin (Spinus spinus L.) and goldcrest (Regulus regulus L.) numbers were remarkably higher in our study may refer to a favourable year for those species. In various clear-cuts of the same site types in Estonia, Rosenvald and Lõhmus (2007) documented 1.6–3.7 pairs ha−1, while our estimate was 3.2 pairs ha−1. Moreover, using the multiple-visit mapping method, Lõhmus (2020) found similar densities in mid-aged spruce plantations (5.5 pairs vs our 6 pairs ha−1).
Also, the effects of forest structure were broadly consistent on different bird assemblage characteristics (question II; Fig. 3). An enrichment was provided by old-forest elements (downed dead wood, large and old trees; Nilsson 1979b; Mag and Ódor 2015) combined with multiple tree species, notably the deciduous component. Those structures are likely to improve various conditions, notably food resources, shelter options and nest sites for birds (Nilsson 1997; Rosenvald et al. 2011; Vélová et al. 2021). Interestingly, the deciduous component had an independent effect on bird species richness while accounting for tree species richness. This may indicate a role of certain deciduous species, e.g., for cavity nesters or arboreal foragers. Similarly, living tree stock had a positive influence on resident bird density, even when accounting for the stand age that has been previously stressed as important for resident bird communities (Imbeau et al. 2001; Zawadzka, et al. 2018). In contrast, stand age or living tree stock fully explained the effect of downed dead wood, supporting the idea that the downed dead wood rather reflects a complex of successional changes (Lõhmus et al. 2021). Contrary to numerous studies (e.g., Gutzat and Dormann 2018; Lewandowski et al. 2021), snag abundance was not related with bird assemblage characteristics.
As expected, late-successional stands hosted the largest numbers of characteristic species (question III; Table 2), but new information was that these were largely resident species. Characteristic migrants nevertheless included some previously identified focal species for conservation, such as the red-breasted flycatcher (Ficedula parva Bechst.) (Lõhmus et al. 2020) and greenish warbler (Phylloscopus trochiloides Sund.) (Väli and Vaan 2020; Lõhmus et al. 2021). However, several species identified as old-growth associated by Rosenvald et al. (2011) were rarely encountered in our study, e.g., three-toed woodpecker (Picoides tridactylus L.) and long-tailed tit (Aegithalos caudatus L.). A reason may be that most of our late-successional sites were small fragments retained in production forest landscapes. Thus, a high density and species richness in our late-successional stands might be due to the surrounding managed landscape (Wesolowski 2007) and edge effects (Gimenes and Dos Anjos 2003; Batáry et al. 2014).
4.2 Implications for sustainable forest management
Our main implications are related to the overall bird abundance along forest succession, and to the specific factors shaping habitat quality in spruce plantations. First, we found that by clear-cutting a late-successional stand, on average 6–8 pairs are lost per hectare in the productive sites studied. The post-clearcut bird densities recovers rapidly up to ca 30-year-old stands when the densities are, based on our study, roughly 65% of late-successional levels and the compositions remain impoverished. After that, the recovery rate is only about one pair ha−1 per 20 years up to at least 100 years of stand age, when the forests still lack most old-growth specialist species (Rosenvald et al. 2011). Such a pattern appears despite a structurally rapid forest recovery on fertile soils (Nilsson 1997; Lõhmus et al. 2020; 2021). We thus concur with other studies (Schmiegelow and Mönkkönen 2002; Fraixedas et al. 2015; Zawadzka et al. 2018) on the importance of old-growth reserves, but we separately highlight the issue of overall losses of bird abundance on production forest landscapes.
Our silvicultural implications are most relevant for even-aged systems. It is well known and confirmed in our study (see also Hansson 1994; Lõhmus 2004), that clear-cuts have distinct bird assemblages of mostly common, but also some declining open habitat or shrubland species (such as Carpodacus erythrinus Pall.). A fundamental difference emerges in the thicket phase, depending on whether the stand is naturally regenerated or planted with spruce. Integrating planted stands into landscapes and retaining a share of natural regeneration would constitute landscape-scale precautionary approaches against biotic homogenization.
Also, the habitat value of spruce plantations can still be facilitated through later management. The ultimate range of techniques depends on the economic options available for the managers, but our study helps prioritize the actions by their ecological effectiveness. A priority for birds would be to establish deciduous gaps in homogenous conifer plantations (Lindbladh et al. 2017; Horák et al. 2019; Vélová et al. 2021), with a long-term aim to increase tree species richness and abundance of deciduous trees. Another major effect (notably on resident birds) is provided by the presence of large trees, which can be achieved through long-term planning of the retention forestry techniques (Rosenvald and Lõhmus 2007; Mag and Ódor 2015; Rosenvald et al. 2019; Gustafsson et al. 2020). Thus, managing even-aged successional forests for birds starts from the clear-cutting stage. The thinning operations in older stands can be crucial for the development of dead wood pools and retaining cavity trees, but their potential has already been shaped by earlier silvicultural decisions.
5 Conclusions
1. Bird assemblages in planted spruce forests were compositionally distinct compared with naturally regenerated mixed stands and impoverished compared with late-successional forests.
2. Due to relatively rapid stand development on productive soils, avian assemblages can host some old-forest species within even-aged systems, although full old-growth assemblages probably further develop in stands well over 100 years old and some species cannot inhabit the production landscape mosaics.
3. Both in planted and naturally regenerated stands, the main silvicultural targets of bird habitat quality are similar (tree species mixtures; large-tree retention; dead wood).
Availability of data and materials
The datasets generated and/or analyzed during the current study are available in the Zenodo repository. https://doi.org/10.5281/zenodo.6323328
References
Batáry P, Fronczek S, Normann C, Scherber C, Tscharntke T (2014) How do edge effect and tree species diversity change bird diversity and avian nest survival in Germany’s largest deciduous forest? For Ecol Manag 319:44–50. https://doi.org/10.1016/j.foreco.2014.02.004
Ahti T, Hämet-Ahti L, Jalas J (1968) Vegetation zones and their sections in northwestern Europe. Ann Bot Fenn 5:169–211
Bergner A, Avci M, Eryigit H et al (2015) Influences of forest type and habitat structure on bird assemblage of oak (Quercus spp.) and pine (Pinus ssp.) stands in southwestern Turkey. For Ecol Manag 336:137–147. https://doi.org/10.1016/j.foreco.2014.10.025
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:354–366. https://doi.org/10.1890/0012-9615(1997)067[0345:SAAIST]2.0.CO;2
Felton A, Andersson E, Ventorp D, Lindbladh M (2011) A comparison of avian diversity in spruce monocultures and spruce-birch polycultures in southern Sweden. Silva Fenn 45(5):1143–1150. https://doi.org/10.14214/sf.92
Felton A, Löfroth T, Angelstam P, Gustafsson L, Hjältén J, Felton AM, Simonsson P, Dahlberg A, Lindbladh M, Svensson J, Nilsson U, Lodin I, Hedwall PO, Sténs A, Lämås T, Brunet J, Kalén C, Kriström B, Gemmel P, Ranius T (2020) Keeping pace with forestry: multi-scale conservation in a changing production forest matrix. Ambio 49(5):1050–1064. https://doi.org/10.1007/s13280-019-01248-0
Felton A, Hedwall PO, Trubins R, Lagerstedt J, Felton A, Lindbladh M (2021) From mixtures to monocultures: Bird assemblage responses along a production forest conifer-broadleaf gradient. For Ecol Manag 494:119299. https://doi.org/10.1016/j.foreco.2021.119299
Fraixedas S, Lindén A, Lehikoinen A, Henderson I (2015) Population trends of common breeding forest birds in southern Finland are consistent with trends in forest management and climate change. Ornis Fenn 92:187–203
Fuller RJ (2003) Bird life of woodland and forest. Cambridge University Press, Cambridge
Georgiev KB, Chao A, Castro J, Chen YH, Choi CY, Fontaine JB, Hutto RL, Lee EJ, Müller J, Rost J, Żmihorski M, Thorn S (2020) Salvage logging changes the taxonomic, phylogenetic and functional successional trajectories of forest bird communities. J Appl Ecol 57(6):1103–1112. https://doi.org/10.1111/1365-2664.13599
Gimenes MR, dos Anjos L (2003) Effects of forest fragmentation on bird communities. Acta Sci Biol Sci 25:392–402
Gjerde I, Saetersdal M (1997) Effects on avian diversity of introducing spruce Picea spp. plantations in the native pine Pinus sylvestris forests of western Norway. Biol Conserv 79(2-3):241–250. https://doi.org/10.1016/S0006-3207(96)00093-6
Gregory RD, van Strien A (2010) Wild bird indicators: Using composite population trends of birds as measures of environmental health. Ornithol Sci 9(1):3–22. https://doi.org/10.2326/osj.9.3
Gustafsson L, Bauhus J, Asbeck T, Augustynczik ALD, Basile M, Frey J, Gutzat F, Hanewinkel M, Helbach J, Jonker M, Knuff A, Messier C, Penner J, Pyttel P, Reif A, Storch F, Winiger N, Winkel G, Yousefpour R, Storch I (2020) Retention as an integrated biodiversity conservation approach for continuous-cover forestry in Europe. Ambio 49(1):85–97. https://doi.org/10.1007/s13280-019-01190-1
Gutzat F, Dormann CF (2018) Decaying trees improve nesting opportunities for cavity-nesting birds in temperate and boreal forests: A meta-analysis and implications for retention forestry. Ecol Evol 17(16):1–11. https://doi.org/10.1002/ece3.4245
Hansson L (1994) Vertebrate distributions relative to clear-cut edges in a boreal forest landscape. Landsc Ecol 9:105–115. https://doi.org/10.1007/BF00124377
Horák J, Brestovanská T, Mladenović S, Kout J, Bogusch P, Halda JP, Zasadil P (2019) Green desert? Biodiversity patterns in forest plantations. For Ecol Manage 433:343–348. https://doi.org/10.1016/j.foreco.2018.11.019
Hughes JW, Hudson FK (1997) Songbird nest placement in Vermont Christmas tree plantations. Can Field-Nat 111:580–585
Imbeau L, Mönkkönen M, Desrochers A (2001) Long-term effects of forestry on birds of the Eastern Canadian boreal forests: a comparison with Fennoscandia. Conserv Biol 15(4):1151–1162. https://doi.org/10.1046/j.1523-1739.2001.0150041151.x
Kebrle D, Zasadil P, Hošek J, Barták V, Šťastný K (2021) Large trees as a key factor for bird diversity in spruce-dominated production forests: Implications for conservation management. For Ecol Manag 496:119460. https://doi.org/10.1016/j.foreco.2021.119460
Klein J, Thor G, Low M, Sjögren J, Lindberg E, Eggers S (2020) What is good for birds is not always good for lichens: Interactions between forest structure and species richness in managed boreal forests. For Ecol Manag 473:118327. https://doi.org/10.1016/j.foreco.2020.118327
LaManna JA, Martin TE (2016) Logging impacts on avian species richness and composition differ across latitudes and foraging and breeding habitat preferences. Biol Rev 92: 1657–1674, 3, DOI: https://doi.org/10.1111/brv.12300
Lewandowski P, Przepiora F, Ciach M (2021) Single dead trees matter: Small-scale canopy gaps increase the species richness, diversity and abundance of birds breeding in a temperate deciduous forest. For Ecol Manag 481:118693. https://doi.org/10.1016/j.foreco.2020.118693
Liira J, Sepp T (2009) Indicators of structural and habitat natural quality in boreo-nemoral forests along the management gradient. Ann Bot Fenn 46(4):308–325. https://doi.org/10.5735/085.046.0407
Lindbladh M, Lindström Å, Hedwall PO, Felton A (2017) Avian diversity in Norway spruce production forests – How variation in structure and composition reveals pathways for improving habitat quality. For Ecol Manag 397:48–56. https://doi.org/10.1016/j.foreco.2017.04.029
Lindbladh M, Petersson L, Hedwall PO, Trubins R, Holmström E, Felton A (2019) Consequences for bird diversity from a decrease in a foundation species—replacing Scots pine stands with Norway spruce in southern Sweden. Reg Environ Change 19(5):1429–1440. https://doi.org/10.1007/s10113-019-01480-0
Lõhmus E (1984) Estonian forest site types. Eesti NSV Agrotööstuskoondise Info- ja juurutusvalitsus, Tallinn (in Estonian)
Lõhmus A (2004) Breeding bird communities in two Estonian forest landscapes: are managed areas lost for biodiversity conservation. Proc Estonian Acad Sci Biol Ecol 53(1):52–67. https://doi.org/10.3176/biol.ecol.2004.1.05
Lõhmus A, Kraut A (2010) Stand structure of hemiboreal old-growth forests: Characteristic features, variation among site types, and a comparison with FSC-certified mature stands in Estonia. For Ecol Manag 260(1):155–165. https://doi.org/10.1016/j.foreco.2010.04.018
Lõhmus A, Rosenvald R (2005) Breeding bird fauna of the Järvselja Primeval Forest Reserve: long-term changes and an analysis of inventory methods. Hirundo 18:18–30
Lõhmus A (2020) Population densities of breeding birds in forests surrounding the drained Kripsi mire (East Estonia). Hirundo 33:30–52
Lõhmus A, Kont R, Runnel K, Vaikre M, Remm L (2020) Habitat models of focal species can link ecology and decision-making in sustainable forest management. Forests 11(7):1–27. https://doi.org/10.3390/f11070721
Lõhmus A, Nellis R, Pullerits M, Leivits M (2016) The potential for long-term sustainability in seminatural forestry: A broad perspective based on woodpecker populations. Environ Manage 57(3):558–571. https://doi.org/10.1007/s00267-015-0638-2
Lõhmus A, Runnel K, Palo A, Leis M, Nellis R, Rannap R, Remm L, Rosenvald R, Lõhmus P (2021) Value of a broken umbrella: biodiversity in single-species reserves established for, but abandoned by, the black stork (Ciconia nigra). Biodivers Conserv 30(12):3647–3664. https://doi.org/10.1007/s10531-021-02268-7
MacKay A, Allard M, Villard MA (2014) Capacity of older plantations to host bird assemblages of naturally-regenerated conifer forests: A test at stand and landscape levels. Biol Conserv 170:110–119. https://doi.org/10.1016/j.biocon.2013.12.023
Mag Z, Ódor P (2015) The effect of stand-level habitat characteristics on breeding bird assemblages in Hungarian temperate mixed forests. Community Ecol 16(2):156–166. https://doi.org/10.1556/168.2015.16.2.3
McKinney M, Lockwood J (2011) PC-ORD. Multivariate analysis of ecological data. MjM Software, Oregon
Nellis R, Volke V (2019) Changes in abundances of forest birds during the period of 1983 to 2018. Hirundo 32:63–80
Nilsson SG (1979a) Density and species richness of some forest bird communities in South Sweden. Oikos 33(3):392–401. https://doi.org/10.2307/3544327
Nilsson SG (1979b) Effect of forest management on the breeding bird community in southern Sweden. Biol Conserv 16(2):135–143. https://doi.org/10.1016/0006-3207(79)90061-2
Nilsson SG (1997) Forests in the temperate-boreal transition: natural and man-made features. Ecol Bull 46:61–71
Oettel J, Lapin K (2021) Linking forest management and biodiversity indicators to strengthen sustainable forest management in Europe. Ecol Indic 122:107275. https://doi.org/10.1016/j.ecolind.2020.107275
Pass E, Kont R, Lõhmus A (2022) A dataset of bird assemblages in different successional pathways– a comparative study in hemiboreal mixed forests. Zenodo. [dataset]. V1. https://doi.org/10.5281/zenodo.6323328.
Põldveer E, Korjus H, Kiviste A, Kangur A, Paluots T, Laarmann D (2020) Assessment of spatial stand structure of hemiboreal conifer dominated forests according to different levels of naturalness. Ecol Indic 110:105944. https://doi.org/10.1016/j.ecolind.2019.105944
Raudsaar M, Valgepea M (2020) Yearbook Forest 2019. Estonian Environmental Agency, Tallinn
Roberge J-M, Virkkala R, Mönkkönen M (2018) Boreal forest bird assemblages and their conservation. In: Fuller RJ, Roberge J-M (eds) Mikusiński G. Cambridge University Press, Ecology and conservation of forest birds, pp 183–230
Rosenvald R, Lõhmus A (2007) Breeding birds in hemiboreal clear-cuts: Tree-retention effects in relation to site type. Forestry 80(5):503–516. https://doi.org/10.1093/forestry/cpm027
Rosenvald R, Lõhmus A, Kraut A, Remm L (2011) Bird communities in hemi-boreal old-growth forests: The roles of food supply, stand structure, and site type. For Ecol Manag 262(8):1541–1550. https://doi.org/10.1016/j.foreco.2011.07.002
Rosenvald R, Lõhmus P, Rannap R, Remm L, Rosenvald K, Runnel K, Lõhmus A (2019) Assessing long-term effectiveness of green-tree retention. For Ecol Manag 448:543. https://doi.org/10.1016/j.foreco.2019.06.034
Schmiegelow FKA, Mönkkönen M (2002) Habitat loss and fragmentation in dynamic landscape: avian perspectives from the boreal forest. Ecol Appl 12(2):375–389. https://doi.org/10.2307/3060949
Spiecker H (2003) Silvicultural management in maintaining biodiversity and resistance of forests in Europe - temperate zone. J Environ Manag 67(1):55–65. https://doi.org/10.1016/S0301-4797(02)00188-3
Steverding M, Leuschner C (2002) Effects of Norway spruce monocultures on the structure of bird communities in a submontane-montane forested landscape of Central Germany. Forstwiss Central 121(2):83–96. https://doi.org/10.1046/j.1439-0337.2002.00083.x
Swanson ME, Franklin JF, Beschta RL, Crisafulli CM, DellaSala DA, Hutto RL, Lindenmayer DB, Swanson FJ (2011) The forgotten stage of forest succession: early-successional ecosystems on forest sites. Front Ecol Environ 9(2):117–125. https://doi.org/10.1890/090157
Sweeney OFMD, Wilson MW, Irwin S, Kelly TC, O’Halloran J (2010) Are bird density, species richness and community structure similar between native woodlands and non-native plantations in an area with a generalist bird fauna? Biodivers Conserv 19(8):2329–2342. https://doi.org/10.1007/s10531-010-9844-7
Swenson JE, Angelstam P (1993) Habitat separation by sympatric forest grouse in Fennoscandia in relation to boreal forest succession. Can J Zool 71(7):1303–1310. https://doi.org/10.1139/z93-180
Thorn S, Chao A, Bernhardt-Römermann M, Chen YH, Georgiev KB, Heibl C, Müller J, Schäfer H, Bässler C (2020) Rare species, functional groups, and evolutionary lineages drive successional trajectories in disturbed forests. Ecology 101(3):e02949. https://doi.org/10.1002/ecy.2949
Tomiałojć L, Lontkowski J (1989) A technique for censusing territorial song thrushes Turdus philomelos. Ann Zool Fennici 26:235–244
Väli Ü, Vaan PK (2020) Greenish warbler (Phylloscopus trochiloides viridanus): An overlooked indicator of old-growth forest? Ornis Fennica 97:165–176
Vélová L, Véle A, Horák J (2021) Land use diversity and prey availability structure the bird communities in Norway spruce plantation forests. For Ecol Manag 480:118657. https://doi.org/10.1016/j.foreco.2020.118657
Voříšek P, Schwarz M, Raši R (2020) Indicator 4.10 Common forest bird species. In: Forest Europe 2020: State of Europe’s Forests. Forest Europe, Bratislava
Wechsler S (2018) Automating the analysis of territory mapping data in bird monitoring. University of Salzburg, Austria, Master’s thesis
Wesolowski T (2007) Primeval conditions – what can we learn from them? Ibis Suppl 149:64–77. https://doi.org/10.1111/j.1474-919X.2007.00721.x
Williams DT, Straw N, Fielding N, Jukes M, Price J (2017) The influence of forest management systems on the abundance and diversity of bark beetles (Coleoptera: Curculionidae: Scolytinae) in commercial plantations of Sitka spruce. For Ecol Manag 398:196–207. https://doi.org/10.1016/j.foreco.2017.05.014
Yamaura Y, Lindenmayer D, Yamada Y, Gong H, Matsuura T, Mitsuda Y, Masaki T (2019) A spatially-explicit empirical model for assessing conservation values of conifer plantations. For Ecol Manag 444:393–404. https://doi.org/10.1016/j.foreco.2019.04.038
Zawadzka D, Drozdowski S, Zawadzki G, Zawadzki J, Mikitiuk A (2018) Importance of old forest stands for diversity of birds in managed pine forests – A case study from Augustów Forest (NE Poland). Pol J Ecol 66(2):162–181. https://doi.org/10.3161/15052249PJE2018.66.2.007
Acknowledgements
We thank Liisi Peets for assistance in the bird mapping, Annabel Runnel for fieldwork assistance, and Maarja Vaikre for statistical help and fieldwork assistance.
Code availability
Not applicable
Funding
This research was funded by the Estonian Research Council (grant SLTOM19456 and PRG1121) and the Estonian Center for Environmental Investments (grant 16288).
Author information
Authors and Affiliations
Contributions
Conceptualization: Eliisa Pass, Asko Lõhmus. Methodology: Asko Lõhmus. Formal analysis and investigations: Eliisa Pass, Raido Kont. Writing—original draft preparation: Eliisa Pass. Writing—review and editing: Asko Lõhmus. Funding acquisition: Asko Lõhmus. Resources: Asko Lõhmus. Supervision: Asko Lõhmus. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The authors declare that they obtained the approval of Estonian Environmental Board for conducting the study in Haavakannu Natural Reserve. Not applicable.
Consent for publication
All authors approved publication of the manuscript
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Aurélien Sallé
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Pass, E., Kont, R. & Lõhmus, A. Spruce (Picea abies L.) planting leads post-clearcut bird assemblages to a novel successional pathway—a comparative study in hemiboreal mixed forests. Annals of Forest Science 79, 20 (2022). https://doi.org/10.1186/s13595-022-01138-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13595-022-01138-8