
Abstract
Caturoidea is a clade of Mesozoic predatory ray-finned fishes which lived mainly in the Jurassic. The clade has a few records in the earliest Cretaceous and only two in the Triassic. Among the latter, specimen MPCA 632 Caturus sp. doubtfully from continental Early Triassic of Argentina, i.e., outside Europe, was particularly problematic in the light of the known fossil record of the group, which suggested their origin in the Western Tethys. The micropaleontological and geochemical analysis of bulk-rock samples of MPCA 632 allowed us to correct the provenance of the specimen which corresponds to Tithonian marine outcrops of the Vaca Muerta Formation, Neuquén, Argentina. Specimen MPCA 632 is excluded from Caturus and reclassified as Caturoidea sp. MPCA 632 might be a specimen of Catutoichthys olsacheri, the only caturoid known from the Vaca Muerta Formation (Los Catutos Member), but the fossils are not comparable and, thus, this hypothesis needs further study. Additionally, the first-hand study of the type material of the only other alleged Triassic caturoid, Furo insignis, in the Norian of Seefeld, Austria, led to the exclusion of this taxon from the Caturoidea. Consequently, the clade Caturoidea is restricted to the Jurassic–Lowest Cretaceous. After a modest evolution during the Early Jurassic, the group had its initial radiation and westward dispersion across the Hispanic Corridor during the Middle Jurassic and reached its maximal diversity during the Kimmeridgian and Tithonian.
Similar content being viewed by others

Introduction
The superfamily Caturoidea (Grande & Bemis, 1998) includes small to medium size Mesozoic predatory ray-finned fishes. They constitute the sister-group of Amiioidea within the order Amiiformes (Neopterygii: Holostei), which is represented today by only two species of the genus Amia, which are restricted to freshwater systems of North America (Brownstein et al., 2022; Wright et al., 2022). The larger clade Halecomorphi embraces the Amiiformes and other extinct groups (ophiopsids, ionoscopids, panxianichthyiforms, parasemionotiforms, etc.) whose classification and phylogenetic relationships are still debatable (Ebert, 2018; Sun et al., 2017; Xu, 2019). Although they are so poorly represented today, halecomorphs were very successful, diverse and abundant during the Mesozoic, and the reasons for their severe decline during the Cenozoic remain unclear.
The known fossil record of the Halecomorphi starts in the Lower Triassic and the oldest record of Amiiformes is reported from the Upper Triassic (Norian) (López-Arbarello & Ebert, 2023; López-Arbarello & Sferco, 2018). A recent taxonomic revision of the Caturoidea (López-Arbarello & Ebert, 2023) shows that the known fossil record of the group is almost restricted to the Jurassic, and revealed an important diversification of this group during the Late Jurassic that is not only indicated by the increase in the number of taxa, but also by the dispersal of the group outside Europe, which had already started in the Middle Jurassic. Despite reaching their maximal diversity and dispersion in the Tithonian, the youngest records of the group in the earliest Cretaceous are limited to western Europe (Berriasian of Swanage, Dorset, United Kingdom; Berriasian–lower Valanginian of El Montsec, Lérida, Spain), suggesting a very rapid decline of the group at that time.
Before the Jurassic, the only records of Caturoidea were Caturus insignis (Kner, 1866), in the Norian of Seefeld, Austria, and Caturus sp. from the freshwater Triassic Los Menucos Group in Argentina (Bogan et al., 2013). Considering the known fossil record of this group, López-Arbarello and Ebert (2023) threw doubts on the provenance of the latter fossil because the presence of a caturoid in freshwater deposits at such an early stage and outside Europe appeared extremely odd. After careful revision of the information about the finding and the micropaleontological analysis of the bearing rock, the provenance of the Argentinean fossil is conclusively corrected in this contribution. Furthermore, the referral of Caturus insignis (Kner, 1866) to the genus Caturus is also revised and corrected.
Geological setting
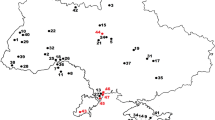
The fossil caturid focus of this study was published with provenance in the Triassic Vera Formation of the Los Menucos Group (Bogan et al., 2013). It will be shown here that the fossil was collected in Jurassic outcrops of the Mendoza Group at Las Lajas, Neuquén (Fig. 1). Therefore, brief descriptions of the geological settings of the Los Menucos and Mendoza groups are necessary to understand the context in which the fossil sample has been analyzed.

Geographical setting: a, general location; b, simplified map showing outcrops of the Mendoza and Los Menucos groups in the provinces of Neuquén and Río Negro. Outcrops of the Mendoza Group are redrawn from the regional scheme in the geological map HG 3969-I Zapala (Leanza et al., 2001). Outcrops of the Los Menucos Group are redrawn from the regional scheme (esquema regional) in the geological map HG 4169-II Los Menucos (Cuchi et al., 2001)
Los Menucos Group
The continental volcanic and sedimentary rocks around the locality Los Menucos, Río Negro province, Argentina, correspond to the Los Menucos Group (Fig. 1; Labudía & Bjerg, 2001). The Los Menucos Group is characterized by recurrent volcanic activity, and it is divided in three units: Puesto Tscherig, Puesto Vera, and Sierra Colorada formations (Falco et al., 2020). Contrary to the previously accepted Middle–Late Triassic age, the stratigraphic and geochronological limits for the Los Menucos Group have been recently established between the Changhsingian (upper Lopingian, Upper Permian; U–Pb zircon age of 253 ± 2 Ma) and the Olenekian (upper Lower Triassic; U–Pb zircon age of 248 ± 2 Ma) (Falco et al., 2020; Luppo et al., 2018). These units are constituted by volcanic, volcaniclastic and pyroclastic and siliciclastic rocks. Based on their lithofacies, megaflora and microfossil content, the whole Los Menucos Group has been assigned to continental environments (Falco et al., 2020).
The Puesto Tscherig and Puesto Vera formations contain a rich megaflora, including the classic coristosperm taxa of the "Dicroidium Flora" together with phyllicines, pleuromeiales, pteridosperms, cycadales, bennettitiales, and ginkgoales (Artabe, 1985, 1986), and tetrapod ichnites of the genus Dicynodontipus (Citton et al., 2018; Díaz-Martínez & De Valais, 2014).
Mendoza Group
The Mendoza Group includes marine lithostratigraphic units that began with a Tithonian transgression in the Neuquén Basin which was a retro-arc depocenter connected to the palaeo-Pacific Ocean and located between the South American subsiding foreland to the east and a volcanic arc to the west, and evolved uninterruptedly through the Jurassic-Cretaceous boundary. The group expands over the province of Neuquén and the south of the Mendoza province (Fig. 1), and its outcrops are classified in several formations distributed as follows: Carrín Curá Formation (marginal marine or fluvial coastal sediments), Picún Leufú Formation (shallow marine and coastal sediments), Mulichinco Formation (shallow marine and coastal sediments), and Agrio Formation (mid-shelf sediments) in the southern part of the Neuquén province; Vaca Muerta Formation (mid-shelf sediments; Otharán et al., 2020), and Mulichinco and Agrio formations (shallow marine and coastal sediments or mid-shelf sediments) in the central and northern part of the Neuquén province.
The Vaca Muerta Formation encompasses an Upper Jurassic–Lower Cretaceous sequence of alternating marine organic shales and limestones (Rodriguez Blanco et al., 2020, and references therein) that developed from the early Tithonian to the early Valanginian (Leanza et al., 2011; Legarreta & Uliana, 1996; Legarreta & Villar, 2015; Scasso et al., 2005). Within the Vaca Muerta Formation, the carbonate Los Catutos Member, defined by Leanza and Zeiss (1990) is particularly well-known for its rich paleontological content, including fishes and tetrapods (Cione et al., 1987; Codorniú & Gasparini, 2013; Gasparini et al., 2015).
Materials and methods
Repositories and institutional abbreviations
Specimen MPCA 632 is deposited in the Museo Carlos Ameghino (MPCA), Paraje Pichi Ruca, Río Negro, Argentina. Sample material and smear slides are stored in the Paleontological Collections of the Department of Geological Sciences, University of Buenos Aires, under the acronym BACF-NP 4220–4224.
Other institutions cited in the text are: AMNH, American Museum of Natural History, New York, U.S.A.; NMNH, National Museum of Natural History (Smithsonian Institution), Washington, DC, U.S.A.; TLM, Sammlungs- und Forschungszentrum der Tiroler Landesmuseen, Hall in Tirol, Austria.
Micropaleontological analysis
The rock MPCA 632 was examined for calcareous nannofossils and resulted productive. The sample was prepared on smear slides using standard techniques (Edwards, 1963). Additional slides were prepared through gravity settling (Bown & Young, 1998) and fixed with UV curing Norland Optical Adhesive. Special care was taken to prepare the samples as uniformly as possible to have comparable slides.
Nannofossil identification and photography were performed with a standard LEICA DMLP petrographic microscope under 1000 × magnification and polarized light.
Carbon- and oxygen-isotopic analyses
Oxygen and carbon stable isotopes of bulk-rock samples of MPCA 632 have been analyzed at the GeoZentrum Nordbayern, Friedrich-Alexander Universität Erlangen-Nürnberg. Sampling was restricted to lime mudstones-wackestones. The samples were prepared and analyzed following standard preparation techniques. Carbonate powders were reacted with 100% phosphoric acid at 70°C using a Gasbench II connected to a ThermoFisher Delta V Plus mass spectrometer at Erlangen University.
All values are reported in per mil relative to V-PDB. Reproducibility and accuracy were monitored by replicate analysis of laboratory standards calibrated by assigning a δ13C of + 1.95‰ internationally accepted International Atomic Energy Association carbonate standard NBS-19 (Friedman, 1982). NBS19 and − 47.3‰ to IAEA-CO9 and a δ18O of − 2.20‰ to NBS19 and − 23.2‰ to NBS18. Reproducibility for δ13C and δ18O was ± 0.02 and ± 0.04 (1 std. dev.), respectively. Standard deviations in the MPCA 632 samples are 0.02‰ for carbon and 0.01‰ for oxygen. Standard NBS 19 (n = 2) was additionally analyzed as quality control sample with δ13C = 1.98‰ and δ18O = − 2.19‰. The only carbonate mineral recognized in the analyzed simple set was calcite. The δ13C and δ18O data are reported in the conventional delta notation with respect to V-PDB.
Systematic paleontology
Actinopterygii Cope, 1887
Neopterygii Regan, 1923
Holostei Müller, 1845, sensu Huxley (1861)
Halecomorphi Cope, 1871, sensu Grande (2010)
Amiiformes Hay, 1929
Caturoidea Grande and Bemis, 1998
Caturidae Owen, 1860, sensu Grande and Bemis (1998)
Caturidae indeterminate genus and species
Material: MPCA 632 (Fig. 2a).

Specimen MPCA 632 from the Jurassic of Neuquén, Argentina: a, fossil of Caturidae sp., including disarticulated skull bones described in Bogan et al. (2013); b, corresponding inventory card of the Museo Carlos Ameghino (MPCA)
Occurrence: Jurassic outcrops at Las Lajas, Neuquén (Fig. 1b).
Remarks: MPCA 632 was described in detail in Bogan et al. (2013) and there is no need to complete or modify this description. However, the referral of the fossil to the genus Caturus is not supported here.
The limits of Caturidae are poorly defined and the family is currently under revision (López-Arbarello & Ebert, 2023). Lambers (1992, 1994) revised the caturids from the Upper Jurassic of Southern Germany, but did not give a definition or diagnosis of the family. Grande and Bemis (1998) found only one character supporting the clade Caturidae, formed by the genera Caturus and Amblysemius in their phylogenetic analysis of amiiforms. This feature, the presence of paired, block-like ural neural arch ossifications, cannot be evaluated in MPCA 632 because only skull bones are preserved. However, the extremely slender maxilla, the marginal teeth with sharply carinate and labiolingually compressed acrodin caps and the general shape of the preopercle most closely resemble Caturus and Amblysemius amongst ray-finned fishes.
Comparing with these two caturid genera, Bogan et al. (2013) discussed morphological differences between MPCA 632 and the species of Amblysemius, and concluded that the Argentinean fish is a species of Caturus.
Although the morphological discussion of Bogan et al. (2013) is correct, the fish preserved in MPCA 632 clearly differs from the species of Caturus and most probably represents a different caturid genus. The main difference is found in the shape of the maxilla. Although very slender, the maxilla of Caturus forms a distal, laterally compressed posteroventral expansion, completely garnished with teeth, and the straight posterior border forms a small but well-defined postmaxillary process (Lambers, 1992, chapter 5: figs. 1, 4). The very slender and rod-like maxilla in MPCA 632 lack such a distal expansion. Instead, it has an oblong posterior end without a well-defined posterior border or postmaxillary process (Fig. 2a; Bogan et al., 2013: figs. 3, 5A).
Results
Microfossils
The micropaleontological analysis of BACF-NP 4220–4224 (from MPCA 632) through positive results, although the taxonomic richness of the recovered nannoflora is very low. The moderately to poorly preserved nannoflora is constituted by rare specimens of Polycostella beckmanni and abundant watznauerids, Watznaueria fossacincta, Watznaueria barnesiae, Watznaueria biporta, and Cyclagelosphaera margerelii, and Crepidolithus sp. (Fig. 3). Among them, Watznaueria biporta is clearly recognized by the two well-defined pores separated by a conjunct transverse bar, and Polycostella beckmannii is clearly distinguishable by its globular and stellate nannolith shape composed by five to eight elements of calcite (Thierstein, 1971). The aforementioned species are the only taxa recovered in BACF-NP 4220-4224 (MPCA 632). No other species has been found and, therefore, it is not possible to characterize the assemblage more precisely.

Nannoflora identified in the sediment of MPCA 632: Cyclagelosphaera margerelii (1–4), Crepidolithus sp. (5–6), Watznaueria barnesiae (7–8), Polycostella beckmannii (9–12), Watznaueria britannica (13), Watznaueria fossacincta (14), Watznaueria biporta (15–16)
In general, watznauerids with moderately-to well-preserved specimens may suggest oligotrophic surface waters (Tremolada et al., 2009). However, other authors consider that Watznaueria is a robust eutrophic taxon rather than a true oligotrophic indicator (Lees et al., 2005). This means that fluctuations in the percentage of abundances of this taxon probably do not represent a direct response to environmental changes, but may reflect a measure of the success of other species (Aguado et al., 2014; Omarini et al., 2020) or diagenetic processes. Attending to the high percentage of micritic carbonate and rhombohedral crystals of calcite in the studied sample, we consider that diagenesis is responsible for the absence of other microfossil taxa. The placoliths found in BACF-NP 4220–4224 (MPCA 632) present a high degree of corrosion of their shields, which is due to dissolution, however, some of them exhibit high birefringence indicating strong overgrowth. Therefore, the high abundances of watznauerids, rare specimens of Crepidolithus sp., and the scarce presence of the massive nannolith Polycostella beckmannii are understood as the consequence of strong diagenetic alteration and dissolution of fragile taxa.
Geochemistry
Our data reveal that the δ13C values of the MPCA 632 sample range from 1,97 to 1,99‰ with the average being 1,98‰ (Additional file 1). On the other hand, the values of δ18O in the MPCA 632 sample range from –2.20 to –2,18‰, with an average of –2,19‰ (Additional file 1).
Discussion
Provenance of MPCA 632
The presence of calcareous nannofossils in the MPCA 632 sample leaves no doubts about the unfeasibility that MPCA 632 was collected in the Early Triassic Vera Formation of the Los Menucos Group as indicated by Bogan et al. (2013). Coccolithophores and calcareous nannofossils first appear in the fossil record in Late Triassic marine sediments (Bown et al., 2004; Demangel et al., 2023). During the late Carnian (approximately 227 Ma) they had a consistent presence, increasing their richness and abundance during the Norian and Rethian times, followed by a significant extinction at the end of the Triassic. Rates of speciation were significantly high in the Late Triassic, Early Jurassic and Tithonian-Berriasian (Bown et al., 2004). Therefore, according to the most recent geochronological dating of the Vera Formation in the Lower Triassic (Falco et al., 2020), the provenance of MPCA 632 from this geological unit is untenable. Furthermore, the fossiliferous content and tectonic setting of the Vera Formation are clearly continental.
The inventory card corresponding to MPCA 632 was recently found at the Museo Carlos Ameghino. The locality indicated in this card is “Las Lajas, Neuquen” (Fig. 2b). Geological surveys summarized in Zapala geological map 3969-I (Leanza et al., 2005) and a Field Guide of the Jurassic of the Neuquén Basin (Gulisano & Gutierrez Pleimling, 1994) show that there are no Triassic outcrops in the vicinity of that locality (Fig. 1). The only outcrops in the area are Jurassic and Cretaceous in age.
Polycostella beckmannii Thierstein is considered a reliable Late Jurassic nannofossil biomarker for the Neuquén Basin (Leanza et al., 2020). Its First Occurrence (FO) defines the base of the NJ20-B calcareous nannofossil Zone in the early Tithonian (Bralower et al., 1989). This nannolith has been recognized in the subsurface of Neuquén Basin (Vennari et al., 2017; Concheyro in Aguirre-Urreta et al., 2019) and in two sections of the Los Catutos Member of the Vaca Muerta Formation. In these sections, Polycostella beckmannii correlates with the Windhauseniceras internispinosum ammonite Zone which has been restricted to the late Tithonian (Riccardi et al., 2011; Zeiss & Leanza, 2010). Recently, the first occurrence (FO) of P. beckmanni has been reported in the lower Tithonian of the Tethyan areas, in the NJT15 calcareous nannofossil biozone (Casellato & Erba, 2021), and is confirmed in the Neuquén Basin (Concheyro & Lescano, 2021). Therefore, due to its stratigraphic value, the sole presence of P. beckmanni in BACF-NP 4220–4224 strongly supports the referral of MPCA 632 to the Tithonian.
The fossil record of Watznaueria fossacincta, W. barnesiae, and Cyclagelosphaera margerelii extend from the Bajocian to the Maastrichtian (Fig. 4). They are long ranging taxa, characteristic of marine environments from the Middle Jurassic until the end of the Cretaceous, and are almost always present in Cretaceous nannofloras. They are also cosmopolitan taxa. Watznaueria biporta, is frequent in marine Cretaceous sediments, but it has also been recorded in the Late Jurassic (Tithonian; Bown et al., 1998; Nannotax 3). In addition, the presence of scarce specimens of Crepidolithus sp. confirms the Jurassic age of the sample, since the genus biochron extends from the Hettangian to the top of Tithonian (Bown & Cooper, 1998; Nannotax 3; Fig. 4).

Stratigraphic distribution of the nannotaxa identified in MPCA 632. Plots von Nannotax 3
The association of Watznaueria fossacincta, Watznaueria barnesiae, Watznaueria biporta, Cyclagelosphaera margerelii, Polycostella beckmannii and Crepidolithus most closely resembles the nannofossil association found in the Los Catutos Member, Vaca Muerta Formation, at Cantera Loma Negra (Scasso & Concheyro, 1999). These same taxa together with the strictly Jurassic Ethmorhabdus gallicus, Crepidolithus crassus, and Schizosphaerella punctulata characterize the nannoflora of the Los Catutos Member, supporting a Tithonian age for this unit (Scasso & Concheyro, 1999). It should be noticed that Ethmorhabdus gallicus, Crepidolithus crassus, and Schizosphaerella punctulate, recognized in Los Catutos (Scasso & Concheyro, 1999) are fragile taxa which would not resist the strong diagenetic process inferred for MPCA 632.
Jurassic outcrops in the area of Las Lajas, Neuquén, are composed by sediments belonging to Cuyo and Mendoza groups (Leanza et al., 2005). Among them, the micritic limestone in MPCA 632 is macroscopically very similar to the 40 samples from three profiles of the Los Catutos Member of the Vaca Muerta Formation analyzed by Scasso and Concheyro (1999).
Compared with the limestones and marls of the Los Catutos Member of the Vaca Muerta formation, the isotopic composition of the MPCA 632 samples is slightly different. Scasso et al. (2005) obtained δ18O values between –3.5 and –5.5‰, with average values of −4.87‰ for limestones and −4.43‰ for marls, and δ13C values between −1 and + 1.55‰ with average of 0.93‰ for limestones and 1.025‰ for marls.
The MPCA 632 samples gave values of δ18O ranged from −2.20‰ to −2.18‰, and the average was −2.19 showing a variation with respect to those (−3.5 to −5.5‰) obtained by Scasso et al. (2005). The oxygen isotopic composition of carbonates is quite sensitive to temperature. Therefore, the negative δ18O values of carbonates would be the result of isotopic fractionation as consequence of an increase in temperature during diagenesis. The isotopic carbon variation observed by Scasso et al. (2005) seems to be caused by local paleoenvironmental conditions during the initial marine transgression in the Neuquén Basin.
The δ13C values of Los Catutos limestones and marls samples representing the marine facies of Los Catutos Member (Scasso et al., 2005) are restricted in a very narrow range varying from −1 to 1.5‰. Our data reveal that the δ13C values of the samples ranged from 1.97 to 1.99‰ with the average being 1.98‰. This carbon isotopic composition was similar to the global seawater of the Tithoniann (Veizer et al., 1999). These values are reflecting optimal conditions for the development of an abundant fauna of marine invertebrates as well as a continuous burial of organic carbon.
In addition, the study on carbonate isotopes carried out by Rodriguez Blanco et al. (2018) in samples from the Tithonian-Early Valanginian succession in the Neuquén Basin show few variations and coincide with the global isotope values (0 and + 3) reports for these stratigraphic interval (Katz et al., 2005). In such a way, the data mentioned in this contribution, although scarce, can be considered similar to those published by the last-mentioned authors.
Considering provenance indicated in the inventory card (Fig. 2b), the nannofossil association, very especially the presence of Polycostella beckmannii (Figs. 3–4), the lithology, the isotopic composition (Additional file 1), and plotting the δ13C values obtained for the MPCA 632 samples together with the extensive data provided by Rodriguez Blanco et al. (2022), it is most likely that MPCA 632 was collected from outcrops of the Los Catutos Member of the Vaca Muerta Formation.
Gouiric-Cavalli (2016) described a new caturid taxon from the Jurassic of Neuquén (Tithonian Los Catutos Member, Vaca Muerta Formation) under the name Catutoichthys olsacheri. Unfortunately, the holotype and only known specimen of Catutoichthys olsacheri is an incomplete fish, including only a few poorly preserved skull bones and a comparison with MPCA 632 is not possible. However, it is likely that MPCA 632 represents a specimen of this taxon.
Distribution of Caturoidei
Confirming the suspicions of López-Arbarello and Ebert (2023), the micropaleontological, chemical and lithological analyses of MPCA 632 demonstrate that the caturoid specimen described by Bogan et al. (2013) does not come from the Lower Triassic of Los Menucos Group. Instead, the specimen must have been collected from outcrops of the Upper Jurassic–Lower Cretaceous Vaca Muerta Formation, most probably from its Tithonian Los Catutos Member.
Eugnathus insignis Kner, 1866, from the Norian of Seefeld, Austria, is the only other record of a caturoid in the Triassic (López-Arbarello & Ebert, 2023). The species was referred to the genus Caturs by Woodward (1895). After close examination of the holotype TLM F.117, the senior author has been able to confidently exclude “Caturus” insignis (Kner, 1866) from the genus Caturus, the family Catuidae or the superfamily Caturoidei (López-Arbarello, pers. obs.). The superfamily Caturoidea is diagnosed by the presence of sharply carinate acrodin tooth caps on the larger jaw teeth; an extremely slender rod-like maxilla; a relatively high number of branchiostegal rays (22 or more on each side); haemal spines broadly spatulate in the transverse plane; preural haemal and neural spines near the caudal peduncle region strongly inclined to a nearly horizontal orientation (Grande & Bemis, 1998). None of these features is present in the holotype of Eugnathus insignis Kner, 1866, which nevertheless share an overall resemblance with caturoids and might be an early member on the lineage leading to this superfamily (López-Arbarello, pers. obs.). The generic name Eugnathus was preoccupied and replaced with Furo Gistel, 1848. Therefore, excluded from Caturus and pending its systematic revision, the name of the species is Furo insignis (Kner, 1866).
With the exclusion of Furo insignis from the group, the fossil record of Caturoidea is restricted to the Jurassic–Lowest Cretaceous (López-Arbarello & Ebert, 2023: table 1). Except for the youngest records in the Lower Cretaceous of Spain (El Montsec and Las Hoyas, Martín-Abad & Poyato-Ariza, 2013), all other caturoid records are from marine environments. Caturoid Bauplan, streamlined body and narrow caudal peduncle, indicate a pelagic habitat (Friedman et al., 2020). The first confident record of a caturoid outside todays Europe is “Caturus” dartoni Eastman, 1899, from the Bathonian (lower Sundance Formation) of Hot Springs, South Dakota, in North America. The holotype of “C.” dartoni (NMNH 4792) is a very incomplete specimen and none of the diagnostic features of the Caturoidea is preserved. However, other specimens from the Sundance and Wanakah formations were studied by Schaeffer and Patterson (1984), who discussed the resemblance of this species with European species of Caturus, including several features: the morphology of the jaws, dentition, paired fins and scales, the presence of at least 24 branchiostegal rays and hemichordacentra. In particular they highlighted the fusion of hypurals 1–3, a feature shared with the Early Jurassic Caturus heterurus from Lyme Regis, UK, and C. smithwoodwardi, from Holzmaden, Germany.
The next younger record of a Caturoid west of the Tethys is Caturus deani Gregory, 1923, from the Upper Jurassic Jagua Formation (Oxfordian) of Cuba (Iturralde-Vinent & Ceballos Izquierdo, 2015). The holotype of this species, an imperfectly preserved skull AMNH FF 6371 (7930), shows a very slender maxilla distinct of caturoid fishes. Later on, in the Tithonian, caturoids are recorded in the Vaca Muerta Formation of Argentina (Gouiric-Cavalli, 2016 and this study). Therefore, according to their known fossil record, the dispersal of caturoids towards the west most probably occurred through the Hispanic Corridor (Fig. 5) and started in the Middle Jurassic as suggested by López-Arbarello and Ebert (2023).

Simplified global paleogeograhic reconstruction during the Late Jurassic, based on Scotese (2014: Map 35, Oxfordian) showing potential dispersal routes of caturoids through the Hispanic Corridor. Stars represent the only known records of caturoids outside Europe: 1, “Caturus” dartoni Eastman, 1899, from the lower Sundance Formation in South Dakota (Bathonian); 2, Caturus deani Gregory, 1923, from the Jagua Formation of Cuba (Oxfordian); 3, Catutoichthys olsacheri and Caturoidea sp. from the Vaca Muerta Formation of Argentina (Tithonian)
The Hispanic Corridor started developing during the Sinemurian, probably intermittently at those early stages, and it was well-established as a shallow marine connection by the Pliensbachian–Toarcian (Damborenea et al., 2012). Studies on bivalves show that, during the Early Jurassic, the Hispanic Corridor functioned as a filter, allowing the dispersion of benthonic littoral species and simultaneously representing a barrier for neritic species (Aberham, 2001; Damborenea et al., 2012). The Hispanic Corridor became an effective dispersal route for neritic taxa only during the Middle Jurassic, during the rift-drift transition and the opening of the Caribbean Seaway (Damborenea et al., 2012; Gasparini et al., 2000; Pindell et al., 2021). Therefore, the first West appearance of a caturoid, “Caturus” dartoni in the Sundance Sea coincides with the establishment of this very successful spreading path which had driven the evolution of other vertebrate groups, in particular the marine reptiles (Bardet et al., 2014).
Conclusions
The micropaleontological and geochemical analyses resulted in the identification of the bearing rock in MPCA 632 as sediments of the Vaca Muerta Formation (Upper Jurassic), probably its Los Catutos Member (Tithonian) of Argentina. The closest resemblance of the nannoflora found in the slides BACF-NP 4220–4224 with the association reported for the Los Catutos Member and, very specially, the presence of the Late Jurassic biomarker Polycostella beckmannii definitely indicate both the marine origin and the Tithonian age of specimen MPCA 632. Furthermore, the oxygen and carbon stable isotopic composition of bulk-rock samples of MPCA 632 agree with the values known for the Late Jurassic in general, and those of the Vaca Muerta Formation in particular. These results, combined with the locality “Las Lajas, Neuquén” indicated in the inventory card, and the macroscopic comparison of the lithology unquestionably exclude the possibility that MPCA 632 was found in continental Lower Triassic outcrops of the Vera Formation (Los Menucos Group), and warrant its provenance from outcrops of the Upper Jurassic Vaca Muerta Formation (probably Los Catutos Member; Mendoza Group).
The anatomical comparison of the fossil fish specimen MPCA 632 led to its exclusion from the genus Caturus, although its referral to the superfamily Caturoidea is well supported. The presence of Catutoichthys olsacheri in the Los Catutos Member of the Vaca Muerta Formation (Gouiric-Cavalli, 2016) strongly suggest that MPCA 632 might be a specimen of this species. However, since the anatomical structures preserved in the holotype and only known specimen of Catutoichthys olsacheri and MPCA 632 are not comparable, this hypothesis will have to be tested when additional material becomes available.
After excluding the Triassic Furo insignis from the group, and correcting the record of MPCA 632, the superfamily Caturoidea is restricted to the Jurassic–lowest Cretaceous (López-Arbarello & Ebert, 2023: table 1). As far as currently known, caturoids inhabited the Western Tethys during the Early Jurassic and dispersed westward into the Paleo-Pacific across the Hispanic Corridor during the Middle and Late Jurassic. This pattern of dispersion and the known fossil record of caturoids strongly indicates that after a modest evolution during the Early Jurassic, the group had its initial radiation during the Middle Jurassic and reached its maximal diversity during the Kimmeridgian and Tithonian.
Availability of data and materials
All data generated or analyzed during this study are included in this published article [and its Additional files].
References
Aberhan, M. (2001). Bivalve palaeobiogeography and the Hispanic Corridor: Time of opening and effectiveness of a proto-Atlantic seaway. Palaeogeography, Palaeoclimatology, Palaeoecology, 165(3–4), 375–394.
Aguado, R., Company, M., O’dogherty, L., Sandoval, J., & Tavera, J. M. (2014). Late Hauterivian-Early Barremian calcareous nannofossil biostratigraphy, palaeoceanography, and stable isotope record in the Subbetic domain (southern Spain). Cretaceous Research, 49, 105–124.
Aguirre-Urreta, B., Naipauer, M., Lescano, M., López-Martínez, R., Pujana, I., Vennari, V., De Lena, L. F., Concheyro, A., & Ramos, V. A. (2019). The Tithonian chrono-biostratigraphy of the Neuquén Basin and related Andean areas: A review and update. Journal of South American Earth Sciences, 92, 350–367.
Artabe, A. E. (1985). Estudio sistemático de la Tafoflora triásica de Los Menucos, Provincia de Río Negro, Argentina. Parte I, Sphenophyta. Filicophyta y Pteridospermophyta. Ameghiniana, 22, 3–22.
Artabe, A. E. (1986). Estudio sistemático de la Tafoflora triásica de Los Menucos, Provincia de Río Negro, Argentina. Parte II. Cycadophyta. Ginkgophyta y Coniferophyta. Ameghiniana, 22, 159–280.
Bardet, N., Falconnet, J., Fischer, V., Houssaye, A., Jouve, S., Suberbiola, X. P., Pérez-García, A., Rage, J.-C., & Vincent, P. (2014). Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Research, 26(3–4), 869–887.
Bogan, S., Taverne, L., & Agnolin, F. (2013). First Triassic and oldest record of a South American amiiform fish: Caturus sp. from the Los Menucos Group (lower Upper Triassic), Rio Negro province. Argentina. Geologica Belgica, 16(3), 191–195.
Bown, P. R., & Cooper, M. K. E. (1998). Jurassic. In P. R. Bown (Ed.), Calcareous Nannofossil Biostratigraphy (pp. 34–85). British Micropalaeontological Society Publication Series, London.
Bown, P. R., Lees, J., & Young, J. (2004). Calcareous nannoplankton evolution and diversity through time. In H. R. Thierstein & J. R. Young (Eds.), Coccolithophores from molecular processes to global impact (pp. 481–508). Springer.
Bown, P. R., Rutledge, D. C., Crux, J. A., & Gallagher, L. T. (1998). Early Cretaceous. In P. R. Bown (Ed.), Calcareous Nannofossil Biostratigraphy (pp. 86–131). British Micropalaeontological Society Publications Series, London.
Bown, P. R., & Young, J. R. (1998). Techniques. In P. R. Bown (Ed.), Calcareous Nannofossil Biostratigraphy (pp. 16–28). British Micropalaeontological Society Publications Series, London.
Bralower, T. J., Monechi, S., & Thierstein, H. R. (1989). Calcareous nannofossil zonation of the Jurassic-Cretaceous boundary interval and correlation with the geomagnetic polarity timescale. Marine Micropaleontology, 14, 153–235.
Brownstein, C. D., Kim, D., Orr, O. D., Hogue, G. M., Tracy, B. H., Pugh, M. W., Singer, R., Myles-McBurney, C., Mollish, J. M., Simmons, J. W., David, S. R., Watkins-Colwell, G., Hoffman, E. A., & Near, T. J. (2022). Hidden species diversity in an iconic living fossil vertebrate. Biological Letters. https://doi.org/10.1098/rsbl.2022.0395
Casellato, C., & Erba, E. (2021). Reliability of calcareous nannofossil events in the Tithonian–early Berriasian time interval: implications for a revised high resolution zonation. Cretaceous Research. https://doi.org/10.1016/j.cretres.2020.104611
Cione, A., Gasparini, Z., Leanza, H. A., & Zeiss, A. (1987). Marine Oberjurassische Plattenkalke in Argentinien. Archaeopteryx, 5, 13–22.
Citton, P., Díaz-Martínez, I., de Valais, S., & Cónsole-Gonella, C. (2018). Triassic pentadactyl tracks from the Los Menucos Group (Río Negro province, Patagonia Argentina): possible constraints on the autopodial posture of Gondwanan trackmakers. PeerJ. https://doi.org/10.7717/peerj.5358
Codorniú, L., & Gasparini, Z. (2013). The Late Jurassic pterosaurs from northern Patagonia, Argentina. Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 103, 1–10.
Concheyro, A., & Lescano, A. (2021). Calcareous nannofossil assemblages and their significance in defining the Jurassic/Cretaceous boundary. Simposios CAPA 2021: Libro de Resúmenes, 86.
Cope, E. D. (1871). Observations on the systematic relations of the fishes. The American Naturalist, 5(8/9), 579–593.
Cope, E. (1887). Zittel’s manual of palaeontology. American Naturalist, 21(1014), e1019.
Cuchi, R., Busterors, A., & Lema., H. (2001). Hoja Geológica 4169-II, Los Menucos. Provincia de Río Negro. Boletín 265, Instituto de Geología y Recursos Minerales, Servicio Geológico Minero Argentino, Buenos Aires.
Damborenea, S. E., Echevarría, J., & Ros-Franch, S. (2012). Southern hemisphere palaeobiogeography of Triassic-Jurassic marine bivalves. SpringerBriefs Seaways and Landbridges: Southern Hemisphere Biogeographic Connections Through Time. Springer.
Demangel, I., Kovács, G., & S., Krystyn, L., Piller, W, Baldermann, A., & Richoz, S. (2023). Fate of calcareous nannofossils during the Rhaetian Late Triassic): Evidence from the Northern Calcareous Alps. Austria. Lethaia, 56(1), 1–24.
Díaz-Martínez, I., & de Valais, S. (2014). Estudio de la variabilidad en la conservación de huellas de tetrápodos del Triásico Superior de Los Menucos, Río Negro. Argentina. Ameghiniana, 52, 8.
Eastman, C. R. (1899). Jurassic fishes from Black Hills of South Dakota. Geological Society of America Bulletin, 10(1), 397–408.
Ebert, M. (2018). Cerinichthys koelblae, gen. et sp. Nov., from the Upper Jurassic of Cerin, France, and its phylogenetic setting, leading to a reassessment of the phylogenetic relationships of Halecomorphi (Actinopterygii). Journal of Vertebrate Paleontology, 38(1), e1420071.
Edwards, A. (1963). A preparation technique for calcareous nannoplankton. Micropaleontology, 9, 103–104.
Falco, J. I., Bodnar, J., & Del Río, D. (2020). Revisión Estratigráfica del Grupo Los Menucos, Pérmico tardío - triásico temprano del Macizo Nordpatagónico, Prov. de Río Negro, Argentina. Revista De La Asociación Geológica Argentina, 77, 532–555.
Friedman, G.M. (1982). Two new carbonate stable-isotope standards. Geostandards Newsletter, 6, 11–12.
Friedman, S. T., Price, S. A. , Corn, K. A., Larouche, O., Martinez, C. M., & Wainwright, P. C. (2020). Body shape diversification along the benthic–pelagic axis in marine fishes. Proceedings of the Royal Society B, 287, 20201053.
Gasparini, Z., Fernández, M. S., De La Fuente, M., Herrera, Y., Codorniú, L., & Garrido, A. (2015). Reptiles from lithographic limestones of the Los Catutos Member (middle–upper Tithonian), Neuquén Province, Argentina: An essay on its taxonomic composition and preservation in an environmental and geographic context. Ameghiniana, 52, 1–28.
Gasparini, Z., Vignaud, P., & Chong, G. (2000). The Jurassic Thalattosuchia (Crocodyliformes) of Chile; a paleobiogeographic approach. Bulletin De La Société Géologique De France, 171(6), 657–664.
Gistel, J. (1848). Naturgeschichte des Thierreichs für höhere Schulen, X. Pisces. Stuttgart: Hoffman'sche Verlags-Buchhandlung.
Gouiric-Cavalli, S. (2016). A new Late Jurassic halecomorph fish from the marine Vaca Muerta Formation, Argentina, southwestern Gondwana. Fossil Record, 19(2), 119–129.
Grande, L., & Bemis, W.E. (1998). A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy: An empirical search for interconnected patterns of natural history. Memoirs of the Society of Vertebrate Paleontology, 4.
Grande, L. (2010). An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy (p. 6). American Society of Ichthyologists and Herpetologists, Special Publication.
Gregory, W. K. (1923). A Jurassic fish fauna from western Cuba: With an arrangement of the families of holostean ganoid fishes. Bulletin of the American Museum of Natural History, 48, 223–242.
Gulisano, C.A., & Gutiérrez Pleimling, A. (1994). Field guide: The Jurassic of the Neuquén Basin. a) Neuquén province. Serie E –N| 2, 111 pgs. Asociación Geológica Argentina.
Hay, O.P. (1929). Second bibliography and catalogue of the fossil Vertebrata of North America. Carnegie Institution of Washington, Publication 390, vol. 1.
Huxley, T. H. (1861). Preliminary essay upon the systematic arrangement of the fishes of the Devonian epoch. Memories of the Geological Survey of the United Kingdom, 10, 1–40.
Iturralde-Vinent, M., & Ceballos Izquierdo, Y. (2015). Catalogue of Late Jurassic vertebrate (Pisces, Reptilian) specimens from western Cuba. Paleontología Mexicana, 3(65), 24–39.
Katz, M. E., Wright, J. D., Miller, K. G., Cramer, B. S., Fennel, K., & Falkowski, P. G. (2005). Biological overprint of the geological carbon cycle. Marine Geology, 217(3–4), 323–338.
Kner, R. (1866). Die Fische der bituminösen Schiefer von Raibl in Kärnthen. Sitzungsberichte der mathematisch-naturwissenschaftlichen Classe der Kaiserlichen Akademie der Wissenschaften Wien, 53, 152–197.
Labudía, C. H., & Bjerg, E. A. (2001). El Grupo Los Menucos: Redefinición estratigráfica del Triásico superior del Macizo Nordpatagónico. Revista De La Asociación Geológica Argentina, 56, 404–407.
Lambers, P. H. (1992). On the ichthyofauna of the Solnhofen Lithographic Limestone (Upper Jurassic. University of Groningen.
Lambers, P. H. (1994). The halecomorph fishes Caturus and Amblysemius in the lithographic limestone of Solnhofen (Tithonian), Bavaria. Geobios, 27, 91–99.
Leanza, H.A., Hugo, C., Repol, D., González, R., Danieli, J. C., & Lizain, A. (2001). Hoja Geológica 3969-I, Zapala. Provincia del Neuquén. Boletín 275, Servicio Geológico Minero Argentino. Instituto de Geología y Recursos Minerales.
Leanza, H.A., Vennari, V., Aguirre–Urreta, M.B., Concheyro, A., Lescano, M. Ivanova, D., Kietzmann, D., López–Martínez, R., Martz, P., Melisa, M., Paolillo, A., Guler, V., Pujana, I., & Paz, M. (2020). Relevant marine paleobiological markers of the Vaca Muerta Formation. In D. Minisini, M. Fantín, I. Lanusse, & H. Leanza (Eds.), Integrated Geology of Unconventionals: The Case of the Vaca Muerta Play, Argentina (pp. 61–98). American Association Petroleum Geologists, Memoir 121.
Leanza, H. A., Hugo, C. A., Repol, D., Gonzalez, R., & Danieli, J. C. (2005). Hoja Geológica 3969-I, Zapala. Instituto De Geología y Recursos Minerales, Boletin, 275, 1–131.
Leanza, H. A., Sattler, F., Martínez, R. S., & Carbone, O. (2011). La Formación Vaca Muerta y sus equivalents (Jurásico Tardío-Cretácico Temprano) en la Cuenca Neuquina. In H. A. Leanza, C. Arregui, O. Carbone, J. C. Danieli, & J. M. Vallés (Eds.), Geología y Recursos Minerales de la Provincia del Neuquén: Relatorio del XVIII Congreso Geológico Argentino (pp. 113–129). Asociación Geológica Argentina.
Leanza, H. A., & Zeiss, A. (1990). Upper Jurassic lithographic limestones from Argentina (Neuquén Basin): Stratigraphy and fossils. Facies, 22, 169–186.
Lees, J., Bown, P., & Mattioli, E. (2005). Problem with proxies? Cautionary tales of calcareous nannofossil paleoenvironmental indicators. Micropaleontology, 51(5), 333–343.
Legarreta, L., & Uliana, M. A. (1996). The Jurassic succession in west-central Argentina: Stratal patterns, sequencesand paleogeographic evolution. Palaeogeography, Palaeoclimatology, Palaeoecology, 20, 303–330.
Legarreta, L., & Villar, H. J. (2015). The Vaca Muerta Formation (Late Jurassic – Early Cretaceous), Neuquén Basin, Argentina: Sequences, facies and source rock characteristics. Unconventional Resources Technology Conference (URTeC). https://doi.org/10.15530/urtec-2015-2170906
López-Arbarello, A., & Ebert, M. (2023). Taxonomic status of the caturid genera (Halecomorphi, Caturidae) and their Late Jurassic species. Royal Society Open Science, 10, 221318.
López-Arbarello, A., & Sferco, E. (2018). Neopterygian phylogeny: The merger assay. Royal Society Open Science, 53, 172337.
Luppo, T., López de Luchi, M., Rapalini, A., Martínez Dopico, C., & Fanning, M. (2018). Geochronologic evidence of a large magmatic province in northern Patagonia encompassing the Permian-Triassic boundary. Journal South American Earth Sciences, 86, 335–346.
Martín-Abad, H., & Poyato-Ariza, F. J. (2013). Historical patterns of distribution in pycnodontiform and amiiform fishes in the context of moving plates. Geologica Belgica, 16(4), 217–226.
Müller, J. (1845). Über den Bau und die Grenzen der Ganoiden und das natürliche System der Fische. Abhandlungen Der Akademie Der Wissenschaften Zu Berlin, Aus Dem Jahr, 1844, 177–216.
Omarini, J., Lescano, M., Odino-Barretod, A., Campetella, D., Tunik, M., Garbán, G., Brea, F., Erra, G., Aguirre-Urreta, B., & Martinez, M. (2020). Paleoenvironmental conditions for the preservation of organic matter during the late Hauterivian in the Neuquén Basin (Western Argentina). Marine and Petroleum Geology, 120, 104469.
Otharán, G., Zavala, C., Arcuri, M., Di Meglio, M., Zorzano, A., Marchal, D., & Köhler, G. (2020). Análisis de facies en depósitos de grano fino asociados a flujos de fango. Formación Vaca Muerta (Tithoniano-Valanginiano), Cuenca Neuquina central. Argentina. Andean Geology, 47, 384–417.
Owen, R. (1860). Palaeontology, or a Systematic Summary of Extinct Animals and Their Geological Relations. Adam and Charlos Black, Edinburgh.
Pindell, J., Villagómez, D., Molina-Garza, R., Graham, R., & Weber, B. (2021). A revised synthesis of the rift and drift history of the Gulf of Mexico and surrounding regions in the light of improved age dating of the Middle Jurassic salt. Geological Society, London, Special Publications, 504(1), 29–76.
Regan, C.T. (1923). The skeleton of Lepidosteus, with remarks on the origin and evolution of the lower neopterygian fishes. Proceedings of the Zoological Society of London, 445–461.
Riccardi, A. C., Damborenea, S. E., Manceñido, M. O., & Leanza, H. A. (2011). Megainvertebrados del Jurásico y su importancia geobiológica. In H. A. Leanza, C. Arregui, O. Carbone, J. C. Danieli, & J. M. Vallés (Eds.), Geología y Recursos Minerales de la Provincia del Neuquén: Relatorio del XVIII Congreso Geológico Argentino (pp. 441–464). Asociación Geológica Argentina.
Rodriguez Blanco, L., Eberli, G.P., Swart, P.K., Weger, R.J., & Tenaglia, M. (2018). Stable Isotope Signature of the Tithonian-Valanginian Strata of the Vaca Muerta-Quintuco System in the Neuquen Basin. 10 Congreso de Exploracion y Desarrollo de Hidrocarburos, Mendoza, Argentina. Actas Simposio de Recursos No Convencionales, 43–59.
Rodriguez Blanco, L., Eberli, G. P., Weger, R. J., Swart, P., Tenaglia, M., Rueda, L., & McNeill, D. (2020). A Periplatform Ooze within a mixed Siliciclastic-Carbonate System—Vaca Muerta Formation, Neuquén Basin. Sedimentary Geology. https://doi.org/10.1016/j.sedgeo.2019.105521
Rodriguez Blanco, L., Swart, P. K., Eberli, G. P., & Weger, R. J. (2022). Negative δ13Ccarb values at the Jurassic-Cretaceous boundary – Vaca Muerta Formation, Neuquén Basin, Argentina. Palaeogeography, Palaeoclimatology, Palaeoecology, 603, 111208.
Scasso, R. A., Alonso, M. S., Lanés, S., Villar, H. J., & Laffitte, G. (2005). Geochemistry and petrology of a Middle Tithonian limestone in the Neuquén Basin, Argentina: depositional and burial history. In G. D. Veiga, L. A. Spalletti, J. A. Howell, & E. Schjwarz (Eds.), The Neuquén Basin, Argentina: A Case Study in Sequence Stratigraphy and Basin Dynamics (pp. 207–229) (p. 252). London, Special Publications: Geological Society.
Scasso, R., & Concheyro, A. (1999). Nanofósiles calcáreos, duración y origen de ciclos caliza-marga (Jurásico tardío de la Cuenca Neuquina). Revista De La Asociación Geológica Argentina, 54(3), 290–297.
Schaeffer, B., & Patterson, C. (1984). Jurassic fishes from the Western United States with comments on Jurassic fish distribution. American Museum Novitates, 2796, 1–86.
Sun, Z. Y., Tintori, A., Xu, Y. Z., Lombardo, C., Ni, P. G., & Jiang, D. Y. (2017). A new non-parasemionotiform order of the Halecomorphi (Neopterygii, Actinopterygii) from the Middle Triassic of Tethys. Journal of Systematic Palaeontology, 15(3), 223–240.
Thierstein, H. R. (1971). Tentative Lower Cretaceous Calcareous Nannoplankton Zonation. Eclogae Geologicae Helvetiae, 64, 459–488.
Tremolada, F., Erba, E., de Bernardi, B., & Cecca, F. (2009). Calcareous nannofossil fluctuations during the Late Hauterivian in the Cismon core (Venetian Alps, northeastern Italy) and in selected sections of the Umbria-Marche basin (central Italy): Paleoceanographic implications of the Faraoni level. Cretaceous Research, 30, 505–514.
Veizer, J., Ala, D., Azmy, K., Bruckschen, P., Buhl, D., Bruhn, F., Carden, G. A. F., Diener, A., Ebneth, S., Godderis, Y., Jasper, T., Korte, Ch., Pawellek, F., Podlaha, O. G., & Strauss, H. (1999). 87Sr/86Sr, d 13C and d 18O evolution of Phanerozoic seawater. Chemical Geology, 161, 59–88.
Vennari, V., Lescano, M., Aguirre-Urreta, B., Fantín, M., Vallejo, M.D., Depine, G., Sagasti, G., & Ambrosio, A. (2017). Bioestratigrafía de alta resolución de la Formación Vaca Muerta: amonites y nanofósiles calcáreos integrando datos de subsuelo y de afloramientos. XX Congreso Geológico Argentino, S4 Prospección y desarrollo de yacimientos convencionales y no convencionales y desarrollo en campos maduros (S.M. de Tucumán), Actas, 168–172.
Woodward, A. S. (1895). Catalogue of the fossil fishes in the British Museum (Natural History) (Vol. 3). London: British Museum.
Wright, J. J., Bruce, S. A., Sinopoli, D. A., Palumbo, J. R., & Stewart, D. J. (2022). Phylogenomic analysis of the bowfin (Amia calva) reveals unrecognized species diversity in a living fossil lineage. Scientific Reports, 12, 16514.
Xu, G.-H. (2019). Osteology and phylogeny of Robustichthys luopingensis, the largest holostean fish in the Middle Triassic. PeerJ, 7, e7184.
Zeiss, A., & Leanza, H. A. (2010). Upper Jurassic (Tithonian) ammonites from the lithographic limestones of the Zapala region, Neuquén Basin, Argentina. Beringeria, 41, 25–75.
Acknowledgements
Sergio Bogan and Sebastián Apesteguía provided helpful information on the fossil MPCA 632. Ignacio Cerda located the fossil and the register card at the MPCA, and provided the rock samples for the analyses. Maria Schaffhauser (TLM) for access to the material under her care. Yanina Herrera and very specially Susana Damborenea for very helpful consultations about the evolution of the Hispanic Corridor. Patricia Alvarez for the preparation of micropaleontological samples and Michael Joachimski for the geochemical analysis of the bulk-rock samples. We wish to thank the editors, an anonymous reviewer, and very especially Martin Ebert for his very helpful comments on an earlier version of our paper.
This is the contribution R-458 of the Instituto de Estudios Andinos “Don Pablo Groeber”.
Funding
Open Access funding enabled and organized by Projekt DEAL. German Science Foundation (DFG) through project LO1405/8-1 to ALA.
Author information
Authors and Affiliations
Contributions
ALA, BAU, designed research. ALA, studied the fossil fish. AC, carried out the micropaleontological analysis. RP, discussed the results of the geochemical analysis. All authors contributed to writing the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Lionel Cavin.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
The early fossil record of Caturoidea (Halecomorphi: Amiiformes): biogeographic implications.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
López-Arbarello, A., Concheyro, A., Palma, R.M. et al. The early fossil record of Caturoidea (Halecomorphi: Amiiformes): biogeographic implications. Swiss J Palaeontol 142, 33 (2023). https://doi.org/10.1186/s13358-023-00297-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-023-00297-z