
Abstract
Understanding the diversity of deep-sea fish fauna based on otoliths in the tropical and subtropical West Pacific has been limited, creating a significant knowledge gap regarding regional and temporal variations in deep-sea fish fauna. To address this gap, we collected a total of 122 bulk sediment samples from the Lower Pliocene Gutingkeng Formation in southwestern Taiwan to reconstruct the otolith-based fish fauna. Using planktonic foraminiferal biostratigraphy, we determined the age of the samples to be 5.6 to 3.1 Ma. A total of 8314 otoliths were assigned to 64 different taxa from 33 families, including the discovery of one new genus, Gutingichthys gen. nov., and three new species: Benthosema duanformis sp. nov., Benthosema parafibulatum sp. nov., and Gutingichthys changi sp. nov. Comparisons with other regional otolith-based assemblages highlighted the exceptional diversity of our collection, making it the most diverse fossil fish fauna reported from Taiwan to date. Otolith diversity analysis revealed very few taxa were dominant in the assemblage, particularly the mesopelagic Myctophidae, with a wide variety of minor taxa. The co-occurrence of shallow-water elements suggests episodic storm events as a potential source. The predominance of deep-sea and oceanic fishes indicated an outer-shelf to upper slope environment, resembling the modern outer-shelf and upper slope fish fauna in the region. Our findings suggest an early establishment and persistent presence of the mesopelagic fish community since the Early Pliocene. Further investigations of the Upper Miocene and Pleistocene sections of the Gutingkeng Formation would provide valuable insights into the evolution of deep-sea fish fauna in the area.
Similar content being viewed by others

Introduction
Marine fish fossils in Taiwan have primarily come from shallow-water deposits, representing nearshore, coastal, and shallow-water communities (Lin et al., 2018; Lin & Chien, 2022; Lin et al., 2022a, 2022b, 2022c). Fish otoliths have emerged as the richest source of fish fossils in Taiwan, surpassing skeletal records due to better preservation potential (Lin et al., 2021). However, our understanding of paleo-deep-water and pelagic fish communities in Taiwan remains limited. One possible deep-water fish fossil is a large otolith of Coelorinchus discovered in the Plio–Pleistocene Maanshan Mudstone of the Hengchun Peninsula, as depicted by Hu (1990, pl. 8, figs. 2, 8, mistakenly cited as Hu, 1989 in Lin et al., 2021).
Existing records of relevant otoliths from the northwest Pacific are primarily confined to temperate Japan with significant studies conducted by Ohe (1977, 1981, 1983). Comprehensive reviews of otolith-based fossils from mesopelagic lanternfish (Myctophidae) in Japan have been provided by Schwarzhans and Ohe (2019) and Schwarzhans et al. (2022). Notably, van Hinsbergh and Helwerda (2019) conducted an extensive study on a deep-sea Plio–Pleistocene otolith assemblage in the northwest Philippines. In southern Taiwan, the Gutingkeng Formation, a thick mudstone series, is well-known for its well-preserved foraminifera and nannofossils (Horng & Shea, 1994; Hu, 1989; Shieh et al., 2002), as well as ostracods and solitary corals (Hu, 1989; Hu & Tao, 1982). This formation is believed to have been deposited in a relatively deeper-water setting compared to other shallow-water layers in the Western Foothills of Taiwan (Chen & Yu, 2016) and the formation has the potential to yield deep-sea fossil assemblages.
In this study, we conducted quantitative sampling from the lower part of the Gutingkeng Formation in the Tainan and Kaohsiung areas (Fig. 1) between 2020 and 2022. The collection, which was dated using planktonic foraminiferal biostratigraphy, provides one of the most comprehensive and richest records of outer-shelf and pelagic fish communities from the Zanclean (Early Pliocene) subtropical West Pacific.

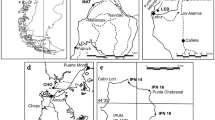
Maps showing the sampling sites. a Overview of geological map of Taiwan (modified after Chen, 2016). b Detailed geological map of study area (map extracted from National Geological Data Warehouse, Central Geological Survey, Ministry of Economic Affairs, Taiwan). c–d Detail maps of the sampling sites in Chochen, Tainan (c) and Tianliao, Kaohsiung (d)
Geological setting
The Gutingkeng Formation is exposed vastly in the southwestern region of Taiwan (Torii, 1932), spanning an area of approximately 750 square kilometers in the Tainan and Kaohsiung areas (Keng, 1981; Chen & Yu, 2016; Fig. 1; Additional file 1: Figs. S1, S2). This formation displays considerable thickness, reaching 2280 m in Tainan and over 3600 m in the Kaohsiung area (Chen & Yu, 2016). It primarily consists of massive dark gray mudstone and shale, occasionally intercalated with thick beds of muddy sandstone (Keng, 1981; Lin, 1990; Luo, 1988). Within the formation, several notable thick-bedded sandstone lenses can be observed. The extensive barren badlands formed by the massive dark gray mudstone have earned the local nickname “Moon World”.
During the Late Miocene to Early Pliocene, the southwestern basin of Taiwan was predominantly filled with deep-water deposits. However, as the Late Pliocene and Pleistocene progressed, the basin gradually filled and became shallower (Chou, 1971; Covey, 1984; Luo, 1988). The continuous deposition of the Gutingkeng Formation is overlain by the Chiting Formation (Fig. 2). Nannofossil dating has provided evidence of the formation’s age spanning from the Late Miocene to the Pleistocene, although exposures of the Miocene layers are comparatively limited in comparison to the more extensive Plio–Pleistocene deposits (Chi, 1979). Notably, exposures of the latest Miocene and the Miocene–Pliocene boundary can be found along the Erhjen River in Kaohsiung, but investigations at this site have focused solely on nannofossils (Horng & Shea, 1994).

Stratigraphic correlation of the southern part of the Western Foothills in Taiwan (modified after Chen, 2016). The studied formation, the Gutingkeng Formation, is indicated in yellow
The overall sedimentary environment of the lower Gutingkeng Formation is characterized as an outer-shelf to slope setting (Keng, 1981; Lin, 1990; Luo, 1988). Studies of benthic foraminiferal assemblages have indicated the dominance of taxa such as Bolivina, Siphogenerina, Quinqueloculina, and Rotalia, suggesting deposition in a moderately deep, outer-shelf and open-sea environment (Oinomikado, 1955).
Material and methods
Sampling sites and foraminiferal biostratigraphy
Site 1 (Chochen, Tainan; Fig. 1c; Additional file 1: Fig. S1).
Location GPS coordinates:
23°02′25.16''N, 120°25′47.62''E.
Stratigraphy:
Lower Gutingkeng Formation, Early Pliocene.
Sediment Dark gray mudstone.
Foraminifera Presence of Pulleniatina primalis indicates an age of approximately 3.7 Ma (last appearance datum, LAD). The composition of benthic foraminiferal assemblage suggests a mid-shelf environment, slightly shallower than Site 2 (see below).
Available material Five bulk samples weighing a total of approximately 25 kg.
Site 2 (Chochen, Tainan; Fig. 1c; Additional file 1: Fig. S1).
Location GPS coordinates:
23°00′33.31''N, 120°25′24.18''E.
Stratigraphy:
Lower Gutingkeng Formation, Early Pliocene.
Sediment Dark gray mudstone, with abundant fossils.
Foraminifera Presence of P. primalis indicates an age of approximately 3.7 Ma (LAD). The site represents a mid-shelf to outer-shelf environment with depths ranging from 50–200 m.
Available material 53 bulk samples weighing a total of approximately 266 kg.
Site 3 (Chochen, Tainan; Fig. 1c; Additional file 1: Fig. S1).
Location GPS coordinates:
22°59′36.89''N, 120°26′53.24''E.
Stratigraphy:
Lower Gutingkeng Formation, Late Pliocene.
Sediment Dark gray mudstone, rich in foraminifera and pyrites, but no otoliths were found.
Foraminifera Presence of Globoconella inflata indicates an age of approximately 3.13 Ma (first appearance datum, FAD). The site represents an outer-shelf environment with depths approximating 200 m.
Available material Three bulk samples weighing a total of approximately 13.7 kg.
Site 4 (Chochen, Tainan; Fig. 1c; Additional file 1: Fig. S1).
Location GPS coordinates:
23°00′06.20''N, 120°27′15.22''E.
Stratigraphy:
Lower Gutingkeng Formation, Early Pliocene.
Sediment Dark gray mudstone, containing foraminifera, but no otoliths were found.
Foraminifera Presence of Globoturborotalita nepenthes indicates an age of approximately 4.4 Ma (LAD). The site suggests a possible outer-shelf environment with depths approximating 200 m.
Available material Three bulk samples weighing a total of approximately 14.5 kg.
Site 5 (Tianliao, Kaohsiung; Fig. 1d; Additional file 1: Fig. S2).
Location GPS coordinates:
22°50′40.54''N, 120°24′43.41''E.
Stratigraphy:
Lower Gutingkeng Formation, Early Pliocene.
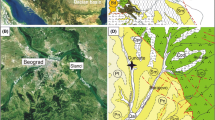
Sediment The exposure at the site is approximately 3 m in height (Fig. 3; Additional file 1: Fig. S2). The top portion is covered, while the lower part consists of thick mudstone interbedded with thin sand layers. The mudstone can be roughly divided into an upper layer of about 1 m in thickness and a lower layer of about 1.8 m in thickness. The upper mud layer is sandwiched by two very thin sand layers, each about 2 cm thick. The upper and lower mud layers are separated by another sand layer, which is about 20 cm thick. The mud is bluish-gray in color and devoid of sedimentary structures. Shell fragments and otoliths can be found on the surface of exposures of the sediments of the lower mud layer, and bulk samples were taken. The sand layers, which are yellow to brownish in color, are unconsolidated. The lowest sand layer includes argillaceous sand, interbedded mud and argillaceous sand, and bioclastic sand. The bioclastic sands are light brown and contain a large amount of biological debris, including shells, fish otoliths, and shark teeth. Both mud and sand layers were sampled for otolith analysis in this study.

Measured section of the outcrops in Site 5 (see text for description). Sampling horizons are indicated
Foraminifera mud layer: Co-occurrence of P. primalis and Sphaeroidinella dehiscens indicates an age range of 5.6 to 3.7 Ma. The site suggests a mid-shelf environment. Bioclastic sand layers: Co-occurrence of Globorotalia menardii and P. primalis indicates an Early Pliocene age. The environment is interpreted as mid-shelf to outer-shelf.
Available material Six bulk samples weighing a total of approximately 27.2 kg were collected from the bioclastic sand layers, while 57 bulk samples weighing a total of approximately 255 kg were collected from the mud layer.
Site 6 (Tianliao, Kaohsiung; Fig. 1d; Additional file 1: Fig. S2).
Location GPS coordinates:
22°51′04.25''N, 120°25′51.66''E.
Stratigraphy:
Lower Gutingkeng Formation, Early Pliocene.
Sediment Dark gray mudstone.
Foraminifera Co-occurrence of S. dehiscens and Sphaeroidinellopsis paenedehiscens indicates an age range of 5.6 to 3.1 Ma. The site suggests a possibly outer-shelf environment with depths approximating 200 m.
Available material Seven bulk samples weighing a total of approximately 35.5 kg.
Otolith preparation and identification
A comprehensive collection of 122 bulk sediment samples, weighing over 600 kg in total, was obtained from various locations in Tainan (n = 58) and Kaohsiung (n = 64). The initial step involved drying and weighing the bulk sediments, followed by the allocation of five kg of sediment for each sample to facilitate subsequent diversity analyses. The sediment was then immersed in water and sieved through a 500-μm mesh screen. Otoliths larger than the 500-μm screen were meticulously hand-picked under a stereomicroscope. The images of the otoliths were captured using a digital camera adapted to a Nikon SMZ1270 stereomicroscope. For each otolith, a stacked image was created using the Helicon Focus software to account for varying depths of field. Subsequently, these images were digitally retouched using Adobe Photoshop.
For otolith identification, various pertinent references were consulted. These included Ohe (1985), Rivaton and Bourret (1999), Schwarzhans (1999, 2013), Schwarzhans and Aguilera (2013), Lin and Chang (2012), and Nolf (2013). In addition, a direct comparison was made with our laboratory’s modern otolith collection, registered under the code CHLOL. Whenever possible, otoliths were identified to the species level. All collected materials are currently stored in the Biodiversity Research Museum, Academia Sinica, Taiwan, and the imaged specimens are archived under the registration code ASIZF.
Diversity analysis
To assess the abundance and distribution of otoliths in the sampled sediments, we calculated the abundance (density) of otoliths in the sediments by dividing the otolith count by the dry sediment weight (kg) for each sample. The number of identified taxa was also compared with respect to sediment weight. We conducted separate analyses for samples from Tainan and Kaohsiung, and the six sand layer samples from Site 5 were analyzed separately from the rest of the mud layer samples, due to their difference in the abundance of otoliths. The results of these analyses were visualized using box plots.
Otolith diversity was evaluated using Hill numbers, which are widely employed in ecology and paleoecology to quantify biological diversity (Hill, 1973). The Hill numbers were calculated for three orders: q = 0 (0D), q = 1 (1D), and q = 2 (2D). The Hill number of 0D represents species richness (number of species), the 1D Hill number represents Shannon diversity (number of abundant species), and the 2D Hill number represents Simpson diversity (number of dominant species) (Chao et al., 2014). As the observed Hill numbers depend on sample coverage, otolith diversity was further estimated using a specimen-based rarefaction method, which provides reliable diversity indices independent of sample sizes (Chao & Jost, 2012; see also Lin et al., 2023). Confidence intervals (95%) were calculated based on 300 bootstrap replicates following the approach described by Hsieh et al. (2016). Rarefaction curves were analyzed and visualized using the iNEXT package in R software (Chao et al., 2014; Hsieh et al., 2016). Additionally, the rank abundance of identified taxa is presented to illustrate the taxonomic composition and relative abundance of the fossil assemblages.
Systematic paleontology
A list of taxa and their abundance from the collection are presented in Table 1. The classification scheme generally follows that of Nelson et al. (2016), while following Davis et al. (2016) for Acropomatiformes (see also Ghedotti et al., 2018; Smith et al., 2022). In addition, the family Microdesmidae is distinguished from the Gobiidae sensu Nelson et al. (2016) due to the distinct small otoliths they possess, which differ from the typical gobiid otoliths. The authorship of higher taxa was determined using the method described by van der Laan et al. (2014), while for genus and species, Fricke et al. (2023) were followed. In the descriptions, the term “otolith” specifically refers to the saccular otolith (sagitta), unless stated otherwise. The general otolith terminology followed the conventions outlined by Nolf (1985, 2013) and Smale et al. (1995). Descriptions and diagnoses for new species and remarks on other taxa are concise, and the synonymy list is limited to nominal species and relevant records from Taiwan and neighboring areas based on published images.
Order Anguilliformes Regan, 1909
Family Congridae Kaup, 1856
Genus Bathycongrus Ogilby, 1898
Bathycongrus sp.
Figure 4a–b

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–b Bathycongrus sp., Site 2, ASIZF 0100597–598. c–d Engraulis japonicus Temminck & Schlegel, 1846, ASIZF 0100599–600, c Site 5, mud layer, d Site 5, sand layer. e, Stolephorus sp., Site 2, ASIZF 0100601. f–g Polyipnus sp., Site 5, sand layer, ASIZF 0100602–603. h Neoscopelus macrolepidotus Johnson, 1863, Site 5, mud layer, ASIZF 0100604. i Gonostomatidae indet., Site 5, sand layer, ASIZF 0100605. 1, ventral view; 2, inner view
Remarks All specimens in the here described collection represent juvenile individuals. These otoliths exhibit typical Bathycongrus characteristics, which include a prominent dorsal depression, a unified sulcus that narrows and extends anteriorly with its colliculum, and an angular ventral rim (Schwarzhans, 2019b).
Order Clupeiformes Bleeker, 1859
Family Engraulidae Gill, 1861
Genus Engraulis Cuvier, 1816
Engraulis japonica Temminck & Schlegel, 1846
Figure 4c–d
1981 Engraulis cf. E. japonicus—Ohe: pl. 1, Fig. 6 [Japan].
1983 Engraulis japonicus—Ohe: pl. 1, Fig. 1 [Japan].
2021 Engraulis japonica—Mitsui et al.: Fig. 4J [Japan].
2023Engraulis japonica—Mitsui et al.: Fig. 4D [Japan].
Remarks The otoliths of Engraulis japonicus can be easily distinguished by their extremely elongated shape (OL/OH = 2.1–2.5), slender profile, funnel-like sulcus, and protruding rostrum (Lin & Chang, 2012: pl. 71). While this species is not uncommon in the collection, the anterior tip and ventral denticles are often not well-preserved.
Genus Stolephorus Lacepède, 1803
Stolephorus sp.
Figure 4e
Remarks The otoliths of Stolephorus exhibit notable similarities to those of E. japonicus (as described above). However, they can be differentiated from the latter by their relatively more compact outline (less elongated, with OL/OH < 1.9) and greater otolith height (Lin & Chang, 2012: pl. 71).
Order Stomiiformes Regan, 1909
Family Gonostomatidae Cocco, 1838
Gonostomatidae indet.
Figure 4i
Remarks The sulcus type of the otolith bears a striking resemblance to that of Gonostoma (Nolf, 2013: pl. 48). However, due to the lack of preservation in the anterior portion of the otolith, a definitive identification cannot be made with certainty.
Family Sternoptychidae Duméril, 1805
Genus Polyipnus Günther, 1887
Polyipnus sp.
Figure 4f, g
Remarks The otoliths of the studied specimens show a distinctive tall shape and thick posterior margin as depicted in Lin et al (2017: Fig. 4C–E). However, identifying them below the genus level may pose challenges due to overlapping interspecific variations and the absence of comprehensive modern otolith reference material. Nevertheless, within our collection, at least two types of otoliths with varying heights were observed (Fig. 4f with OL/OH > 0.7 vs Fig. 4g with OL/OH < 0.6), suggesting the presence of two distinct taxa.
Order Myctophiformes Regan, 1909
Family Neoscopelidae Jordan, 1901
Genus Neoscopelus Johnson, 1863
Neoscopelus macrolepidotus Johnson, 1863
Figure 4h
Remarks A large and thin otolith, characterized by a posterior angle and a vertical posterior rim, is attributed to Neoscopelus macrolepidotus. Comparative modern otoliths of all three species of Neoscopelus from Taiwan are presented in Fig. 5. The otoliths of N. macrolepidotus can be differentiated from other congeners by their more extended rostral part and large ventral area inclining towards anterior, especially evident in larger specimens (Fig. 5a). In addition, the postero-dorsal angle is rounded and less pronounced than that of other congeners. Although the anterior part of the rostrum in our fossil otolith is not preserved, the general outline of the fossil specimen agrees well with those of N. macrolepidotus.

Extant Neoscopelus otoliths from Taiwan. Scale bars = 1 mm. Images are inner views. a–d Neoscopelus macrolepidotus Johnson, 1863, a 25 Mar. 2022, 110.61 mm SL, CHLOL 15935, b 25 Mar. 2022, 75.78 mm SL, CHLOL 16588, c, 25 Mar. 2022, 75.55 mm SL, CHLOL 15902, d, 25 Mar. 2022, 60.10 mm SL, CHLOL 16592. e–h Neoscopelus microchir Matsubara, 1943, e, 21 Jan. 2021, 171.07 mm SL, CHLOL 3283, f 21 Jan. 2021, 116.36 mm SL, CHLOL 3287, g 19 Feb. 2020, 60.75 mm SL, CHLOL 819, h 29 Jul. 2021, 95.46 mm SL, CHLOL 8208. i–l Neoscopelus porosus Arai, 1969, i 24 Jan. 2021, 120.58 mm SL, CHLOL 2127, j 29 Dec. 2021, 100.18 mm SL, CHLOL 13389, k 24 Jan. 2021, 55.30 mm SL, CHLOL 2136, l 19 Feb. 2020, 78.81 mm SL, CHLOL 824
Family Myctophidae Gill, 1893
Genus Benthosema Goode & Bean, 1896
Remarks Benthosema otoliths typically exhibit certain characteristics, including a robust antirostrum and a specific arrangement of ventral denticles. This arrangement consists of 1–3 anterior denticles separated from a single, strong posterior denticle by a smooth gap along the ventral margin (Schwarzhans & Ohe, 2019; Schwarzhans, 2019a). However, we found that the presence of a consistent smooth gap may vary within species such as Benthosema pterotum (Alcock, 1890). These intra-specific variations are mainly attributed to ontogeny (Fig. 7).

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–g Benthosema parafibulatum sp. nov., ASIZF 0100606–612, a holotype, Site 5, mud layer, b–g paratypes, b, e–g Site 5, mud layer, c–d Site 5, sand layer. h–k Benthosema duanformis sp. nov., ASIZF 0100613–616, h holotype, Site 5, mud layer, i–k paratypes, i, k Site 5, mud layer, j Site 2. l–n Benthosema pterotum (Alcock, 1890), Site 5, mud layer, ASIZF 0100617–619. 1, ventral view; 2, inner view

Extant Benthosema otoliths from Taiwan. Scale bars = 1 mm. Images are inner views. a–c, Benthosema fibulatum (Gilbert & Cramer, 1897), a 14 Mar. 2023, 61.75 mm SL, CHLOL 22578, b 31 May 2022, 68.16 mm SL, CHLOL 22364, c 18 Feb. 2021, 57.15 mm SL, CHLOL 9210. d–h Benthosema pterotum (Alcock, 1890), d 30 Jun. 2022, 52.06 mm SL, CHLOL 18836, e 30 Jun. 2022, 36.88 mm SL, CHLOL 18840, f 30 Jun. 2022, 37.34 mm SL, CHLOL 18832, g 30 Jun. 2022, 42.91 mm SL, CHLOL 18854, h 29 Dec. 2021, 29.88 mm SL, CHLOL 13838
Three species of Benthosema, B. fibulatum, B. pterotum, and B. suborbitale (Gilbert, 1913), are known in Taiwanese waters today (Shao, 2023). Among them, the otoliths of B. suborbitale exhibit distinct characteristics, including a salient depression on the postero-dorsal rim and two consistently spaced ventral denticles separated by a smooth gap (Schwarzhans, 2019a: Fig. 49–4). The otoliths of B. fibulatum feature a more pronounced posterior rim that extends further backward and a gently curved ventral rim (Fig. 7a–c). The otoliths of B. pterotum are characterized by equal dorsal and ventral areas with both rostrum and antirostrum similar in size (Fig. 7d–h). Juvenile otoliths of B. pterotum are more rounded (Fig. 7h), while in adults, the otoliths are semi-rectangular and elongated (Fig. 7d). The presence of a smooth gap on the ventral margin may become more evident in larger specimens.
Benthosema duanformis sp. nov. Lin
Figure 6h–k
?2019 Benthosema aff. fibulatum—van Hinsbergh & Helwerda: pl. 3, Fig. 32, 34 [Philippines].
2019 Benthosema aff. fibulatum—van Hinsbergh & Helwerda: pl. 3, Fig. 33 [Philippines].
Holotype ASIZF 0100613 (Fig. 6h), Site 5, mud layer, Tianliao (Kaohsiung), Taiwan. Early Pliocene.
Paratypes Three specimens: two specimens ASIZF 0100614, 0100616 (Fig. 6i, k), Site 5, mud layer, Tianliao (Kaohsiung), Taiwan; one specimen ASIZF 0100615 (Fig. 6j), Site 2, Chochen (Tainan), Taiwan. Early Pliocene.
Etymology From duan (Chinese) = short, referring to the compact outline of the otolith.
Diagnosis OL/OH = 1.11–1.16, OL/OT = 4.33–4.44, OsL/CaL = 1.67–1.42. Rounded otoliths with vertical posterior rim and truncated posterior part of the otolith. Outline very compact. Two strong ventral denticles separated by a smooth gap.
Description The otoliths are generally characterized by their large, regularly rounded shape, with a vertical posterior rim that results in a truncated appearance of the otoliths. The dorsal rim is gently curved to flat, while the ventral rim is curved. There is a prominent antero-ventral denticle located just before the midline of the otolith, and a much stronger posterior denticle positioned at the postero-ventral corner. These two denticles are separated by a smooth gap, and there may be some minute denticles between the rostrum and the antero-ventral denticle (Fig. 6h, j). The antirostrum is slightly smaller than the rostrum, and both are pointed. An angle is present at the postero-dorsal corner. The sulcus is well-divided, very wide, and centrally positioned on the inner face of the otolith. The ostium is slightly greater in height and longer than the cauda. The dorsal and ventral areas of the otolith are similar in size.
Remarks This newly described species is characterized by having otoliths with the lowest aspect ratio (OL/OH = 1.11–1.16) compared to other closely related species in the region (B. fibulatum: 1.28–1.33; B. pterotum: 1.21–1.26; B. parafibulatum sp. nov.: 1.15–1.26, see below). However, it is likely that the species can only be reliably identified in adult individuals, as the otoliths of juveniles (typically OL < 2 mm) may be indistinguishable from those of other closely related species. The otoliths of B. duanformis are similarly compressed to those of Benthosema minutum Schwarzhans, 2019 from the Early Miocene of New Zealand (Schwarzhans, 2019a), but B. minutum otoliths are typically small-sized, have nearly equally long ostium and cauda (vs OsL/CaL = 1.67–1.42) and are thicker.
Occurrence Taiwan: Lower Pliocene Gutingkeng Formation in Chochen (Tainan) and Tianliao (Kaohsiung), southwest Taiwan. Philippines: Upper Pliocene–Lower Pleistocene Santa Cruz Formation in Cabarruyan, northwest Philippines.
Benthosema parafibulatum sp. nov. Lin
Figure 6a–g
?2019 Benthosema pterotum—Schwarzhans & Ohe: Fig. 5D, E, non Fig. 5F–J [Japan].
?2019 Benthosema aff. fibulatum—van Hinsbergh & Helwerda: pl. 3, Fig. 28, 29 [Philippines].
2019 Benthosema aff. fibulatum—van Hinsbergh & Helwerda: pl. 3, Fig. 31 [Philippines].
Holotype ASIZF 0100606 (Fig. 6a), Site 5, mud layer, Tianliao (Kaohsiung), Taiwan. Early Pliocene.
Paratypes Six specimens: four specimens ASIZF 0100607, 0100610–0100612 (Fig. 6b, e–g), Site 5, mud layer, Tianliao (Kaohsiung), Taiwan; two specimens ASIZF 0100608, 0100609 (Fig. 6c–d), Site 5, sand layer, Tianliao (Kaohsiung), Taiwan. Early Pliocene.
Etymology The species is named after its close relationship (based on otolith morphology) to the extant species Benthosema fibulatum.
Diagnosis OL/OH = 1.15–1.26, OL/OT = 5.22–5.75, OsL/CaL = 1.00–1.29. Oval otoliths with curved and extended posterior rim. Two strong ventral denticles separated by a smooth gap.
Description The otoliths are oval-shaped with curved dorsal and ventral rims. The posterior rim is also curved and extends backward. There is a noticeable antero-ventral denticle positioned just before the midline of the otolith, and a stronger, sometimes protruding posterior denticle located at the postero-ventral corner. These two denticles are often separated by a smooth gap, and there may be 2 or 3 min denticles between the rostrum and the antero-ventral denticle. The rostrum is broader than the antirostrum, and both are pointed. An angle is present at the postero-dorsal corner. The sulcus is well-divided and wide in height, while the ostium is longer than the cauda. The dorsal area is similar in size to the ventral one.
Remarks Benthosema parafibulatum exhibits otoliths that closely resemble those of modern species B. fibulatum (Fig. 7a–c, but see Schwarzhans, 2019a: Fig. 49–1 for an image of a more compact B. fibulatum otolith from Hawaii). The main differences lie in the lower aspect ratio (1.15–1.26 in B. parafibulatum vs 1.28–1.33 in B. fibulatum) and less extended posterior rim observed in the fossil species. The otoliths of B. parafibulatum also differ from those of B. pterotum in having a curved dorsal rim and a more consistent presence of a smooth gap between the two major ventral denticles. A Pliocene otolith previously described as B. aff. fibulatum from the Philippines by van Hinsbergh and Helwerda (2019) can reasonably be assigned to this newly identified species, although another undescribed species with larger dorsal area may also be involved (van Hinsbergh & Helwerda, 2019: pl. 3, Fig. 28, 29).
Occurrence Taiwan: Lower Pliocene Gutingkeng Formation in Chochen (Tainan) and Tianliao (Kaohsiung), southwest Taiwan. Philippines: Upper Pliocene–Lower Pleistocene Santa Cruz Formation in Cabarruyan, northwest Philippines.
Benthosema pterotum (Alcock, 1890)
Figure 6l–n
?1981 Diogenichthys sp.—Ohe: pl. 2, Fig. 13 [Japan].
2019 Benthosema pterotum—Schwarzhans & Ohe: Fig. 5F–J, non Fig. 5D [Japan].
2021 Benthosema pterotum—Lin et al.: Fig. 4g [Taiwan].
Remarks See remarks under the genus Benthosema.
Benthosema suborbitale (Gilbert, 1913)
Figure 8a, b

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–b Benthosema suborbitale (Gilbert, 1913), ASIZF 0100620–621, a Site 5, mud layer, b Site 2. c, Bolinichthys supralateralis (Parr, 1928), Site 5, mud layer, ASIZF 0100622. d–e Ceratoscopelus warmingii (Lütken, 1892), Site 5, sand layer, ASIZF 0100623–624. f Diaphus effulgens (Goode & Bean, 1896), Site 2, ASIZF 0100625. g–h Diaphus chrysorhynchus Gilbert & Cramer, 1897, ASIZF 0100626–627, g Site 6, h Site 5, sand layer
2021 Benthosema suborbitale—Mitsui et al.: Fig. 5B.
Remarks See remarks under the genus Benthosema.
Genus Bolinichthys Paxton, 1972
Bolinichthys supralateralis (Parr, 1928)
Figure 8c
Remarks Four otoliths characterized by oblique anterior and posterior rims and a straight dorsal rim exhibit the closest resemblance to those of Bolinichthys supralateralis (Smale et al., 1995: pl. 17, fig. B1–3). This represents the first recorded fossil occurrence of the genus in the region.
Genus Ceratoscopelus Günther, 1864
Ceratoscopelus warmingii (Lütken, 1892)
Figure 8d, e
2019 Ceratoscopelus warmingii—van Hinsbergh & Helwerda: pl. 4, Fig. 39, 40 [Philippines].
2021 Ceratoscopelus warmingii—Mitsui et al.: Fig. 5C [Japan].
2023 Ceratoscopelus warmingii—Mitsui et al.: Fig. 5A [Japan].
Remarks The otoliths of Ceratoscopelus can be readily distinguished from all other myctophid genera by their large and protruding rostrum, the presence of a tiny or nearly absent antirostrum, and a curved posterior rim (Smale et al., 1995: pl. 17, fig. C1–2; Rivaton & Bourret, 1999: pl. 111, Fig. 1–10). However, the otoliths of C. warmingii are hardly distinguished from those of Ceratoscopelus townsendi (Eigenmann & Eigenmann 1889). The latter species appear to have a more pronounced postero-dorsal angle (see Schwarzhans & Ohe, 2019 for further discussion of the otoliths of the two species).
Genus Diaphus Eigenmann & Eigenmann, 1890
Remarks Diaphus otoliths represent the second most abundant myctophid in the collection. While most of the otoliths belong to juvenile individuals and were not further identified, at least two taxa have been recognized: Diaphus chrysorhynchus Gilbert & Cramer, 1897 and Diaphus effulgens (Goode & Bean, 1896) (see below).
Diaphus chrysorhynchus Gilbert & Cramer, 1897
Figure 8g–h
Remarks These otoliths show many similarities to those of the recent species Diaphus chrysorhynchus, Diaphus garmani Gilbert, 1906 and Diaphus regani Tåning, 1932 (see Schwarzhans, 2013: pl. 11, Fig. 1–5, pl. 10, Fig. 7–11, and 12–16, respectively), as well as the Miocene species Diaphus hataii Ohe & Araki, 1973 (see Schwarzhans et al., 2022: Fig. 9J–AC). These similarities include a marked elevation in the anterior part of the dorsal rim and a wide in height sulcus. However, our fossil otoliths possess a more elongated shape with a more extended posterior rim and are most similar to those of D. chrysorhynchus.
Diaphus effulgens (Goode & Bean, 1896)
Figure 8f
Remarks Among the otoliths, a single large and tall specimen closely resembles those of D. effulgens (Schwarzhans, 2013: pl. 13, Fig. 1–7). It has a curved ventral rim and a relatively flat dorsal rim with a large dorsal area and a somewhat short posterior rim. Although the species is not recorded in the Fish Database of Taiwan (Shao, 2023), it has been reported to have a distribution ranging from 0° and 29°N in the West Pacific (Froese & Pauly, 2023).
Genus Diogenichthys Bolin, 1939
Diogenichthys sp.
Figure 9a

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a Diogenichthys sp., Site 5, sand layer, ASIZF 0100628. b–c Myctophum sp., Site 5, sand layer, ASIZF 0100629–630. d Notoscopelus resplendens (Richardson, 1845), Site 5, sand layer, ASIZF 0100631. e Lampanyctus alatus Goode & Bean, 1896, Site 5, sand layer, ASIZF 0100632. f–h Coelorinchus sp., ASIZF 0100633–635, f–g Site 2, h Site 5, sand layer. 1, ventral view; 2, inner view
Remarks A single specimen with a high-bodied outline (OL/OH = 1.03) and curved dorsal and ventral rims is assigned to the genus Diogenichthys. Currently, three Diogenichthys extant species are known (see Schwarzhans & Aguilera, 2013: pl. 1, Fig. 19–21 and pl. 2, Fig. 1–2), but the distinctions among these species are subtle.
Genus Lampanyctus Bonaparte, 1840
Lampanyctus alatus Goode & Bean, 1896
Figure 9e
2019 Lampanyctus alatus—Schwarzhans & Ohe: Fig. 7L–N [Japan].
2019 Lampanyctus alatus—van Hinsbergh & Helwerda: pl. 6, Fig. 52–55.
Remarks The otoliths are rounded and semi-square in shape with equal dorsal and ventral areas. Among our specimens, their outline closely resembles the otoliths of Lampanyctus alatus as depicted by Smale et al., (1995: pl. 22, fig. D) and Rivaton and Bourret (1999: pl. 120, Fig. 1–7).
Genus Myctophum Rafinesque, 1810
Myctophum sp.
Figure 9b–c
Remarks Myctophum otoliths are only represented by two specimens in our collection, and both exhibit very general features that are characteristic of the genus only (see Schwarzhans & Aguilera, 2013: pl. 3, Fig. 9–21 for reference otoliths). Further specimens are required to conduct a more detailed taxonomic characterization.
Genus Notoscopelus Günther, 1864
Notoscopelus resplendens (Richardson, 1845)
Figure 9d
2021 Notoscopelus resplendens—Mitsui et al.: Fig. 5O [Japan].
Remarks The otoliths of Notoscopelus are typically characterized by their highly elongated shape, with OL/OH ratio often reaching 1.8 and higher (Schwarzhans & Ohe, 2019: Fig. 13K–U). However, among the congeners, the otoliths of Notoscopelus resplendens appear to be more compact. They have OL/OH ratios ranging from 1.30 to 1.59 as documented by Smale et al., (1995: pl. 25, fig. F1–2) and Rivaton and Bourret (1999: pl. 123, Fig. 3–8). Our single specimen falls within this range with an OL/OH ratio of 1.32.
Order Gadiformes Goodrich, 1909
Family Macrouridae Bonaparte, 1831
Genus Coelorinchus Giorna, 1809
Coelorinchus sp.
Figure 9f–h
Remarks The small to medium-sized otoliths of Coelorinchus display similar outline to those of Coelorinchus formosanus Okamura, 1963, characterized by having a notable elevation in the anterior part of the dorsal rim (Fig. 10a–e). However, it is important to note that other closely related Coelorinchus species exist. Therefore, when it comes to species identification based on juvenile or subadult individuals, we adopt a conservative approach.

Extant Coelorinchus and Hymenocephalus otoliths from Taiwan. Scale bars = 1 mm. Images are inner views. a–e, Coelorinchus formosanus Okamura, 1963, a 26 Oct. 2015, 119.00 mm PAL, CHLOL 366, b 29 Dec. 2020, 125.00 mm PAL, CHLOL 1827, c 21 Jan. 2022, 103.95 mm PAL, CHLOL 14683, d 21 Jan. 2022, 76.16 mm PAL, CHLOL 14687, e 21 Jan. 2022, 56.02 mm PAL, CHLOL 14662. f–g Hymenocephalus striatissimus Jordan & Gilbert, 1904, f 18 Feb. 2021, 34.93 mm PAL, CHLOL 2388, g 26 Apr. 2022, 36.99 mm PAL, CHLOL 17424. h–j Hymenocephalus lethonemus Jordan & Gilbert, 1904, h 25 Mar. 2022, 49.12 mm PAL, CHLOL 17108, i 26 Apr. 2022, 24.64 mm PAL, CHLOL 17423, j, 26 Apr. 2022, 35.21 mm PAL, CHLOL 17204. k–l Hymenocephalus longibarbis (Günther 1887), k 12 May 2022, 42.66 mm PAL, CHLOL 17563, l 25 Mar. 2022, 41.28 mm PAL, CHLOL 17663
Genus Hymenocephalus Giglioli, 1884
Hymenocephalus aff. papyraceus Jordan & Gilbert, 1904
Figure 11h–i

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a Hymenocephalus striatissimus Jordan & Gilbert, 1904, Site 5, sand layer, ASIZF 0100636. b–d Bregmaceros japonicus Tanaka, 1908, Site 5, sand layer, ASIZF 0100637–639. e–f Onuxodon sp., Site 2, ASIZF 0100640–641. g Encheliophis sp., Site 2, ASIZF 0100642. h–i Hymenocephalus aff. papyraceus Jordan & Gilbert, 1904, Site 5, sand layer, ASIZF 0100643–644. j–k Bregmaceros nectabanus Whitley, 1941, Site 5, mud layer, ASIZF 0100645–646. 1, ventral view; 2, inner view
1983 Hymenocephalus striatissimus—Ohe: pl. 13, Fig. 8 [Japan].
Remarks The twelve specimens of Hymenocephalus aff. papyraceus exhibit distinct features such as a salient antero-dorsal lobe, rounded ventral and posterior rims, well-separated but closely aligned collicula, and a well-delineated collicular crest located in the middle of the otolith and just above the crista inferior. These characteristics bear a strong resemblance to those of H. papyraceus (Schwarzhans, 2014: Fig. 14I), a species currently known only from the southeastern coast of Japan (Nakayama, 2020; Schwarzhans, 2014). The presence of these otoliths suggests a possible southward extension of the species during the Early Pliocene. However, we note that our specimens represent younger individuals and display a much shorter, less pointed posterior rim and a less oblique ventral area compared to the otolith depicted by Schwarzhans (2014: Fig. 14I), which belongs to a larger individual. Extant otoliths of other species of Hymenocephalus found in Taiwan are presented in Fig. 10f–l for comparison.
Hymenocephalus striatissimus Jordan & Gilbert, 1904
Figure 11a
Remarks The otolith of Hymenocephalus striatissimus, represented by a single specimen, features a high-bodied shape with a fused colliculum and a prominent antero-dorsal lobe that extends to approximately one third of the otolith’s height (OH). These characteristics align well with the otoliths of the extant H. striatissimus (Schwarzhans, 2014: Fig. 32; Fig. 10f–g). Other species within the genus Hymenocephalus also possess very similar otoliths, such as H. megalops Iwamoto & Merrett, 1997 and H. torvus Smith & Radcliffe, 1912 (see Schwarzhans, 2014 for figures), but these species appear to have considerable geographic separation from the fossil sites. In addition, the otolith images depicted by Ohe as H. striatissimus (1983: pl. 13, Fig. 7, 8) and H. striatissimus hachijoensis (1983: pl. 12, Fig. 9, pl. 13, Fig. 4) are not clear enough for species identification, but they certainly do not belong to H. striatissimus, as the otoliths possess a well-divided sulcus.
Family Bregmacerotidae Gill, 1872
Genus Bregmaceros Thompson, 1840
Remarks Bregmaceros otoliths are relatively common in the collection, although many of them are represented by juveniles and/or poorly preserved specimens. The taxonomy of extant Bregmaceros species in the region has been subject to ongoing revision (Ho et al., 2011; Koeda & Ho, 2019), with new species still being named (Ho et al., 2020). It has been suggested that Bregmaceros mcclellandi Thompson, 1840 may not exist outside of the northern Indian Ocean (Froese & Pauly, 2023) and that previous records of B. mcclellandi in Taiwan is a misidentified species of Bregmaceros pseudolanceolatus Torii, Javonillo & Ozawa, 2004 (Ho et al., 2011). Consequently, the identification of corresponding Bregmaceros otoliths also requires updating. For example, the otoliths of B. mcclellandi in Lin and Chang (2012) contain misidentifications, and those in Rivaton and Bourret (1999) may represent a mixture of several Bregmaceros species. Images of extant Bregmaceros otoliths from Taiwan are provided in Fig. 12 for comparative purposes.

Extant Bregmaceros otoliths from Taiwan. Scale bars = 1 mm. Images are inner views. a–c Bregmaceros japonicus Tanaka, 1908, a 26 Apr. 2022, 86.24 mm SL, CHLOL 16742, b 25 Mar. 2022, 70.69 mm SL, CHLOL 15727, c 26 Jun. 2020, 102.08 mm SL, CHLOL 937. d–g Bregmaceros lanceolatus Shen, 1960, d 21 Jan. 2022, 93.75 mm SL, CHLOL 14858, e 21 Jan. 2022, 84.66 mm SL, CHLOL 14863, f, 21 Jan. 2022, 61.09 mm SL, CHLOL 14889, g 21 Jan. 2022, 48.44 mm SL, CHLOL 14884. h, i Bregmaceros nectabanus Whitley, 1941, h 21 Jan. 2022, 47.35 mm SL, CHLOL 14957, i 24 Oct. 2021, 56.01 mm SL, CHLOL 10071. j–k Bregmaceros pseudolanceolatus Torii, Javonillo & Ozawa 2004, 3 Feb. 2023, 91.53 mm SL, CHLOL 22181
In the present collection, otoliths of Bregmaceros japonicus Tanaka, 1908 and Bregmaceros nectabanus Whitley, 1941 are recorded (see below). Both species are characterized by a robust and wide dorsal lobe, distinguishing them from other Bregmaceros species in the region that possess a narrower dorsal lobe or a pointed, antenna-like dorsal denticle (Fig. 12d–g, j–k). The distinction between B. japonicus and B. nectabanus is subtle, but B. japonicus typically exhibits a broader and more extended posterior lobe positioned just behind the caudal depression, resulting in a stockier otolith outline. In contrast, B. nectabanus displays a less pronounced posterior lobe located above the caudal depression, giving the otolith a slender outline that inclines towards the anterior.
Bregmaceros japonicus Tanaka, 1908
Figure 11b–d
2019 Bregmaceros cf. mcclellandi—van Hinsbergh & Helwerda: pl. 7, Fig. 63 [Philippines].
Remarks See remarks under the genus.
Bregmaceros nectabanus Whitley, 1941
Figure 11j–k
2019 Bregmaceros cf. mcclellandi—van Hinsbergh & Helwerda: pl. 7, figs. 61, 62, 64.
Remarks See remarks under the genus.
Order Ophidiiformes Berg, 1937
Family Carapidae Poey, 1867
Genus Encheliophis Müller, 1842
Encheliophis sp.
Figure 11g
1983 Carapus sp.—Ohe: pl. 9, Fig. 2 [Japan].
Remarks Two triangular, massive otoliths with a fused colliculum, a flat inner face, and a predominantly convex and thickened outer face are confidently classified as belonging to the carapid family. The outline of the otoliths and the proportion of the sulcus to the otolith closely resemble those of the genus Encheliophis (Smale et al., 1999: pl. 33, fig. E), suggesting their affiliation with this genus.
Genus Onuxodon Smith, 1955
Onuxodon sp.
Figure 11e–f
Remarks These otoliths share many characteristics with those of Encheliophis (see above), particularly in terms of the sulcus and overall outline. However, these specimens are generally smaller in size, displaying a much shorter posterior part and a less convex outer face, which aligns more closely with the otoliths of the genus Onuxodon.
Family Ophidiidae Rafinesque, 1810
Genus Neobythites Goode & Bean, 1885
Neobythites sp.
Figure 13a–b

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–b Neobythites sp., Site 2, ASIZF 0100647–648. c, Bythitidae indet., Site 2, ASIZF 0100649. d–j Gutingichthys changi sp. nov., ASIZF 0100650–656, d holotype, Site 2, e–j, paratypes, e, g, h, j Site 2, f, i Site 5, sand layer. k Microdesmidae indet., Site 5, mud layer, ASIZF 0100657. l Jaydia sp., Site 5, mud layer, ASIZF 0100658. m–p Myersina filifer (Valenciennes, 1837), Site 5, sand layer, ASIZF 0100659–662. 1, ventral view; 2, inner view
Remarks Two otoliths belonging to the genus Neobythites are present in the collection. While they bear similarities to Neobythites sivicola (Jordan & Snyder, 1901) (see Lin & Chang, 2012: pl. 81), a common modern species found in the area, the poor preservation of the specimens prevents a definitive identification at the species level.
Family Bythitidae Gill, 1861
Genus Gutingichthys gen. nov. Lin
Type species Gutingichthys changi sp. nov.
Etymology The genus is named after the type horizon, the Gutingkeng Formation.
Diagnosis OL/OH = 1.79–2.75, OL/OT = 4.11–4.55. Elongate, rectangular otoliths with vertical or obtuse posterior and anterior rims and nearly horizontal, straight dorsal and ventral rims. Sulcus filled with a single oblong colliculum, entirely surrounded by elevated cristae forming a broad oval bulge around the sulcus.
Remarks The distinct characteristics of the elongated otoliths, including the vertical posterior and anterior rims, oblong sulcus that is not inclined, and the presence of thick and swollen cristae, are highly characteristic. Such swollen cristae are rarely observed in extant bythitids, with an example being Tuamotuichthys bispinosus Møller et al., 2004 (see Møller et al., 2004 for figures). These otoliths clearly represent a fossil species belonging to an extinct genus (Schwarzhans, pers. comm. 2022).
Composition Gutingichthys changi sp. nov. Lin.
Gutingichthys changi sp. nov. Lin
Figure 13d–j
Holotype ASIZF 0100650 (Fig. 13d), Site 2, Chochen (Tainan), Taiwan. Early Pliocene.
Paratypes Six specimens: four specimens ASIZF 0100651, 0100653, 0100654, 0100656 (Fig. 13e, g–h, j), Site 2, Chochen (Tainan), Taiwan; two specimens ASIZF 0100652, 0100655 (Fig. 13f, i), Site 5, sand layer, Tianliao (Kaohsiung), Taiwan. Early Pliocene.
Etymology The name is derived from the name of Dr. Chih-Wei Chang (National Academy of Marine Research, Ocean Affairs Council, Kaohsiung, Taiwan), who has made significant contributions to the field of ichthyology, ocean science, and fish otolith studies in Taiwan. The name is a tribute to Dr. Chang's expertise and his role as a mentor to the author (CHL).
Diagnosis See diagnosis under the genus.
Description The otoliths of this species exhibit an elongated to rectangular shape. In juveniles, the shape is more rounded and obtuse with an aspect ratio (OL/OH) of approximately 1.8. However, in larger individuals, the otoliths become more elongated with OL/OH ranging from 2.3–2.7. The posterior and anterior rims of the otoliths are variable, with some specimens bearing two angles, as seen in the holotype (Fig. 13d2), while others have nearly straight and almost vertical rims (Fig. 13f2, i2). The ventral and dorsal rims are flat and straight, contributing to the overall rectangular shape. The otoliths are characterized by their thickness. The sulcus of these otoliths is oblong, centrally positioned. It is completely filled with a single, large colliculum, which is entirely surrounded by swelling and highly elevated cristae. This distinctive feature sets them apart from other species.
Remarks See remarks under the genus.
Occurrence Currently only known from the Lower Pliocene Gutingkeng Formation in Chochen (Tainan) and Tianliao (Kaohsiung), southwest Taiwan.
Bythitidae indet
Figure 13c
Remarks This single, moderately thick otolith has a regular oblong shape with tapering ends, an oval sulcus, and similar-sized dorsal and ventral areas. The thickness, outline, and sulcus type are typical characteristics shared by several genera, including Cataetyx and Grammonus. However, the single specimen available is not sufficient to make a definitive identification beyond the level of family. Further specimens and additional diagnostic features would be necessary for a more specific classification.
Order Kurtiformes Jordan, 1923
Family Apogonidae Günther, 1859
Genus Jaydia Smith, 1961
Jaydia sp.
Figure 13l
Remarks A single massive otolith with its typical apogonid-type outline and sulcus is assigned to the genus Jaydia. Its distinctive features include a prominent antero-dorsal lobe, a smaller lobe in the dorsal rim, a truncated posterior rim, and a curved ventral rim. The ostium is wider in height than the oblong cauda (OsL/CaL = 1.5). It closely resembles the Jaydia otoliths depicted in Lin and Chang (2012: pl. 99, as Apogon).
Order Gobiiformes Günther, 1880
Family Gobiidae Cuvier, 1816
Remarks Gobiid otoliths are commonly found from the studied sites and rank as the second most abundant (Table 1). Taiwan is home to a considerable number of extant gobiid species (Shao, 2023), which share a similar otolith morphology. However, the lack of knowledge about the otoliths of extant gobiid species in the region poses a significant challenge and complicates the identification of relatively young fossils, as described in this study. Therefore, it can be challenging to assign otoliths to specific species or even genus. In this study, a conservative approach was taken, and only otoliths with distinct characteristics were identified to lower taxonomic levels, while the remaining otoliths were assigned to the family level only (e.g., 14 h–k).
Genus Myersina Herre, 1934
Myersina filifer (Valenciennes, 1837)
Figure 13m–p
1981 Cryptocentrus cf. C. filifer—Ohe: pl. 8, Fig. 7 [Japan].
Remarks The otoliths of Myersina filifer exhibit distinct characteristics that allow for their easy identification. They are characterized by their tall and high-bodied outline (OL/OH = 0.76–0.86), which sets them apart from other gobiids. Additionally, the otoliths typically feature crenulated dorsal rims, concave anterior and posterior rims, and a centrally located sulcus (Lin & Chang, 2012: pl. 125). The antero-ventral corner of the otoliths shows variation in shape, which can be either rounded or, less frequently, angular.
Genus Priolepis Valenciennes, 1837
Priolepis sp.
Figure 14a–d

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–d Priolepis sp., ASIZF 0100663–666, a, d Site 2, b–c Site 5, sand layer. e–f Parachaeturichthys sp., Site 2, ASIZF 0100667–668. g Suruga aff. fundicola Jordan & Snyder, 1901, Site 2, ASIZF 0100669. h–k Gobiidae indet., ASIZF 0100670–673, h–j Site 5, sand layer, k Site 2. l Carangidae indet., Site 2, ASIZF 0100674. m Exocoetidae indet., 1901, Site 5, sand layer, ASIZF 0100675. 1, ventral view; 2, inner view
Remarks The 16 thickset gobiid otoliths, characterized by their rectangular shape, massive swelling cristae, and oblong sulcus, are tentatively assigned to the genus Priolepis. The identification is based on comparison with similar otoliths depicted by van Hinsbergh and Helwerda (2019: Fig. 91) and several Priolepis otoliths by Schwarzhans et al., (2020: pl. 8, Fig. 2–9). It is important to note that these otoliths demonstrate varied outlines, particularly the variations in the shape of the dorsal rim, and it is likely that our fossil otoliths represent multiple taxa within the genus. Further taxonomic investigation and comparison with additional specimens and reference materials will be necessary to provide a more precise identification and potentially discern the presence of different species within the collected Priolepis otoliths.
Genus Parachaeturichthys Bleeker, 1874
Parachaeturichthys sp.
Figure 14e–f
Remarks The otoliths of Parachaeturichthys are characterized by a large dorsal lobe that extends in the postero-dorsal direction. While similar enlarged lobes can also be observed in the otoliths of Amblychaeturichthys and Chaeturichthys, the size of the specimens in question more closely resembles those of Parachaeturichthys. This similarity can be observed in previous references such as Ohe (1985: p. 132–133), Iizuka and Katayama (2008: p. 145), and Ohe et al., (2020: Fig. 7).
Genus Suruga Jordan & Snyder, 1901
Suruga aff. fundicola Jordan & Snyder, 1901
Figure 14g
?1983 Suruga fundicala—Ohe: pl. 10, Figs. 1, 2, 3 and 4 [Japan].
2019 Suruga fundicola—van Hinsbergh & Helwerda: pl. 8, figs. 84–88 [Philippines].
Remarks This single specimen displays a squarish shape and a deeply incised sulcus, which bears a strong resemblance to those depicted by Ohe (1983: pl. 10, Fig. 1–4) and van Hinsbergh and Helwerda (2019: figs. 84–88), although they did not provide any otolith images of this extant temperate species. It is highly likely that the specimen belongs to the same taxon as those depicted in the references. However, due to the limited availability of only one otolith, identification to the species level was only tentative.
Family Microdesmidae Regan, 1912
Microdesmidae indet.
Figure 13k
Remarks The very small otolith described (< 1 mm), with its tall outline (OL/OH < 0.63) and distinct features, can be confidently assigned to the family Microdesmidae (e.g., Nolf, 2013: pl. 325). The otolith exhibits a very straight and vertical posterior rim, as well as a parallel lower half of the anterior rim. In addition, the otolith displays elevated and thickset cristae.
Order Beloniformes Berg, 1973
Family Exocoetidae Risso, 1827
Exocoetidae indet.
Figure 14m
Remarks The otolith is tentatively assigned to the family Exocoetidae based on its distinctive features. Although the anterior half of the otolith is not preserved, the presence of a thick fringe ventral rim that curves towards the outer face and a wide cauda that is only slightly bent at the tip are characteristic traits observed in otoliths of exocoetids. However, without further details or additional better-preserved specimens, it is not possible to make a more specific identification.
Order Carangiformes Patterson, 1993
Family Carangidae Rafinesque, 1815
Carangidae indet.
Figure 14l
Remarks The otoliths exhibit thinness, a pronounced concavity in the outer face, and typical percomorph sulcus type, and show similarities to otoliths of the family Carangidae. However, due to the limited preservation of the otoliths, it is not possible to assign them to a particular species or genus within the Carangidae.
Order Pleuronectiformes Bleeker, 1859
Family Bothidae Smitt, 1892
Genus Arnoglossus Bleeker, 1862
Arnoglossus sp.
Figure 15a–b

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–b Arnoglossus sp., Site 5, sand layer, ASIZF 0100676–677. c Pardachirus sp., Site 5, mud layer, ASIZF 0100678. d Heteromycteris sp., Site 5, sand layer, ASIZF 0100679. e, Cynoglossus sp., Site 5, mud layer, ASIZF 0100680. f, Parapercis sp., Site 5, mud layer, ASIZF 0100681. g Callionymidae indet., Site 2, ASIZF 0100682. h Champsodon sp., Site 2, ASIZF 0100683. i Trichiuridae indet., Site 5, sand layer, ASIZF 0100684. j Labridae indet., Site 2, ASIZF 0100685. 1, ventral view; 2, inner view
Remarks The otoliths exhibit characteristic features, including a thick and convex structure (biconvex) with both faces of the otolith being convex, especially the outer face. They also have a well-divided sulcus that is deeply incised and surrounded by very thick cristae. These features comply with those of Arnoglossus (Lin & Chang, 2012; Schwarzhans, 1999). It is worth noting that Arnoglossus is the most abundant flatfish genus in the collection. However, it is also important to acknowledge that a more comprehensive reference collection for this genus in Taiwan is still under development.
Family Soleidae Bonaparte, 1833
Genus Heteromycteris Kaup, 1858
Heteromycteris sp.
Figure 15d
Remarks Two oblong otoliths exhibit a vertical posterior rim and an obtuse anterior rim. They are also characterized by a flat dorsal rim, a slightly elevated antero-dorsal part, and an oblique antero-ventral rim. The sulcus is elongated and divided into a straight ostium and a short, squarish cauda. The described otoliths align with the otoliths of Heteromycteris as described by Schwarzhans (1999: figs. 876–880). Although an extant species, Heteromycteris matsubarai Ochiai, 1963, is known from Taiwan according to Shao (2023), but the otoliths of this species are not known.
Genus Pardachirus Günther, 1862
Pardachirus sp.
Figure 15c
Remarks Otoliths of Pardachirus are small (< 1.5 mm). The key features include a slightly enlarged antero-dorsal part, a deep ventral area, and a sulcus that is only slightly divided into an elongate ostium and an oval cauda (Schwarzhans, 1999, figs. 872–875), although this is not clearly seen in our fossil. Given the similarities in morphology and the presence of characteristic features, it is plausible that the otoliths belong to Pardachirus.
Family Cynoglossidae Jordan, 1888
Genus Cynoglossus Hamilton, 1822
Cynoglossus sp.
Figure 15e
Remarks The triangular shape, flat dorsal rim, curved anterior and ventral rims, and vertical posterior rim are typical features observed in Cynoglossus otoliths (Schwarzhans, 1999). Additionally, the narrow and channel-like ostium and the wide, triangular cauda further support the assignment to the genus. However, due to the limited preservation of the specimens, it is not possible to provide a more specific identification at the species level.
Order Callionymiformes sensu Nelson et al., 2016
Family Callionymidae Bonaparte, 1831
Callionymidae indet.
Figure 15g
Remarks The single otolith, despite being small (< 2 mm) and showing some abrasion, exhibits diagnostic characters that align with the family Callionymidae. These include a triangular outline, marked middorsal angle, and the slightly inclined sulcus opening antero-ventrally. Furthermore, the presence of a short posterior rim without an extended posterior part suggests a closer resemblance to the genus Repomucenus (Lin & Chang, 2012: pl. 124).
Order Scombriformes Rafinesque, 1810
Family Trichiuridae Rafinesque, 1810
Trichiuridae indet.
Figure 15i
Remarks The otolith, with its distinct sulcus widening towards the caudal tip and a thick dorsal rim, suggests an affiliation with the family Trichiuridae. Specifically, it shows similarities to the otoliths of the genus Trichiurus as depicted in Lin and Chang (2012: pl. 129). However, due to the limited preservation and the lack of the anterior part of the otolith, further taxonomic assignment is not possible at this time.
Order Trachiniformes sensu Nelson et al., 2016
Family Champsodontidae Jordan & Snyder, 1902
Genus Champsodon Günther, 1867
Champsodon sp.
Figure 15h
Remarks The two otoliths are well-recognized as Champsodon by a pentagonal outline that is pointed at both anterior and posterior ends, a high angled mediodorsal rim, two angles on the ventral rim, a deeply incised and oblong ostium, and a narrow, shallow, and downward oriented, but rather indistinct, cauda (Schwarzhans, 2019c: pl. 2, figs. 1–8). It should be noted that identifying Champsodon species based on otoliths alone can be challenging as the differences between species are often subtle (Schwarzhans, 2019c).
Family Pinguipedidae Günther, 1860
Genus Parapercis Bleeker, 1863
Parapercis sp.
Figure 15f
Remarks Paraperci otoliths are typically triangular with pointed anterior and posterior ends and a marked dorsal angle in the middle of the dorsal rim. The well-delineated sulcus is characterized by a short widening ostium that opens anteriorly and is accompanied by a short rostrum. The long cauda is slightly bent downward and widened posteriorly. Additionally, a deep and oval dorsal depression and an upward curved ventral furrow are observed (Lin & Chang, 2012: pl. 122; Schwarzhans, 2019c: pl. 1, figs. 1–10). While these otoliths are easily recognized as Parapercis, there can be overlapping inter- and intra-specific variations within this species-rich genus. Thus, no species determination was possible.
Order Labriformes Kaufman & Liem, 1982
Family Labridae Cuvier, 1816
Labridae indet.
Figure 15j
Remarks A small and triangular otolith displays characteristics consistent with the family Labridae. It has a distinctly elevated collum that separates the ostium and cauda, both of which are deeply incised. This sulcus type is typical of Labridae. Lin and Chang (2012) have provided otolith images of various genera and species of Labridae from Taiwan; however, it is important to note that the specimen appears to be from a juvenile individual and lacks specific features necessary for identification at the generic or specific level.
Order Acropomatiformes sensu Davis et al., 2016
Family Acropomatidae Gill, 1893
Genus Acropoma Temminck & Schlegel, 1843
Acropoma hanedai Matsubara, 1953
Figure 16a–b

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a–b Acropoma hanedai Matsubara, 1953, Site 2, ASIZF 0100686–687. c–d Acropoma japonicum Günther, 1859, Site 5, sand layer, ASIZF 0100688–689. e–g Malakichthys sp., Site 2, ASIZF 0100690–692. h Synagrops japonicus (Döderlein, 1883), Site 5, sand layer, ASIZF 0100693. i Serranidae indet., Site 5, sand layer, ASIZF 0100694. j Upeneus sp., Site 2, ASIZF 0100695. 1, ventral view; 2, inner view
Remarks Acropoma otoliths are not uncommon and quite recognizable by their notable postero-dorsal and mediodorsal angles, nearly straight sulcus, and large ventral area. The sulcus follows a typical percomorph configuration with well-developed cristae, an oval and short ostium that is mostly filled with a colliculum, and an elongate cauda that bends only slightly ventrally at the tip. Two species, Acropoma hanedai Matsubara, 1953 and Acropoma japonicum Günther, 1859, are known from Taiwan (Shao, 2023). The otoliths of these two species can be distinguished based on the shape of the posterior rim in larger specimens. In A. japonicum, the posterior rim is gently curved and exhibits a less pronounced postero-dorsal angle, whereas A. hanedai has a straighter and more vertical posterior rim with a more salient angle, resulting in a more compact otolith outline appearance (Fig. 17 for comparative figures). Both species have been found in our Gutingkeng collection, but A. hanedai is much rarer compared to A. japonicum.

Extant Acropoma otoliths from Taiwan. Scale bars = 1 mm. Images are inner views. a–d Acropoma hanedai Matsubara, 1953, a 29 Aug. 2021, 80.65 mm SL, CHLOL 8683, b 24 Jan. 2021, 106.97 mm SL, CHLOL 2533, c, 19 Feb. 2020, 29.48 mm SL, CHLOL 908, d 29 Jul. 2021, 68.94 mm SL, CHLOL 8299. e–h Acropoma japonicum Günther, 1859, e, 21 Jan. 2022, 103.40 mm SL, CHLOL 14810, f 21 Jan. 2022, 146.07 mm SL, CHLOL 14808, g 29 Jul. 2021, 65.95 mm SL, CHLOL 8296, h 21 Jan. 2022, 42.00 mm SL, CHLOL 14800
Acropoma japonicum Günther, 1859
Figure 16c–d
2018 Acropoma japonicum—Lin et al.: pl. 2, Figs. 5, 6 [Taiwan].
?2019 Sparidae indeterminate—van Hinsbergh & Helwerda: pl. 9, Fig. 93 [Philippines].
Remarks See remarks under Acropoma hanedai.
Family Malakichthyidae Jordan & Richardson, 1910
Genus Malakichthys Döderlein, 1883
Malakichthys sp.
Figure 16e–g
2021 Malakichthys sp.—Lin et al.: Fig. 5b–d [Taiwan].
Remarks The otoliths of Malakichthys are characterized by their massive and thickset structure as well as their high-bodied and pentagonal outline. They share some similarities of the sulcus with the otoliths of Acropoma, particularly in juveniles (see above). However, one distinguishing feature of Malakichthys otoliths is that the ostium appears to bend upwards more strongly. In Taiwan, there are four known species of Malakichthys (Shao, 2023), but their otoliths exhibit subtle or no distinct differences that can be used for species-level identification (Lin & Chang, 2012: pl.319). Therefore, it is challenging to assign these otoliths to a specific species within the genus based on their morphology alone.
Family Synagropidae Smith, 1961
Genus Synagrops Günther, 1887
Synagrops japonicus (Döderlein, 1883)
Figure 16h
Remarks Otoliths of Synagrops japonicus (Döderlein, 1883) are very distinctive by having a protruding and oval ostium that forms the anterior part of the otolith, and a long and narrow cauda that flexes strongly towards the antero-ventral direction (Lin & Chang, 2012: pl. 94). In the case of our two fossil otoliths, the overall shape and surface are heavily abraded and not well-preserved. However, despite the poor condition, the peculiar ostium characteristic of S. japonicus otoliths is still visible.
Order Perciformes sensu Nelson et al., 2016
Family Mullidae Rafinesque, 1815
Genus Upeneus Cuvier, 1829
Upeneus sp.
Figure 16j
Remarks Two juvenile otoliths are reasonably assigned to Upeneus based on the sulcus. The presence of a triangular ostium, which opens widely towards the anterior, and a cauda with a long straight part and a downward-bent posterior tip forming a deep expansion are indicative of Upeneus otoliths (Lin & Chang, 2012: pl. 111). However, the expansion of the caudal tip (bulb) is difficult to see clearly because of preservation.
Family Serranidae Swainson, 1839
Serranidae indet.
Figure 16i
Remarks A juvenile otolith is provisionally allocated to the family Serranidae. It shares similarities with those of Chelidoperca (Lin & Chang, 2012: pl. 95) and Pseudanthias (Rivaton & Bourret, 1999: pl. 18, Fig. 7–24), but further identification was not attempted.
Order Scorpaeniformes Garman, 1899
Family Bembridae Kaup, 1873
Bembridae indet.
Figure 18a

Fish otoliths from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Scale bars = 1 mm. Images are inner views unless otherwise indicated. a Bembridae indet., Site 5, sand layer, ASIZF 0100696. b Antigonia sp., Site 2, ASIZF 0100697. c Pagrus major (Temminck & Schlegel, 1843), Site 5, sand layer, ASIZF 0100698. d–e Atrobucca nibe (Jordan & Thompson, 1911), ASIZF 0100699–700, d Site 5, sand layer, e Site 5, mud layer. f utricular otolith, Site 5, sand layer, ASIZF 0100701. 1, ventral view; 2, inner view
Remarks A small, triangular otolith resembles otoliths of the family Bembridae. Despite being a juvenile specimen, its overall outline and sulcus resemble those of Bembras or Parabembras, as depicted in Lin and Chang (2012: pl. 92). The otoliths of Bembridae can display ontogenetic variations, particularly the overall outline, making accurate identification based on a single specimen more difficult.
Order Acanthuriformes Berg, 1930 sensu Nelson et al., 2016
Family Sciaenidae Cuvier, 1829
Genus Atrobucca Chu, Lo & Wu, 1963
Atrobucca nibe (Jordan & Thompson, 1911)
Figure 18d–e
1983 Atrobucca cf. A. nibe —Ohe: pl. 8, Fig. 7 [Japan].
2018 Atrobucca sp.—Lin et al.: pl. 2, Fig. 9 [Taiwan].
2021 Atrobucca sp.—Lin et al.: Fig. 5h–j [Taiwan].
Remarks Atrobucca nibe (Jordan & Thompson, 1911), the only species of the genus Atrobucca known in Taiwan (Shao, 2023), is known to have very thick and distinct triangular otoliths with a nearly flat dorsal rim and an angled ventral rim that creates a large ventral area. The sulcus is characterized by a triangular ostium, which is further distinguished by a large postero-ventral lobe that extends backward. The cauda of the sulcus is strongly flexed postero-ventrally and widened at the tip. These features align with the known otolith morphology of A. nibe (Schwarzhans, 1993: figs. 356–357; C.-H. Lin et al., 2022b: Fig. 6g).
Order Spariformes Akazaki, 1962
Family Sparidae Rafinesque, 1818
Genus Pagrus Cuvier, 1816
Pagrus major (Temminck & Schlegel, 1843)
Figure 18c
?2021 Pagrus major—Mitsui et al.: Fig. 9F [Japan].
Remarks Two fusiform otoliths with strong postero-dorsal and postero-ventral angles share similarities with the otoliths of the family Sparidae. The otoliths of 12 species and all genera of Sparidae found in Taiwan are provided in Figs. 19, 20. Although the surface erosion and incomplete preservation of the rostral part make a definitive identification challenging, the general outline, convexity of the inner face, concavity of the outer face, and sulcus characteristics align with those of P. major. The otolith reported by Mitsui et al. (2021) has a somewhat taller outline, but it appears to belong to a juvenile individual.

Extant otoliths of Sparidae from Taiwan. Scale bars = 1 mm. Images are inner views. a–b, Acanthopagrus chinshira Kume & Yoshino, 2008, a 24 Dec. 2020, 232.86 mm SL, CHLOL 1937, b 28 Aug. 2019, 182.27 mm SL, CHLOL 621. c–d Acanthopagrus latus (Houttuyn, 1782), c, 31 Oct. 2019, 226.97 mm SL, CHLOL 674, d 31 Oct. 2019, 233.84 mm SL, CHLOL 688. e–f Acanthopagrus pacificus Iwatsuki, Kume & Yoshino 2010, e 28 Aug. 2019, 300.32 mm SL, CHLOL 625, f 10 Feb. 2020, 230.00 mm SL, CHLOL 737. g–h Acanthopagrus schlegelii (Bleeker, 1854), g 28 Aug. 2019, 156.12 mm SL, CHLOL 622, h 19 Jan. 2020, 173.88 mm SL, CHLOL 718. i–j Acanthopagrus sivicolus Akazaki, 1962, i 31 Oct. 2019, 223.51 mm SL, CHLOL 683, j 31 Oct. 2019, 217.26 mm SL, CHLOL 684. k Acanthopagrus taiwanensis Iwatsuki & Carpenter, 2006, 24 Dec. 2020, 223.56 mm SL, CHLOL 1938. l–n Rhabdosargus sarba (Forsskål, 1775), l 31 Oct. 2019, 203.79 mm SL, CHLOL 697, m 18 Dec. 2020, 144.89 mm SL, CHLOL 1643, n 21 Aug. 2019, 172.10 mm SL, CHLOL 578

Extant otoliths of Sparidae from Taiwan. Scale bars = 1 mm. Images are inner views. a–d Dentex hypselosomus Bleeker, 1854, a 3 Sep. 2021, 133.64 mm SL, CHLOL 12954, b 25 Jul. 2020, 211.22 mm SL, CHLOL 955, c, 15 Aug. 2020, 260.48 mm SL, CHLOL 1190, d 3 Sep. 2021, 89.04 mm SL, CHLOL 12951. e–h Argyrops bleekeri Oshima, 1927, e 4 Apr. 2022, 43.44 mm SL, CHLOL 17282, f 16 Apr. 2022, 54.79 mm SL, CHLOL 17425, g 17 Jun. 2022, 320.41 mm SL, CHLOL 18574, h 3 Jun. 2022, 96.95 mm SL, CHLOL 18528. i–l Evynnis cardinalis (Lacepède, 1802), i 28 Aug. 2019, 130.88 mm SL, CHLOL 643, j 3 Sep. 2021, 100.93 mm SL, CHLOL 12236, k, 3 Sep. 2021, 59.38 mm SL, CHLOL 11976, l 31 Oct. 2019, 162.74 mm SL, CHLOL 695. m–p Evynnis tumifrons (Temminck & Schlegel, 1843), m 25 Jul. 2020, 192.32 mm SL, CHLOL 956, n 3 Sep. 2021, 125.10 mm SL, CHLOL 11388, o 26 Oct. 2021, 84.32 mm SL, CHLOL 10206, p 15 Aug. 2020, 242.99 mm SL, CHLOL 1159. q–s Pagrus major (Temminck & Schlegel, 1843), q 24 Apr. 2021, 260.38 mm SL, CHLOL 5451, r 17 Aug. 2022, 108.88 mm SL, CHLOL 19786, s 24 Apr. 2021, 183.25 mm SL, CHLOL 5443
Order Caproiformes sensu Rosen, 1984
Family Caproidae Bonaparte, 1835
Genus Antigonia Lowe, 1843
Antigonia sp.
Figure 18b
Remarks This small (< 1.5 mm) and tall otolith is characterized by an oblique posterior rim, lobated rim margins, and a deep excisure. The sulcus is well-divided into a deep, short ostium with a salient ventral lobe and a straight, elongate cauda. These combined features comply well with those of Antigonia (Lin & Chang, 2012: pl. 131), but it is worth noting that our identified otolith is from a juvenile specimen.
Utricular otoliths
Figure 18f
Remarks Utricular otoliths, represented by 76 specimens in the Gutingkeng Formation collection, are relatively common. However, it is important to note that these otoliths do not possess significant taxonomic characteristics that can be used for identification. As a result, the utricular otoliths in question were not subjected to an identification attempt.
Results and discussion
Abundance, diversity, and sample coverage
In this study, the extensive collection of 122 bulk sediment samples from the study sites provided a unique opportunity to conduct a comprehensive evaluation of otolith-based fish abundance and diversity. Unfortunately, no fish otoliths were found in the samples from sites 3 and 4. Nevertheless, our biostratigraphic analysis allowed for age determination for both sites, which is valuable for documentation purposes (Material and methods). It is worth noting that the distribution of otoliths in sediments is likely to be uneven (Agiadi et al., 2022), and to capture a more representative faunal assemblage, multiple replicates with appropriate sampling coverage are necessary (Lin & Chien, 2022). While previous studies may not have routinely incorporated diversity assessment and sample coverage analysis, our adoption of this robust sampling strategy has significantly improved the reliability and accuracy of our findings.
The assessment of otolith abundance in the sediments, or otolith density, provided a general view of fossil distribution in the present study (Fig. 21). The abundance and diversity of otoliths were found to be low in sites 1 and 6 (Table 1), and the preservation of otolith specimens was also less favorable compared to sites 2 and 5. Notably, the mud samples exhibited much lower otolith density and diversity compared to the bioclastic sand layers. The sand layers showed over one order of magnitude higher otolith density and two times greater diversity (Fig. 21).

Differences in density of otoliths (a–b) and species richness (c–d) for the sampled sites. Box plots with lower (25th percentile), median, and upper (75th percentile) boundaries, and outliers (solid circle) outside of 10th and 90th percentiles are presented. Note change of scale for samples from the sand layers in Kaohsiung (c–d)
The results from the rarefaction curves revealed interesting patterns in three different diversity indices (Fig. 22). While a total of 64 taxa were observed in all assemblages (Table 1), the rarefaction curve for species richness (0D) indicated a higher diversity of approximately 75 taxa with increased sampling effort. These additional taxa would likely be represented by a wide variety of marginal species with considerable low individual numbers. In contrast, the rarefaction curves for Shannon (1D) and Simpson (2D) diversity indices reached asymptotes suggesting that our sample coverage for abundant and dominant taxa was sufficient. It is likely that there may be up to five abundant taxa and only three dominant taxa present in the entire Lower Gutingkeng Formation. Overall, these findings suggest a fish community with very few highly dominant fish species and numerous minor species during the Early Pliocene in southwestern Taiwan.

Rarefaction curves of otolith diversity (Hill numbers) among three sampling sites and all sites combined in terms of species richness (0D), Shannon diversity (1D), and Simpson (2D) diversity. Shaded areas represent 95% confidence interval for their respective sites based on 300 bootstrap replicates
Taxonomic composition and paleoecological significance
The otolith assemblages from the Lower Pliocene Gutingkeng Formation exhibit a taxonomic composition comprising 8,314 otoliths assigned to 64 different taxa from 33 families (Table 1). Remarkably, it contains one new fossil genus and three new fossil species, highlighting the significance of the otolith collection in expanding our understanding of the fossil fish fauna in Taiwan. In terms of taxonomic diversity, this collection represents the most diverse fossil fish fauna documented from Taiwan so far (Lin et al., 2021).
Rank abundance analysis of the otolith assemblages from the Gutingkeng Formation indicates strong dominance of Benthosema (Myctophidae), Gobiidae, and Polyipnus (Sternoptychidae) (Fig. 23). Among these, the mesopelagic fish family Myctophidae alone constitutes over 85% of the otolith count across all samples. Additionally, there are several marginal taxa with lower individual counts, including Gonostomatidae, Macrouridae, Champsodon (Champsodontidae), and Acropomatiformes, which are characteristic of typical deep-sea inhabitants (Nelson et al., 2016). The presence of Atrobucca nibe (Sciaenidae) also suggests slightly deeper water depths compared to other sciaenids known as shallow-water inhabitants (Shao, 2023). Overall, the habitat preferences of these deep-sea and oceanic fishes align well with the paleoenvironmental interpretation of an outer-shelf to upper slope environment, as supported by previous studies (Keng, 1981; Lin, 1990; Luo, 1988; Oinomikado, 1955).

Rank abundance of otolith taxa from the Lower Pliocene Gutingkeng Formation, southern Taiwan. Numbers of otoliths are indicated in the parenthesis
However, a notable difference in taxonomic composition is observed between samples from the mud and bioclastic sand layers. In the sand layer samples, in addition to outer-shelf fishes, otoliths of shallow-water taxa derived from various depth settings are frequently found (Table 1). These include gobies (Gobiidae), flounders (Bothidae), cardinalfishes (Apogonidae), and seabreams (Sparidae), which are typically associated with shallow-water and shelf environments (Froese & Pauly, 2023; Nelson et al., 2016). The sand layer samples contain a considerable abundance of gobiid otoliths indicating a diverse gobiid fish fauna (Table 1). However, further research integrating otolith morphology with molecular data may be helpful to gain a comprehensive understanding of ancient goby fauna in the area (Gierl et al., 2022). The co-occurrence of various mollusks and even shark teeth in the sand layers is also noteworthy as these are not commonly observed or present in the mud samples (Fig. 3). The sand layers can potentially originate from either turbidity currents or storm events. Distinguishing between episodic storm deposits and turbidite deposits can be challenging, as both processes involve sediment transport from shallower-water to deeper settings and deposition by underwater currents. However, certain characteristics commonly found in turbidite deposits, such as alternating layers of coarser and finer sediments, well-defined graded bedding, and distinctive sedimentary structures like cross-bedding and ripple marks, are not evident within these sand layers. While turbidite deposits typically exhibit relatively thicker beds compared to episodic storm deposits, it's worth noting that the observed sand layers are relatively thin in our case (Fig. 3; Additional file 1: Fig. S2). Based on these observations, it can be inferred that the sand layers likely represent episodes of storm events (distal tempestites) or periods of strong water currents, resulting in the transportation and deposition of sediments from shallower depths.
Establishment of mesopelagic fish fauna over 3 million years in southwestern Taiwan
Based on our biostratigraphic analysis, the bulk sediment samples containing otoliths are dated to 5.6 to 3.1 Ma, indicating an Early Pliocene Zanclean age. While a comprehensive comparison with adjacent otolith-based assemblages is not yet feasible due to data gaps in time and space and inconsistent sampling methods, our findings indicate a higher species diversity compared to the deep-sea Plio–Pleistocene fish otolith assemblage from the Cabarruyan region in northwest Philippines, where 53 taxa have been described (Van Hinsbergh & Helwerda, 2019). In contrast, the well-known Late Pliocene Kakegawa assemblages from central Japan exhibit rich and diverse tropical to subtropical fauna with more than 100 otolith taxa being reported (Ohe, 1977, 1981, 1983). However, aside from the revision of myctophid otoliths by Schwarzhans and Ohe (2019), extensive revision are needed for these reports.
Among the identified species in our collection, the myctophid Benthosema pterotum stands out as the most abundant fish (Table 1), mirroring its status as one of the most abundant species observed in modern mesopelagic fish communities in the area (Wang & Chen, 2001). The annual bycatch from deep-sea trawling fisheries alone is estimated to exceed 3,000 tons for this species in waters off southwestern Taiwan (Chiou et al., 2020). Other common mesopelagic fish taxa found in our fossil collection include Diaphus, Bregmaceros, Polyipnus, and Acropoma (Fig. 23, Table 1), which are also frequently encountered in the discards of mesopelagic trawlers (Koeda & Ho, 2019; Wu et al., 2004).
When excluding the shelf and nearshore elements derived from the bioclastic sand layers, the overall composition of the Gutingkeng fish assemblages at the family level already resembles the modern outer-shelf and upper slope fish fauna in southwestern Taiwan (Koeda & Ho, 2019; Shao et al., 2008, 2012). The dominant mesopelagic taxa and their relative abundance further suggest a close affinity and a potential early establishment and persistence of modern mesopelagic fish fauna in southwestern Taiwan and the northern part of the South China Sea since at least the Early Pliocene. However, it is also important to note that many otoliths described here are labeled under open nomenclature and have not being identified to species. Thus, they may or may not be directly comparable to the modern fish fauna at the genus or species level. Future works focusing on the Late Miocene and Pleistocene portions of the lower and upper Gutingkeng Formation would provide a more comprehensive understanding of the succession of deep-sea fish fauna in the area.
Conclusions
Our study provides valuable insights into the diversity and composition of otolith-based deep-sea fish fauna in the Pliocene West Pacific, specifically in Taiwan. Through the analysis of 122 bulk sediment samples from the Lower Pliocene Gutingkeng Formation, we reconstructed a previously overlooked yet highly diverse fish fauna consisting of 64 taxa, including one new genus and three new species. The dominance of mesopelagic Myctophidae and deeper-water taxa suggest an outer-shelf to upper slope environment, while the co-occurrence of shallow-water elements indicates episodes of storm events. Our findings highlight the early establishment and persistence of the mesopelagic fish community in southwestern Taiwan since the Early Pliocene. This unique deep-sea fish fauna, exclusively documented through fossil otolith data due to preservation conditions, contributes to filling the knowledge gap in the paleoichthyological fauna in the West Pacific. The baseline data provided by our study will facilitate future research comparing coeval assemblages across the entire West Pacific.
Availability of data and materials
All materials and figured specimens have been registered and deposited in public scientific institution in Taiwan (Biodiversity Research Museum, Academia Sinica, Taiwan). All data generated or analyzed during this study are either included in this published article or available from the corresponding author on reasonable request.
Abbreviations
- CaL:
-
Cauda length
- FAD:
-
First appearance datum
- LAD:
-
Last appearance datum
- OH:
-
Otolith height
- OL:
-
Otolith length
- OsL:
-
Ostium length
- OT:
-
Otolith thickness
- PAL:
-
Fish preanal length
- SL:
-
Fish standard length
References
Agiadi, K., Azzarone, M., Hua, Q., Kaufman, D. S., Thivaiou, D., & Albano, P. G. (2022). The taphonomic clock in fish otoliths. Paleobiology, 48(1), 154–170. https://doi.org/10.1017/pab.2021.30
Chao, A., Chiu, C.-H., & Jost, L. (2014). Unifying species diversity, phylogenetic diversity, functional diversity, and related similarity and differentiation measures through Hill numbers. Annual Review of Ecology, Evolution, and Systematics, 45(1), 297–324. https://doi.org/10.1146/annurev-ecolsys-120213-091540
Chao, A., & Jost, L. (2012). Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology, 93(12), 2533–2547. https://doi.org/10.1890/11-1952.1
Chen, W.-S. (2016). Preface. In W.-S. Chen, An introduction to the geology of Taiwan. Geological Society of Taiwan.
Chen, W.-S., & Yu, N.-T. (2016). Geology of the western foothills. In W.-S. Chen (Ed.), An introduction to the geology of Taiwan (pp. 53–90). Geological Society of Taiwan.
Chi, W.-R. (1979). A biostratigraphic study of the late Neogene sediments in the Kaohsiung area based on calcareous nannofossils. Proceedings of the Geological Society of China, 22, 121–144.
Chiou, W.-D., Chen, K.-W., & Hsu, C.-C. (2020). Estimates of periodicity of otolith ring formation and age and growth of Benthosema pterotum in the coastal waters off southwestern Taiwan. Journal of the Fisheries Society of Taiwan, 47(4), 191–204. https://doi.org/10.29822/JFST.202012_47(4).0001
Chou, J.-T. (1971). A preliminary study of the stratigraphy and sedimentation of the mudstone formations in the Tainan area, southern Taiwan. Petroleum Geology of Taiwan, 8, 187–219.
Covey, M. (1984). Lithofacies analysis and basin reconstruction, Plio-Pleistocene western Taiwan foredeep. Petroleum Geology of Taiwan, 20, 53–83.
Davis, M. P., Sparks, J. S., & Smith, W. L. (2016). Repeated and widespread evolution of bioluminescence in marine fishes. PLoS ONE, 11(6), e0155154.
Fricke, R., Eschmeyer, W. N., & Van der Laan, R. (2023, June 30). Eschmeyer’s catalog of fishes: genera, species, references. Retrieved June 30, 2023, from http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp
Froese, R., & Pauly D. (2023, August 12). FishBase. World Wide Web electronic publication. Retrieved August 12, 2023, from http://fishbase.org
Ghedotti, M. J., Gruber, J. N., Barton, R. W., Davis, M. P., & Smith, W. L. (2018). Morphology and evolution of bioluminescent organs in the glowbellies (Percomorpha: Acropomatidae) with comments on the taxonomy and phylogeny of Acropomatiformes. Journal of Morphology, 279(11), 1640–1653. https://doi.org/10.1002/jmor.20894
Gierl, C., Dohrmann, M., Keith, P., Humphreys, W., Esmaeili, H. R., Vukić, J., Šanda, R., & Reichenbacher, B. (2022). An integrative phylogenetic approach for inferring relationships of fossil gobioids (Teleostei: Gobiiformes). PLoS ONE, 17(7), e0271121. https://doi.org/10.1371/journal.pone.0271121
Hill, M. O. (1973). Diversity and evenness: A unifying notation and its consequences. Ecology, 54(2), 427–432. https://doi.org/10.2307/1934352
Ho, H.-C., Choo, J.-Y., & Teng, P.-Y. (2011). Synopsis of codlet fishes (Gadiformes: Bregmacerotidae) in Taiwan. Platax, 2011, 25–40. https://doi.org/10.29926/PLATAX.201112.0004
Ho, H.-C., Endo, H., & Chu, T.-W. (2020). A new species of the codlet genus Bregmaceros from the western Pacific Ocean (Gadiformes: Bregmacerotidae). Zootaxa, 4786(4), 565–573. https://doi.org/10.11646/zootaxa.4786.4.8
Horng, C.-S., & Shea, K.-S. (1994). Study of nannofossil biostratigraphy in the eastern part of the Erhjen-Chi section, southwestern Taiwan. Special Publication of the Centre Geological Survey, 8, 181–204.
Hsieh, T.-C., Ma, K.-H., & Chao, A. (2016). iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods in Ecology and Evolution, 7(12), 1451–1456. https://doi.org/10.1111/2041-210X.12613
Hu, C.-H. (1989). Manual for ten geological routes in central Taiwan—route 7: Geology along the Nanheng Highway—Tsailiao, Peiliao, Chiahsien. In Faculty members of the Department of Earth Sciences, National Taiwan Normal University (Ed.). Field manual of the geology of Taiwan: Vol. 2 (pp. 105–163). Department of Earth Sciences, National Taiwan Normal University; p. 105–163.
Hu, C.-H. (1990). Manual for ten geological routes in southern Taiwan—route 1: Geology of Hengchun, Eluanbi, and Longpan. In Faculty members of the Department of Earth Sciences, National Taiwan Normal University (Ed.), Field manual of the geology of Taiwan: Vol. 3 (pp. 5–72). Department of Earth Sciences, National Taiwan Normal University.
Hu, C.-H., & Tao, S.-J. (1982). A new species of solitary corals and associated fossils from the lower Gutingkeng Formation of Taiwan. Ti-Chih, 4(1), 39–46.
Iizuka, K., & Katayama, S. (2008). Otolith morphology of teleost fishes of Japan. Bulletin of Fisheries Research Agency, 25, 1–222.
Keng, W.-P. (1981). Geology of the Tainan Hills, southern Taiwan. Bulletin of the Central Geological Survey, 1, 1–31.
Koeda, K., & Ho, H.-C. (2019). Fishes of Southern Taiwan. Marine Biology and Aquarium, Pingtung, Taiwan.
Lin, C.-H., Brzobohatý, R., Nolf, D., & Girone, A. (2017). Tortonian teleost otoliths from northern Italy: Taxonomic synthesis and stratigraphic significance. European Journal of Taxonomy, 322, 1–44. https://doi.org/10.5852/ejt.2017.322
Lin, C.-H., & Chang, C.-W. (2012). Otolith atlas of Taiwan fishes. National Museum of Marine Biology and Aquarium.
Lin, C.-H., & Chien, C.-W. (2022). Late Miocene otoliths from northern Taiwan: Insights into the rarely known Neogene coastal fish community of the subtropical northwest Pacific. Historical Biology, 34(2), 361–382. https://doi.org/10.1080/08912963.2021.1916012
Lin, C.-H., Chien, C.-W., Lee, S.-W., & Chang, C.-W. (2021). Fish fossils of Taiwan: A review and prospection. Historical Biology, 33(9), 1362–1372. https://doi.org/10.1080/08912963.2019.1698563
Lin, C.-Y., Lin, C.-H., & Shimada, K. (2022a). A previously overlooked, highly diverse early Pleistocene elasmobranch assemblage from southern Taiwan. PeerJ, 10, e14190. https://doi.org/10.7717/peerj.14190
Lin, C.-H., Ou, H.-Y., Lin, C.-Y., & Chen, H.-M. (2022b). First skeletal fossil record of the red seabream Pagrus major (Sparidae, Perciformes) from the late Pleistocene of subtropical West Pacific, southern Taiwan. Zoological Studies, 61, e10. https://doi.org/10.6620/ZS.2022.61-10
Lin, C.-H., Wang, L.-C., Wang, C.-H., & Chang, C.-W. (2018). Common early Pleistocene fish otoliths from Niubu in Chia-Yi County, southwestern Taiwan. Journal of the National Taiwan Museum, 71(3), 47–68.
Lin, C.-H., Wang, Y.-C., Ribas-Deulofeu, L., Chang, C.-W., & Li, K.-T. (2022c). Changes in marine resource consumption over the past 5000 years in southwestern Taiwan revealed by fish otoliths. Journal of Archaeological Science: Reports, 42, 103400. https://doi.org/10.1016/j.jasrep.2022.103400
Lin, C.-H., Wei, C.-L., Ho, S. L., & Lo, L. (2023). Ocean temperature drove changes in the mesopelagic fish community at the edge of the Pacific Warm Pool over the past 460,000 years. Science Advances, 9(27), eadf0656. https://doi.org/10.1126/sciadv.adf0656
Lin, Z.-Y. (1990). Sedimentology of sandstones in the Gutingkeng formation, southern Taiwan [Unpublished master’s thesis]. National Sun Yat-sen University.
Luo, J.-Y. (1988). Sedimentological study of mudstones in the Gutingkeng formation, southern Taiwan [Unpublished master’s thesis]. National Sun Yat-sen University.
Mitsui, S., Lin, C.-H., Taru, H., & Shibata, K. (2023). Fish otolith record reveals possible tropical-subtropical fish community in temperate Japan during the exceptionally warm Last Interglacial Period. Historical Biology. https://doi.org/10.1080/08912963.2023.2201933
Mitsui, S., Taru, H., Ohe, F., Lin, C.-H., & Strüssmann, C. A. (2021). Fossil fish otoliths from the Chibanian Miyata Formation, Kanagawa Prefecture, Japan, with comments on the paleoenvironment. Geobios, 64, 47–63. https://doi.org/10.1016/j.geobios.2020.11.003
Møller, P. R., Schwarzhans, W., & Nielsen, J. G. (2004). Tuamotuichthys bispinosus, a new genus and species from off Tuamotu Islands, South Pacific Ocean (Ophiidiformes, Bythitidae). Ichthyological Research, 51(2), 146–152.
Nakayama, N. (2020). Grenadiers (Teleostei: Gadiformes: Macrouridae) of Japan and adjacent waters, a taxonomic monograph. Megataxa, 3(1), 1–383.
Nelson, J. S., Grande, T. C., & Wilson, M. V. H. (2016). Fishes of the World. John Wiley & Sons. https://doi.org/10.1002/9781119174844
Nolf, D. (1985). Otolithi Piscium. In H. P. Schultze (Ed.), Handbook of Paleoichthyology (Vol. 10, pp. 1–145). Gustav Fischer Verlag.
Nolf, D. (2013). The diversity of fish otoliths, past and present. Royal Belgian Institute of Natural Sciences.
Ohe, F. (1977). Otoliths from Dainichi sand and Hosoya Tuffaceous beds of Kakegawa Group (Pliocene), Shizuoka Prefecture, central Japan. Kaseki No Tomo, 16, 13–19.
Ohe, F. (1981). Fish-otoliths from the Dainichi sand and the Fosoya Tuffaceous members of the Pliocene Kakegawa Group, Shizuoka Prefecture, Central Japan. Bulletin of the Senior High School Attached to the Aichi University, 8, 125–194.
Ohe, F. (1983). On the otoliths of deep water fishes from Pliocene Hijikata mud formation exposed in the southern Part of Kakegawa Prefecture, central Japan. Bulletin of the Senior High School Attached to the Aichi University, 10, 1–54.
Ohe, F. (1985). Marine fish-otoliths of Japan. The Senior High School Attached to the Aichi University Education.
Ohe, F., Ando, Y., Ugai, H., Hirose, K., & Kawano, S. (2020). An otolith assemblage of the pleistocene ogushi formation, Itsuwa-machi, Amakusa City, Kumamoto Prefecture, Japan. Bulletin of Goshoura Cretaceous Museum, 2, 1–20.
Oinomikado, T. (1955). Chinese Petroleum Company Paleontology Laboratory Report: Vol. 7. Micropaleotological investigation of the Chishan standard section, near Tainan, Taiwan. Chinese Petroleum Company.
Rivaton, J., & Bourret, P. (1999). Les otolithes des poissons de l’Indo-Pacifique. Institut de recherche pour le développement.
Schwarzhans, W. (1993). A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). Verlag F.
Schwarzhans, W. (1999). A comparative morphological treatise of recent and fossil otoliths of the order Pleuronectiformes. Verlag F.
Schwarzhans, W. (2013). A comparative morphological study of the recent otoliths of the genera Diaphus, Idiolychnus and Lobianchia (Myctophidae). Palaeo Ichthyologica, 13, 41–82. https://doi.org/10.13140/2.1.2872.3843
Schwarzhans, W. (2014). Head and otolith morphology of the genera Hymenocephalus, Hymenogadus and Spicomacrurus (Macrouridae), with the description of three new species. Zootaxa, 3888, 1–73.
Schwarzhans, W. (2019a). Reconstruction of the fossil marine bony fish fauna (Teleostei) from the Eocene to Pleistocene of New Zealand by means of otoliths. Memorie Della Società Italiana Di Scienze Naturali e Del Museo Di Storia Naturale Di Milano, 46, 3–326.
Schwarzhans, W. (2019b). A comparative morphological study of recent otoliths of the Congridae, Muraenesocidae, Nettastomatidae and Colocongridae (Anguilliformes). Memorie Della Società Italiana Di Scienze Naturali e Del Museo Di Storia Naturale Di Milano, 46, 327–354.
Schwarzhans, W. (2019c). A comparative morphological study of recent otoliths of the so-called Trachinoidei. Memorie Della Società Italiana Di Scienze Naturali e Del Museo Di Storia Naturale Di Milano, 46, 371–388.
Schwarzhans, W., & Aguilera, O. (2013). Otoliths of the Myctophidae from the Neogene of tropical America. Palaeo Ichthyologica, 13, 83–150.
Schwarzhans, W., Brzobohatý, R., & Radwańska, R. (2020). Goby otoliths from the Badenian (middle Miocene) of the Central Paratethys from the Czech Republic, Slovakia and Poland: A baseline for the evolution of the European Gobiidae (Gobiiformes; Teleostei). Bollettino Della Societa Paleontologica Italiana, 59(2), 125–173. https://doi.org/10.4435/BSPI.2020.10
Schwarzhans, W., & Ohe, F. (2019). Lanternfish otoliths (Teleostei, Myctophidae) from the Pliocene and Pleistocene of Japan. Rivista Italiana Di Paleontologia e Stratigrafia, 125(2), 355–400. https://doi.org/10.13130/2039-4942/11670
Schwarzhans, W., Ohe, F., Tsuchiya, Y., & Ujihara, A. (2022). Lanternfish otoliths (Myctophidae, Teleostei) from the Miocene of Japan. Zitteliana, 96, 103–134. https://doi.org/10.3897/zitteliana.96.83571
Shao, K. T. (2023, July 07). The Fish Database of Taiwan. Biodiversity Research Center, Academia Sinica, Taiwan. Retrieved July 07, 2023, from http://fishdb.sinica.edu.tw
Shao, K.-T., Ho, H.-C., Lin, P.-L., Lee, P.-F., Lee, M.-Y., Tsai, C.-Y., Liao, Y.-C., Lin, Y.-C., Chen, J.-P., & Yeh, H.-M. (2008). A checklist of the fishes of southern Taiwan, northern South China Sea. The Raffles Bulletin of Zoology, 19, 233–271.
Shao, K.-T., Lin, J., Wu, C.-H., Yeh, H.-M., & Cheng, T.-Y. (2012). A dataset from bottom trawl survey around Taiwan. ZooKeys, 198, 103–109.
Shieh, Y.-T., You, C.-F., Shea, K.-S., & Horng, C.-S. (2002). Identification of diagenetic artifacts in foraminiferal shells using carbon and oxygen isotopes. Journal of Asian Earth Sciences, 21(1), 1–5. https://doi.org/10.1016/S1367-9120(01)00083-9
Smale, M. J., Watson, G., & Hecht, T. (1995). Otolith atlas of southern African marine fishes. J.L.B. Smith Institute of Ichthyology.
Smith, W. L., Ghedotti, M. J., Domínguez-Domínguez, O., McMahan, C. D., Espinoza, E., Martin, R. P., Girard, M. G., & Davis, M. P. (2022). Investigations into the ancestry of the Grape-eye Seabass (Hemilutjanus macrophthalmos) reveal novel limits and relationships for the Acropomatiformes (Teleostei: Percomorpha). Neotropical Ichthyology, 20(3), e210160. https://doi.org/10.1590/1982-0224-2021-0160
Torii, K., & Honma, U. (1932). Reports on the geology of the Hsinhua oil field. Tainan. Government General of Taiwan.
van der Laan, R., Eschmeyer, W. N., & Fricke, R. (2014). Family-group names of recent fishes. Zootaxa, 3882(1), 1–230. https://doi.org/10.11646/zootaxa.3882.1.1
van Hinsbergh, V. W. M., & Helwerda, R. A. (2019). Fish otoliths from the Cabarruyan Piacenzian-Gelasian fauna found in the Philippines. Zootaxa, 4563(3), 401–443. https://doi.org/10.11646/zootaxa.4563.3.1
Wang, J.T.-M., & Chen, C.-T. (2001). A review of lanternfishes (Families: Myctophidae and Neoscopelidae) and their distributions around Taiwan and the Tungsha Islands with notes on seventeen new records. Zoological Studies, 40(2), 103–126.
Wu, C.-C., Lin, J.-C., Huang, C.-S., Hsieh, S.-H., & Su, W.-C. (2004). Studies on the composition and catching rate of trash fishes by small trawlers of Tungkang in Taiwan. Journal of Taiwan Fisheries Research, 12(1), 11–23.
Acknowledgements
We thank all the people who have helped in the field: Meng-Chen Ko, Li-You Lin, Li-Li Lin, Li-Man Lin, Junko Kurosaki, and the members of Marine Paleontology Lab, BRCAS (https://otolithlin.biodiv.tw/). Hsin-Wei Liu is thanked for preparing the otolith images. This manuscript has been improved by the revisions of Werner Schwarzhans and an anonymous reviewer.
Funding
This study was supported by the National Science and Technology Council, Taiwan (Grant No. 110-2116-M-001-009, 111-2116-M-001-033, 112-2116-M-001-017-MY3) and Academia Sinica, Taipei, Taiwan to C-HL.
Author information
Authors and Affiliations
Contributions
C-HL conceived and conceptualized the study; C-HL, S-MW, and C-YL collected the samples; C-HL processed the material, prepared and identified otoliths and took images; C-HL and S-MW performed formal analyses, compiled, validated, and visualized the data; C-YL made geological maps; C-WC performed biostratigraphic analyses; C-HL wrote the draft and was a major contributor in writing the manuscript. All authors contributed and approved the final manuscript.
Corresponding author
Additional information
Editorial handling: Daniel Marty.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Field photographs showing the Gutingkeng Formation and sampling sites 1–4 in Chochen, Tainan (see text for further descriptions). Figure S2. Field photographs showing the Gutingkeng Formation and sampling sites 5–6 in Tianliao, Kaohsiung (see text for further descriptions).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lin, CH., Wu, SM., Lin, CY. et al. Early Pliocene otolith assemblages from the outer-shelf environment reveal the establishment of mesopelagic fish fauna over 3 million years ago in southwestern Taiwan. Swiss J Palaeontol 142, 23 (2023). https://doi.org/10.1186/s13358-023-00288-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-023-00288-0