
Abstract
Here we report new data on the Upper Triassic Polzberg Konservat-Lagerstätte in the Northern Calcarous Alps of Lower Austria. We examined new fossil material obtained from bed-by-bed collections of the well-laminated Reingraben Shales. Over 5290 new fossils of various marine taxa were collected within the fossiliferous layers from the Early Carnian (Julian 2 Ib, Austrotrachyceras austriacum Zone). The newly collected assemblage comprises ammonoids (Austrotrachyceras, Paratrachyceras, Carnites, Simonyceras), belemnoids (Phragmoteuthis, Lunzoteuthis), bivalves (Halobia, div. indet taxa), gastropods (caenogastropods/heterobranchs), thylacocephalan arthropods (Austriocaris, Atropicaris), crustaceans (the decapod Platychela and isopods Obtusotelson, Discosalaputium), branchiopods (Euestheria), polychaetes (Palaeoaphrodite sp. and an unidentified eunicid polychaete), acytinopterygians (Saurichthys, Polzbergia, Peltopleurus, Habroichthys), cartilaginous fishes (Acrodus), coelacanth fish (“Coelacanthus”), a lungfish (Tellerodus), and numerous conodont clusters (Mosherella). Bromalites (coprolites and regurgitalites) produced by piscivorous actinopterygians and durophagous fish accompany the Polzberg palaeobiota along with rare plant remains (Voltzia, div. indet plants). Based on new findings the palaeobiota characterises an intermittent colonisation by abundant benthic halobiid bivalves and a predator-dominated (fish, belemnoids) nektic community. The prerequisites for high-quality preservation—calm and oxygen-depleted conditions—prevailed at the sea floor of the Polzberg Konservat-Lagerstätte. Normal marine conditions prevail in the Reifling Basin, occasionally interrupted by freshwater influx. New in situ findings of benthic and nektic taxa highlight the great value of the unique Polzberg palaeobiota and the autochthonous deposition of the inhabitants within the palaeohabitat. The fauna and flora from the Polzberg Konservat-Lagerstätte, deposited during the Carnian Pluvial Episode or Carnian Wet Intermezzo, points to a carbonate platform decline followed by the deposition of laminated deposits in warmer and wetter conditions.
Similar content being viewed by others

Introduction
Much work has been conducted on the Late Triassic palaeobiota from the Lower Carnian Polzberg Konservat-Lagerstätte (Allison, 1988; Seilacher, 1970; see also Forchielli & Pervesler, 2013; Krystyn, 1991). This includes examining the palaeoenvironment and the trophic food web (Lukeneder et al., 2020; Lukeneder & Lukeneder, 2021), the taphonomy, trophic history and taxonomy of ammonoids (Lukeneder & Lukeneder, 2022a; Lukeneder & Lukeneder, 2023), and new findings of unique mineralized belemnoid cartilage (Lukeneder & Lukeneder, 2022b). These publications were based on historical and recent palaeontological material housed in the official Austrian collections and material from private collections.
Fossils from Konservat-Lagerstätten form valuable sources of palaeobiological information. Thus, new excavations with bed-by-bed collecting were done during four field campaigns (40 day sampling) in 2021. The resulting details on the fauna and flora yielded new insights into the Polzberg palaeocommunity of the Northern Calcareous Alps of Lower Austria and corroborated the assumptions made by Lukeneder and Lukeneder (2021). More details are in preparation on distinct taxonomic groups (thylacocephalans, bristle worms, conodonts, bromalites, and phragmoteuthids), the geochemistry, geophysics and lithological aspects. Lukeneder et al. (2020) and Lukeneder and Lukeneder (2021) discussed food chains and dietary habits, as well as trophic interactions between the inhabitants from the Upper Triassic marine ecosystems here. Known for over 140 years, this Konservat-Lagerstätte with its fossil assemblages generate new data based on previously unrecorded taxa and associated lithological data. This biota was deposited during the Carnian Julian 2 Ib (Austrotrachyceras austriacum Zone, A. minor biohorizon), within a period known as the worldwide Upper Triassic Carnian Pluvial Episode (CPE; Lukeneder et al., 2012; Ruffel et al., 2015; Dal Corso et al., 2018; Simms & Ruffel, 2018). The CPE features an enhanced humidification of the Carnian climate, resulting in the deposition of fossiliferous Reingraben Shales at Polzberg. The climate inducing low oxygenation of bottom sediments, that led to the deposition of laminated sediments. Such deposits provide excellent conditions for the preservation of entire and soft body fossils, a prerequisite for Konservat-Lagerstätten. The biota reflects a marine environment, nonetheless with intermittent shallow-water influence indicated by the findings of freshwater inhabitants, such as a single specimen of the lungfish Tellerodus (Lehman, 1975; Lukeneder & Lukeneder, 2021; Martin, 1982; Stur, 1886; Teller, 1891) and the branchiopod Euestheria (Lukeneder & Lukeneder, 2021; Stur, 1874).
In the present paper, we report all known and new taxa of the Polzberg palaeobiota, from invertebrates and vertebrate traces to plants. The study also presents the detailed bed-by-bed biota recently collected from a single outcrop (Polzberg) and reveals new details of the Carnian food webs. New insights into the Late Triassic (lower Carnian) marine ecosystem are presented and the significance of the assemblage is outlined.
Materials and methods
The authors collected approximately 5290 fossils in 2021 at the Polzberggraben ravine (Lunz Nappe, Northern Calcareous Alps) near Polzberg (= Schindelberggraben; or given as Polzberg locality in numerous collections; Lukeneder & Lukeneder, 2021). More specifically, bed-by-bed sampling was possible only in the lower calcareous parts, whereas in the upper argillaceous parts, the sampling interval was 20 cm due to the soft nature of the deposits. This great amount of new fossil data was compared to the relevant historical material in collections (6397 fossil remains) in the Natural History Museum of Vienna (NHMW), the Geological Survey of Austria (GBA; since 2023 GeoSphere Austria), and the Departments of Earth Sciences at the University of Vienna (DES). Excavation campaigns to obtain those fossil collections were organized by the GBA in 1885 and the NHMW in 1909 (Griffith, 1977; Stur, 1886; Teller, 1891).
Geologic setting and lithology
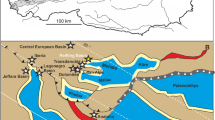
The Upper Triassic outcrops at Polzberg are located on the western slope of Mount Schindelberg (1066 m), north of the Ois River, 4 km northeast of Lunz am See in Lower Austria. Assignment of fossils and samples to the locality Schindelberg is synonymous with the locality Polzberg (= Pölzberg; Lukeneder & Lukeneder, 2021; Stur, 1874, 1886; 1:50 000, geological map, sheet 71 Ybbsitz; Ruttner & Schnabel, 1988, and sheet 72 Mariazell; Bauer & Schnabel, 1997; Fig. 1). The northernmost tectonic elements of the Northern Calcareous Alps (NCA) in Lower Austria are the Frankenfels Nappe, followed to the south by the Lunz Nappe. Within the Lunz Nappe in Lower Austria, the Reifling Basin (Krystyn, 1991; Lukeneder & Lukeneder, 2021)—an intraplatform basin during the Late Triassic—is located between Polzberg and Großreifling. The exact position of the fossiliferous locality in the southern area of the Lunz Nappe within the lower fossiliferous part of the Reingraben Shales (Piller et al., 2004; Tollmann, 1976) was determined by GPS (global positioning system): N 47°53′4.98″ and E 15°4′28.15″, municipality Gaming, federal district Scheibbs.

A Locality map of the Lunz am See area in Lower Austria and the Austrian Northern Calcareous Alps (in grey). B Stratigraphic position of the Polzberg deposits during the Lower Carnian. White asterisk: position of the Konservat-Lagerstätte Polzberg (POLZ). C The Polzberg section with Reingraben Shales. D Fine lamination of the Reingraben Shales at Polzberg. Black asterisk and grey bar: stratigraphic age. Adapted after Lukeneder et al. (2021)
Biostratigraphy of the Polzberg Konservat-Lagerstätte
The Reingraben Shales of Polzberg consist of a single ammonoid assemblage, representing the Lower Carnian Austrotrachyceras austriacum Zone (Julian 2; Fig. 1). The lowermost, fossiliferous part of the Reingraben Shales are dated based on the occurrence of Austrotrachyceras minor, Paratrachyceras haberfellneri and the accompanying Carnites floridus and Simonyceras simonyi as Julian 2 (Julian 2/Ib). The presence of these ammonoid members together with the mass occurrence of the benthic bivalve Halobia rugosa within the fossiliferous layers strengthen the biostratigraphical assignment as Julian age (Lukeneder & Lukeneder, 2021, 2022a, b; Mueller et al., 2016, in press). Concerning the ammonoid zonation and the specific duration of ammonoid zones in the late Carnian (Dal Corso et al., 2020; Mueller et al., 2016; Ogg, 2015a) the analysed section embraces deposits of approximately 500,000 years.
Results
Bed-by-bed collected palaeobiota
The Late Triassic Polzberg Konservat-Lagerstätte, with deposits of black, finely laminated Reingraben Shales, appears in historical collections with 37 fossil marine genera comprising 6,397 specimens from invertebrates to vertebrates. Recent excavations provided an additional 5,290 specimens of various taxonomic groups. Based on lithology and facies, our findings clearly demonstrate that the entire historical material derived from the lowermost three calcareous meters of the Reingraben Shales. No fossils in the historical collections come from the overlying soft argillaceous deposits, resulting in a major sampling bias. We sampled in detail mainly one section 50 m downstream of the historical, collapsed adits. The new section at Polzberg (Po) comprises the basal and fossiliferous 390 cm (Po − 50 cm to Po 340 cm; Fig. 2) high outrcop. The enormous number of fossils (11687; all specimens) and their quality (entire, fragile parts, soft parts) yield special insights into the morphology of such otherwise rarely preserved fossils. The above number is exclusive of the uncountable masses of rock-forming bivalves (mainly Halobia rugosa). The palaeobiota shows a nekton-dominated fauna with abundant fish and cephalopods (Lukeneder et al., 2020).

Detailed Polzberg section with indicated sampling layers (sample numbers Polz − 50 cm to Polz 340 cm) within the lower Carnian Reingraben Shales. Pie charts for compilation of fossil taxa and composition of Polzberg palaeobiota (percentage) throughout the section. Enlarged pie chart at the top represents entire fauna (except halobiids) at Polzberg. Pie chart legend: dark blue Austrotrychyceras minor, grey Paratrachyceras heberfellneri, orange Anaptychus, yellow bromalites, light blue actinopterygians, green Phragmoteuthis bisinuata, ochre gastropods, pink Euestheria. Range and numbers (= n) of the occurring fauna and flora with the taxa detected by bed-by-bed sampling. Diameter of black circles increase with quantity from n = 1 to n > 1,000
Cephalopods dominate the nekton here with 72.2% ammonoids and 4.5% coleoids, followed by 6.0% fish (actinopterygians, coelocanths, conodontophorids), accompanied by less frequent undetermined bivalves (1.0%), gastropods (1.6%), arthropods (thylacocephalans, isopods, platychelids) (3.0%), polychaetes (0.1%), coprolites and regurgitalites (10.1%) as well as allochthonous plant remains (1.3%). The autochthonous macrobenthic community is dominated by countless halobiid bivalves. These small, thin shelled clams dominate throughout the section, both with sporadic appearance and in rock-forming accumulations within distinct layers.
The main faunal elements (Figs. 3, 4) are the benthic bivalves Halobia rugosa, and the ceratitid ammonoids Austrotrachyceras minor (= Trachyceras triadicum var. minor kursiv; Krystyn, 1991; Mojsisovics, 1893a and 1893b with CLXXXVI = 186, Figs. 1–3, p. 682) and Paratrachyceras haberfellneri (= Trachyceras haberfellneri; Mojsisovics, 1893a and 1893b with CLXXXVI = 186, Figs. 6–7, CLXXXVII = 187, Figs. 1, 2, p. 691). The main constituents from the recent collection are (see Table 1 for n of recent and historical collections) trachyceratid ammonoids with Austrotrachyceras and Paratraychceras, Carnites floridus, Simonyceras simonyi, anaptychi and upper jaws of trachyceratids, belemnoids (proostraca, phragmocones, hooks, cartilage), halobiid bivalves and undetermined bivalves, gastropods, arthropods, thylacocephala with Austriocaris and Atropicaris, isopods, euestheriids, polychaetes, bromalites (coprolites and regurgitalites), conodontophorids (clusters), actinopterygian fish, chondrichtyes (questionable), a single lungfish, coelacanths, Coniferopsida with Voltzia and undetermined plant remains. A. minor and P. haberfellneri appear frequently with partially preserved buccal apparatuses of anaptychus-type lower jaws (Trauth, 1935) and upper jaws (Lukeneder & Lukeneder, 2022a). In numerous specimens of Phragmoteuthis bisinuata (Doguzhaeva & Summesberger, 2012; Doguzhaeva et al., 2007b; Lukeneder & Lukeneder, 2022b), the tripartite proostracum and the phragmocones appear with black bituminous sheets of the ink sac, along with black amorphous cartilage and arm hook structures (Lukeneder & Lukeneder, 2021, 2022b). For the questionable belemnoid Lunzoteuthis schindelbergensis from the same locality see Doguzhaeva et al. (2006). Within the abundant ceratitid ammonoids Austrotrachyceras minor and Paratrachyceras haberfellneri, a clear trend from dominating A. minor towards the smaller, less tuberculated P. haberfellneri is evident (Fig. 2). A. minor dominates within the interval from Po -50 cm up to Po 180 cm with values up to 88%. From Po 180 cm (with 17%) on upwards, the ratio of A. minor decreases, and from Po 260 cm on upwards (with 11%), A. minor is gradually replaced by the smoother and smaller P. haberfellneri, with values increasing from 13% to 64% at the top at Po 340 cm (Fig. 2). A. minor has diameters ranging up to 77 mm (mean 16.6 mm) with maximum diameter in layer Po 0–20 cm and the minima in layers Po -50–0 cm and Po 300–320 cm (Lukeneder & Lukeneder, 2022a). The smaller P. haberfellneri exhibits a diameter ranging from 3 to 33 mm (mean 16.9 mm), with a maximum abundance in layer Po − 50–0 cm and a minimum in Po 320–340 cm.

Invertebrate members of the Lower Carnian (Upper Triassic) Polzberg palaeobiota. A Austrotrachyceras minor, positive, lateral view, note change in adult shell ornamentation and black material near aperture representing the in situ Anaptychus, layer Po -50 cm to 0 cm, NHMW 2021/0123/0132a. B Paratrachyceras haberfellneri, positive, lateral view, adult specimens, fatally bitten ammonoid with dislocated shell fragments, layer Po -50 cm to 0 cm, NHMW 2021/0123/0153. C Carnites floridus, lateral view of an adult entire specimen with narrow umbilicus with rounder shoulder, note fine growth lines on body chamber, historical collection, NHMW 2012/0228/0226. D Simonyceras simonyi, ventrolateral view of an adult fragmented specimen with narrow round flank and venter, with original shell, historical collection, NHMW 2012/0228/0360. E Phragmoteuthis bisinuata, proostracum fragment, layer Po -50 cm to 0 cm, NHMW 2021/0123/0195. F Belemnoid cartilage, carbonised fragments, layer Po 60 cm to 80 cm, NHMW 2021/0123/0196. G Arm hooks of P. bisinuata, layer Po 280 cm to 300 cm, NHMW 2021/0123/0197. H Upper jaw of A. minor, positive, lateral view, isolated specimen, layer Po -50 cm to 0 cm, NHMW 2021/0123/0151. I Anaptachus lunzensis, lower jaw of A. minor, positive, lateral view, isolated specimen, layer Po -50 cm to 0 cm, NHMW 2021/0123/0149. J Halobia rugosa, adult specimen, butterfly preservation, layer Po -50 cm to 0 cm, NHMW 2021/0123/0198. K H. rugosa, juvenile specimens, butterfly preservation, layer Po 120 cm to 140 cm, NHMW 2021/0123/0199. L H. rugosa, midaged specimen, accumulated in butterfly preservation, layer Po 120 cm to 140 cm, NHMW 2021/0123/0200. M Bivalvia indet, butterfly preservation, historical collection, NHMW 2012/0228/3612. N Bivalvia indet, butterfly preservation, historical collection, NHMW 2012/0228/3613. O Gastropoda indet, historical collection, layer Po 50 cm to 0 cm, NHMW 2021/0123/0201. P Atropicaris striata, lateral view, layer Po 300 cm to 320 cm, NHMW 2021/0123/0202. Q Austriocaris carinata, lateral view, layer Po 0 cm, NHMW 2021/0123/0203. R Platychela trauthi, lateral view, historical collection, exhibition hall 8 NHM Vienna, NHMW 1910/0015/0018. S Obtusotelson summesbergeri, positive, lateral view, holotype, historical collection, NHMW 2020/0003/0001. T Euestheria minuta, positive, lateral view, layer Po 300 cm, NHMW 2021/0123/0204. U Palaeoaphrodite sp., positive, lateral view, historical collection, NHMW 2018/0103/0003a. V Eunicid bristle worm, positive, lateral view, layer Po 0 cm, NHMW 2021/0123/0056a. Scale bars: 10 mm, except H, I, G, P, T scale bar: 5 mm

Vertebrate members and flora of the Lower Carnian (Upper Triassic) Polzberg palaeobiota. A Nannolepis elegans, lateral view, layer Po 120 cm to 140 cm, NHMW 2021/0123/0205a. B N. elegans, lateral view, layer Po − 50 cm to 0 cm, NHMW 2021/0123/0206a. C Thoracopterus niederristi, lateral view, layer Po − 50 cm to 0 cm, NHMW 2021/0123/0207a. D Skull of Saurichthys calcaratus, note corresponding operculum nearby, lateral view, historical collection, basal part, NHMW 2021/0123/0208a. E Tellerodus sturii, lateral view, cast, historical collection of GBA Vienna, GBA 1891/001/0001. F “Coelacanthus” lunzensis, tail fragment, basal part, historical collection, NHMW 2012/0228/3611. G Coprolite with fish scales, produced by fish, lateral view, layer Po 0 cm, NHMW 2021/0123/0209. H Bromalite mass with A. minor, A. haberfellneri and Anaptychus lunzensis, top view, entire and fragmented shells, layer Po -50 cm to 0 cm, NHMW 2021/0123/0163. I Microcoprolite with accumulated ramiform conodonts of Conodontophorida indet, layer Po -50 cm to 0 cm, NHMW 2021/0123/0210. J Voltzia sp., lateral view, Po -50 cm to 0 cm, NHMW 2021/0123/0001. K Coniferophyta, lateral view, Po -50 cm to 0 cm, NHMW 2021/0123/0033. Scale bars: 10 mm, except I scale bar: 1 mm
Halobia rugosa (1–30 mm length) appears in all ontogenetic stages and, in layer/rock-forming quantities, mostly in double-valved butterfly position. The Reingraben Shales at Polzberg show microfossils, such as ostracods, gastropods and radiolaria in thin sections. The main Polzberg collections are housed at the NHMW and the GBA.
Discussion
Historical summary and legacy of the Carnian Pluvial Episode—the Polzberg Konservat-Lagerstätte
The Carnian Pluvial/Humid Episode (CPE; Dal Corso et al., 2018, 2020; Hornung & Brandner, 2005; Kozur & Bachmann, 2010; Lukeneder et al., 2012; Lukeneder & Lukeneder, 2021; Mueller et al., 2016; Ogg, 2015b; Ruffel et al., 2015; Simms & Ruffell, 1990) was characterized by an abrupt and increased siliciclastic transport towards the north-western branch of the Tethys, resulting in the breakdown of the carbonate factory (Hornung & Brandner, 2005). The ‘Reingraben turning point’ is reflected by changes or adaptations of lithologies, facies, palaeobiota, biofacies, and evolutionary events mirrored in all facies belts of the entire NW Tethyan continental margin (Hornung & Brandner, 2005; Lukeneder & Lukeneder, 2021; Schlager & Schöllnberger, 1974). Global warming and enhanced humidification during the Early Carnian, combined with the deposition of large amounts of volcanogenic material, triggered a worldwide climate change (Dal Corso et al., 2020; Hornung & Brandner, 2005; Hornung et al., 2007a, b; Mueller et al., 2016; Lukeneder & Lukeneder, 2021). Data on palynomorphs (Mueller et al., 2016) support a general trend from a dry climate in the Julian 1 to more humid and warmer conditions during the early Julian 2, corresponding to the deposition of the Göstling Member and the Reingraben Shales (Lukeneder & Lukeneder, 2021). The increasing humidification is reflected by a change in lithology from the basal Julian sequence in the Polzberg area. In the Late Triassic, during the Carnian stage, the Polzberg Subbasin and the Reifling Basin were located at the north-western Tethyan rim in latitudes of 15°N to 30°N (Lukeneder et al., 2012; Lukeneder & Lukeneder, 2021; Mueller et al., 2016). The lowermost nodular limestones of the Reifling Formation were deposited on the palaeoslope of the Reifling Basin. At the base of the Julian 2, the Reifling Formation (Trachyceras aonoides Zone, T. aonoides Subzone) is replaced by the limestone deposits with organic-rich mudstones of the Göstling Member in the Austrotrachyceras austriacum Zone (A. austriacum Subzone, biohorizon of A. triadicum), overlain by deposits of the Reingraben Shales (A. austriacum Subzone, biohorizon of A. minor).
New aspects on the Polzberg palaeobiota revealed by recent findings
Konservat-Lagerstätten characterized by stagnate environments, with their excellent preservation of invertebrate and vertebrate fossils, provide the special conditions required for the formation (Bernardi et al., 2011; Briggs, 2014; Lukeneder & Lukeneder, 2021; Seilacher, 1970). In contrast to fossils housed in historical collections investigated by Lukeneder and Lukeneder (2021), the recent data were gained by bed-by-bed sampling. The historical material derives from two adits for fossil mining in 1885 and 1909. New findings stem from a single additional small excavation site located nearby, approximately 50 m downstream of the abandoned tunnels in the same ravine. The lithology of the historical specimens indicates that the material was derived from the lowermost two meters of the Reingraben Shales, directly located above the Göstling Member. Hence, the recently collected fossil material is correlatable with the historical fossil collections. Benthic bivalves with Halobia rugosa, accompanied by frequent other bivalve taxa (indet) and gastropods, clearly dominate throughout the section. Thylacocephalan arthropods (Austriocaris, Atropicaris), crustaceans (Platychela), branchiopods (Euestheria), isopods (Obtusotelson, Discosalaputium), and polychaetes (Palaeoaphrodite, Eunicidae indet) are benthic members of the marine ecosystem (Fig. 5).

Carnian Polzberg palaeobiota and assumed mode of life and environment. Mode of life based on direct and indirect evidence from the fossil record of the Polzberg Konservat-Lagerstätte. For more details see interpretations in the text. Co-occurring members within the Polzberg assemblage are dominated by the rock-forming thin-shelled benthic bivalves of Halobia rugosa. Nekton is dominated by trachyceratitid ammonites followed by abundant actinopterygian fish and frequent coleoids
Nektic members are dominated by ammonoids with the trachyceratids Austrotrachyceras and Paratrachyceras as well as rare Carnites, ?Sageceras, and Simonyceras. Frequent nektic coleoids (Phragmoteuthis bisinuata) complete the cephalopod community in the lower Carnian Polzberg subbasin. Nektic acytinopterygid fish appear throughout the section with Saurichthys, Polzbergia, Peltopleurus, and Habroichthys. Other taxa include remnants of several coelocanthid fish (“Coelacanthus”), the lungfish Tellerodus, and numerous conodont clusters with Mosherella.
The preservation of benthic (epifaunal and infaunal) along with nektic taxa, and the completeness of the bivalves (still articulated, little fragmentation), gastropods and most of ammonoids points to a autochthonous deposition of the animals or their remnants, with minimal or no subsequent post-mortem drift or transport. Taphonomic evidence, exceptional soft body preservation, in situ buccal masses, completeness of shells, no boturbation) suggests that the Polzberg palaeobiota developed in oxygen-depleted basinal waters (Ekdale & Mason, 1988; Lukeneder et al., 2020; Savrda & Bottjer, 1991; Wignall & Hallam, 1991) during calm conditions in the water column and near the sea floor, without major transport or reorientation of the fossil material.
Comparable sites of Triassic age with similar facies and preservational features are rare but known from Seefeld in Tyrol (Brandner & Poleschinski, 1986) or Wiestal in Salzburg (Hornung et al., 2019), both within the Norian Seefeld Formation, and from the Middle Triassic (late Anisian to Ladinian) Konservat-Lagerstätte of Monte San Giorgio (Ticino, Switzerland; Brinkmann, 1982, 1997; Furrer, 2003; Furrer & Vandelli, 2014).
New findings and well-preserved fossils of the Polzberg Konservat-Lagerstätte mirror the trophic conditions of the palaeo-food web at the time of deposition (Briggs, 2014; Lukeneder & Lukeneder, 2021; Seilacher, 1970; Fig. 6). Due to the oxygen depleted bottom water conditions the shells or carcasses are not or only little affected by benthic scavengers or bacterial decay (see also Beardmore & Furrer, 2016; Furrer 2004; Tintori, 1992). Additional and frequent findings of bitten shell fragments crushed by nektic predators, along with numerous bromalites (coprolites and regurgitalites; Hunt & Lucas, 2012; Hunt et al., 2012), shed light on the palaeobiota here (Lukeneder & Lukeneder, 2021; Lukeneder et al., 2020). Ammonoid shell fragments and entire ammonoids within the bromalite masses are solely from the trachyceratid species A. minor and P. haberfellneri (Lukeneder & Lukeneder, 2022a). Coleoid fragments belong exclusively to the belemnoid Phragmoteuthis bisinuata (Lukeneder & Lukeneder, 2022b). Abundant coprolite masses are dominated either by fish scales, fragmented or entire ammonoid shells, coleoid hooks or carbonized cartilage remains. This suggests that durophaguous fish fed on these cephalopods and fish (Griffith, 1977; Lukeneder & Lukeneder, 2021; Lukeneder et al., 2020). Near or at the sea floor thylacocephalan arthropods (Austriocaris, Atropicaris), crustaceans (the decapod Platychela and isopods, such as Obtusotelson, Discosalaputium) and polychaetes (Palaeoaphrodite sp. and eunicid polychaetes) occurred. On the sediment surface and in the sediment organic material was decomposed by bacteria or fungi (cf. Ramanan et al., 2016).

Hypothesised trophic food web of the Carnian Polzberg palaeobiota based on direct and indirect evidence from the fossil record of the Polzberg Konservat-Lagerstätte. Direct evidence from bromalites, indirect evidence as generalized interpretation of occurring members within the Polzberg assemblage. Figured members of palaeobiota not to scale
A single specimen of the lungfish Tellerodus sturii is known from historical collections (Lukeneder & Lukeneder, 2021; Stur, 1886; Teller, 1891). Mesozoic dipnoans were restricted to freshwater environments, and their remains found in marine deposits are commonly interpreted as a result of post-mortem transport from freshwater ecosystems (Stur, 1886; Teller, 1891). Conchostraca appear frequently with Euestheria in the upper, more argillaceous part of the Polzberg section. Euestheriids typically inhabit freshwater or at least brackish environments (Montagna et al., 2018; Tintori, 1990). Both elements—the lungfish Tellerodus and the conchostracan shells—indicate a sporadic influx of freshwater or sedimentation from surrounding shallow-water or terrestrial areas into the restricted Reifling Basin with the Polzberg Subbasin (Fig. 7). A possible adaptation of these new conchostracan species to marine environments cannot be fully excluded. Plant remains with foliated trunks of the Coniferophyta member Voltzia support the interpretation of sporadic freshwater influx into the fully marine basin.

A Detailed palaeogeography of the Reifling Basin with the Polzberg locality POLZ (asterisk) with the laminated Reingraben Shales during the Upper Triassic, Carnian. B Reconstruction of the palaeoenvironment and members of the Carnian Polzberg palaeobiota. 1 Austrotrachyceras minor, 2 Austrotrachyceras haberfellneri, 3 Phragmoteuthis bisinuata, 4 Halobia rugosa, 5 caenogastropod, 6 Palaeoaphrodite, 7 Antrimpos, 8 Platychela trauthi, 9 conodonts, 10 “Coelacanthus” lunzensis, 11 Polzbergia brochatus, 12 Habroichthys gregarius, 13 Nannolepis elegans, 14 Saurichthys calcaratus, 15 Acrodus, 16 Thoracopterus niederristi, 17 potential Sauropterygia, 18 Tellerodus sturii. Figured members of palaeobiota not to scale. A: Artwork by Mathias Harzhauser, NHMW. B: Artwork based on Fig. 6 by AL, Final artwork by 7reasons; www.7reasons.net; adapted after Lukeneder & Lukeneder, 2021. WCP Wetterstein Carbonate Platform. Hds Hallstatt deep swell. A and B: not to scale
Fatal injuries to ammonoids and phragmoteuthids, along with bromalite contents, document the prevailing predator–prey and other synecological relationships among ammonoids, phragmoteuthids and fish. Bromalites (regurgitalites, coprolites) and ammonoid shell fragment clusters confirm that ammonoid shells were frequently fatally bitten by actinopterygian fish, such as Saurichthys or phragmoteuthids. A high percentage (22.7%) of the detected ammonoid shells are fragmented, with size ranges from three-fourths of the shell down to small pieces measuring only 2–3 mm (Lukeneder & Lukeneder, 2022a). Calm and stagnate environments with oxygen-depleted conditions were hostile to scavengers at the sea floor (Furrer, 2004), creating the prerequisites for excellent preservation of in situ buccal masses (n = 57) with anaptychi (Anaptychus lunzensis) and upper jaws within or close to the body chambers of A. minor and P. haberfellneri. Frequent ammonoids (n = 216) exhibit an elongated area showing a black mass at the posterior end of the body chamber near the final septa, approximately a half whorl distance from the aperture (Lukeneder & Lukeneder, 2022a). The dark-coloured organic remains preserved as a black mass in the posterior body chamber are interpreted as muscle remains in the body chambers of the small trachyceratid genera Austrotrachyceras and Paratrachyceras (Lukeneder & Lukeneder, 2022a).
The palaeoenvironment of the Polzberg Konservat-Lagerstätte
The Reingraben Shales with dark, laminated deposits were formed in a relatively deep marine environment within an intra-platform basin, the Reifling Basin. This interpretation is inferred from the dominance of benthic halobiids and a diverse nektic fauna with abundant ammonoids and fish (Forchielli & Pervesler, 2013; Griffith, 1977; Lukeneder & Lukeneder, 2021; Lukeneder et. al., 2020; Fig. 7). The bottom conditions during the deposition of the Reingraben Shales were dysoxic to anoxic. This conclusion is based on the preservation of abundant entire carcasses of fish with fragile skeletons (Tintori, 1992), masses of double-valved bivalves, in situ buccal masses in ammonoids, the soft-bodied fauna of bristle worms and ammonoid soft parts (carbonisation, phosphatisation), the abundance of organic material in the sediment, the presence of common framboidal pyrite, the absence of sessile organisms, and the lack of bioturbation (Glaessner, 1931, 1965; Lukeneder & Lukeneder, 2021, 2022a, b; Lukeneder et al., 2020). The Polzberg Subbasin within the Reifling Basin was mainly marine with ephemeral and limited freshwater input (Griffith, 1977; Lukeneder & Lukeneder, 2021; 2022a, b). Limited or absent bottom water currents (low energy) on the sea floor and dysaerobic conditions, which prevented predators from separating the ammonoid shells from the jaw apparatuses (cf. Lukeneder & Tanabe, 2002; Lukeneder, 2015; Tintori, 1992), led to the extraordinary preservation of the Polzberg palaeobiota. Exceptional preservation features, such as articulated hard parts, fragile and entire specimens (fish, cf. Tintori, 1992), and soft body preservation are the prerequisite for Konservat-Lagerstätten (Lukeneder & Lukeneder, 2021; Seilacher, 1970). When oxygen concentrations drop below a critical threshold level (0.1 ml/l dissolved oxygen), bioturbation virtually ceases and laminated, organic-rich deposits accumulate as observed in Polzberg with the Reingraben Shales (Lukeneder & Lukeneder, 2021; Savrda & Bottjer, 1991). Based on geochemical data (Lukeneder, A. et al., in prep.), the oxygen availability was highly variable during the deposition of the Carnian sediments of this basin. Except for the transported freshwater lungfish, the frequent branchiopod Euestheria and terrestrial plant material (Voltzia), the autochthonous character of the Polzberg deposits is strengthened by the preservation of fragile parts and the extraordinary preservation of in situ buccal (anaptychi) associations within or close to the body chambers of A. minor and P. haberfellneri. The geochemical results (Lukeneder, A. et al., in prep.), along with the laminated fabric and facies, abundant organic matter and high amounts of sulphur, indicate that the assemblage was deposited under conditions of intermittent oxygen depletion associated with stable water masses. A dynamic environment, controlled by short- and long-term fluctuations in oxygen levels, along with poor circulation of bottom-water currents within an isolated, basin-like region, led to the accumulation of the Austrotrachyceras abundance zone (= ‘Trachyceras Schichten’).
Trophic food web of the Polzberg palaeobiota
The great amount (> > 10,000, historical and recent findings) and diversity of fossil remains (37 taxa) enable detailed conclusions on the palaeo-food web (Figs. 6, 7). The marine fauna is dominated by benthic halobiid bivalves, nektonic actinopterygian fish and ceratitid ammonoids (Lukeneder & Lukeneder, 2021). Pulses of freshwater influx are documented by rare euestheriids (Euestheria) and a single lungfish, completing the picture of a diverse Lower Carnian food web (Fig. 7). Based on new abundant and high-quality fossil material from this Konservat-Lagerstätte the data presented earlier on the food web, food chains and predator–prey relations, between fish and cephalopods in Lukeneder and Lukeneder (2021), are confirmed.
Primary producers are represented by algae and bacteria. Microfilms and microbes were grazed by primary consumers such as gastropods and arthropods from the sea floor and filtered by benthic bivalves. Low-level consumers were preyed upon by secondary consumers or nektic predators including different actinopterygian fish, ammonoids and coleoids. Smaller fish and the abundant cephalopods served as prey for larger carnivorous fish, such as Saurichthys. Thylacocephalan arthropods, represented by Austriocaris and Atropicaris, probably either exhibited a scavenging mode of life near the sea floor, as shown for Ostenocaris (Jobbins et al., 2020; Pinna, 1985), or actively hunted in the water column for conodonts, small cephalopods (Broda et al., 2019) and, as shown for Concavicaris, on other small organisms (see also Broda et al., 2019; Cross et al., 2018; Lukeneder & Lukeneder, 2021). These small arthropods, in turn, were hunted by chondrichthyans or other large fishes (Broda et al., 2019; Ji et al., 2020), thus serving as an important food source for numerous fish taxa (Lukeneder & Lukeneder, 2021).
Important information is gained from bromalites, represented by coprolites and regurgitalites (Lukeneder et al., 2020 and references therein). The regurgitalites here were produced by large durophagous predators, but the cephalopods and arthropods appear to be too small to produce bromalites up to 100 mm in size. The rich ichthyofauna and lack of reptile remains point to predatory fishes as bromalite producers. Krystyn (1991) and Lukeneder and Lukender (2021) noted the occurrence the actinopterygian predators Elpistoichthys, Gigantopterus, Saurichthys, Thoracopterus, Habroichthys, Nannolepis and Peltopleurus. Griffith (1977) stated that the Upper Triassic ichthyofauna of the Polzberg region is characterized by abundant flying fish, which, according to that author, suggests strong predation pressure in this marine ecosystem. The majority of marine fish genera known from Polzberg were predatory (Griffith, 1977; Lukeneder & Lukeneder, 2021).
The regurgitalites from Polzberg typically show specialization of the producer to a cephalopod prey, because they consist of ammonoid shells of A. minor and P. haberfellneri as well as coleoids hooks and cartilage material from P. bisinuata. In contrast, the belemnoid Phragmoteuthis could have fed on actinopterygiid fish and actively hunted other belemnoids or small and slow ceratitid ammonoids. Klug et al. (2021a) described a Lower Jurassic (Toarcian) taphocoenosis with Jeletzkyteuthis coriaceus which fed on a smaller octobrachian Parabelopeltis flexuosa. Conversely, evidence for actinopterygian fish predation on Clarkeiteuthis elsewhere is available from the Lower Jurassic Posidonia Shale (Přikryl et al., 2012), whose deposits also yield evidence for the predation of vampyromorph coleoids on other vampyromorphs (Klug et al., 2021a, b).
No evidence is currently available for marine reptiles at Polzberg (Fig. 7). Nonetheless, late Triassic ichthyosaurs and nothosaurs are known (Arthaber, 1896; Dzik & Sulej, 2007; Huene, 1916; Liu et al., 2014; Neenan et al., 2013; Scheyer et al., 2014; Summesberger & Wagner, 1972) from the Reifling Basin.
Taphonomic mechanisms
Conditions at the sea floor in the constricted basin (Lukeneder et al., 2020; Lukeneder & Lukeneder, 2021;) hindered or decelerated alteration processes, such as scavenging, decay, disarticulation, abrasion, bioerosion and corrosion. Unique preservational features highlight the significance of the Polzberg Konservat-Lagerstätte and the importance of such depositional Lagerstätten in general.
Black remnants of carbonised cephalic retractor and hyponome retractor muscles were described as black areas within trachyceratitid ammonoids A. minor and P. haberfellneri (Lukeneder & Lukeneder, 2022a). The assumed nature of that black structures correlate with lateral muscle attachment scars for the hyponome retractor in Jurassic ammonoids from Russia (Mironenko, 2015a, b), muscle scars in Cretaceous Aconeceras from Russia (Doguzhaeva & Mutvei, 1996; see also Doguzhaeva et al., 2004), strengthened by new data on 3D neutron tomography and X-ray tomography of the position of soft parts in Mid-Jurassic ammonoids (Cherns et al., 2022).
An additional special feature of the Polzberg deposits is the preservation of in situ buccal masses described in detail by Trauth (1935) and more recently by Lukeneder and Lukeneder (2022a). Biostratinomically, such entirely preserved ammonoids exhibiting in situ buccal masses are interpreted as quasi autochthonous faunal elements, where the intact shells sank after death of the animal and neither surfaced nor drifted far from their original habitats (Lukeneder, 2015; Korn & Klug, 2002; Maeda & Seilacher, 1996; Yacobucci, 2018). The anaptychi mentioned herein are interpreted as non-mineralised trachyceratid lower (Anaptychus lunzensis) and upper jaws of the ammonoids A. minor and P. haberfellneri (Lukeneder & Lukeneder, 2022a). This scenario is also strengthened by the faunal (entire fish carcasses, see Tintori, 1992 well-preserved bristle worms), the sedimentological (lamination, no bioturbation, black) and the geochemical data (total organic carbon, pyrite; Lukeneder et al., in prep.). As noted by Doguzhaeva et al., (2007a, b) Lehmann (1980), Lukeneder (2015), Lukeneder & Tanabe (2002), Reboulet et al. (2003) and Schlögl et al. (2011), the cephalopod versus aptychi ratio (even in situ) is a useful criterion for evaluating the postmortem transport of ammonoids.
The quantity and type of shell damage in the palaeobiota are significant indicators for predator–prey interactions (Hoffmann & Keupp, 2015; Keupp, 2012; Kowalewski, 2002; Lukeneder, 2015). Fragmented shells of ammonoids and phragmoteuthids are abundant and bear sharp edges caused by attacks (Lukeneder & Lukeneder, 2023) of actinopterygian fish or coleoids. Marine reptiles are currently excluded as possible predators of ammonoids (see above). Vertically and subvertically preserved ammonoid shell remains from Polzberg are exclusively bitten fragments, isolated and separated after being attacked from the rest of the shell on the bedding planes (see also Lukeneder & Lukeneder, 2022a; Olivero, 2007).
Here, the trophic interactions involve different predators and numerous prey species. Cephalopods, mainly the dominant ammonoid genera Austrotrachyceras and Paratraychyceras, as well as phragmoteuthids, are both predators and prey. Similar sublethal or lethal shell damage is well-known from recent and fossil cephalopods (Klug et al., 2021a; Lukeneder, 2015; Mapes & Chaffin, 2003). In general, predation in the water column can involve ichthyosaurs, mosasaurs, nothosaurs, cartilaginous or bony fish [pycnodonts (Ward, 1981); holostei (Mehl, 1978, Richter, 2009); teleosts (Keupp, 2000), semionitids (Hoffmann & Keupp, 2015)] and invertebrates including other ammonoids, nautiloids and coleoids (Hoffmann & Keupp, 2015; Klug et al., 2021a; Lukeneder, 2015; Mehl, 1978). Actuopalaeontological analyses and observations show that modern cephalopods (squids, cuttlefish, octopuses) serve as prey for predatory fish worldwide (Smale, 1996). As documented for a variety of Mesozoic ammonoid groups (Klompmaker et al., 2009, 2019) and for ventral bite marks (Andrew et al., 2010, 2015), most attacks were probably caused by coleoids actively preying on the living ammonoids (Klug et al., 2021b; Lukeneder, 2015 Lukeneder & Lukeneder, 2022a). The Polzberg palaeobiota supports this interpretation: numerous specimens are preserved with ventrally and fatally bitten shells. As observed in modern Nautilus, parrotfish (Saunders et al., 1987), triggerfish and groupers (Mapes & Chaffin, 2003) break parts off the nautiloid aperture to expose the soft body. Phragmoteuthids, in turn, attacked the ventral parts of the ammonoids. Injuries inflicted by extant fish can be sublethal or lethal, in contrast to the always fatal bite attacks by coleoid members indicated by the fossil evidence from the Polzberg Konservat-Lagerstätte. Ammonoid shell clusters, stemming from breakage during bite attacks (Mehl, 1978), are frequent in the Polzberg palaeobiota.
Predatory genera are dominated by actinopterygians within the fish community (Lukeneder & Lukeneder, 2021; Lukeneder et al., 2020; Simms & Ruffel, 1990). The largest predator of that group is the agile Saurichthys, which actively hunted other actinopterygians (Argyriou et al., 2015) and probably also ceratitid ammonoids. In contrast, the belemnoid Phragmoteuthis fed on actinopterygians and hunted small and slow austrotrachyceratid ammonoids (Lukeneder & Lukeneder, 2022a). Such strategies were reported for Jurassic Plesioteuthis and Trachyteuthis (Hoffmann et al., 2020), exhibiting stomach contents with ammonoid (i.e., lamellaptychi) and actinopterygian remains. Additional evidence is available for actinopterygian predation on Phragmoteuthis (Přikryl et al., 2012) and for predation of coleoids on other coleoids (Klug et al., 2021a; b). Saurichthys is an important predator based on its size, abundance and predatory behaviour (Lukeneder et al., 2020; Lukeneder & Lukeneder, 2021).
Polzberg bromalites contain all ammonoid size classes from juveniles to adults. These are either entirely preserved or, in other accumulations, dominated by fragments to shell hash. For the two genera Austrotraychyceras and Paratrachyceras, the presence of the shell along with anaptychi and upper jaws suggests that the entire animal was swallowed (Lukeneder et al., 2020, Lukeneder & Lukeneder, 2021; 2022a). Ammonoid shell accumulations consisting of phragmocones or fragments have been described in the literature and interpreted as reflecting predation (Lukeneder, 2015; Lukeneder et al., 2020). Research on bromalites, stomach contents and palaeopathologies in ammonoids (Hoffmann et al., 2020) led to the conclusion that coleoids (e.g., Plesioteuthis and Trachyteuthis) were among the key predators of ammonoids in Mesozoic marine ecosystem (Andrew et al., 2010, 2015; Klompmaker et al., 2009; Klug et al., 2021a, b; Mehl, 1978).
The excellent preservation of entire organisms and corresponding bromalites documents trophic interactions (food chains and food web) and predator–prey relationships (Klug et al., 2021a, b; Lukeneder et. al., 2020; Lukeneder & Lukeneder, 2021; Lukeneder & Lukeneder, 2022a; Přikryl et al., 2012; Vullo, 2011). Nektic members of the Carnian assemblage such as ammonoids (trachyceratids), coleoids (phragmoteuthids) or fish (mostly actinopterygians) form the main predatorsof the Polzberg palaeobiota and palaeo-food web (Lukeneder et al., 2020; Fig. 7).
Conclusions
Since former material from the Polzberg Konservat-Lagerstätte stems solely from historical collections, new excavations with bed-by-bed collecting were done during four field campaigns in 2021. We present data based on thousands of new fossils from the Late Triassic (Early Carnian) that provide new details and insights into the Polzberg palaeocommunity, supporting hypotheses presented by the authors in earlier papers. We show that the Polzberg locality yields producers, consumers, as well as small and large predators in the Reifling intraplatform basin from the Carnian Pluvial Episode. The Reingraben Shales were deposited at the epipelagic to upper mesopelagic sea floor of the Reifling Basin, which was periodically disconnected from oxygenated bottom currents. In the low-oxygen ecosystems at Polzberg, halobiid bivalves were the dominant (up to rock-forming) epifaunal elements. In the overlying oxygenated water column, ceratitid nektic/nektobenthic ammonoids and nektic actinopterygian fish prevailed. Fish and phragmoteuthids dominated among nektic predators. The changing ecological conditions over 500,000 years caused a shift from the dominant nektic ceratitid member Autrotrachyceras minor to the smaller and less ornamented ceratitid Paratrachyceras haberfellneri. Frequent findings of shell fragments left behind by nektic predators, along with numerous bromalites (coprolites and regurgitalites), shed light on the predator–prey relations in the water column. The effect of scavengers and grazers (thylacocephalan arthropods, crustaceans, polychaetes) was followed by decomposition processes of organic material by bacteria or fungi. The entire fossil assemblage provides new insights into the Late Triassic food webs of this Carnian marine ecosystem. The preservation of fragile skeletons of nektic and benthic taxa and the preservation of entire carcasses as well as soft parts point to unique palaeoenvironmental conditions in the Carnian dysoxic bottom water of the Reifling Basin. The occasional freshwater influx from the surrounding Wetterstein Platform resulted in terrigenous siliciclastic and argillaceous sedimentation, accompanied by terrestrial plant material, a single freshwater dipnoid lungfish and frequent branchiopods.
Availability of data and materials
All sources of information are provided in the text. Raw data related to the fossil material from Polzberg are available from the corresponding author upon request. Data will be made available upon publication on the project homepage (https://www.nhm-wien.ac.at/forschung/geologie__palaeontologie/forschungsprojekte/polzberg) connected to a server of the Natural History Museum Vienna. Images or additional informations are available upon request from AL. The fossil material is incorporated in the collections of the Natural History Museum Vienna (NHMW 2012/0288/0001-6397; NHMW, NHMW 2021/0123/5290, NHMW 2022/0123/0001-0169).
References
Abel, O. (1906). Fossile Flugfische. Abhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 56, 1–88.
Allison, R. A. (1988). Konservat-Lagerstätten: Cause and classification. Palaeobiology, 14, 331–344.
Andrew, C., Hoew, P., Paul, C. R. C., & Donovam, S. K. (2010). Fatally bitten ammonoids from the lower Lias Group (Lower Jurassic) of Lyme Regis, Dorset. Proceedings of the Yorkshire Geological Society, 58(2), 81–94.
Andrew, C., Howe, P., & Paul, C. (2015). Fatally bitten ammonites from septarian concretions of the ‘Marston Marble’ (Lower Jurassic, Sinemurian), Somerset, UK, and their taphonomy. Proceedings of the Geologists’ Association., 126(1), 119–129.
Argyriou, A., Clauss, M., Maxwell, E. E., Furrer, H., & Sánchez-Villagra, M. R. (2015). Exceptional preservation reveals gastrointestinal anatomy and evolution in early actinopterygian fishes. Scientific Reports, 6, 18758. https://doi.org/10.1038/srep18758
Arthaber, G. (1896). Die Cephalopodenfauna der Reiflinger Kalke. Beiträge Zur Paläontologie Und Geologie Österreich-Ungarns Und Des Orients, 10, 1–112.
Bauer, F. K., & Schnabel, W. (1997). Geologische Karte der Republik Österreich. Sheet Mariazell 72, 1:50.000. Geologische Bundesanstalt.
Beardmore, S., & Furrer, H. (2016). Taphonomic analysis of Saurichthys from two stratigraphic horizons in the Middle Triassic of Monte San Giorgio, Switzerland. Swiss Journal of Geosciences, 109, 1–16.
Bernardi, M., Avanzini, M., & Bizzarini, F. (2011). Vertebrate fauna from the San Cassiano Formation (early Carnian) of the Dolomites region. Geo Alp, 8, 122–127.
Brandner, R., & Poleschinski, W. (1986). Stratigraphie und Tektonik am Kalkalpensüdrand zwischen Zirl und Seefeld in Tirol. Jahresberichte Und Mitteilungen Des Oberrheinischen Geologischen Vereins, 68, 67–92.
Briggs, D. E. (2014). Konservat-Lagerstätten 40 years on: the exceptional becomes mainstream. In M. Laflamme, J. D. Schiffbauer, & S. Darroch (Eds.), Reading and Writing of the Fossil Record: Preservational Pathways to Exceptional Fossilization The Paleontological Society Papers (Vol. 20, pp. 1–14). Cambridge University Press.
Brinkmann, W. (1982). Die Ichthyosaurier (Reptilia) aus der Grenzbitumenzone (Mitteltrias) des Monte San Giorgio (Tessin, Schweiz) – neue Ergebnisse. Vierteljahresschrift Der Naturforschenden Gesellschaft in Zürich, 143(4), 165–177.
Brinkmann, W. (1997). Die Ichthyosaurier (Reptilia) aus der Mitteltrias des Monte San Giorgio (Tessin, Schweiz) und von Besano (lombardei, Italien) – der aktuella Forschungsstand. Vierteljahresschrift Der Naturforschenden Gesellschaft in Zürich, 142(2), 69–78.
Broda, K., Marynowski, L., Rakociński, M., & Zatoń, M. (2019). Coincidence of photic zone euxinia and impoverishment of arthropods in the aftermath of the Frasnian-Famennian biotic crisis. Scientfic Reports, 9, 16996. https://doi.org/10.1038/s41598-019-52784-4
Cherns, L., Spencer, A. R., Rahman, I. A., Garwood, R. J., Reedman, C., Burca, G., Turner, M. J., Hollingworth, N. T., & Hilton, J. (2022). Correlative tomography of an exceptionally preserved Jurassic ammonite implies hyponome-propelled swimming. Geology, 50(4), 397–401.
Cross, S. R. R., Ivanovski, N., Duffin, C. J., Hildebrandt, C., Parker, A., & Benton, M. J. (2018). Microvertebrates from the basal Rhaetian Bone Bed (latest Triassic) at Aust Cliff, S.W. England. Proceedings of the Geologists’ Association, 129, 635–653.
Dal Corso, J., Bernardi, M., Sun, Y., Song, H., Seyfullah, L. J., Preto, N., Gianolla, P., Ruffell, A., Kustatscher, E., Roghi, G., Merico, A., Hohn, S., Schmidt, A. R., Marzoli, A., Newton, R. J., Wignall, P. B., & Benton, M. J. (2020). Extinction and dawn of the modern world in the Carnian (Late Triassic). Science Advances. https://doi.org/10.1126/sciadv.aba0099
Dal Corso, J., Ruffell, A., & Preto, N. (2018). The Carnian Pluvial Episode (Late Triassic): New insights into this important time of global environmental and biological change. Journal of the Geological Society London, 175, 986–988.
Doguzhaeva, L. A., Mapes, R. H., Summesberger, H., & Mutvei, H. (2007a). The preservation of body tissues, shell, and mandibles in the ceratitid ammonoid Austrotrachyceras (Late Triassic), Austria. In H. N. Landman, R. A. Davis, & R. H. Mapes (Eds.), Cephalopods present and past. New insight and fresh perspectives (pp. 221–238). Springer.
Doguzhaeva, L. A., & Mutvei, H. (1996). Attachment of the body to the shell in ammonoids. In N. H. Landman, K. Taneba, & R. A. Davis (Eds.), Ammonoid paleobiology. Topics in geobiology (Vol. 13, pp. 43–63). Plenum Press.
Doguzhaeva, L. A., Mutvei, H., Summesberger, H., & Dunca, E. (2004). Bituminous soft body tissues in the bodychamber of the Late Triassic ceratitid Austrotrachyceras from the Austrian Alps. Mitteilungen Aus Dem Geologisch-Paläontoligischen Institut Der Universität Hamburg, 88, 37–50.
Doguzhaeva, L. A., & Summesberger, H. (2012). Pro-ostraca of Triassic belemnoids (Cephalopoda) from Northern Calcareous Alps, with observations on their mode of preservation in an environment of northern Tethys which allowed for carbonization of non-biomineralized structures. Neues Jahrbuch Für Geologie Und Palaontologie, Abhandlungen, 266, 31–38.
Doguzhaeva, L., Summesberger, H., & Mutvei, H. (2006). An unique upper Triassic coleoid from the Austrian Alps reveals pro-ostracum and mandibule ultrastructure. Acta Universitatis Carolinae Geologica, 49, 69–82.
Doguzhaeva, L. A., Summesberger, H., Mutvei, H., & Brandstaetter, F. (2007b). The mantle, ink sac, ink, arm hooks and soft body debris associated with the shells in Late Triassic coleoid cephalopod Phragmoteuthis from the Austrian Alps. Palaeoworld, 16, 272–284.
Dzik, J., & Sulej, T. (2007). A review of the early Late Triassic Krasiejów biota from Silesia, Poland. Palaeontologica Polonica, 64, 3–27.
Ekdale, A. A., & Mason, T. R. (1988). Characteristic trace-fossil associations in oxygen-poor sedimentary environments. Geology, 16, 720–723.
Forchielli, A., & Pervesler, P. (2013). Phosphatic cuticle in thylacocephalans: A taphonomic case study of (Arthropoda, Thylacocephala) from the Fossil-Lagerstätte Polzberg (Reingraben shales, Carnian, Upper Triassic, Lower Austria). Austrian Journal of Earth Sciences, 106, 46–61.
Furrer, H. D. (2003). Monte San Giorgio im Südtessin—Vom Berg der Saurier zur Fossil-Lagerstätte internationaler Bedeutung. Neujahrsblatt Der Naturforschenden Gesellschaft in Zürich, 206, 1–64.
Furrer, H., & Vandelli, A. (2014). Guide to the Museum of fossils from Monte San Giorgio Meride (pp. 1–127). Fondazione del Monte San Giorgio.
Glaessner, M. F. (1931). Eine Crustaceen fauna aus den Lunzer Schichten Niederösterreichs. Jahrbuch Der Geologischen Bundesanst, 81, 467–486.
Glaessner, M. F. (1965). Vorkommen fossiler Dekapoden (Crustacea) in Fisch-Schiefern. Senckenbergiana Lethaea, 46, 111–122.
Griffith, J. (1977). The Upper Triassic fishes from Polzberg bei Lunz. Zoological Journal of the Linnean Society, 60, 1–93.
Hoffmann, R., & Keupp, H. (2015). Ammonoid paleopathology. In C. Klug, D. Korn De Baets, K. I. Kruta, & R. Mapes (Eds.), Ammonoid Paleobiology: From Anatomy to ecology. Topics in Geobiology (Vol. 43, pp. 877–926). Springer.
Hoffmann, R., Stevens, K., Keupp, H., Simonsen, S., & Schweigert, G. (2020). Regurgitalites—a winodw into the trophic ecology of fossil cephalopods. Journal of the Geological Society London, 177, 82–102.
Hornung, T., & Brandner, R. (2005). Biochronostratigraphy of the Reingraben Turnover (Hallstatt facies belt): Local black shale events controlled by regional tectonism, climatic change and plate tectonics. Facies, 51, 460–479.
Hornung, T., Brandner, R., Krystyn, L., Joachimski, M. M., & Keim, L. (2007a). Multistratigraphic constraints on the NW Tethyan “Carnin Crisis.” The Global Triassi., 41, 59–67.
Hornung, T., Kogan, I., Wolf, G., & Moosleitner, G. (2019). The Norian fish deposits of Wiestal (“Seefeld Member”, Northern Calcareous Alps, Salzburg, Austria): Taxonomy and palaeoenvironmental implications. Austrian Journal of Earth Sciences, 112, 125–165.
Hornung, T., Krystyn, L., & Brandner, R. A. (2007b). Tethys-wide mid-Carnian (Upper Triassic) carbonate productivity crisis: Evidence for the Alpine Reingraben Event from Spiti (Indian Himalaya)? Journal of Asian Earth Sciences, 30(2), 285–302.
Huene, F. (1916). Beiträge zur Kenntnis der Ichthyosaurier im deutschen Muschelkalk. Palaeontographica, 62, 1–68.
Hunt, A. P., & Lucas, S. G. (2012). Descriptive terminology of coprolites and recent feces, Vertebrate Coprolites. New Mexico Museum of Natural History and Science Bulletin., 57, 153–160.
Hunt, A. P., Miln, J., Lucas, S. G., & Spielmann, J. A. (2012). Vertebrate coprolites. New Mexico Museum of Natural History and Science Bulletin, 57, 1–387.
Ji, C., Tintori, A., Jiang, D., Montani, R., & Confortini, F. (2020). New Thylacocephala (Crustacea) assemblage from the Spathian (Lower Triassic) of Majiashan (Chaohu, Anhui Province, South China). Journal of Paleontology, 95(2), 305–319.
Jobbins, M., Haug, C., & Klug, C. (2020). First African thylacocephalans from the Famennian of Morocco and their role in Late Devonian food webs. Scientific Reports, 10, 5129. https://doi.org/10.1038/s41598-020-61770-0
Keupp, H. (2000). Ammoniten (p. 165). Paläobiologische Erfolgsspiralen.
Keupp, H. (2012). Atlas zur Paläopathologie der Cephalopoden. Berliner Paläobiologische Abhandlungen, 12, 1–392.
Klompmaker, A. A., Kelley, P. H., Chattopadhyay, D., Clements, J. C., Huntley, J. W., & Kowalewski, M. (2019). Predation in the marine fossil record: Studies, data, recognition, environmental factors, and behavior. Earth-Science Reviews, 194, 472–520.
Klompmaker, A. A., Waljaard, N. A., & Fraaije, R. H. B. (2009). Ventral bite marks in Mesozoic ammonoids. Palaeogeography Palaeoclimatology Palaeoecology, 280, 245–257.
Klug, C., Schweigert, G., Fuchs, D., & De Baets, K. (2021a). Distraction sinking and fossilized coleoid predatory behaviour from the German Early Jurassic. Swiss Journal of Palaeontology. https://doi.org/10.1186/s13358-021-00218-y
Klug, C., Schweigert, G., Tischlinger, H., & Pochmann, H. (2021b). Failed prey or peculiar necrolysis? Isolated ammonite soft body from the Late Jurassic of Eichstätt (Germany) with complete digestive tract and male reproductive organs. Swiss Journal of Palaeontology, 140(3), 1–15. https://doi.org/10.1186/s13358-020-00215-7
Korn, D., & Klug, C. (2002). Ammoneae devonicae. In W. Riegraf (Ed.), Fossilium catalogus, animalia I (pp. 1–375). Backhuys.
Kowalewski, M. (2002). The fossil record of predation: An overview of analytical methods. The fossil record of predation. Paleontological Society Special Papers., 8, 1–42.
Kozur, H. W., & Bachmann, G. H. (2010). The middle Carnian Wet Intermezzo of the Stuttgart Formation (Schilfsandstein), Germanic Basin. Palaeogeography Palaeoclimatology Palaeoecology, 290, 107–119.
Krystyn, L. (1991). Die Fossillagerstätten der alpinen Trias. In W. Vasicek, L. Krystyn, & R. Golebiowski (Eds.), Exkursionen im Jungpaläozoikum und Mesozoikum Österreichs. Österreichische Paläontologische Gesellschaft.
Lehman, J.-P. (1975). A propos de Ceratodus sturii Teller, 1891. Bulletin Du Muséum Mational D’histoire Naturelle Paris, 345, 241–246.
Lehman, J.-P. (1978). Note sur les Poissons du Trias de Lunz. I. Thoracopterus Bronn. Annalen Des Naturhistorischen Museums Wien, 82, 53–66.
Lehmann, U. (1980). Ammonite jaws and soft parts. In M.R. House & J. R. Senior (Eds.), The Ammonoidea. The evolution, classification, mode of life and geological usefulness of a major fossil group. The Systematics Association, spec (Vol. 18, pp 275–287). Academic Press, London.
Liu, J., Hu, Sx., Rieppel, O., Jiang, D., Benton, M. J., Kelley, N. P., Aitchison, J. C., Zhou, C., Wen, W., Huang, J., Xie, T., & Lv, T. (2014). A gigantic nothosaur (Reptilia: Sauropterygia) from the Middle Triassic of SW China and its implication for the Triassic biotic recovery. Scientific Reports, 4, 7142. https://doi.org/10.1038/srep07142
Lukeneder, A. (2015). Ammonoid habitats and life history. In C. Klug, D. Korn, K. De Baets, I. Kruta, R. H. Mapes (Eds.), Ammonoid paleobiology: From anatomy to ecology, topics in geobiology, (Vol. 43, pp. 689–791). Springer Science + Business Media Dodrecht.
Lukeneder, A., & Lukeneder, P. (2021). The Upper Triassic Polzberg palaeobiota from a marine Konservat-Lagerstätte deposited during the Carnian Pluvial Episode in Austria. Nature Research, Scientific Reports, 11, 16644. https://doi.org/10.1038/s41598-021-96052-w
Lukeneder, A., & Lukeneder, P. (2022a). Taphonomic history and trophic interactions of an ammonoid fauna from the Upper Triassic Polzberg palaeobiota. Scientific Reports, 12, 7455. https://doi.org/10.1038/s41598-022-11496-y
Lukeneder, P., & Lukeneder, A. (2022b). Mineralized belemnoid cephalic cartilage from the Late Triassic Polzberg Konservat-Lagerstätte (Austria). PLoS ONE, 17(4), e0264595. https://doi.org/10.1371/journal.pone.0264595
Lukeneder, A., & Lukeneder, P. (2023). Ammonite taxonomy of the Carnian Polzberg Konservat-Lagerstätte in Austria. Wien: Jahrbuch der Geologischen Bundesanstalt.
Lukeneder, A., Surmik, D., Gorzelak, P., Niedźwiedzki, R., Brachaniec, T., & Salamon, M. A. (2020). Bromalites from the Upper Triassic Polzberg section (Austria); insights into trophic interactions and food chains of the Polzberg palaeobiota. Scientific Reports, 10, 20545. https://doi.org/10.1038/s41598-020-77017-x
Lukeneder, A., & Tanabe, K. (2002). In situ finds of aptychi in the Barremian of the Alpine Lower Cretaceous (Barremian, Northern Calcareous Alps, Upper Austria). Cretaceous Research, 23, 15–24.
Lukeneder, S., Lukeneder, A., Harzhauser, M., İslamoğlu, Y., Krystyn, L., & Lein, R. (2012). A delayed carbonate factory breakdown during the Tethyan-wide Carnian Pluvial Episode along the Cimmerian terranes (Taurus, Turkey). Facies, 58, 279–296.
Maeda, H., & Seilacher, A. (1996). Ammonoid taphonomy. In N. H. Landman, K. Taneba, & R. A. Davis (Eds.), Ammonoid paleobiology. Topics in geobiology (Vol. 13, pp. 543–578). Plenum Press.
Mapes, R. H., & Chaffin, D. T. (2003). Predation on cephalopods. A Genaral overview with case study from the Upper Carboniferous of Texas. In T. A. Hansen, P. H. Kelley, & M. Kowalewski (Eds.), Predator-prey interaction in the fossil record (pp. 177–213). Kluwer Academic/Plenum Publishers.
Martin, M. (1982). Revision von Tellerodus sturii (Teller) 1891. Verhandlungen Der Geologischen Bundesanstalt, 1982, 21–31.
Mehl, J. (1978). Anhäufungen scherbenartiger Fragmente von Ammonitenschalne im süddeutschen Lias und Malm und ihre Deutung als Frassreste. Berichte der naturforschenden Gesellschaft zu Freiburg Breisgau, 68, 75–93.
Mironenko, A. A. (2015a). The soft-tissue attachment scars in Late Jurassic ammonites from Central Russia. Acta Palaeontologica Polonica, 60(4), 981–1000.
Mironenko, A. A. (2015b). Soft-tissue preservation in the Middle Jurassic ammonite Cadoceras from Central Russia. Swiss Journal of Palaeontology, 134, 281–287.
Mojsisovics, J. A. E. (1893a). Die Cephalopoden der Hallstätter Kalke - Band I. Abhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 6, 1–835.
Mojsisovics, J. A. E. (1893b). Die Cephalopoden der Hallstätter Kalke - Band II. Abhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 6, 1–524.
Montagna, M., et al. (2018). The Middle Triassic Lagerstätte of Monte SANG Giorgio reveals the oldest lace bugs (Hemiptera: Tingidae): Archetingis ladinica gen. n. Sp. n. Rivista Italiana Die Paleontologia e Stratigrafia, 124, 35–44.
Mueller, S., Krystyn, L., & Kürschner, W. M. (2016). Climate variability during the Carnian Pluvial Phase—a quantitative palynological study of the Carnian sedimentary succession at Lunz am See, Northern Calcareous Alps, Austria. Palaeogeography Palaeoclimatology Palaeoecology, 441, 198–211.
Neenan, J. M., Klein, N., & Scheyer, T. M. (2013). European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications, 4, 1621. https://doi.org/10.1038/ncomms2633
Ogg, J. G. (2015a). Triassic. Chapter 25. In F. M. Gradstein, J. G. Ogg, M. D. Schmitz, & G. M. Ogg (Eds.), The geologic time scale 2012 (Vol. 2, pp. 681–730). Elsevier.
Ogg, J. G. (2015b). The mysterious Mid-Carnian “Wet Intermezzo” global event. Journal of Earth Science, 26, 181–191.
Olivero, E. B. (2007). Taphonomy of ammonites from the Santonian-Lower Campanian Santa Marta Formation, Antarctica: Sedimentological controls on vertically embedded ammonites. Palaios, 22, 586–597.
Piller, W. E., Egger, H., Erhart, C. W., Gross, M., Harzhauser, M., Hubmann, B., van Husen, D., Krenmayr, H.-G., Krystyn, L., Lein, R., Lukeneder, A., Mandl, G., Rögl, F., Roetzel, R., Rupp, C., Schnabel, W., Schönlaub, H. P., Summesberger, H., Wagreich, M., & Wessely, G. (2004). Die Stratigraphische Tabelle von Österreich 2004 (sedimentäre Schichtfolgen). Kommission für die Paläontologische und stratigraphische Erforschung Österreichs. Wien: Österreichische Akademie der Wissenschaften und Österreichische Stratigraphische Kommission.
Pinna, G. (1985). Exceptional preservation in the Jurassic of Osteno. Philosophical Transaction of the Royal Society London, B, 311, 171–180.
Přikryl, T., Košt’ák, M., Mazuch, M., & Mikuláš, R. (2012). Evidence for fish predation on a coleoid cephalopod from the Lower Jurassic Posidonia Shale of Germany. Neues Jahrbuch Für Geologie Und Palaontologie, Abhandlungen, 263, 25–33.
Ramanan, R., Kim, B. H., Cho, D. H., Oh, H. M., & Kim, H. S. (2016). Algae–bacteria interactions: Evolution, ecology and emerging applications. Biotechnology Advances, 34(1), 14–29.
Reboulet, S., Mattioli, E., Pittet, B., Baudin, F., Olivero, D., & Proux, O. (2003). Ammonoid and nannoplankton abundance in Valanginian (early Cretaceous) limestone–marl successions from the southeast France Basin: Carbonate dilution or productivity? Palaeogeography Palaeoclimatology Palaeoecology, 201, 113–139.
Reis, O. M. (1900). Coelacanthus lunzensis Teller. Jahrbuch Der Kaiserlich-Königlichen Geologischen ReichsanstAlt, 50, 187–192.
Richter, A. E. (2009). Ammoniten-Gehäuse Mit Bissspuren. Berliner Paläontologische Abhandlungen, 10, 297–305.
Ruffell, A., Simms, M. J., & Wignall, P. B. (2015). The Carnian Humid Episode of the late Triassic: A review. Geological Magazine, 153, 271–284.
Ruttner, A., & Schnabel, W. (1988). Geologische Karte der Republik Österreich. Sheet Ybbsitz 71, 1:50.000. Wien: Geologische Bundesanstalt.
Saunders, W. B., Spinosa, C., & Davies, L. E. (1987). Predation on Nautilus. In W. B. Saunders & N. H. Landman (Eds.), Nautilus. The biology and paleobiology of a Living Fossil Topics in Geobiology (Vol. 6, pp. 201–212). Springer Press.
Savrda, C. E., & Bottjer, D. J. (1991). Oxygen-related biofacies in marine strata: An overview and update. Geological Society London, 58, 201–219.
Schlögl, J., Chirat, R., Balter, V., Joachimski, M., Hudáčková, N. & Quillévéré, F. (2011). Aturia from the Miocene Paratethys: An exceptional window on nautilid habitat and lifestyle. Palaegeography Palaeoclimatology Palaeoecology, 308, 330–338.
Schädel, M. V., Eldijk, T., Winkelhorst, H., Reumer, J. W., & Haug, J. (2020). Triassic Isopoda – three new species from Central Europe shed light on the early diversity of the group. Bulletin Geosciences, 95(2), 145–166.
Scheyer, T. M., Romano, C., Jenks, J., & Bucher, H. (2014). Early Triassic Marine Biotic Recovery: The Predators’ Perspective. PLoS ONE, 9(3), e88987. https://doi.org/10.1371/journal.pone.0088987
Schlager, W., & Schöllnberger, W. (1974). Das Prinzip stratigraphischer Wenden in der Schichtfolge der Nördlichen Kalkalpen. Mitteilungen Der Geologischen Gesellschaft Wien, 66, 165–193.
Schultz, O. (2013). Pisces. In W. E. Piller (Ed.), Catalogus Fossilium Austriae (Vol. 3, pp. 1–576). Austrian Academy of Sciences.
Seilacher, A. (1970). Begriff und Bedeutung der Fossil-Lagerstätten (Concept and meaning of fossil lagerstätten). Neues Jahrbuch Für Geologie Und Paläontologie, Abhandlungen, 1970, 34–39.
Simms, M. J., & Ruffell, A. H. (1990). Climatic and biotic change in the late Triassic. Journal of the Geological Society London, 147, 321–327.
Simms, M. J., & Ruffell, A. H. (2018). The Carnian Pluvial Episode: From discovery, through obscurity, to acceptance. Journal of the Geological Society, 175(6), 989–992.
Smale, M. J. (1996). Cephalopods as prey. IV. Fishes. Philosophical Transactions Oft He Royal Society London, Biological Science, B, 351, 1067–1081.
Stur, D. (1874). Neue Aufschlüsse im Lunzer Sandsteine bei Lunz und ein neuer Fundort von Wengerschiefer im Pölzberg zwischen Lunzersee und Gaming. Verhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 1, 271–273.
Stur, D. (1886). Vorlage des ersten fossilen Schädels von Ceratodus aus den ober triadischen Reingrabner Schiefern von Pölzberg nördlich bei Lunz. Verhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 15, 381–383.
Summesberger, H., & Wagner, L. (1972). Der Stratotypus des Anis (Trias). Annalen Des Naturhistorischen Museums Wien, 76, 515–538.
Teller, F. (1891). Über den Schädel eines fossilen Dipnoers, Ceratodus Sturii nov. spec. aus den Schichten der oberen Trias der Nordalpen. Abhandlungen Der Kaiserlich-Königlichen Geologischen Reichsanstalt, 15, 1–38.
Tintori, A. (1990). Estherids from the Kalkschieferzone (Triassic) of Lombardy (N. Italy). In E. Robba (Ed.), Atti del Quarto Simposio di Ecologia e Palecologia delle Comunità’ Bentoniche Sorrento (Vol. 1988, pp. 95–105). Museo Regionale di Science Naturali.
Tintori, A. (1992). Fish taphonomy and Triassic anoxic basins from the Alps: A case history. Rivista Italiana Di Paleontologia e Stratigrafia, 97, 393–408.
Tollmann, A. (1976). Analyse des klassischen nordalpinen Mesozoikums (pp. 1–580). Franz Deuticke.
Trauth, F. (1935). Die Aptychen der Trias Sitzungsberichte der Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Klasse. Abteilung., 144, 455–483.
Vullo, R. (2011). Direct evidence of hybodont shark predation on Late Jurassic ammonites. Naturwissenschaften, 98, 545–549.
Ward, P. D. (1981). Shell sculpture as a defensive adaptation in ammonoids. Paleobiology, 7, 96–100.
Wignall, P. B., & Hallam, A. (1991). Biofacies, stratigraphic distribution and depositional models of British onshore Jurassic black shales. Modern and Ancient Continental Shelf Anoxia. Geological Society London, 58, 291–309.
Yacobucci, M. M. (2018). Postmortem transport in fossil and modern shelled cephalopods. PeerJ, 6, e5909. https://doi.org/10.7717/peerj.5909
Acknowledgements
We are particularly grateful to the owners of the outcrop area, Franziska and Hermann Hofreiter, Eva and Karl Jagersberger (all Gaming) for sampling and digging permission. The work was done within the framework of and financially supported by projects of the Austrian Academy of Sciences (headquarters in Vienna, project Polzberg Lukeneder 2021) represented by the National Committee for Geo/Hydro Sciences (Earth System Sciences Programme), the Federal Government of Lower Austria (K3-F-964/001-2020 and K3-F-964/002-2023, Department Science and Research; headquarters in St. Pölten), and the Triassic project “Fenster in die Karnische Krise” of the society of friends of the Natural History Museum Vienna (Freunde des Naturhistorischen Museums Wien, CPE Lukeneder 2023). The manuscript greatly benefited from valuable comments of three anonymous reviewers, the handling editor Christian Klug (Zurich) and the editor-in-chief Daniel Marty (Basel).
Funding
Open access funding enabled and project financially supported by the project ÖAW (Polzberg Lukeneder 2021), the projects Land NÖ (K3-F-964/001-2020, K3-F-964/002-2023), and the Triassic project “Fenster in die Karnische Krise” of the society of friends of the Natural History Museum Vienna (Freunde des Naturhistorischen Museums Wien, CPE Lukeneder 2023) (to AL). The authors are responsible for the contents of this publication. The funder had no impact on conceptualization, design, data collection, analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
AL, designed research. AL, PL collected the new material. AL, PL provided documentation on fossil material. AL, PL contributed to writing the paper; both authors contributed equally to this work, and both edited the final version of MS. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Handling editor: Christian Klug
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lukeneder, A., Lukeneder, P. New data on the marine Upper Triassic palaeobiota from the Polzberg Konservat-Lagerstätte in Austria. Swiss J Palaeontol 142, 9 (2023). https://doi.org/10.1186/s13358-023-00269-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-023-00269-3