
Abstract
Studies of morphology and developmental patterning in adult stages of many invertebrates are hindered by opaque structures, such as shells, skeletal elements, and pigment granules that block or refract light and necessitate sectioning for observation of internal features. An inherent challenge in studies relying on surgical approaches is that cutting tissue is semi-destructive, and delicate structures, such as axonal processes within neural networks, are computationally challenging to reconstruct once disrupted. To address this problem, we developed See-Star, a hydrogel-based tissue clearing protocol to render the bodies of opaque and calcified invertebrates optically transparent while preserving their anatomy in an unperturbed state, facilitating molecular labeling and observation of intact organ systems. The resulting protocol can clear large (> 1 cm3) specimens to enable deep-tissue imaging, and is compatible with molecular techniques, such as immunohistochemistry and in situ hybridization to visualize protein and mRNA localization. To test the utility of this method, we performed a whole-mount imaging study of intact nervous systems in juvenile echinoderms and molluscs and demonstrate that See-Star allows for comparative studies to be extended far into development, facilitating insights into the anatomy of juveniles and adults that are usually not amenable to whole-mount imaging.
Similar content being viewed by others


Introduction
There is a wealth of anatomical diversity across metazoans that can inform questions on the evolution of animal body plans and organ systems. However, modern research in developmental biology has largely focused on a subset of model organisms, introducing some level of bias in our understanding of animal macroevolution. A way to solve this issue is through comparative observations in phylogenetically diverse taxa to expand our understanding of the evolution of organs systems and body plans [1, 2]. Development of broadly applicable, adaptable research approaches is crucial for achieving this goal.
A pivotal challenge in biological research is visualizing cells or molecules within their native tissue context, as well as imaging entire tissues and organisms. This often requires dissection or serial sectioning of tissues followed by reconstruction, which is both destructive and labor-intensive. Such tasks are particularly demanding for complex structures like the nervous system, where it is exceptionally hard to piece together fragmented neurons and intricate neural projections [3, 4]. While sophisticated microscopy techniques can provide high-resolution images of thin samples, achieving detailed images of entire nervous systems, or other organ systems, across whole organisms continues to be a significant obstacle, especially outside of well-characterized model organisms. Compounding this difficulty is the scattering of light due to varying refractive indices (RIs) among biological molecules—such as water, lipids, and proteins—and the absorption of light by endogenous pigments and calcified structures [3]. For some organisms, such as shelled or heavily calcified marine invertebrates, this challenge is insurmountable, making the observation of internal structures in adult stages impossible without dissection or sectioning. In the context of evolutionary developmental biology, this has caused studies to often be restricted to developmental stages that are optically clear, such as embryos and larvae, due to imaging limitations.
Recent advances in tissue clearing methodologies have largely resolved the challenge of varying RIs to enable three-dimension observation of intact mammalian tissues in unprecedented detail, but these tools have received limited use and adaptation in non-mammalian systems [3, 4]. These methods typically aim to: (1) remove lipids from optically dense, fatty tissues like the central nervous system; and (2) match the RI within tissues using specialized media to minimize light scattering. Solvent-based strategies, such as 3DISCO [5] and EZ Clear [6], and strategies relying on detergents or other hydrophilic reagents, such as CUBIC [7, 8], CLARITY [9], and SeeDB [10] for delipidation have enabled molecular interrogation of intact neural circuits within mammalian brains (reviewed in Tainaka et al., [3]). Hydrogel-based strategies, like CLARITY, in which tissues are crosslinked to a polyacrylamide matrix through fixation and gelation, can provide a structural scaffold to support delicate tissues, and have been adapted for tissue clearing of decalcified bones [11]. While some efforts have been made to adapt tissue clearing protocols to non-model systems [12, 13], none have demonstrated the capacity to preserve morphology in delicate, heavily calcified organisms.
Here, we describe a robust and simple method for visualizing organ systems in calcified and pigmented marine invertebrates. This method, which we term See-Star, combines elements from multiple protocols, including hydrogel crosslinking, decalcification, and tissue clearing, into a single protocol. See-Star preserves tissue integrity of delicate, highly calcified marine invertebrates following chemical decalcification. We demonstrate the efficacy of the See-Star protocol on echinoderms and molluscs, two phyla of highly pigmented and calcified animals. We show that See-Star is compatible with common molecular techniques, including immunohistochemistry (IHC) and in situ hybridization (ISH). It enables imaging of molecular labels across scales in large samples, from high-resolution imaging of cellular structures to whole-body imaging of organ systems. Thus, See-Star is a versatile approach that will enable molecular characterization of intact organ systems in developmental stages typically challenging for whole-mount imaging, such as juveniles or young adults, in a variety of non-model organisms.
Results
See-Star renders calcified invertebrates transparent while preserving tissue integrity
Heavily calcified marine invertebrates often become very fragile after decalcification. We reasoned that a hydrogel-crosslinking fixation could provide adequate support to maintain tissue structure through this process. Our goal was to create a protocol that combines robust fixation to preserve delicate tissues with decalcification to eliminate opaque skeletal elements, while also achieving efficient tissue clearing. We hypothesized that using distinct buffers for fixation and gelation would offer the flexibility to independently optimize each step (Fig. 1A). Unlike CLARITY, and related hydrogel-based clearing methods [9, 11], which use a single buffer for both the fixation and gelation steps, our approach allows for this necessary flexibility.

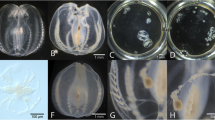
See-Star renders calcified invertebrates transparent while preserving tissue integrity. A Principle of the See-Star clearing procedure, with approximate timing given for each step. B Representative images of Strongylocentrotus purpuratus juveniles before and after preparation with varying concentrations of acrylamide. Scale bar = 1 cm. C Quantification of sample fragmentation observed across varying concentrations of acrylamide. n = 5 per condition. D Brightfield images of representative cleared echinoderm juveniles (left: Patiria miniata, asteroid; right: S. purpuratus, echinoid) imaged before clearing (following fixation) and after clearing. Squares = 600 µm. E Quantification of transparency before and after the clearing process for the species shown in D. n = 3 per condition. In C, E statistical significance determined by Wilcoxon test; *p < 0.05 , **p < 0.01, ***p < 0.001
In preliminary experiments, we observed that the standard 4% acrylamide fixation used in CLARITY, and related hydrogel-based clearing protocols, was insufficient to preserve tissue integrity in adult echinoderms following decalcification (Fig. 1B). We hypothesized that increasing the acrylamide concentration in the fixation step would increase tissue robustness. To test this, we prepared samples of juvenile purple sea urchins, Stronglyocentrotus purpuratus, across a range of acrylamide concentrations. We found that acrylamide concentration correlated with tissue integrity: samples prepared with no or low percentages of acrylamide were severely fragmented following decalcification, whereas samples with the highest percentages of acrylamide survived decalcification and delipidation intact (Fig. 1B, C). To verify that samples fixed with 30% acrylamide could still be rendered optically transparent, we next measured the transparency of samples following refractive index matching. We found that for two different species of echinoderms, S. purpuratus and the sea star Patiria miniata, large (> 1 cm3) juveniles could be rendered nearly transparent using See-Star (Fig. 1D, E).
Comparison of See-Star fluorescence microscopy performance with other clearing methods
To examine the utility of See-Star for fluorescence microscopy, we compared its performance to previously published clearing techniques. We prepared small P. miniata juveniles (150–200 µm thickness) using the See-Star protocol across a range of acrylamide concentrations (4, 15, and 30%) to compare how acrylamide concentration affects imaging depth. For comparison, we also prepared samples with other clearing protocols, including CUBIC and EZ-Clear, and with conventional preparations and mounting media, including 70% glycerol and 80% fructose (Fig. 2A). We stained all samples with DAPI, a highly permeable nucleic acid dye, to assess the impact of optical clarity on imaging depth. We hypothesized that differences in imaging quality would stem from variations in refractive index matching and/or depigmentation, influencing light penetration and scattering.

Comparison of See-Star fluorescence microscopy performance with other clearing methods. A Representative images of Patiria miniata juveniles following different clearing techniques visualized in DIC (A′) or in confocal microscopy after DAPI nuclei staining (Aʺ, Aʺ′). Control samples were imaged in PBS buffer and See-Star cleared samples (4% Acry, 15% Acry, 30% Acry) were imaged in CUBIC mount. For DAPI-stained samples, color-coded (surface: red, base: blue) depth projections indicate the relative depth range for each sample. All samples were imaged from the aboral side of the juveniles, with corresponding orthogonal views shown below (Aʺ). To clearly visualize the entire imaging depth, individual contrast adjustments were done for each condition. Close-ups on surface nuclei in representative areas are shown with identical contrast parameters (A’’’). In A′, Aʺ, scale bar = 200 µm. In Aʺ′, scale bar = 20 µm. B Representative orthogonal projections from the center of the samples shown in Aʺ. C Box plot of average imaging depth at which signal was lost (intensity < average background of 5 a.u. intensity). Grey box indicates the size range of juvenile P. miniata samples, 150–200 µm. D Quantification of mean DAPI fluorescence intensity within nuclei of surface epidermis following different clearing procedures. n = 5 per condition
We found that the See-Star protocol, particularly at 15% and 30% acrylamide concentrations, provided superior optical clarity compared to other methods, followed by 4% acrylamide See-Star and EZ-Clear; these conditions outperformed conventional methods, which were still heavily pigmented following clearing (Fig. 2A′). Imaging depth was greatest in samples prepared using See-Star and did not appear to vary across acrylamide concentrations (Fig. 2Aʺ, B, C). Both See-Star and EZ-Clear enabled imaging across the full depth of samples, unlike other methods where imaging was confined to surface layers (Fig. 2Aʺ, B, C; Supp. Figure 1). We also measured the normalized brightness of the DAPI signal throughout the samples and found that peak signal intensity with See-Star occurred at approximately 50% depth, whereas for other methods, the highest intensity was near the surface, and decreased greatly with depth (Fig. 2C; Supp. Fig. 1). Additionally, to evaluate the effect of mounting media on absolute signal intensity, we observed the brightness of nuclei at the surface, and found that See-Star and CUBIC produced the brightest signal, with negligible difference across acrylamide concentrations (Fig. 2A′ʺ, D). As the 30% acrylamide fixation provided optimal physical integrity (Fig. 1B, C) without diminishing optical clarity (Fig. 2A–C) compared to other conditions, we adopted this as a standard condition for all further samples (Figs. 3 and 4).

See-Star is compatible with immunohistochemistry. IHC in echinoderms (A–N) and molluscs (O–R) following See-Star clearing. A–E Depth projections of acetylated α-tubulin IHC in 2-TF (A), 7-TF (B), and 22-TF (C–E) Patiria miniata juveniles. D Magnification of the distal part of the arm outlined in C. E Magnification of the tube foot papilla outlined in D. F–K Comparison of α-tubulin IHC in a 11-TF P. miniata juvenile (F–H) and a 16-TF Henricia sp. juvenile (I–K). In F, G, I, J, acetylated α-tubulin staining is shown as a color-coded depth projection, with the depth range indicated by the colorbar, so that the nerve ring on the oral side appears in red while the papulae on the aboral side appear in blue. G, J Magnification of the right side of the arms outlined in F and I, respectively. H, K Magnification of an aboral sub-stack of the papulae outlined in F and I, respectively. In H, K, the cyan asterisks indicate the position of the papula. In K, the white arrows indicate condensations of basiepidermal neurites. L-N Depth projections of acetylated α-tubulin IHC in a 11-TF Strongylocentrotus purpuratus juvenile. In L-Lʺ, distinct depth projections along the oral–aboral axis show the oral surface (L), the oral half (L’), and the equator (Lʺ) of the same juvenile. M Magnification of the radial nerve cord outlined in L’. N Magnification of the radial nerve cord outlined in Nʺ. O, P Depth projection of serotonin IHC in Doryteuthis opalescens paralarvae. P Close-up on the region of the stellate ganglion outlined in O. Q, R Acetylated α-tubulin IHC in a juvenile chiton (Lepidozona sp.). The acetylated α-tubulin staining is shown as a color-coded depth projection, with the depth range indicated by the colorbar, so that the dorsal side appears in red while the ventral side appears in blue. R Magnification of the sensory aesthetes outlined in Q. In A-E, H, K–P, nuclei are counter-stained with DAPI. ec: epineural canal; ga: giant axon; irn: interradial nerves; ln: lateral nerve; m: mouth; mn: marginal nerve; mtn: mantle nerve; nr: nerve ring; ob: olfactory bulb; pa: papula; rnc: radial nerve cord; sg: stellate ganglion; sp: spine; st: stomach; tf: tube foot

See-Star is compatible with in situ hybridization. ISH in Patiria miniata (A–F) and Doryteuthis opalescens (G–I) following See-Star clearing. A, B Colorimetric ISH for nkx2.1 in a large P. miniata juvenile. Note that this sample was mounted in SSC buffer and not in RI-matching medium to allow for better contrast with the NBT-BCIP precipitate. B Magnification of the arm outlined in A. C–E Depth projections of HCR FISH for opn-4 in a 8-TF P. miniata juvenile. D Magnification of the distal part of the arm outlined in C. E Magnification of the tube foot papilla outlined in D. F Depth projection of HCR FISH for MHC in a 8-TF P. miniata juvenile. Note that in A–C, F, background signal is present in the digestive tract. G Depth projection of HCR FISH for GFLN1 in a D. opalescens paralarva. Upper panel shows a lateral view, lower panel shows a dorsal view. H, I Double HCR FISH for GFLN1 and SqGluR in a D. opalescens paralarva. H Close-up on the brain region outlined in G. I Close-up on the region of the stellate ganglia outlined in G. In C-I, nuclei are counter-stained with DAPI. gfl: giant fiber lobe; oc: optic cushion; ol: optic lobe; psm: posterior subesophageal mass; rnc: radial nerve cord; sg: stellate ganglion; st: stomach; tf: tube foot
See-Star is compatible with immunohistochemistry in diverse marine invertebrates
Next, we tested whether See-Star was compatible with two of the most commonly used techniques for visualizing molecules, IHC and ISH. For IHC, we used a cross-reactive commercial anti-acetylated α-tubulin antibody, which allows for visualization of neurons and cilia in a broad range of invertebrate species [14,15,16,17,18,19,20]. We first assayed this antibody in cleared P. miniata post-metamorphic juveniles, defined by bearing only two pairs of secondary tube feet in each ray (2-TF stage) (Fig. 3A; Fig. S2A). Neurite tracts within the circumoral nerve ring, the radial nerve cords as well as cilia on the epidermis, in the digestive tract and in the water vascular system were labeled, consistent with previous description of acetylated α-tubulin immunoreactivity in P. miniata at similar developmental stages [21]. However, P. miniata post-metamorphic juveniles are small (diameter ~ 500 µm; thickness ~ 200 µm) and can be easily decalcified by a short EDTA treatment to enable detailed microscopy. To test how See-Star enables whole-mount IHC in more difficult samples, we compare acetylated α-tubulin immunoreactivity of post-metamorphic juvenile stages with larger P. miniata juveniles at the 7-TF stage (diameter ~ 5 mm; thickness ~ 500 µm) and 22-TF stage (diameter ~ 1 cm; thickness ~ 2 mm), which are much more opaque and calcified and would typically be impractical for whole-mount imaging (Fig. 3B, C; Fig. S2B, C). At 7-TF and 22-TF stages, features similar to post-metamorphic stages were labeled, including the circumoral nerve ring, the radial nerve cords, and cilia associated with the digestive tract. Additional structures were labeled in the nervous system at these later stages. At the 7-TF stage, marginal nerves located on either side of the ambulacral regions [22] became clearly visible (Fig. 3B). In addition, series of nerves that we refer to as interradial nerves branched off regularly from the marginal nerves at the level of each tube foot and extended laterally in the interradial areas towards the edges of the oral surface (Fig. 3B). The most proximal interradial nerves coalesced with their counterparts from the neighboring rays along the midline of the interradial area. Despite the large size of the specimens used, high-resolution imaging of smaller anatomical structures was still possible. For instance, we looked at the detail of the acetylated α-tubulin labeling in individual tube feet, showing that the lateral nerves branching off the radial nerve cords were prolonged along the stem of tube feet up to the base of the papilla, where a neurite network lined the entire epidermis, with a condensation facing the medial part of the arm (Fig. 3D, E). These observations are consistent with previous reports of tube feet neuroanatomy in various asteroid species [21, 23, 24]. Additionally, acetylated α-tubulin immunoreactivity also showed the presence of cilia inside the coelomic lining of the tube feet (Fig. 3E), as observed previously [21].
We then validated the compatibility of See-Star with IHC in other echinoderm species by comparing acetylated α-tubulin immunoreactivity in cleared P. miniata with Henricia sp. juveniles (diameter ~ 5 mm; thickness ~ 750 µm), which belong to a distinct asteroid order (Fig. 3F–K; Fig. S2D, E). Both species exhibited similar neural organization, including the nerve ring and radial nerve cords, with marginal and interradial nerves displaying lateral branching at each tube foot. However, Henricia sp. had significantly shorter interradial nerves, aligning with its more compact interradial areas compared to P. miniata (Fig. 3G, J). A notable difference between the two species was the number of dermal papulae present on the aboral surface. In asteroids, papulae are outgrowths of the aboral coelom penetrating through the endoskeleton and protruding in the aboral epidermis (Fig. S3A), likely serving a respiratory function [25]. At the stages investigated, P. miniata juveniles only exhibited a pair of papulae at the proximal end of each arm, while Henricia sp. juveniles had a larger number of papulae spanning the entire surface of each arm (Fig. 3F, I; Fig.S2B, C). In both cases, the lining of the coelom forming the papulae was covered in strongly immunoreactive cilia. Closer examination of the papulae in Henricia sp. also revealed condensations of basiepidermal neurites around and between the base of the coelomic protrusion, which were not observed in P. miniata (Fig. 3H, K; Fig. S2D, E). In both cases, the samples were imaged from the oral side, while the papulae are located on the aboral surface, demonstrating the value of See-Star to allow high-resolution morphological investigation across a thick layer of tissue.
To extend our study outside of asteroid species, we investigated the same antibody in 11-TF S. purpuratus juveniles (diameter ~ 2.5 mm; thickness ~ 1 mm). Unlike post-metamorphic echinoid juveniles that have a thin test and can be observed following short EDTA treatments and simple clearing methods [21, 26], older echinoid juveniles are extensively calcified and are completely opaque, offering a significant challenge for whole-mount imaging. Their fluid-filled cavity encased in a rigid test make them difficult samples for semi-thin sections as well, so that there are no straightforward methods for IHC on internal structures in this type of sample. Here, using See-Star we were able to observe for the first time the structure of the nervous system inside intact late echinoid juveniles (Fig. 3L–N; Fig. S2F), revealing the array of nerves innervating the spines (Fig. 3L), the circumoral nerve ring (Fig. 3L), the serial branching of the lateral nerves along the radial nerve cords (Fig. 3L′, M) and the epineural canal (Fig. 3Lʺ, N). These observations indicate that the general organization of the central and peripheral nervous systems at this stage are very similar to earlier descriptions of post-metamorphic juveniles [21, 27], although complexified by the higher number of appendages each provided with their own innervation. Altogether, our survey of acetylated α-tubulin immunoreactivity in asteroid and echinoid highlights the value of See-Star for comparing intact late developmental stages of large, opaque and highly calcified specimens that would otherwise only be accessible using destructive methods like dissection or semi-thin sections.
We next applied See-Star to two mollusc species to test the utility of the protocol outside of echinoderms in another phylum of heavily pigmented and calcified animals. First, to assess whether See-Star could adequately clear heavily pigmented samples, we attempted to visualize the nervous system of a cephalopod, the California market squid, Doryteuthis opalescens. Cephalopods have a variety of pigment-bearing cell types in the skin that enable the color-changing properties this group is known for [28]. However, this abundance of pigment creates a challenge for observation of underlying structures by light microscopy. We found that D. opalescens paralarvae (length ~ 6 mm; thickness ~ 1.5 mm) processed with See-Star were nearly optically transparent, and a majority of the pigment had been removed (Fig. S3A). Using a commercial cross-reactive anti-serotonin (5HT) antibody, See-Star enabled visualization of the entire serotonergic nervous system of paralarvae (Fig. 3O; Fig. S2G), and also observation of fine structures, such as neural connections within the stellate ganglion (Fig. 3P). We also tested the utility of See-Star as a post-fixation on D. opalescens samples previously fixed in 4% paraformaldehyde and dehydrated in ethanol, and found that it performed equally well on freshly fixed samples and samples dehydrated for long-term storage.
We reasoned that See-Star could also be applied to study structures that closely underlie or pass through shells, such as the sensory aesthetes of chitons, which are neural sensory organs that reside in hollow channels within the valves (shell plates) [29,30,31]. While heavily pigmented and calcified in life, chitons processed with See-Star were rendered nearly transparent (Fig. S3B). Anti-acetylated α-tubulin staining in juvenile chitons (length ~ 5 mm; thickness ~ 500 µm) revealed for the first time the intact organization of the nervous system (Fig. 3Q; Fig. S2H), including neural processes in close proximity to the shell. We were able to observe the ultrastructure of aesthetes by confocal microscopy (Fig. 3R), which until now had only been observed by epoxy casting and electron microscopy.
See-Star is compatible with colorimetric and chain reaction in situ hybridizations
ISH is another technique classically used to detect the localization of mRNAs, and which is highly amenable for non-model species, including in a wide range of marine invertebrates. However, hydrogel-based clearing techniques, such as CLARITY, on which See-Star is based, are not compatible with ISH without additional fixation steps [9, 32]. To test whether the standard 30% acrylamide fixation is sufficient to make See-Star compatible with ISH, we used digoxigenin-labeled riboprobes to analyze the expression of the transcription factor nkx2.1 in large (> 1 cm2) cleared P. miniata juveniles (Fig. 4A, B). We found nkx2.1 to be expressed along the entire length of the radial nerve cords, which is consistent with expression patterns in earlier juvenile stages [33], but also confirmed the expression pattern predicted using RNA tomography at a similar stage [33]. In addition, nkx2.1 was also expressed in the digestive tract, which again was consistent with its expression in 2-TF stages [33]. We then tested the compatibility of the See-Star clearing protocol with the Hybridization Chain Reaction (HCR) split-probe design, a new elaboration of fluorescent ISH (FISH) which uses short DNA probe pairs instead of full-length riboprobes, allowing cloning-free probe synthesis and versatile gene multiplexing [34, 35]. We used two different HCR FISH probe sets for opn-4 and myosin heavy chain (MHC) in 8-TF P. miniata juveniles (Fig. 4C–F). We found that opn-4, which code for a rhabdomeric photopigment [36], was expressed at the tip on each arm in the optic cushion (Fig. 4C, D). This is consistent with the presence in this organ of multiple photosensitive ocelli [37, 38]. In addition, we also found opn-4 to be expressed in discrete cells located at the tip of each tube foot papilla (Fig. 4E). Opsin expression in the tube feet has also been reported in echinoids [39, 40], reinforcing the idea that echinoderm tube feet are important sensory organs in addition to their locomotory role. In addition to opn-4, we looked at the expression of MHC, which is a terminal muscle differentiation marker expressed in striated muscles in all echinoderms [41,42,43]. In P. miniata juveniles, we found this gene to be highly expressed in the water vascular system lining of the tube feet (Fig. 4F), consistent with the presence of longitudinal muscle fibers in this tissue [21]. MHC was also expressed in interradial muscles at the junction of the arms, and in the lining of the digestive tract (Fig. 4F).
To validate that ISH is also compatible with the See-Star protocol outside of echinoderms, we designed HCR FISH probe sets in D. opalescens for two neural markers, the sodium channel GFLN1 [44] and the glutamate receptor, SqGluR [45]. We then investigated the expression of these genes in D. opalescens paralarvae (Fig. 4G–I). The sequence for GFLN1 was originally described from cDNA isolated from stellate ganglion extracts, and it was inferred to be expressed in the giant fin lobe of the stellate ganglion [44]; however, its complete expression pattern was unknown. Here, using See-Star and HCR FISH, we find that GFLN1 is highly expressed within the stellate ganglion, but also broadly in the brain and eyes (Fig. 4G). Our observations confirm that GFLN1 is expressed in the cell bodies within the stellate ganglion that produce the giant fibers of the giant axon. SqGluR is specifically expressed in subregions of the brain, and in the giant fiber lobe of the stellate ganglion. SqGluR expression is enriched in the posterior of the brain, in a sub-region of the posterior subesophageal mass that is largely non-overlapping with expression of GFLN1 (Fig. 4H). Within the stellate ganglion, SqGluR expression is restricted to the giant fiber lobe (Fig. 4I).
Discussion
See-Star is a new clearing protocol which effectively combines hydrogel crosslinking, chemical decalcification, and detergent-based delipidation into a single protocol to enable robust fixation and tissue clearing in large (> 1 cm3), shelled and pigmented invertebrates, while preserving tissue integrity and morphology. The application of See-Star to large specimens, demonstrating deep-tissue imaging and preservation of cellular structures, underscores its potential to push the boundaries of what can be studied in non-model organisms, but also in model organisms outside of the developmental stages which are typically surveyed. This is particularly relevant in the context of many marine invertebrates such as echinoderms or spiralians, in which embryos and larvae are often transparent, while later developmental stages corresponding to the formation of the adult body plan are much more challenging to image. See-Star is amenable for both heavily calcified and soft-bodied organisms, indicating that it is a versatile protocol that is useful for organisms of different body compositions and represents a significant advancement in the visualization of internal structures within calcified organisms, a long-standing obstacle in the field of evo-devo. Another key strength of See-Star, as demonstrated in our comparative microscopy studies, is its compatibility with both IHC and ISH, enabling the detailed visualization of protein and mRNA localization across entire organ systems. This compatibility allows for the exploration of gene expression patterns in stages of development previously inaccessible to whole-mount imaging techniques, particularly in large juvenile and adult stages. In particular, See-Star’s compatibility with HCR FISH makes it highly suitable for comparative studies. Unlike IHC, which relies on the availability of target-specific primary antibodies, HCR FISH enables the exploration of expression patterns of any gene with readily available DNA probes which can be synthesized directly from cDNA sequences without molecular cloning, making it particularly attractive to use in emerging models. These properties make our See-Star protocol an invaluable tool to extend comparative studies in the field of evo-devo and provide new insights into the molecular mechanisms underlying animal body plans and organ systems.
While we have demonstrated that See-Star is ideal for whole mount preparations of calcified marine invertebrates, it may not be suitable for all cases requiring tissue clearing. Therefore, examining the similarities and differences between See-Star and other tissue clearing protocols is useful. See-Star is most closely related to CLARITY in terms of chemistry and procedure and should be viewed as an extension and optimization of this technique for marine invertebrates, especially heavily calcified or shelled species. In CLARITY and related protocols, amine-fixable molecules are chemically crosslinked to acrylamide monomers, which are then polymerized into polyacrylamide chains in a heat-induced reaction to create a biomolecule:polyacrylamide hydrogel meshwork [9]. See-Star relies on the same underlying acrylamide chemistry, but with several major differences including: (1) the separation of the acrylamide crosslinking and hydrogel gelation steps to allow for different buffers with variable acrylamide concentrations; (2) increased concentrations of acrylamide during the fixation step (30% versus 4%); (3) optimization of fixation buffer chemistry to be compatible with the osmolarity of seawater; and (4) the incorporation of chemical decalcification. Unlike Bone-CLARITY, See-Star performs chemical decalcification after acrylamide crosslinking and hydrogel gelation, providing a robust scaffold that enables delicate tissues to survive the decalcification process [11]. Thus, See-Star incorporates a number of innovations that make it uniquely suited for clearing large calcified marine invertebrate samples. However, for soft-bodied animals and small samples, traditional CLARITY or other protocols such as EZClear or CUBIC may be sufficient.
There are several possible further optimizations that could be made to See-Star for tackling difficult tissues beyond what we tested. See-Star enables imaging of thick samples of at least 2 mm thickness (the maximum tested here) by conventional laser-scanning confocal microscopy, but other imaging modalities, such as light sheet microscopy, may enable imaging of even larger samples. Consistent with other studies, we observed that depending on the samples, detergent-based delipidation did not always completely remove all light-absorbing pigments [13]. This was for instance the case in the squid eye and chiton digestive tract (Fig. S4). While the small amount of pigment retained in some samples did not hinder our observations, in other heavily pigmented species it may be necessary to incorporate a bleaching step, such as H2O2 treatment [12, 13]. We also observed that background and autofluorescence can be an issue. This was typically the case for sea star samples larger than 5 mm, in which the gut exhibited some level of autofluorescence despite having been starved for up to one week before the fixation (e.g., Fig. 4C, F). Therefore, for some applications extended washing and addition of an autofluorescence quenching step may be necessary [11]. Lastly, we did not test whether endogenous GFP fluorescence is retained through the See-Star protocol. While this is not an issue in many non-model species where GFP transgenics are not available, it would limit the utility of the method for some model-system applications. However, based on the maintenance of GFP fluorescence through the CLARITY and Bone-CLARITY protocols that have similar chemistry to See-Star, it is likely that GFP would survive the See-Star process [9, 11].
In conclusion, See-Star is a powerful tool for comparative studies in a variety of fields, including developmental biology, neuroscience, and morphology, offering a new lens to examine the diversity of life. By enabling the detailed visualization of internal structures in non-model organisms, See-Star not only enhances our ability to conduct comparative studies, but also opens up new possibilities for the investigation of molecular mechanisms and processes across a broader range of life history stages.
Methods
Animal collection
Adult specimens of Patiria miniata and egg cases of Doryteuthis opalescens were collected off the coast of Monterey Bay, California, US, and kept in circulating seawater tanks. For P. miniata, gravid adults were spawned in the laboratory by injecting 1 mL of 1 mM 1-methyladenine in each gonad. Following in vitro fertilization, P. miniata embryos and larvae were cultured at 14 °C in UV-sterilized filtered seawater (FSW) at a density of about 1 per mL. The seawater was renewed every 2 or 3 days and the larvae were fed ad libitum with freshly grown Rhodomonas lens microalgae. After reaching metamorphosis, the juveniles were fed biofilm cultivated on circulating sea-water tables and supplemented by R. lens until they reached the appropriate size. For D. opalescens, paralarvae were hatched live from egg cases and maintained in flowing seawater. In addition, juvenile Strongylocentrotus purpuratus, Henricia sp., and Lepidozona sp., were collected directly at the appropriate size from the intertidal zone. All animals used in this study were collected with appropriate state permits.
Fixation and clearing
A detailed See-Star protocol is provided in Supplementary Information 1. In short, animals were incubated in clean glass containers with FSW and without any food for at least two days and in some cases up to one week, in order to clear their gut from autofluorescent content. The samples were then anesthetized in a 1:1 mix of 7.5% MgCl and FSW on a flat petri dish before being fixed in 3.3× phosphate buffer saline (PBS) containing 4% paraformaldehyde and 30% acrylamide overnight at 4 °C with gentle agitation. After the fixation, the samples were washed extensively in 1× PBS and incubated in gelation solution (1× PBS, 4% acrylamide, 0.25% VA-044) overnight at 4 °C in septum-capped containers. Following the transfer in gelation solution and after the overnight incubation the containers were de-gassed using a syringe connected to a vacuum line. The gelation itself was performed subsequently by incubating the samples in gelation solution for 2 h at 37 °C. After gelation, the samples were carefully transferred to a decalcification solution (200 mM NaCl, 500 mM EDTA, 50 mM Tris–HCl at pH 8.5) and incubated at 37 °C until any biomineralized structure had disappeared upon visualization under a stereoscope. This step typically took between 1 and 3 days to complete, and for highly calcified samples the incubation temperature was raised to 55 °C. Once biomineralized structures were completely dissolved, the samples were transferred to a clearing solution (200 mM SDS, 200 mM NaCl, 50 mM Tris–HCl at pH 8.5) and incubated at 37 °C until they became partially translucent. This step typically took between 1 and 3 days to complete, and for larger samples the incubation temperature was raised to 55 °C. The samples were then washed extensively in 1× PBS and were ready for subsequent applications. Some of the samples used for IHC and ISH were initially fixed in 4% formaldehyde and dehydrated into 100% ethanol for long-term storage. These samples were rehydrated, post-fixed with acrylamide and cleared following the same procedure.
Immunohistochemistry
After fixation and clearing, specimens were washed extensively with 1× PBS containing 0.5% Tween-20 (PBST) and then incubated for two hours in blocking solution (superblock (ThermoFisher) supplemented with 0.05% triton X-100). The samples were then incubated for 24 to 48 h at 4 °C in blocking solution with the appropriate concentration of primary antibodies. The primary antibodies used in this study were a mouse anti-acetylated tubulin antibody (Sigma-Aldrich #T7451) diluted at 1:200 and a goat anti-serotonin antibody (Immunostar #20080) diluted 1:200. Following primary antibody incubation, the samples were washed extensively in PBS and then incubated overnight at 4 °C in blocking solution with secondary antibodies diluted at 1:500 and with DAPI (Invitrogen) diluted at 1:1000. The secondary antibodies used in this study were goat anti-mouse IgG and goat anti-rabbit IgG antibodies coupled to either Alexa488, Alexa546 or Alexa647 fluorophores (ThermoFisher). Following secondary antibody incubation, the samples were washed extensively in PBST, and then transferred to a refractive index matching mounting solution (50% weight/volume sucrose, 25% weight/volume urea, 25% weight/volume quadrol) modified from the CUBIC clearing protocol [8].
HCR fluorescent in situ hybridization
For HCR FISH, P. miniata and D. opalescens orthologues of opn-4, MHC, GFLN1, and SqGluR were identified from published literature [36, 42, 44, 45] and short antisense DNA probe sets were ordered from Molecular Instruments using the full-length cDNA sequence as input.
HCR FISH was performed as follows. After clearing and extensive washes in PBST, samples were permeabilized in detergent solution (1.0% SDS, 0.5% Tween-20, 150 mM NaCl, 1 mM EDTA, 50 mM Tris–HCl at pH 7.5) for one hour. The samples were then washed in PBST, and then in 5× saline sodium citrate buffer containing 0.1% Tween-20 (SSCT), before being pre-hybridized in hybridization buffer (Molecular Instruments) for 1 h at 37 °C. The probes were then added to the hybridization buffer at a final concentration of 0.05 µM and the samples were let to hybridize overnight at 37 °C under gentle agitation. Following hybridization, the samples were washed 4 times 30 min in probe wash buffer (Molecular instruments) at 37 °C and then in 5× SSCT at room temperature. They were then pre-amplified in amplification buffer (Molecular Instruments) for 30 min. Meanwhile, H1 and H2 components of the HCR amplifiers coupled to Alexa647 fluorophores (Molecular Instruments) were incubated separately at 95 °C for 90 s, cooled down to room temperature in the dark and then pooled together before being added to the amplification buffer at a final concentration of 60 nM. The amplification was then performed overnight at room temperature. The samples were subsequently washed 4 times 30 min in 5× SSCT and incubated overnight in PBST containing DAPI (Invitrogen) diluted at 1:1000. Finally, the samples were washed in PBST and transferred to mounting solution before imaging.
Colorimetric in situ hybridization
For colorimetric ISH, the P. miniata orthologue of nkx2.1 was identified from published literature [46], amplified from mixed stage P. miniata cDNA and cloned into pGEM-T Easy (Promega). Inserts were PCR amplified using M13 primers and template plasmid was digested with DpnI before PCR purification (Qiagen). Digoxigenin-labeled antisense probes were synthesized using SP6 or T7 RNA polymerase (Promega), and purified using LiCl precipitation.
Colorimetric ISH was performed as follows. After clearing and extensive washes in PBST, samples were pre-hybridized in hybridization buffer (100 µg mL−1 heparin, 1 mg mL−1 yeast RNA, 1× Denhardt’s, 5 mM EDTA, 0.1% CHAPS, 0.1% Tween-20, 50% formamide, 5× SSC) at 65 °C for 4 h. The samples were then incubated in hybridization buffer with 1 ng µL−1 of probe overnight at 65 °C. After hybridization, they were successively washed two times 1 h in 2× SSCT at 65 °C, three times 1 h in 0.2× SSCT at 65 °C, and three times 15 min in maleic acid buffer containing 0.5% Tween-20 (MABT) at room temperature. The samples were then blocked in blocking solution (maleic acid buffer containing 2% Boehringer-Mannheim blocking reagent) and incubated overnight at 4 °C in blocking solution containing anti digoxigenin antibody (Roche) diluted at 1:2000. Following antibody incubation, the samples were washed extensively in MABT and then in alkaline phosphatase buffer (100 mM NaCl, 50 mM MgCl2, 0.1% Tween-20, 100 mM Tris–HCl at pH 9.5). The colorimetric reaction was performed by adding NBT/BCIP reagents (Roche) to the alkaline phosphatase buffer, and stopped at the appropriate time with MABT washes. Finally, samples were transferred to mounting solution before imaging.
Image acquisition
Images of cleared samples before and after transfer to mounting solution and colorimetric ISH were taken with either a Zeiss discovery v12 stereoscope or a Zeiss Axioimager A2, in both cases using a Canon EOS T2i DSLR camera. Images of IHC and HCR FISH were taken using a Zeiss LSM700 confocal microscope. For confocal imaging, multichannel acquisitions were obtained by sequential imaging and large samples were acquired using the tile scan option.
Image analysis and quantification
Confocal optical sections spanning regions of interest along the oral–aboral axis of the samples were compiled into maximum intensity z-projections using ImageJ v.1.52 g. In some cases, a temporal color code was applied to visualize z-depth. To quantify fragmentation, images were thresholded to produce a binary mask, and fragments were counted using the ‘Analyze particles’ tool in ImageJ. Fragmentation was calculated as the number of fragments divided by the total area of all fragments, then normalized on a zero to one scale. Transparency was calculated as the ratio of the difference in grayscale intensity values between black and white squares underneath samples divided by the same difference between squares outside of the sample. To quantify nuclear staining from fluorescent images, z-stacks of equivalent area were sub-sampled from the center of each image. Intensity of DAPI staining was then measured by segmenting nuclei using the ‘Analyze particles’ tool in ImageJ, and measuring average nuclear intensity per z slice. To quantify normalized fluorescence intensity across sample depth, intensity values and z-depth were normalized per sample, and then averaged within treatments. Absolute fluorescent intensity was measured from a surficial slice and averaged within treatments.
Availability of data and materials
Data is provided within the manuscript or supplementary information files.
References
Brenowitz EA, Zakon HH. Emerging from the bottleneck: benefits of the comparative approach to modern neuroscience. Trends Neurosci. 2015;38:273–8.
Hejnol A, Lowe CJ. Embracing the comparative approach: how robust phylogenies and broader developmental sampling impacts the understanding of nervous system evolution. Philos Trans R Soc B Biol Sci. 2015;370:20150045.
Tainaka K, Kuno A, Kubota SI, Murakami T, Ueda HR. Chemical principles in tissue clearing and staining protocols for whole-body cell profiling. Annu Rev Cell Dev Biol. 2016;32:713–41.
Azaripour A, Lagerweij T, Scharfbillig C, Jadczak AE, Willershausen B, Van Noorden CJF. A survey of clearing techniques for 3D imaging of tissues with special reference to connective tissue. Prog Histochem Cytochem. 2016;51:9–23.
Ertürk A, Becker K, Jährling N, Mauch CP, Hojer CD, Egen JG, et al. Three-dimensional imaging of solvent-cleared organs using 3DISCO. Nat Protoc. 2012;7:1983–95.
Hsu C-W, Cerda J III, Kirk JM, Turner WD, Rasmussen TL, Flores Suarez CP, et al. EZ clear for simple, rapid, and robust mouse whole organ clearing. Elife. 2022;11:e77419.
Susaki EA, Tainaka K, Perrin D, Kishino F, Tawara T, Watanabe TM, et al. Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis. Cell. 2014;157:726–39.
Susaki EA, Tainaka K, Perrin D, Yukinaga H, Kuno A, Ueda HR. Advanced CUBIC protocols for whole-brain and whole-body clearing and imaging. Nat Protoc. 2015;10:1709–27.
Chung K, Wallace J, Kim S-Y, Kalyanasundaram S, Andalman AS, Davidson TJ, et al. Structural and molecular interrogation of intact biological systems. Nature. 2013;497:332–7.
Ke M-T, Fujimoto S, Imai T. SeeDB: a simple and morphology-preserving optical clearing agent for neuronal circuit reconstruction. Nat Neurosci. 2013;16:1154–61.
Greenbaum A, Chan KY, Dobreva T, Brown D, Balani DH, Boyce R, et al. Bone CLARITY: clearing, imaging, and computational analysis of osteoprogenitors within intact bone marrow. Sci Transl Med. 2017;9:eaah6518.
Konno A, Okazaki S. Aqueous-based tissue clearing in crustaceans. Zool Lett. 2018;4:13.
Pende M, Vadiwala K, Schmidbaur H, Stockinger AW, Murawala P, Saghafi S, et al. A versatile depigmentation, clearing, and labeling method for exploring nervous system diversity. Sci Adv. 2020;6:eaba0365.
Formery L, Wakefield A, Gesson M, Toisoul L, Lhomond G, Gilletta L, et al. Developmental atlas of the indirect-developing sea urchin Paracentrotus lividus: from fertilization to juvenile stages. Front Cell Dev Biol. 2022;10: 966408.
Herranz M, Sørensen MV, Park T, Leander BS, Worsaae K. Insights into mud dragon morphology (Kinorhyncha, Allomalorhagida): myoanatomy and neuroanatomy of Dracoderes abei and Pycnophyes ilyocryptus. Org Divers Evol. 2020;20:467–93.
Martín-Durán JM, Hejnol A. The study of Priapulus caudatus reveals conserved molecular patterning underlying different gut morphogenesis in the Ecdysozoa. BMC Biol. 2015;13:29.
Rawlinson KA. Emryonic and post-embryonic development of the polyclad flatworm Maritigrella crozieri; implications for the evolution of spiralian life history traits. Front Zool. 2010;7:1–25.
Santagata S, Resh C, Hejnol A, Martindale MQ, Passamaneck YJ. Development of the larval anterior neurogenic domains of Terebratalia transversa (Brachiopoda) provides insights into the diversification of larval apical organs and the spiralian nervous system. EvoDevo. 2012;3:3.
Schwaha TF, Handschuh S, Ostrovsky AN, Wanninger A. Morphology of the bryozoan Cinctipora elegans (Cyclostomata, Cinctiporidae) with first data on its sexual reproduction and the cyclostome neuro-muscular system. BMC Evol Biol. 2018;18:92.
Zieger E, Candiani S, Garbarino G, Croce JC, Schubert M. Roles of retinoic acid signaling in shaping the neuronal architecture of the developing amphioxus nervous system. Mol Neurobiol. 2018;55:5210–29.
Formery L, Orange F, Formery A, Yaguchi S, Lowe CJ, Schubert M, et al. Neural anatomy of echinoid early juveniles and comparison of nervous system organization in echinoderms. J Comp Neurol. 2021;529:1135–56.
Cuénot L. Traité de Zoologie. Grassé. 1948.
Elia L, Selvakumaraswamy P, Byrne M. Nervous system development in feeding and nonfeeding asteroid larvae and the early juvenile. Biol Bull. 2009;216:322–34.
Moore SJ, Thorndyke MC. Immunocytochemical mapping of the novel echinoderm neuropeptide SALMFamide 1 (S1) in the starfish Asterias rubens. Cell Tissue Res. 1993;274:605–18.
Cobb JLS. An ultrastructural study of the dermal papulae of the starfish, Asterias rubens, with special reference to innervation of the muscles. Cell Tissue Res. 1978. https://doi.org/10.1007/BF00229616.
Thompson JR, Paganos P, Benvenuto G, Arnone MI, Oliveri P. Post-metamorphic skeletal growth in the sea urchin Paracentrotus lividus and implications for body plan evolution. EvoDevo. 2021;12:3.
Burke RD, Angerer LM, Elphick MR, Humphrey GW, Yaguchi S, Kiyama T, et al. A genomic view of the sea urchin nervous system. Dev Biol. 2006;300:434–60.
Cloney RA, Brocco SL. Chromatophore organs, reflector cells, Iridocytes and Leucophores in Cephalopods. Am Zool. 1983;23:581–92.
Boyle PR. The aesthetes of chitons. Cell Tissue Res. 1976;172:379–88.
Varney RM, Speiser DI, Cannon JT, Aguilar MA, Eernisse DJ, Oakley TH. A morphological basis for path-dependent evolution of visual systems. Science. 2024;383:983–7.
Vendrasco MJ, Fernandez CZ, Eernisse DJ, Runnegar B. Aesthete canal morphology in the Mopaliidae (Polyplacophora)*. Am Malacol Bull. 2008;25:51–69.
Sylwestrak EL, Rajasethupathy P, Wright MA, Jaffe A, Deisseroth K. Multiplexed intact-tissue transcriptional analysis at cellular resolution. Cell. 2016;164:792–804.
Formery L, Peluso P, Kohnle I, Malnick J, Thompson JR, Pitel M, et al. Molecular evidence of anteroposterior patterning in adult echinoderms. Nature. 2023;623:555–61.
Choi HMT, Calvert CR, Husain N, Huss D, Barsi JC, Deverman BE, et al. Mapping a multiplexed zoo of mRNA expression. Development. 2016;143:3632–7.
Choi HMT, Schwarzkopf M, Fornace ME, Acharya A, Artavanis G, Stegmaier J, et al. Third-generation in situ hybridization chain reaction: multiplexed, quantitative, sensitive, versatile, robust. Development. 2018;145: dev165753.
D’Aniello S, Delroisse J, Valero-Gracia A, Lowe EK, Byrne M, Cannon JT, et al. Opsin evolution in the Ambulacraria. Mar Genomics. 2015;24:177–83.
Eakin RM, Brandenburger JL. Effects of light on ocelli of seastars. Zoomorphologie. 1979;92:191–200.
Garm A. Sensory biology of starfish—with emphasis on recent discoveries in their visual ecology. Integr Comp Biol. 2017;57:1082–92.
Ullrich-Lüter EM, Dupont S, Arboleda E, Hausen H, Arnone MI. Unique system of photoreceptors in sea urchin tube feet. Proc Natl Acad Sci USA. 2011;108:8367–72.
Ullrich-Lüter EM, D’Aniello S, Arnone MI. C-opsin expressing photoreceptors in Echinoderms. Integr Comp Biol. 2013;53:27–38.
Andrikou C, Iovene E, Rizzo F, Oliveri P, Arnone MI. Myogenesis in the sea urchin embryo: the molecular fingerprint of the myoblast precursors. EvoDevo. 2013;4:33.
Perillo M, Swartz SZ, Pieplow C, Wessel GM. Molecular mechanisms of tubulogenesis revealed in the sea star hydro-vascular organ. Nat Commun. 2023;14:2402.
Wessel GM, Zhang W, Klein WH. Myosin heavy chain accumulates in dissimilar cell types of the macromere lineage in the sea urchin embryo. Dev Biol. 1990;140:447–54.
Rosenthal JJ, Gilly WF. Amino acid sequence of a putative sodium channel expressed in the giant axon of the squid Loligo opalescens. Proc Natl Acad Sci USA. 1993;90:10026–30.
Battaglia AA, Nardi G, Steinhardt A, Novakovic A, Gentile S, Iaccarino Idelson P, et al. Cloning and characterization of an ionotropic glutamate receptor subunit expressed in the squid nervous system. Eur J Neurosci. 2003;17:2256–66.
Yankura KA, Martik ML, Jennings CK, Hinman VF. Uncoupling of complex regulatory patterning during evolution of larval development in echinoderms. BMC Biol. 2010;8:143.
Acknowledgements
We thank Lauren Lubeck, Paul Bump, and students in the Stanford BIOS236 course for help collecting animal specimens, and members of the Lowe laboratory for helpful discussions.
Funding
This work was supported by an NSF grant (1656628) and Chan Zuckerberg BioHub funding to C.J.L.
Author information
Authors and Affiliations
Contributions
DNC and CJL conceptualized the work, and DNC developed the clearing protocol. DNC and LF conducted experiments and drafted the manuscript. CJL provided financial and intellectual support, and edited the text.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13227_2024_228_MOESM1_ESM.docx
Additional file 1. Figure S1. Normalized sample depth of cleared P. miniata juveniles. Quantification of average sample intensity over normalized sample depth for the different clearing methods assayed on P. miniata juveniles. n = 5 per condition. Figure S2. High-resolution images of selected cleared samples used for IHC (A-H) and HCR FISH (I-J). A 2-TF Patiria miniata juvenile from Fig. 3A. B 7-TF P. miniata juvenile from Fig. 3B. C 11-TF P. miniata juvenile from Fig. 3C. D 11-TF P. miniata juvenile from Fig. 3F. E 16-TF Henricia sp. juvenile from Fig. 3I. F 11-TF Strongylocentrotus purpuratus. juvenile from Fig. 3L’. G Dorytheuthis opalescens paralarva from Fig. 3O. H Lepidozona sp. juvenile from Fig. 3Q. I P. miniata juvenile from Fig. 4C. J P. miniata juvenile from Fig. 4F. Figure S3. Papulae in Patiria miniata and Henricia sp. A Brightfield image of the aboral surface of an arm in a live P. miniata juvenile, showing the protruding papulae. B-E Acetylated α-tubulin IHC showing the aboral surface of a 11-TF P. miniata juvenile (B, D) and a 16-TF Henricia sp. juvenile (C, E). D, E Magnifications of the regions outlined in B and C respectively, showing details of the papulae. m: madreporite; pa: papula. Figure S4. See-Star clearing of Doryteuthis opalescens and Lepidozona sp. Brightfield images of representative cleared D. opalescens paralarvae (A) and Lepidozona sp. (B) imaged before clearing (following fixation) and after clearing. Squares = 600 µm.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Clarke, D.N., Formery, L. & Lowe, C.J. See-Star: a versatile hydrogel-based protocol for clearing large, opaque and calcified marine invertebrates. EvoDevo 15, 8 (2024). https://doi.org/10.1186/s13227-024-00228-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13227-024-00228-0