
Abstract
Background
Babesiosis is a tick-borne infection caused by piroplasmid protozoa and associated with anemia and severe disease in humans, domestic animals and wildlife. Domestic cats are infected by at least six Babesia spp. that cause clinical disease.
Methods
Infection with a piroplasmid species was detected by microscopy of stained blood smears in three sick cats from Israel. Genetic characterization of the piroplasmid was performed by PCR amplification of the 18S rRNA, cytochorme B (CytB) and heat shock protein 70 (HSP70) genes and the internal transcribed spacer (ITS) locus, DNA sequencing and phylogenetic analysis. In addition, Haemaphysalis adleri ticks collected from two cats were analyzed by PCR for piroplasmids.
Results
The infected cats presented with anemia and thrombocytopenia (3/3), fever (2/3) and icterus (1/3). Comparison of gene and loci sequences found 99–100% identity between sequences amplified from different cats and ticks. Constructed phylogenetic trees and DNA sequence comparisons demonstrated a previously undescribed Babesia sp. belonging to the Babesia sensu stricto (clade X). The piroplasm forms detected included pear-shaped merozoite and round-to-oval trophozoite stages with average sizes larger than those of Babesia felis, B. leo and B. lengau and smaller than canine Babesia s.s. spp. Four of 11 H. adleri adult ticks analyzed from cat # 3 were PCR positive for Babesia sp. with a DNA sequence identical to that found in the cats. Of these, two ticks were PCR positive in their salivary glands, suggesting that the parasite reached these glands and could possibly be transmitted by H. adleri.
Conclusions
This study describes genetic and morphological findings of a new Babesia sp. which we propose to name Babesia galileei sp. nov. after the Galilee region in northern Israel where two of the infected cats originated from. The salivary gland PCR suggests that this Babesia sp. may be transmitted by H. adleri. However, incriminating this tick sp. as the vector of B. galilee sp. nov. would require further studies.
Graphical Abstract

Similar content being viewed by others


Background
Babesia Starcovici, 1893, is a tick-borne protozoan genus from the phylum Apicomplexa, class Piroplasmea, order Piroplasmida. Babesia spp. infect domestic and wildlife animals and humans and cause severe hemolytic disease which could eventually lead to a fatal outcome [1]. More than 100 different species of Babesia have been described in domestic mammals and wildlife as well as in some avian species, and Babesia spp. are the second most frequently reported blood parasites of mammals after trypanosomes [2]. New Babesia spp. have been described morphologically and molecularly characterized in a variety of wildlife and domestic vertebrates in the past 20 years [1]. Domestic cats are infected with clinical disease by several Babesia spp. and sub-species which have been characterized genetically and described morphologically (Table 1). These include: (i) Babesia felis Davis, 1929, initially described from a wild cat (Felis ocreata) and then from domestic cats [3,4,5]; (ii) Babesia canis presentii Baneth et al., 2004 [6]; (iii) Babesia leo Penzhorn et al., 2001, initially described in lions (Panthera leo) and then from cats [7,8,9]; (iv) Babesia hongkongensis [10]; (v) Babesia lengau Bosman et al., 2010, initially described in cheetahs (Acinonyx jubatus) and thereafter from cats [11, 12]; and (vi) Babesia panickeri Panicker et al., 2020 [13]. In addition, Babesia sp. Cat Western Cape was reported to cause disease in cats in South Africa but it has not been described as a new species yet [8, 14].
Additional Babesia spp. described to infect cats include Babesia cati reported from an Indian wild cat (Felis catus) [15], Babesia pisicii reported from European wild cats (Felis silvestris) [16] and some Babesia spp. which are typically found in canines and have mostly been detected only by PCR and reported based on molecular data including Babesia canis [17], B. vogeli [18], B. gibsoni [19] and B. vulpes [17]. DNA of Babesia microti, which infects rodents and humans, has also been reported in cat surveys from Europe and Pakistan [20, 21].
This study describes severe clinical babesiosis which was detected in cats brought for veterinary care in Israel. Initial PCR and genetic analysis of the infecting organism suggested that the cats are infected with a previously unknown Babesia sp. The aim of this study was to morphologically describe, genetically characterize and record the clinical disease associated with this piroplasmid organism in domestic cats.
Methods
Animal samples
Piroplasm parasites were detected in erythrocytes by light microscopy in blood smears stained by Romanowsky staining solutions in three domestic cats from Israel whose blood was tested at the laboratory for vector-borne diseases, Koret School of Veterinary Medicine, in Rehovot, Israel, during the years 2008–2023. The medical history, physical examination findings and complete blood count (CBC) as well as other diagnostic and laboratory test results were extracted from the medical records of all cats (Table 2). Blood smears were visualized by oil immersion microscopy at 1000 × magnification, and parasite sizes were measured using a micrometer.
Molecular detection of Babesia and other pathogens in animal samples
DNA was extracted from 200 μl blood of EDTA anticoagulated blood using the Qiagen DNeasy Blood & Tissue Kit (Qiagen, USA) following the manufacturer’s protocol. Babesia spp. molecular characterization was done using several PCRs with primers, and conditions are specified in Table 3. PCR was performed in a total volume of 25 μl using the PCR-ready High Specificity mix (Syntezza Bioscience, Jerusalem) with 4 μl of DNA template, 400 nM of each primer and sterile DNase/RNase-free water (Sigma, St. Loius, MO, USA). Amplification was performed by a programmable conventional thermocycler (Biometra, Goettingen, Germany). Initial denaturation at 95 °C for 5 min was followed by 35–45 cycles of denaturation at 95 °C for 30 s, annealing and extension conditions (Table 3) and final extension at 72 °C for 30 s, unless detailed otherwise. The extension step was continued, after the last cycle, the for an additional 5 min. PCR products were electrophoresed on 1.5% agarose gels stained using ethidium bromide and evaluated for the size of amplified fragments under UV light by comparison to a 100-bp DNA molecular weight marker. Positive DNA controls from naturally infected dogs with B. vogeli were run with each corresponding reaction. Non-template control (NTC) reactions were performed by using the same reagents and procedures described above but without DNA added to the reaction to rule out contaminations. PCR was also used for testing the cat samples for infection with Mycoplasma [17], Hepatozoon, Ehrlichia and Anaplasma spp. (Table 4). Samples were also tested for the presence of antibodies for the feline immunodeficiency virus (FIV) and for feline leukemia virus (FeLV) antigenemia using a commercial assay (SNAP FIV/FeLV Combo Test, IDEXX Laboratories, Westbrook, ME, USA).
DNA from all the positive PCR products was sequenced at the Center for Genomic Technologies, Hebrew University of Jerusalem, Israel, using the BigDye Terminator v3.1 Cycle Sequencing Kit and an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA). The resulting sequences were evaluated using ChromasPro software version 2. 1.1 (Technelysium Pty Ltd., Australia) and compared for similarity with sequences available in GenBank using the BLAST program (http://www.ncbi.nlm.nih.gov/BLAST/). New DNA sequences from Babesia-infected cats in this study were deposited as new accessions in GenBank.
The near full length of the piroplasma 18S rRNA gene was sequenced in parts using 3.2 pmol of the following primers: Nbab1F (5′-AAGCCATGCATGTCTAAGTATAAGCTTTT-3′), TB Rev (5′-AATAATTCACCGGATCACTCG-3′), BT 2R (5′-CCCGTGTTGAGTCAAATTAAGCCG-3′), BT 3F (5′-GGGCATTCGTATTTAACTGTCAGAGG-3′), Nbab 4F (5′-CCGTTAACGGAACGAGACCTTAACC-3′) and Nbab 4R (5′-GGTAGGCCAATACCCTACCG-3′) [22]. Sequences were then reconstructed using MEGA version X (http://www.megasoftware.net) [23]. Multiple sequence alignment (MSA) was done using EMBL-EBI—CLUSTAL OMEGA [24].
Tick samples
Ticks collected from two of the cats with babesiosis in the study (cats 1 and 3) were identified morphologically and analyzed for Babesia infection by PCR. Some of the ticks from cat # 3 were dissected, and their salivary glands, guts and remaining body parts were separated. DNA was extracted from ticks and their organs using a commercial kit (DNeasy Blood & Tissue Kit, Qiagen, Hilden, Germany) following the manufacturer's instructions. PCR of ticks was used to detect the presence of Babesia spp. DNA was analyzed using the piroplasmid PCR and primers (Table 3). All positive DNA amplicons were sequenced as detailed above and identified using the BLAST program as described for Babesia sp. sequences from cat blood.
Phylogenetic analysis
Phylogenetic analyses, which included DNA sequences derived in this study, were performed to compare these sequences to other piroplasmid spp. sequences deposited previously in GenBank. Sequences were analyzed using the MEGA version X [23], and phylogenetic trees were constructed using maximum likelihood algorithms. A model for each phylogenetic tree was chosen according to the Aikaike information criterion (AIC) and was specified for each tree separately. Bootstrap replicates were done to estimate node reliability, and values were extracted from 1000 randomly selected samples of the aligned sequence data.
Results
Family Babesiidae Poche, 1913
Genus Babesia Starcovici, 1893
Babesia galileei sp. nov.
Type-host Domestic cat Felis catus Linnaeus, 1758 (Mammalia: Felidae).
Type-locality City of Holon (32°01′01″N 34°46′45″E), Israel.
Other localities Village of Harashim (32°57′24″N 35°19′41″E), Israel and Kibbutz Yehiam (32°59′49″N 35°13′15″E), Israel.
Type-material A stained thin blood smear from an 11-year-old Israeli male domestic cat containing the holotype (cat. no. 2; Fig. 1h) was deposited in the National Natural History Collection of the Hebrew University of Jerusalem, Israel, under accession number HUJINV 1000000006. In addition, genomic DNA extracted from the blood of infected cats no. 2 and 3 was deposited at the Koret School of Veterinary Medicine, Hebrew University of Jerusalem, Rehovot, Israel, under accession numbers 9394(P) and 2358941(C), respectively.

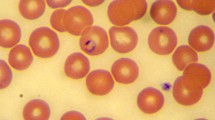
Proposed evolution of Babesia galileei sp. nov. development and division. Round early trophozoite form within a young erythrocyte (a) followed by elongating oval trophozoites (b, c). Trophozoite forms develop a pointed end (d) and begin to divide into merozoites creating a triangular shape (e), which eventually separates into two pear-shaped merozoites which are initially attached to each other (f). Pairs of merozoites gradually become distant from each other (g, h) and may divide again to form four merozoites within the same erythrocyte (i, j). The holotype parasite from cat # 2 is shown in Fig. 1 h. Modified Wright’s and quick Romanowsky staining
Vector Unknown. The ixodid tick Haemaphysalis adleri Feldman-Muhsam, 1951 [25] is suspected.
Representative DNA sequences Present study (GenBank: PP620719-PP62021: 18S rRNA; PP620716-PP620718: ITS; PP24317, PP590338, PP590339: CytB; PP624315, 624316; HSP 70).
ZooBank registration To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN) [26], details of the new species were submitted to ZooBank. The Life Science Identifier (LSID) of the article is urn:lsid:zoobank.org:pub:98EBB21B-88F0-4385-9D64-663DC0E8B975. The LSID for the new name Babesia galileei is urn:lsid:zoobank.org:act:3751D81F-5191-4986-A69D-5D70816E5E65.
Etymology The species is named after the Galilee district in northern Israel where the first cat infected with this parasite was detected.
Description
All measurements are in micrometers ± standard error.
Trophozoites (Measurements based on 84 parasites; see Fig. 1a, d.) Round to oval forms measuring 0.81–2.50 (1.53 ± 0.35) in length and 0.54–1.62 (1.07 ± 0.25) in width (n = 84) with basophilic-staining nuclear material adhering to the parasite’s outer limits. As the trophozoites develop they become more oval and eventually develop a pointed end (Fig. 1d) after which they begin to divide into merozoites creating a triangular shape (Fig. 1e), which eventually separates into two pear-shaped merozoties which are initially attached to each other (Fig. 1f).
Merozoites (Measurements based on 108 parasites; see Fig. 1f, j.) Pear-shaped merozoites with eccentric nuclei presenting as two (Fig. 1f–h) or four intraerythorocytic parasites (Fig. 1i, j). The merozoites are found in different stages of development measuring 0.81–3.05 (1.70 ± 0.37) in length and 0.54–1.39 (0.91 ± 0.25) in width (n = 108) with basophilic staining nuclei in the outer rounded poles of the piroplasm forms (Fig. 1f–h, j). Pairs of merozoites were also seen in erythrocytes that were phagocytosed by large monocytes in the feather edge of the blood smear (Fig. 2).

Two merozoites of Babesia galileei sp. nov. in a phagocytosed erythrocyte within a monocyte. The parasites are present in a monocyte (arrow) at the edge of a blood smear from cat # 1. Modified Wright’s stain. Bar = 5 microns
Differential diagnosis
Intraerythrocytic parasites presented in several shapes. Trophozoites of B. galileei sp. nov. are initially small and round (Fig. 1a) and gradually enlarge and assume an oval shape (Fig. 1b, c). Trophozoite shapes that develop a pointed end (Fig. 1d) begin to divide and widen to a fan-like shape in which two early nuclei may be seen (Fig. 1e) and then complete the division to produce pear-shaped merozoites which are initially attached to each other following binary fission (Fig. 1f) and then separate and detach (Fig. 1g). Merozoites possess basophilic staining dense chromatin nuclei located in the rounded poles of their elongated forms (Fig. 1f–i). Four merozoites were found within some erythrocytes presumably after pairs of merozoites had divided, creating four merozoites which took up about half of the erythrocyte volume (Fig. 1i, j).
The forms of B. galileei sp. nov. described here from cat erythrocytes include pear-shaped merozoites that present in pairs with their pointed ends touching or close to each other, similar to the forms of large Babesia spp. of dogs such as B. vogeli Reichenow, 1937, Babesia canis Pianna & Galli-Vallerio, 1895, and Babesia rossi (Nuttal, 1910) Wenyon, 1926. However, they are much smaller in size than these large-form canine Babesia spp. with the merozoites of B. galileei sp. nov. measuring on average 1.70 × 0.91 µm and the large-form canine Babesia spp. typically measuring 4.5–5.0 × 2.0–2.5 µm [27]. When comparing the merozoites and trophozites of B. galileei sp. nov. to those of other Babesia spp. described in cats, some differences can be noted. The most well-known feline Babesia sp. is B. felis, which was reported by Davis in 1929 from a Sudanese wild cat (Felis ocreata presumably Felis sylvestris) with a diameter of about 1.25 µm [3]. The name B. felis was later ascribed to a small-form Babesia sp. that causes severe disease in domestic cats in South Africa with an average diameter of 1.5 µm [4, 14]. Babesia felis described by Davis in 1929 was found in a naturally infected Sudanese wild cat kitten which did not have clinical signs of disease and maintained the infection with visible parasitemia for at least 12 months, with an episode of anemia which resolved spontaneously. The kitten’s blood was used to experimentally infect 14 cats without causing anemia or clinical disease, except for when experimentally-infected cats were splenectomized and then anemia and hemoglobinuria were detected [3]. Unfortunately, there is no known deposited specimen of this parasite and no genetic characterization of it is available. Babesia felis from South Africa is likely to be a different species from the one described by Davis in Sudan [3] as it appears to be somewhat larger and causes severe and potentially fatal disease in naturally as well as experimentally infected cats [14, 28, 29]. The merozoite stages of B. galileei sp. nov. possess abundant typical pyriform shapes, which are formed following binary fission and present as pairs of parasites attached to each other which then separate often remaining opposing each other as pear shaped couples similar in shape to those seen in large-form canine Babesia spp. In contrast, B. felis was described to rarely form pairs of pyriform or elongated merozoites, and its replication was described as the cruciform formation of four small merozoites in a tetrad shape [4, 29]. The morphology of B. leo is similar to that of B. felis, and they are also very closely related genetically [9]. Babesia lengau also reported from domestic cats in South Africa has merozoties measuring on average 1.91 × 1.1 µm and round to oval forms of 1.3 × 1.3 µm, and it has not been reported to form pairs of pyriform merozoites that present opposing each other [11, 12]. Babesia hongkongensis was described to have round to oval trophozoties with ring forms measuring 1.4 to 1.7 µm resembling small-form Babesia spp. with no description of pyriform merozites to date [10]. Babesia canis subsp. presenti was reported to form pairs of pyriform merozoites and also rounded trophozoites like B. galileei sp. nov. but these forms were larger on average than those measured for B. galileei sp. nov. with merozoites of 2.5 × 1.4 µm and trophozites of 2.7 × 1.7 µm compared to 1.70 × 0.91 µm and 1.53 × 1.07 µm in B. galileei sp. nov. [6]. Babesia panickeri from a domestic cat in southern India was described to have pair-shaped merozoites with average sizes of 2.7 × 1.3 µm, which is larger than those of B. galileei sp. nov. [13]. Lastly, Babesia sp. lynx genotype described from the Eurasian lynx (Lynx lynx) in Turkey had merozoites of 1.9 × 1 µm and trophozoites of 1.87 × 1.55 µm on average, which seem to be similar in shape but slightly bigger than those of B. galileei sp. nov. [30].
Molecular phylogeny
The nearly complete piroplasmid 18S rRNA gene (about 1400 bp) was amplified from the blood of all three cats (GenBank: PP620719-PP620721). Pairwise comparisons showed a 100% identity among the three sequences. As determined by BLAST, the closest match to the cat sequences with 99.8% identity and 97% coverage was a sequence from a Babesia sp. from the Eurasian lynx from Turkey termed Babesia sp. lynx (GenBank: MZ905342) [30] and a sequence from a Haemaphysalis erinacei tick also from Turkey termed Babesia sp. Ankara (GenBank MH504117) [31] with 99.6% identity and 98% coverage (Additional File 1). The 18S rRNA sequences from the Israeli cats in this study were 99.2% identical (100% coverage) to B. rossi (GenBank KY463434), only 95.7% identical (100% coverage) to B. canis presentii (GenBank AY272047) and 95.5% identical (100% coverage) to B. pisicii (GenBank MW939360).
A phylogenetic tree inferred based on 1397-bp sequences of the 18S rRNA gene of B. galileei sp. nov. and other piroplasmids present in GenBank representing a wide range of parasites of the order piroplasmida, including those that have been found to infect cats, was inferred using the maximum likelihood model (Fig. 3). Sequences of the three cats infected with B. galileei sp. nov. grouped within the Babesia s.s. clade (clade X) [32]. They clustered together with each other in a strongly supported [bootstrap (bs) 93] sister clade to Babesia sp. lynx (GenBank: MZ905342) and Babesia sp. Ankara (GenBank MH504117). These Israeli and Turkish piroplasm sequences clustered close yet significantly distinctly to B. rossi from African canines and B. panickeri from an Indian cat and then further away from B. vogeli, B. canis presentii, Babesia pisicii, B. canis and other clade X Babesia s.s. spp. including B. hongkongensis and Babesia sp. Western Cape from domestic cats. The Western group Babesia spp. (clade III), including the feline spp. B. lengau, formed a distinct cluster from the Babesia s.s. (clade X [32]), and the Cytauxzoon spp. infecting domestic cats and wild felines (clade VII) also clustered separately, as well as the B. microti group (clade I) including B. felis and B. leo, which cause disease in domestic cats in southern Africa which branched separately.

An 18S rRNA phylogenetic tree based on the near full length gene. Sequences of Babesia galileei sp. nov. from the cats in this study (black circle) compared to other relevant piroplasmid spp. deposited in GenBank. Clade designation of piroplasmid spp. is shown according to Jalovecka et al., 2019 [32]. The GenBank accession numbers, host and country of origin are included for each sequence. The topology was inferred by using the maximum likelihood method and Tamura-Nei + G model [49]. This analysis involved 44 nucleotide sequences. All positions containing gaps and missing data were eliminated (complete deletion option). There were a total of 1295 positions in the final dataset. Plasmodium falciparum 18S rRNA sequence was used as an outgroup. The scale bar represents the evolutionary distance in the units of the number of nucleotide substitutions per site
Analysis of a 760-bp sequence of the internal transcribed spacer (ITS) which included parts of the 18S rRNA, ITS1 locus, 5.8S rRNA, ITS2 locus and partial 28S rRNA gene [33] showed that all the cats infected with B. galileei sp. nov. had sequences that were 98.94–100% identical to each other (GenBank: PP620716-PP620718). The closest match to the B. galileei sp. nov. cat sequences with 98.75% identity and 84% coverage was a sequence from B. canis susp. presentii (GenBank AY272048), followed by Babesia sp. Ankara (GenBank MH504112; 97% identity; 99% coverage), Babesia sp. lynx (GenBank MZ905341; 95% identity; 100% coverage) and B. rossi (GenBank AF394535; 83.3% identity; 100% coverage) (Additional File 1). Phylogenetic analysis based on a 623-bp-long sequence of the ITS (Fig. 4) showed that B. galileei sp. nov. and B. canis subsp. presentii clustered together and were separated with strong support (bs 85) from Babesia sp. Ankara, which also separated (bs 84) from Babesia sp. lynx. The ITS tree also showed that B. galileei sp. nov. was placed within the Babesia s.s. (clade X) and separately from the B. microti group (clade I).

A phylogenetic tree of ITS piroplasmid sequences. Sequences of Babesia galileei sp. nov. from the cats in this study (black circle) are compared to other relevant piroplasmid spp. ITS sequences deposited in GenBank. Clade designation of piroplasmid spp. is shown according to Jalovecka et al., 2019 [32]. The GenBank accession numbers, host and country of origin are included for each sequence. The topology was inferred by using the maximum likelihood method and HKY + G model [23]. This analysis involved 23 nucleotide sequences. All positions with < 95% site coverage were eliminated, i.e. < 5% alignment gaps, missing data and ambiguous bases were allowed at any position (partial deletion option). There were a total of 449 positions in the final dataset. Theileria annulata ITS sequence was used as an outgroup. The scale bar represents the evolutionary distance in the units of the number of nucleotide substitutions per site
Seven hundred fifty-bp sequences of the B. galileei sp. nov. CytB gene, a mitochondrial gene, were 99.47%–100% identical to each other (GenBank PP624317, PP590338, PP590339). The closest match to the B. galileei sp. nov. cat sequences were CytB sequences from B. pisicii (GenBank MW93962) followed by Babesia sp. lynx (GenBank MZ927094; 96% identity; 100% coverage) and B. rossi (GenBank KC2078230) (Additional File 1). Phylogenetic analysis based on 660-bp sequences of the CytB gene (Fig. 5) showed that B. galileei sp. nov. was separated with strong support (bs 99) from Babesia sp. lynx and also from the canine Babesia s.s. spp. B. rossi, B, vogeli, B. canis and B. gibsoni, corroborating the identity of B. galileei sp. nov. as a novel species.

A phylogenetic tree of CytB gene piroplasmid sequences. Sequences of Babesia galileei sp. nov. from the cats in this study (black circle) are compared to other relevant piroplasmid spp. CytB sequences deposited in GenBank. The GenBank accession numbers, host and country of origin are included for each sequence. The topology was inferred by using the maximum likelihood method and GTR + G model [17]. This analysis involved 16 nucleotide sequences. All positions containing gaps and missing data were eliminated (complete deletion option). There were a total of 498 positions in the final dataset. Cytauxzoon felis cytB sequence was used as an outgroup. The scale bar represents the evolutionary distance in the units of the number of nucleotide substitutions per site
The 330-bp HSP70 gene sequences of B. galileei sp. nov. were derived only from cats # 2 and 3, as PCR for this gene from cat # 1 failed, and were 98.97% identical to each other (GenBank PP624315, PP624316). The closest match to the B. galileei sp. nov. HSP70 sequences was from B. rossi (GenBank AB248737; 95.5% identity, 100% coverage) compared to those sequenced from cat # 2 (Additional File 1). Phylogenetic analysis of a 330-bp DNA sequence of the HSP70 gene with comparison to HSP70 sequences of relevant Babesia spp. found in GenBank (Fig. 6) showed that B. galileei sp. nov. clustered separately with strong support (bs 100) from B. rossi and was also clearly separated from other Babesia s.s. spp. and from other species belonging to different piroplasmid clades.

A phylogenetic tree of HSP70 gene piroplasmid sequences. Sequences of Babesia galileei sp. nov. from the cats in this study (black circle) are compared to other relevant piroplasmid spp. HSP70 sequences deposited in GenBank. The GenBank accession numbers, host and country of origin are included for each sequence. The topology was inferred by using the maximum likelihood method and Kimura two-parameter + G model [50]. This analysis involved 18 nucleotide sequences. All positions with < 95% site coverage were eliminated, i.e. < 5% alignment gaps, missing data and ambiguous bases were allowed at any position (partial deletion option). There were a total of 292 positions in the final dataset. Plasmodium falciparim HSP70 sequence was used as an outgroup. The scale bar represents the evolutionary distance in the units of the number of nucleotide substitutions per site
Overall, the 18S rRNA phylogeny constructed using nearly complete gene sequences and the additional analyses of the ITS locus and CytB and HSP70 genes clearly demonstrated that B. galileei sp. nov. represents a novel distinct individual species with the Babesia s.s. group distinguished from other known piroplasmid spp. including all of those reported to infect felines. The DNA sequences from the Turkish lynx and H. erinaceri tick are the closest piroplasmid sequences to B. galileei sp. nov., yet they form a strongly supported separate sister clade, as shown by the 18S rRNA, ITS and CytB phylogenetic analysis. Babesia canis presentii reported earlier from cats in Israel is also genetically distinct from B. galileei sp. nov., as shown by the 18S rRNA, ITS phylogenetic analysis (Fig. 3).
PCR of ticks
Two adult female ticks were collected from cat # 1, identified morphologically as H. adleri using the identification key of Feldman-Muhsam (1951) [25] and processed as a whole tick for DNA extraction and PCR. The female engorged tick was positive by 18S rRNA PCR for Babesia sp. with a sequence that was 100% identical to B. galileei sp. nov. Eleven ticks, two adult females and nine adult males, were collected from cat # 3. All 11 ticks were identified morphologically as H. adleri. The female ticks from cat # 3 were dissected and were negative by PCR for Babesia in their salivary glands, guts and bodies. Of the male ticks from cat # 3, four were not dissected because of their small size, and DNA extracted from them was 18S rRNA PCR-positive from one tick with a sequence of B. galileei sp. nov. Of the 5 male ticks dissected, three were positive by 18S rRNA PCR, of which two were positive in the salivary glands as well as in their guts, whereas one was positive only in its guts and not in the salivary glands. Altogether, 4 of 11 (36%) H. adelri ticks found on cat # 3 were positive for B. galileei sp. nov. DNA. The presence of B. galileei sp. nov. DNA in the tick’s salivary gland is suggestive of the possibility that H. adleri could be a vector of this pathogen, as Babesia sporozoites develop in the salivary gland before being transferred to the vertebrae host with the tick’s saliva during a blood meal.
Clinical findings in infected cats
The infected cats included three males from northern and central Israel.
Cat # 1 was a 3-year-old castrated male from the rural village Kibbutz Yehiam in the Galilee region of northern Israel who was living indoors with its owners and had free access to the outdoor environment. It was admitted for veterinary medical care on 14 December 2008 in a private veterinary clinic with a history of lethargy and loss of appetite. On physical examination, it had a high rectal body temperature (40.5 ºC; normal range 38.1–39.2 ºC) and was infested with ticks. A complete blood count showed anemia [red blood cell (RBC) 3.66 × 106/ul, reference interval (RI) 6–10.1 × 106//ul; mean cell volume (MCV) 36.4 fL, RI 41.3–52.6 fL; mean corpuscular hemoglobin concentration (MCHC) 38.9 g/d, RI 27–32.8] and thrombocytopenia [platelets (PLT) 90 × 103/ul, RI 156–626 × 103/ul]. A serum biochemistry panel showed azotemia (urea 98 mg/dl RI 35–70 mg/dl; creatinine 3.5 mg/dl, RI 0.4–2 mg/dl) and increased activity of liver and muscle enzymes [alanine aminotransferase (ALT) 131 u/l, RI 15–60; aspartate transaminase (AST) 70 u/l RI 0–60]. The cat tested positive for antibodies to FIV and negative for FeLV using a commercial assay (SNAP FIV/FeLV Combo Test, IDEXX Laboratories, Westbrook, ME, USA) and was negative for Hepatozoon, Anaplasma and Ehrlichia spp. by PCR. A blood smear examination showed infection of RBC with piroplasmid organisms interpreted as Babesia sp., and blood was taken for PCR to verify and genetically characterize the infecting organism. The cat was initially treated with the antibiotic doxycycline (Dexcel Ltd., Israel) at 10 mg/kg orally once daily for 14 days and was then administered an injection of imidocarb dipropionate (Imizol; Schering-Plough, Animal Health) at 3.5 mg/kg subcutaneously once 2 days after admission when the diagnosis of babesiosis was made. It improved clinically 6 days after admission and was visited in the owner’s home 11 days after its initial admission on the 25 December 2008. The cat had recovered clinically but was infested with ticks which were collected for testing, and a second blood sample was collected for Babesia PCR. PCR was positive for Babesia sp. both before imidocarb dipropionate treatment and 9 days after the treatment. Two ticks removed from the cat were identified morphologically as H. adleri [25], one of which also tested PCR positive for the same Babesia sp. found in the cat’s blood.
Cat # 2 was an 11-year-old castrated male from the city of Holon in central Israel that lived mostly indoors and had a history of tick infestation. It was diagnosed in a private veterinary clinic with babesiosis on 26 October 2022 after being treated for 12 days with initial lethargy, loss of appetite, dehydration, fever (40.7 ºC) and anemia. Initial blood tests on 14 October 2022 before the diagnosis of babesiosis revealed normocytic normochromic anemia (RBC 5.2 × 106/ul) and thrombocytopenia (PLT 24 × 103/ul). A serum biochemistry panel showed azotemia [blood urea nitrogen (BUN) 46 mg/dl RI 12–30 mg/dl; creatinine 2.2 mg/dl, RI 0.4–2 mg/dl] and increased activities of the liver enzyme ALT (147 u/l, RI 15–60), and the cat was negative for FIV and FeLV by serology (SNAP FIV/FeLV Combo Test) and for Hepatozoon, Anaplasma and Ehrlichia spp. by PCR. It was treated initially with doxycycline (Dexcel Ltd., Israel) at 10 mg/kg orally once daily for 14 days and then after 4 days when there was deterioration in the cat’s anemia, and it began receiving immune-suppressive therapy because of suspected immune-mediated hemolytic anemia (IMHA) with prednisolone (Rekah Pharmaceutical Products LTD, Holon, Israel) at 4 mg/kg orally once daily and after an additional day also with cyclosporine (Atopica, Novartis Animal Health, Basel, Switzerland) at 5 mg/kg q12h orally every 12 h. On day 11 after the cat’s admission, it was still anemic (RBC 4.71 × 106/ul) and thrombocytopenic (PLT 79 × 103/ul), and no organisms were evident on the blood smear. However, Babesia sp. were visualized within RBC on a blood smear prepared without performing a CBC on the following day. The cat was treated with imidocarb dipropionate at 3.5 mg/kg injected subcutaneously, and it had improved clinically and was more active on the next day, but despite this a blood smear showed Babesia sp. organisms which were still evident in its RBC. The cat improved gradually over the next 14 days and was negative for Babesia in its blood smear but still anemic (RBC 4.32 × 106/ul) when administered a second imidocarb dipropionate 2 weeks after the initial injection of this drug. PCR for Babesia sp. was positive on the 26 October 2022 prior to the first imidocarb dipropionate injection and also a day later when Babesia was detected in the blood smear, but it was negative 2 weeks after the first imidocarb dipropionate treatment and before the second injection. The cat was clinically normal, and its CBC was within RI 25 days after it was diagnosed with babesiosis and received its first imidocarb dipropionate injection.
Cat # 3 was an 11-year-old castrated male from the village of Harashim in the Galilee region of northern Israel who was living indoors with outdoor access. It was admitted to a private veterinary clinic on 8 November 2023 after disappearing from the owner’s house for 8 days and presented with extreme lethargy, loss of appetite and apparent jaundice. It was referred to the Hebrew University Veterinary Teaching Hospital (HUVTH) on the next day where on physical examination it was noted that the cat was dehydrated and thin, had yellow mucous membranes and conjunctivas, a heart murmur, a low rectal body temperature (37.2 ºC) and purulent ocular excretions and was infested with ticks. Blood tests showed anemia (RBC 4.7 × 106/ul, RI 6–10.1 × 106/ul; MCV 39.6 fL, RI 41.3–52.6 fL; MCHC 34.6 g/d, RI 27–32.8, leukocytosis [white blood cells (WBC) 35.9 × 103/ul; RI 6.3–19.6 × 103/ul] composed mainly of neutrophilia (neutrophils 28.9 × 103/ul; RI 3.0–13.4 × 103/ul) and monocytosis (monocytes 2.38 × 103/ul; RI 0–1 × 103/ul; RI) and thrombocytopenia (PLT 68 × 103/ul, RI 156–626 × 103/ul). A blood smear examination showed infection of RBC with piroplasmid organisms interpreted as Babesia sp., and blood was taken for PCR to verify and genetically characterize the infecting organism. A serum biochemistry panel showed increased creatinine (3.57 mg/dl, RI 0.4–2 mg/dl) and increased activity of ALT (82 u/l. RI 15–60) and an extremely high bilirubin level (28.5 mg/dl; RI 0–0.2 mg/dl). The cat was negative for FIV and FeLV by serology (SNAP FIV/FeLV Combo Test), negative for Hepatozoon, Anaplasma and Ehrlichia spp. by PCR and positive by PCR for Candidatus Mycoplasma haemominitum. Eleven ticks which were removed from the cat were identified morphologically as H. adleri and were tested thereafter for infection with Babesia by PCR. The cat was treated with imidocarb dipropionate at 2.5 mg/kg injected subcutaneously 1 day after its admittance to the HUVTH on 10 November 2023. No blood parasites were seen on a blood smear taken 2 days after the imidocarb dipropionate injection on the 12 November 2024; however, blood taken 2 days later on 14 November 2024 was still positive for Babesia sp. DNA by PCR. The cat was hospitalized at the HUVTH for 24 days with the diagnoses of babesiosis, acute kidney disease, pancreatitis and hypertophic cardiomyopathy. During hospitalization, the cat’s anemia gradually improved while its platelet and leukocyte numbers returned to the RI levels within 9 days after admission. Its bilirubin level decreased to 0.67 mg/dl with no visual icterus 17 days after hospitalization but its creatinine levels gradually increased despite fluid treatment and reached the peak of 5.25 mg/dl on 27 November 2023, after which it decreased but did not reach the RI when it was discharged. A month after discharge, the cat was treated at home daily with subcutaneous fluids because of kidney disease that became chronic and was reported to be active with good appetite despite a persistent increased creatinine level of 2.4 mg/dl.
Discussion
This study presents B. galileei sp. nov. as a new taxon by fulfilling the ICZN guideline requirements for a new species [26]. The placement of B. galileei sp. nov. in the genus Babesia, and its segregation in the Babesia s.s. (clade x) within this genus is derived from the molecular phylogenetic analysis of the 18S rRNA, ITS, CytB and HSP70 sequences. The phylogenetic analysis demonstrates the distinct identity of B. galileei sp. nov. as corroborated by the demonstration of a strongly supported clade containing the sequences of the infected domestic cats in each of the constructed phylogenetic trees. Babesia galileei sp. nov. is positioned as a sister clade of Babesia sp. lynx and Babesia sp. Ankara as inferred from the 18S rRNA, ITS and CytB trees. Both of the latter parasites were reported from Turkey, which is situated north of Israel in the Middle East [30, 31]. However, it should be noted that no wildlife lynx species, including the Eurasian lynx, which is described as the host of Babesia sp. lynx, are resident in Israel. Interestingly, both the Turkish Babesia genotypes and B. galileei sp. nov. group are closely related to B. rossi, a canine species, according to the 18S rRNA, ITS and CytB phylogenetic trees. Babesia rossi causes disease in dogs in Africa, and although it has mainly been described as a cause of severe babesiosis in domestic dogs from southern Africa, it is also present as far north in Africa as Sudan [34, 35]. The Black-backed jackal (Canis mesomelas) is infected sub-clinically with B. rossi and is considered the reservoir of this infection in South Africa [35, 36]. The presence of genetically related Babesia spp. along the route from Africa to Turkey, passing Israel on the way, is of evolutionary interest, as there are also animals that originated in Africa and are found following this route such as the rock hyrax (Procavia capensis) [37]. Therefore, the dispersion of Babesia spp. with migrating wildlife hosts is a likely route of parasite distribution.
The tick species H. adleri, which was found on cats # 1 and 3, is a palearctic Ixodid species that was described initially from Israel in 1951 [25] and has since then been reported also from Lebanon [38], the Palestinian Authority [39] and Iraq [40]. The vertebrate hosts of H. adleri include a variety of wildlife and domestic animals including mustelid, canids, suidae and felid spp. [41]. A survey of ectoparasites on cats from Jerusalem, Israel, has found that 3 of 340 cats (0.9%) were infested with H. adleri, and infestation was more common in the winter season [42]. Winter in Israel approaches at the end of October and continues until late March. The cases of babesiosis in the cats reported in this study, in which two cats were infested with H. adleri, occurred between the end of October and mid-December. This agrees with the expected period of H. adleri’s activity. The reported limited geographic distribution of H. adleri, if indeed it is the parasite’s vector, may also explain why B. galileei sp. nov. has not been reported so far from other distant countries except for Israel. The fact that no other species of ticks were found on the infected cats and that 36% of the ticks from cat # 3 were positive by PCR for B. galileei sp. nov. supports the suspicion that H. adleri could be the vector of this parasite. Furthermore, the detection of B. galileei sp. nov. DNA in the salivary glands of two ticks that were also positive in their digestive tracts strengthens the assumption that if the parasite has reached the tick’s salivary glands, it may also be transmitted by the saliva to infect a new host. However, just finding the DNA of a pathogen in a tick that has fed on blood does not indicate that this tick is capable of transmitting the pathogen. Therefore, more studies should be carried out to prove the possible capacity of H. adleri to serve as a vector of B. galileei sp. nov.
Piroplasmid spp., which infect domestic cats, are often parasites of wildlife and in particular wild felids in which they cause sub-clinical infection. Such is the case of B. leo, which infects lions, and B. lengau, whose natural host is the cheetah [9, 11, 14]. Another example is C. felis, which infects bobcats sub-clinically and causes a frequently fatal disease in domestic cats in North America [43]. It is likely that B. galileei sp. nov. is also hosted by some wildlife mammals in the Middle East, although it has not been reported in such a host yet.
The clinical presentation of the three cats with B. galileei sp. nov. infection includes typical findings of babesiosis including anemia, thrombocytopenia, extreme lethargy, fever in two cats and severe icterus in one cat. Co-infection with the immunosuppressive virus FIV in cat # 1 may have played a role in increasing its susceptibility to becoming infected or to progressing from infection to clinical disease. Associations between parasitic infections and FIV in populations of domestic cats have been reported previously and include an association between FIV and Leishmania infantum infection and a relationship between FIV and Toxoplasma gondii seropositivity [44, 45]. The immunosuppressive therapy that cat # 2 received with prednisolone and cyclosporine because of suspected IMHA probably contributed to the development of babesiosis in this cat, as the cat was ill before the immunosuppressive treatment began but Babesia organisms were detected in its blood smears only after cyclosporine treatment was begun. It is possible that the cat had been infected previously but only showed parasitemia after immunosuppression or that babesiosis was the initial cause of disease with very low parasitemia that went unnoticed in the earlier blood tests. Cat # 3 was positive for C. M. haemominutum; however, it is likely that this infection did not contribute to the Babesia infection as C. M. haemominutum is considered mostly non-pathogenic and has only rarely been reported as a cause of clinical disease in cats [46]. Altogether, since two cats with babesiosis included in this report were immunosuppressed by additional factors, it is plausible that immune suppression has a role in increasing the susceptibility of cats to clinical B. galileei sp. nov. infection.
The three cats responded well and recovered clinically following injections of imidocarb dipropionate. This agrees with B. galileei sp. nov. classification as Babesia s.s. sp. and the general responsiveness of parasites from this clade to treatment with imidocarb dipropionate, as found in dogs infected with the Babesia s.s. species, e.g. B. vogeli, B. canis and B. rossi [47], and in horses infected with Babesia caballi [48].
Conclusions
This study describes a new Babesia sp. infecting domestic cats in the Middle East, which places phylogenetically in the Babesia s.s. group and is associated with severe clinical disease. Identifying new Babesia spp. and unraveling their clinical impact and response to drug treatment is imperative for veterinary clinicians and parasitologists. More research is warranted to confirm the tick vector of B. galileei sp. nov and to study other potential animal hosts for this pathogen.
Availability of data and materials
All data generated or analyzed during this study are included in this published article. Analyzed nucleotide sequences used for pairwise comparisons and tree construction were submitted to the GenBank database under the accession numbers PP620719-PP620721(18S rRNA),PP620716-PP620718 (ITS), PP624317, PP590338, PP590339 (CytB) and PP624315, PP624316 (HSP70). The holotype was deposited in the National Natural History Collection of the Hebrew University of Jerusalem, Israel, under the accession number HUJINV 1000000006.
Abbreviations
- HSP:
-
Heat shock protein
- ICZN:
-
International code of zoological nomenclature
- ITS:
-
Internal transcribed spacer
- PCR:
-
Polymerase chain reaction
- s.s.:
-
Sensu stricto
References
Lempereur L, Beck R, Fonseca I, Marques C, Duarte A, Santos M, et al. Guidelines for the detection of Babesia and Theileria Parasites. Vector Borne Zoonotic Dis. 2017;17:51–65.
Antunes S, Rosa C, Couto J, Ferrolho J, Domingos A. Deciphering Babesia-vector interactions. Front Cell Infect Microbiol. 2017;7:429.
Davis LJ. On a piroplasm of the Sudanese wild cat (Felis ocreata). Trans R Soc Trop Med Hyg. 1929;22:523–34.
Jackson C, Dunning FJ. Biliary fever (Nuttalliosis) of the cat: a case in the Stellenbosch district. JS Afr Vet Med Assoc. 1937;8:83–8.
McNeil J. Piroplasmosis of the domestic cat. JS Afr Vet Med Assoc. 1937;8:88–90.
Baneth G, Kenny MJ, Tasker S, Anug Y, Shkap V, Levy A, et al. Infection with a proposed new subspecies of Babesia canis, Babesia canis subsp. presentii in domestic cats. J Clin Microbiol. 2004;42:105.
Bosman AM, Venter EH, Penzhorn BL. Occurrence of Babesia felis and Babesia leo in various wild felid species and domestic cats in Southern Africa, based on reverse line blot analysis. Vet Parasitol. 2007;144:33–8.
Bosman AM, Penzhorn BL, Brayton KA, Schoeman T, Oosthuizen MC. A novel Babesia sp. associated with clinical signs of babesiosis in domestic cats in South Africa. Parasit Vectors. 2019;12:138.
Penzhorn BL, Kjemtrup AM, López-Rebollar LM, Conrad PA. Babesia leo n. sp. from lions in the Kruger National Park, South Africa, and its relation to other small piroplasms. J Parasitol. 2001;87:681–5.
Wong SS, Poon RW, Hui JJ, Yuen KY. Detection of Babesia hongkongensis sp. nov. in a free-roaming Felis catus cat in Hong Kong. J Clin Microbiol. 2012;50:2799–803.
Bosman AM, Oosthuizen MC, Peirce MA, Venter EH, Penzhorn BL. Babesia lengau sp. nov., a novel Babesia species in cheetah (Acinonyx jubatus, Schreber, 1775) populations in South Africa. J Clin Microbiol. 2010;48:2703–8.
, Bosman AM, Oosthuizen MC, Venter EH, Steyl JC, Gous TA, Penzhorn BL. Babesia lengau associated with cerebral and haemolytic babesiosis in two domestic cats. Parasit Vectors. 2013;6:128.
Panicker VP, Sreedharannair AK, Narayanan A, George S, Hameed SV. Molecular identification of a novel species, Babesia panickeri sp. nov., from a naturally infected domestic cat of India and its comparison with canine Babesia isolates. Acta Parasitol. 2020;65:913–8.
Penzhorn BL, Oosthuizen MC. Babesia species of domestic cats: molecular characterization has opened pandora’s box. Front Vet Sci. 2020;7:134.
Mudaliar SV, Achary G, Alwar VS. On a species of Babesia in an Indian wild cat (Felis catus). Ind Vet J. 1950;26:391–5.
Panait LC, Hrazdilová K, Ionică AM, Deak G, Chişamera GB, Adam C, et al. Babesia pisicii n. sp. and Babesia canis Infect European Wild Cats, Felis silvestris, in Romania. Microorganisms. 2021;9:1474.
Criado-Fornelio A, Martinez-Marcos A, Buling-Saraña A, Barba-Carretero JC. Presence of Mycoplasma haemofelis, Mycoplasma haemominutum and piroplasmids in cats from southern Europe: a molecular study. Vet Microbiol. 2003;93:307–17.
Alho AM, Lima C, Latrofa MS, Colella V, Ravagnan S, Capelli G, et al. Molecular detection of vector-borne pathogens in dogs and cats from Qatar. Parasit Vectors. 2017;10:298.
Kelly PJ, Köster L, Li J, Zhang J, Huang K, Branford GC, et al. Survey of vector-borne agents in feral cats and first report of Babesia gibsoni in cats on St Kitts. West Indies BMC Vet Res. 2017;13:331.
Pennisi MG, Alongi A, Agnone A, Vitale F, Reale S, Torina A. Cats as reservoir of Babesia microti. Parassitologia. 2007;49:100.
Akram IN, Parveen T, Abrar A, Mehmood AK, Iqbal F. Molecular detection of Babesia microti in dogs and cat blood samples collected from Punjab (Pakistan). Trop Biomed. 2019;36:304–9.
Oosthuizen MC, Zweygarth E, Collins NE, Troskie M, Penzhorn BL. Identification of a novel Babesia sp. from a sable antelope (Hippotragus niger Harris, 1838). J Clin Microbiol. 2008;46:2247–51.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol. 2018;35:1547–9.
Madeira F, Pearce M, Tivey ARN, Basutkar P, Lee J, Edbali O, et al. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022;50:W276–9.
Feldman-Muhsam B. A note on East Mediterranean species of the genus Haemaphysalis. B Res Counc Israel. 1951;1:96–107.
ICZN. International commission on zoological nomenclature: amendment of articles 8, 9, 10, 21 and 78 of the international code of zoological nomenclature to expand and refine methods of publication. Bull Zool Nomencl. 2012;69:161–9.
Solano-Gallego L, Baneth G. Babesiosis in dogs and cats—expanding parasitological and clinical spectra. Vet Parasitol. 2011;181:48–60.
Potgieter FT. Chemotherapy of Babesia felis infection: efficacy of certain drugs. J S Afr Vet Assoc. 1981;52:289–93.
Jacobson LS, Schoeman T, Lobetti RG. A survey of feline babesiosis in South Africa. J S Afr Vet Assoc. 2000;71:222–8.
Orkun Ö. Description of a novel Babesia sp. genotype from a naturally infected Eurasian lynx (Lynx lynx) in Anatolia, Turkey, with remarks on its morphology and phylogenetic relation to other piroplasmid species. Ticks Tick Borne Dis. 2022;13:102026.
Orkun Ö, Çakmak A, Nalbantoğlu S, Karaer Z. Molecular detection of a novel Babesia sp. and pathogenic spotted fever group rickettsiae in ticks collected from hedgehogs in Turkey: Haemaphysalis erinacei, a novel candidate vector for the genus Babesia. Infect Genet Evol. 2019;69:190–8.
Jalovecka M, Sojka D, Ascencio M, Schnittger L. Babesia life cycle—when phylogeny meets biology. Trends Parasitol. 2019;35:356–68.
Schmid N, Deplazes P, Hoby S, Ryser-Degiorgis MP, Edelhofer R, Mathis A. Babesia divergens-like organisms from free-ranging chamois (Rupicapra r. rupicapra) and roe deer (Capreolus c. capreolus) are distinct from B. divergens of cattle origin—an epidemiological and molecular genetic investigation. Vet Parasitol. 2008;154:14–20.
Oyamada M, Davoust B, Boni M, Dereure J, Bucheton B, Hammad A, et al. Detection of Babesia canis rossi, B. canis vogeli, and Hepatozoon canis in dogs in a village of eastern Sudan by using a screening PCR and sequencing methodologies. Clin Diagn Lab Immunol. 2005;12:1343–6.
Shabangu N, Penzhorn BL, Oosthuizen MC, Vorster I, van Schalkwyk OL, Harrison-White RF, et al. A shared pathogen: Babesia rossi in domestic dogs, black-backed jackals (Canis mesomelas) and African wild dogs (Lycaon pictus) in South Africa. Vet Parasitol. 2021;291:109381.
Penzhorn BL, Vorster I, Harrison-White RF, Oosthuizen MC. Black-backed jackals (Canis mesomelas) are natural hosts of Babesia rossi, the virulent causative agent of canine babesiosis in sub-Saharan Africa. Parasit Vectors. 2017;10:124.
Ben-Moshe N, Rosensaft M, Iwamura T. Land-use changes interact with geology to facilitate dispersal of the rock hyrax (Procavia capensis) and leishmaniasis across Israel and the West Bank. Ecol Evol. 2023;13:e9915.
Guglielmone AA, Nava S, Robbins RG. Geographic distribution of the hard ticks (Acari: Ixodida: Ixodidae) of the world by countries and territories. Zootaxa. 2023;5251:1–274.
Azmi K, Ereqat S, Nasereddin A, Al-Jawabreh A, Baneth G, Abdeen Z. Molecular detection of Theileria, Babesia, and Hepatozoon spp. in ixodid ticks from Palestine. Ticks Tick Borne Dis. 2016;7:734–41.
Shubber HWK, Al-Hassani NAW, Kadhim M. Ixodid ticks diversity in the middle and south of Iraq. Int J Recent Sci Res. 2014;5:1518–23.
Guglielmone AA, Robbins RG, Apanaskevich DA, Petney TN, Estrada-Peña A, Horak IG. The hard ticks of the World (Acari: Ixodida: Ixodidae). Dordrecht: Springer; 2014.
Salant H, Mumcuoglu KY, Baneth G. Ectoparasites in urban stray cats in Jerusalem, Israel: differences in infestation patterns of fleas, ticks and permanent ectoparasites. Med Vet Entomol. 2014;28:314–8.
Cohn LA. Cytauxzoonosis. Vet Clin North Am Small Anim Pract. 2022;52:1211–24.
Iatta R, Furlanello T, Colella V, Tarallo VD, Latrofa MS, Brianti E, et al. A nationwide survey of Leishmania infantum infection in cats and associated risk factors in Italy. PLoS Negl Trop Dis. 2019;13:e0007594.
Chi X, Fang K, Koster L, Christie J, Yao C. Prevalence of feline immunodeficiency virus and Toxoplasma gondii in feral cats on St. Kitts West Indies. Vet Sci. 2021;8:16.
Tasker S. Hemotropic Mycoplasma. Vet Clin North Am Small Anim Pract. 2022;52:1319–40.
Baneth G. Antiprotozoal treatment of canine babesiosis. Vet Parasitol. 2018;254:58–63.
Onyiche TE, Suganuma K, Igarashi I, Yokoyama N, Xuan X, Thekisoe O. A review on equine piroplasmosis: epidemiology, vector ecology, risk factors, host immunity, diagnosis and control. Int J Environ Res Public Health. 2019;16:1736.
Blaschitz M, Narodoslavsky-Gfoller M, Kanzler M, Stanek G, Walochnik J. Babesia species occurring in Austrian Ixodes ricinus ticks. Appl Environ Microbiol. 2008;74:4841–6.
Matjila PT, Leisewitz AL, Oosthuizen MC, Jongejan F, Penzhorn BL. Detection of a Theileria species in dogs in South Africa. Vet Parasitol. 2008;157:34–40.
Tabar MD, Altet L, Francino O, Sánchez A, Ferrer L, Roura X. Vector-borne infections in cats: molecular study in Barcelona area (Spain). Vet Parasitol. 2008;151:332–6.
Tian Z, Jin L, Zheng J, Xie J, Shen H, Yin H, et al. Phylogenetic analysis of Babesia species in China based on cytochrome b (COB) gene. Infect Genet Evol. 2023;13:36–40.
Almeida AP, Marcili A, Leite RC, Nieri-Bastos FA, Domingues LN, Martins JR, et al. Coxiella symbiont in the tick Ornithodoros rostratus (Acari: Argasidae). Ticks Tick Borne Dis. 2012;3:203–6.
Peleg O, Baneth G, Eyal O, Inbar J, Harrus S. Multiplex real-time qPCR for the detection of Ehrlichia canis and Babesia canis vogeli. Vet Parasitol. 2010;173:292–9.
Acknowledgements
This paper has been sponsored by Elanco Animal Health in the framework of the CVBD® World Forum Symposium. The authors thank all the veterinary clinicians who have treated the infected cats.
Funding
The study was funded by the author’s internal resources with no external funding.
Author information
Authors and Affiliations
Contributions
GB conceived the study. GB performed the morphologic characterization of parasites. YNB and GB analyzed the genetic data. AD, IA, SA and YS analyzed the blood smears of the cats and their clinical data, HS and DS dissected the ticks and performed PCR on ticks and blood samples, and KYM identified the ticks morphologically. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
13071_2024_6371_MOESM1_ESM.pptx
Additional file1 Table 1. Identity matches (%) of gene and loci DNA sequences from the Israeli cats compared to relevant close spp. and genotypes (% cover in brackets)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Baneth, G., Nachum-Biala, Y., Dvorkin, A. et al. Description of Babesia galileei sp. nov. A piroplasmid species causing severe disease in domestic cats. Parasites Vectors 17, 297 (2024). https://doi.org/10.1186/s13071-024-06371-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06371-w