
Abstract
Volatile organic compounds (VOCs) are chemicals emitted as products of cell metabolism, which reflects the physiological and pathological conditions of any living organisms. These compounds play a key role as olfactory cues for arthropod vectors such as mosquitoes, sand flies, and ticks, which act in the transmission of pathogens to many animal species, including humans. Some VOCs may influence arthropod behaviour, e.g., host preference and oviposition site selection for gravid females. Furthermore, deadly vector-borne pathogens such as Plasmodium falciparum and Leishmania infantum are suggested to manipulate the VOCs profile of the host to make them more attractive to mosquitoes and sand fly vectors, respectively. Under the above circumstances, studies on these compounds have demonstrated their potential usefulness for investigating the behavioural response of mosquitoes, sand flies, and ticks toward their vertebrate hosts, as well as potential tools for diagnosis of vector-borne diseases (VBDs). Herein, we provide an account for scientific data available on VOCs to study the host seeking behaviour of arthropod vectors, and their usefulness as attractants, repellents, or tools for an early diagnosis of VBDs.
Graphical Abstract

Similar content being viewed by others

Background
Mosquitoes, ticks, and sand flies are among the most important vectors of pathogens worldwide, being responsible for the transmission of infectious agents such as viruses, bacteria, nematodes, and protozoa of human and veterinary concern [1,2,3]. The mechanisms used by these arthropod vectors to locate susceptible vertebrate hosts include visual, olfactory, acoustic, and thermal stimuli. Such mechanisms influence the host seeking behaviour of individual arthropods according to the species characteristics and biology, even within the same taxonomic group. For example, many species of ticks (e.g., Dermacentor spp., Ixodes spp., Rhipicephalus spp.) have a questing behaviour, in which they climb the vegetation and wait for the appropriate host, whereas other species such as Hyalomma spp. are considered hunting ticks, since they actively chase for vertebrate hosts [4,5,6]. Another example is the South American sand fly species Lutzomyia (Lutzomyia) longipalpis, in which females are attracted not only by the host odour, but also by aggregation of sex pheromones of sand fly males that arrive first to the host [7,8,9,10].
Among the cues used by arthropods, the olfactory stimuli have been the most studied, being crucial to identify the preferential hosts, principally for vectors with a high host specificity [11, 12]. Volatile organic compounds (VOCs) are a group of chemicals in a gaseous phase and with high vapor pressure, which are delivered as products of the cell metabolism, reflecting the physiological and pathological conditions of the organisms [13]. The US Environmental Protection Agency (EPA; https://www.epa.gov/) define VOCs as compounds of carbon, excluding gases that participate in atmospheric photochemical reactions, such as carbon monoxide, carbon dioxide (considered an universal attractant for arthropods), carbonic acid, metallic carbides or carbonates, and ammonium carbonate. Importantly, some VOCs affect the behaviour of arthropod vectors, which indicates that studying these molecules is crucial to understand the host preference of mosquitoes, sand flies, and ticks [14,15,16,17]. In addition, such compounds may be applied to improve the trapping methods available for capturing these arthropods [11, 18, 19].
Several VOCs are emitted by vertebrate hosts. However, a relatively small amount of them have an influence on the arthropod behaviour, being defined as ‘allelochemicals’ [11]. The latter include kairomones (attractants) and allomones (repellents) [11]. Kairomones may be applied as selective tools for studying population abundance, surveillance of invasive species and vector-borne pathogens, as well as for predicting pathogen outbreaks [20, 21]. Conversely, the repellent effect of allomones emitted by fungi, bacteria, yeasts, plants, and mammals represents a potential tool for controlling arthropod vectors [22,23,24,25]. The presence of VOCs has also been explored in the diagnosis of cancer and infectious diseases in humans and animals [13, 26,27,28], as well as for managing agricultural pests [21, 29, 30].
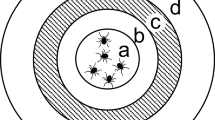
In general, the host preference of blood feeding arthropod vectors is often investigated through the analysis of blood meal by molecular methods [31,32,33,34,35,36,37,38], or by host-choice experiments in laboratory conditions [39,40,41,42]. However, while the above methods are useful to investigate the host preference, they are limited in assessing the factors influencing the attracting capacity of the hosts. Conversely, the interaction between arthropod vectors and vertebrate hosts could be assessed through olfactory cues by determining the attractiveness or repellent effects of VOCs emitted by the hosts (Fig. 1). This is usually investigated at laboratory scale, collecting VOCs from the host through classical extraction methods; this can be done relying to adsorption materials used in solid-phase micro extraction (SPME) and direct-contact sorptive extraction (DCSE). Then, the collected compounds are subsequently identified through gas chromatography (GC), nuclear magnetic resonance (NMR) and mass spectrometry (MS) approaches [43]. Once identified, electroantennography (EAG/EAD; Fig. 2), also coupled with GC and MS (i.e., GC-EAD and GC–MS-EAD) can be used to evaluate which molecules of a VOCs’ bouquet are really perceived by a given arthropod vector. Electroantennography-active VOCs can be therefore tested in behavioural experiments such as olfactometer assays (e.g., Y-tube olfactometer and flight tunnel; Fig. 3) and in field/semi-field tests to shed light on their ecological role and potential use in vector monitoring and management.

Host-borne volatile organic compounds (VOCs) influencing the olfactory behaviour of mosquitoes, sand flies, and ticks

Electroantennography for the evaluation of sand flies chemoreceptivity to VOCs

Y-tube olfactometer used for behavioural experiments with sand flies and other arthropods
Several studies on VOCs and their role as attractants or repellents have been performed for different arthropod vectors including mosquitoes [22, 42, 44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60], ticks [61,62,63,64,65,66,67], phlebotomine sand flies [68,69,70,71,72,73,74,75,76], triatomines [12], and tsetse flies [77, 78]. The latter includes species of the genus Glossina, which are exclusively found in Africa, where field and laboratory studies have been performed and reviewed elsewhere [78]. In the present review, we analysed current knowledge on the host-borne VOCs acting as attractants or repellents for worldwide distributed vectors of major medical and veterinary importance, such as mosquitoes, phlebotomine sand flies, and ticks. Moreover, we discuss the applied implications of these compounds, including their potential usefulness for an early diagnosis of vector-borne diseases (VBDs).
Volatile organic compounds and mosquitoes
Mosquitoes are the most important arthropod vectors worldwide transmitting many deadly pathogens of human and veterinary concern, such as chikungunya, dengue, zika, West Nile virus, Rift Valley fever virus, Japanese encephalitis virus, Plasmodium spp., and Dirofilaria spp. [79,80,81]. Studies on the repellent and attraction effects of these arthropods using olfactory cues have been mostly assessed for Anopheles spp., Aedes aegypti, and Culex spp., which are vectors of deadly human pathogens [45, 47, 51, 82,83,84,85,86,87]. For example, the repellent effect of some VOCs isolated from cattle has been showed for Ae. aegypti, Culex quinquefasciatus and Anopheles coluzzii [22]. In the above study, unsaturated aldehydes (Table 1) extracted from the hair of Holstein heifer cattle showed a repellent effect against these insects under laboratory and field conditions [22]. In laboratory assays, the four-component blends (i.e., (E)-2-hexenal, (E)-2-heptenal, (E)-2-octenal, and (E)-2-nonenal) repelled females with an effect similar to that of commercial repellents that were used as control (i.e., N, N-diethyl-m-toluamide [DEET], ethyl butylacetylaminopropionate [IR3535], p-menthane-3, 8-diol [PMD], icaridin, and d-allethrin). Moreover, under field conditions, traps treated with the four-component blend captured significantly lesser mosquitoes in a site (average reduction of 84.8%) in comparison with the control traps (baited with CO2 and host kairomone), whereas in the second site the average reduction was of 55.5% [22].
The olfactory preferences of mosquitoes may differ according to the species, being an important cue for understanding the host choice and behaviour of these arthropod vectors [88]. Indeed, the attractive effect of VOCs has been assessed for Ae. aegypti, Cx. quinquefasciatus, and Cx. nigripalpus, demonstrating that a higher number of compounds associated with bovine and avian blood (Table 1) were attractive for Ae. aegypti and, to a lesser extent for the other two species (Table 1) [89]. Again, lactic acid, a compound present in vertebrate hosts, was demonstrated to be attractive to Ae. aegypti [89, 90] and Cx. quinquefasciatus [89] but did not elicit any behavioural response for Anopheles gambiae mosquitoes [45, 91]. Nevertheless, the attractiveness of carbon dioxide to An. gambiae was significantly increased when combined with lactic acid in experimental assays [83], suggesting that although this compound is not attractive on its own, it may potentialize the response of other compounds present on the skin [83]. Indeed, the synergism among VOCs is an important factor to be considered when studying the role of these compounds in the behaviour of arthropod vectors, as demonstrated for An. gambiae mosquitoes, for which ammonia, lactic acid, and a mixture of carboxylic acids (Table 1) were tested alone and in combination to assess the olfactory responses of this mosquito species [45, 47, 51]. While ammonia was found to be attractive, the mixture of carboxylic acids was repellent, whereas lactic acid did not attract the mosquitoes. However, when ammonia, carboxylic acids and lactic acid were combined, they were significantly more attractive for An. gambiae mosquitoes, than ammonia alone [45, 47, 51]. In addition, the component blends from the above-mentioned studies became even more attractive to An. gambiae by the addition of butan-1-amine (associated with CO2), and 3-methyl-1-butanol [92]. Similarly, nonanal and CO2 synergized in field experiments in which traps containing the two compounds captured significantly more Cx. quinquefasciatus mosquitoes than traps baited only with CO2 [86], therefore suggesting that host volatiles may have different effects on the host seeking behaviour of arthropods, according to their abundance and combination.
Though not emitted by hosts, some VOCs have also been associated with the selection of oviposition sites by An. gambiae, An. arabiensis and Ae. aegypti gravid females [19, 93, 94], demonstrating that compounds produced in water bodies containing mosquito larvae may influence the selection of oviposition sites [93]. For example, dimethyl disulfide and dimethyl trisulfide detected in emanations from water containing fourth instar larvae (L4) acted as oviposition repellents, also causing retention of eggs within the gravid females [93]. On the other hand, VOCs (i.e., nonane and 2,4-pentanedione) identified in emanations containing first instar larvae (L1) were considered attractants, being the presence of these two compounds in water related with significantly higher oviposition rates when compared to untreated water in semi-field trials [93]. The findings above indicate that the selection of the oviposition site by mosquitoes may be mediated by chemical compounds, and that the presence of larvae instars may impact on the choice of oviposition site by female mosquitoes, with L1 causing an attractive effect, and L4 repellent and inhibition outcomes on oviposition [93]. The oviposition selection sites of Ae. aegypti are also affected by VOCs produced in water bodies containing immature stages [19]. Indeed, by using blends of bioactive VOCs identified in eggs (i.e., 2,4-dimethylhept-1-ene; 2,6-dimethyl-7-octen-2-ol; camphor; decanal), in second instar larvae (L2) (i.e., 4-cyanocyclohexene; (E)-2-octenal; nonanal; decanal 4-(2-methylbutan-2-yl) phenol), in L4 (i.e., 2,4-dimethylhept-1-ene; (E)-2-heptanal; nonanal; camphor; (E)-2-nonenal; (E)-2-decenal; 4-(2-methylbutan-2-yl) phenol), and in pupae exuviae (4-cyanocyclohexene; 2,6-dimethyl-7-octen-2-ol; nonanal) of Ae. aegypti it was demonstrated that gravid females were more stimulated to lay eggs in response to VOCs produced in water bodies containing late-stage larvae [19]. All the above suggested that VOCs may signalize the density of conspecific aquatic stages, influencing on the oviposition site choice by gravid females, which may avoid laying eggs on sites in which the odour indicates high density of conspecific competitors [19].
Volatile organic compounds and sand flies
Sand flies are involved in the transmission of viruses (e.g., Phleboviruses: Naples, Sicilian and Toscana viruses; [95]) bacteria (e.g., Bartonella bacilliformis; [96]), and most importantly, of protozoa of the genus Leishmania to humans and animals [97]. The latter is transmitted by sand flies of the genus Phlebotomus spp. and Lutzomyia spp. in the old and new world, respectively [98]. Studies on the interaction among sand fly species and their hosts have been performed worldwide, and they mainly focused on the evaluation of blood meal by molecular analysis in these insects [36, 99,100,101]. Besides the detection of blood meal, the interaction of sand flies with vertebrate hosts may also be assessed by determining the attractiveness or repellent effect of these insects by VOCs, though few investigations are available. Lutzomyia species (members of subgenera Lutzomyia (Lu.) and Nyssomyia (N.)) were tested with VOCs from humans, dogs, and foxes [68, 69, 72,73,74]. Of the 42 VOCs identified from the hair of 33 human male volunteers in Brazil, seven compounds (i.e., Phenylacetaldehyde; 6-methylhept-5-en-2-one; tetradecane; pentadecane; hexadecane; nonadecane; icosane) were tested for the attractiveness of field captured Lutzomyia spp. (i.e., n = 420 specimens; 75.4% Lutzomyia (N.) intermedia; 4.5% Lutzomyia migonei; 2.8% Lutzomyia (N.) whitmani; 0.9% Lutzomyia fischeri; 0.2% Lutzomyia shannoni; and 16.2% damaged sand flies non-identified) [72]. In the study above, four VOCs were demonstrated to induce a significant activation response in sand flies (i.e., phenylacetaldehyde; 6-methylhept-5-en-2-one; pentadecane; icosane), while three of them (i.e., Phenylacetaldehyde; 6-methylhept-5-en-2-one; icosane) were attractive to female sand flies [72]. Lutzomyia (Lu.) longipalpis also had a behavioural response to VOCs found on humans, with the compounds octenol, nonanol and heptanol activating and attracting male and female sand flies in behavioural experiments [73]. Additionally, Lu. (Lu.) longipalpis and Lu. (N.) intermedia were attracted to octenol and a synthetic human odour BG-Mesh Lure™ (BGML—lactic acid, caproic acid and ammonia) in field experiments using CDC light traps baited with these compounds [69]. The attractiveness of VOCs from the skin of different human individuals for Lu. (Lu.) longipalpis has also showed that there is a significant variation in individual attractiveness of human subjects to sandflies. However, the attractants were not identified in that study [68].
Volatile organic compounds emitted by L. infantum infected dogs were also assessed to test the responses of Lu. (Lu.) longipalpis [74], demonstrating that, when tested individually, octanal, nonanal, decanal and heptadecane activated and attracted male sand flies, whereas only decanal and nonanal showed an activation response of females [74]. Moreover, the blend of octanal, decanal and heptadecane increased both the activation and attraction behaviour in males, while the mixture of octanal and decanal acted only in the activation [74]. The higher attractiveness of these VOCs to male Lu. (Lu.) longipalpis may be related to the biology of this sand fly species, since males are also known to be attracted by the host odours, and under field studies they have been demonstrated to arrive first at the host sites than females [9]. Furthermore, females of this species have been suggested to be more attracted to the host volatile compounds when in combination with male-produced sex pheromone, a strategy for easing blood meal [8, 10]. In contrast to VOCs released from infected dogs, Lu. (Lu.) longipalpis females were demonstrated to be significantly more attracted than males in response to skin emanations from humans under experimental conditions. However, the attractive factors were not identified, advocating further experiments to confirm this observation [102]. In wildlife, a single study on VOCs from foxes (Vulpes vulpes) demonstrated that female Lu. (Lu.) longipalpis were attracted by the natural odour, and by a synthetic blend mimicking the odour of this host, as well as by individual compounds (i.e., benzaldehyde; 4-hydroxy-4-methyl-2-pentanone; 4-methyl-2-pentanone) identified in the fox odour [103].
The attraction of VOCs has also been tested for the species Lu. (N.) neivai, a vector of Leishmania (Viannia) brasiliensis in South America [70, 71], in which octenol [70, 71], pentanol, hexanol, and octanol [71] acted both in activating and attracting field captured female sand flies in behavioural experiments using wind tunnel. The above findings suggest that research on the attractiveness of sand flies by VOCs from mammalian hosts are relevant to assess the host preference of these insects, as well as to enhance the efficiency of conventional traps used for monitoring and controlling phlebotomine sand flies.
The effect of VOCs in sand fly selection of oviposition sites has also been studied for Phlebotomus papatasi, in which volatile semiochemicals produced by bacterial isolates (i.e., Actinobacteria, Bacteroides, Firmicutes, and Proteobacteria) had an attractive response for gravid females at low doses (i.e., 107 cells per ml), and a repellent response at higher concentrations (109 cells per ml) [104]. Though, the identification of the attractive VOCs produced by these bacteria was not completed, data above suggest that the attractiveness of Ph. papatasi to those VOCs is dose dependent.
Volatile organic compounds and ticks
Ticks are important vectors of pathogens of zoonotic concern worldwide, being able to transmit infectious agents to humans, domestic animals, and wildlife [80, 105]. These arthropods are responsible for significant economic losses to the livestock industry, which are directly related with the decrease in animal production and, indirectly, with the transmission of pathogens such as Anaplasma spp., Babesia spp., and Theileria spp. [105, 106]. For this reason, studying the host preference, the pathogen transmission times [107] as well as the mechanisms behind the resistance of some breeds to ticks, is paramount to improve control measures against these ectoparasites.
Studies on VOCs have been employed in the assessment of the attractive and repellent effects [108] of these compounds for ticks (Table 1). For example, the attraction of some volatiles (e.g., methyl salicylate; benzoic acid; ammonium hydroxide; salicylic acid) has been tested in nymphs of Amblyomma sculptum ticks collected on naturally infected horses from Brazil [108]. These data suggested that such compounds may be used as trapping methods for A. sculptum ticks [108]. The same tick species has also been demonstrated to be repelled by benzaldehyde and isobutyric acid, which, in turn, are compounds previously detected in tick resistant mammalian hosts [63, 108]. Moreover, in another study performed on donkeys, the (E)-2-octenal was demonstrated to act as a repellent for this tick species [109]. Of notice, the above compound was found in sebum extracts of donkeys, but not of horses, being the latter less resistant to the infestation by A. sculptum ticks. Such data suggested that the compound (E)-2-octenal could be used as a repellent for reducing A. sculptum populations in susceptible hosts [109].
In dogs, VOCs have also been assessed in breeds resistant to Rhipicephalus sanguineus sensu lato ticks to detect compounds with potential repellent effect [63, 66]. For example, beagles have been demonstrated to be less susceptible to tick infestation when compared to English cocker spaniel breed [110], resulting in a lower attractiveness of beagles’ odour to R. sanguineus s.l. [111]. The observation above was further investigated through the isolation of VOCs from the skin of these two breeds, in which the gas chromatography analysis detected a greater number of compounds on odour extracts collected from beagles than in the extracts from cocker spaniel [63]. In addition, among the compounds detected in beagles, three of the most abundant (i.e., 2-hexanone; benzaldehyde; undecane), showed repellent effect at the behavioural assays [63]. These data confirmed that beagles produce a larger amount of potentially natural repellents against this tick species [63]. This was further supported by another study, in which the VOCs produced by the above-mentioned breeds were compared with the ones produced by the miniature pinscher, which is a putative tick-resistant breed [66]. In the aforementioned study, 2-hexanone and benzaldehyde were detected in the odour extracts of all three breeds, with 6-methyl-5-hepten-2-one (sulcatone), and 1,2,4-trimethylbenzene being more abundant in miniature pinscher dogs. In addition, the overall abundance of these compounds was significantly higher in beagles and miniature pinschers than in cocker spaniels [66]. In the behavioural assays, the compounds 6-methyl-5-hepten-2-one and 1,2,4-trimethylbenzene were repellent for ticks, supporting that both may play a role in the resistance of these breeds to R. sanguineus s.l. ticks [66].
The attractiveness of VOCs from dogs has also been assessed in a study identifying compounds that may be involved in host attraction and localization by R. sanguineus s.l. [67]. In the above study isovaleric acid, hexanal, heptanal, and sulcatone significantly stimulated the olfactory receptors of female ticks using the single sensillum recording technique [67], while in the Y-tube olfactometer bioassays the ticks were attracted only to isovaleric acid and to a blend of hexanal, heptanal, and isovaleric acid [67]. Overall, those data suggested that the latter compounds may be involved in the host location by R. sanguineus s.l. [67].
Volatile organic compounds and vector-borne pathogens
The association of VOCs with pathogens transmitted by arthropods have been assessed for Plasmodium falciparum [112,113,114,115] and for L. infantum [75, 76, 116, 117], transmitted by mosquitoes and sand flies, respectively (Fig. 1). Some VOCs attractive for mosquitoes have been suggested to be produced by P. falciparum as a strategy of this parasite for facilitating its transmission [113, 118,119,120,121,122,123]. For example, the compound (E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate (HMBPP) produced by P. falciparum has been suggested to increase the production and release of An. gambiae attractants from blood of malaria infected individuals, increasing the likelihood of vector bite in infected humans [121]. Indeed, VOCs already known to attract mosquitoes (i.e., α-pinene and 3-carene) were detected at significantly higher levels in the breath of malaria infected children as compared with uninfected ones [123]. Nevertheless, it was later demonstrated that HMBPP does not alter the levels of the mosquito attractant α-pinene, indicating that the mechanism used by Plasmodium spp. parasites to increase attractiveness of infected individuals remains unclear [124]. Moreover, under field conditions, a blend of VOCs (i.e., α-pinene; β-pinene; limonene; octanal; nonanal; decanal) reflecting the odours induced by P. falciparum gametocyte parasitizing the red blood cells, attracted 2.5 times more Anopheles spp. mosquitoes than non-baited traps, indicating that the parasite could manipulate the host-seeking behaviour of malaria vectors [114]. Similarly, experimental trials demonstrated that the aldehydes heptanal, octanal, and nonanal, which are produced in larger amount by infected individuals, enhanced an increased attractiveness of An. gambiae vectors [125], consequently affecting the transmission of the parasite.
The attraction of Anopheles stephensi mosquitoes to mice infected by Plasmodium chabaudi has also been demonstrated in an experimental study, which also showed a clear difference in the VOCs composition between infected and non-infected individuals, identifying 11 VOCs (i.e., N,N-dibutylformamide; tridecane; 2-pyrrolidone; 3-methyl-2-buten-1-ol; 3-methyl butanoic acid; 2-hexanone; benzaldehyde; and 4 unidentified compounds) as indicators of infection during the chronic phase [126]. In addition, 3-methyl butanoic acid, 2-methyl butanoic acid, hexanoic acid, and tridecane were attractive for the mosquitoes, whereas benzothiazole, which decreased in infected mice, caused a significant reduction in mosquito attraction [126].
Volatile organic compounds of symptomatic and asymptomatic malaria human patients has been used as biomarkers for the diagnosis of this disease, demonstrating that some compounds (e.g., 4-hydroxy-4-methylpentan-2-one; nonanal; toluene; 2-ethylhexan-1-ol; ethylbenzene; ethylcyclohexane; propylcyclohexane; hexane) were predictors of malaria infection [127]. Importantly, these volatiles presented a sensitivity of 100% in patients without clinical signs, suggesting their potential in screening of malaria infected individuals [127].
Similarly, VOCs production profile of L. infantum infected vs non-infected dogs has been assessed for the identification of possible biomarkers for the diagnosis of leishmaniasis [116]. In the above study, some VOCs (i.e., octanal; nonanal; undecane; β-hydroxyethylphenyl ether; decanal; tetradecane; nonyl cyclopentane; 8-pentadecanone; heptadecane; 2-ethylhexyl-salicylate) were identified being selectively expressed with different profiles in infected (with and without clinical signs) vs non-infected dogs [116]. In particular, β-hydroxyethylphenyl ether, nonanal, heptadecane, 2-ethylhexyl-salicylate, decanal, and octanal showed a significant variation in production, which could be regarded as potential biomarkers of infection by L. infantum in dogs [116].
The differences in VOCs profiles from L. infantum infected and non-infected individuals also influenced the attractiveness of sand flies, under experimental studies both in dogs [75], and hamsters for Lu. (Lu.) longipalpis [117]. The above studies demonstrated that male and female of Lu. (Lu.) longipalpis were equally attracted to the odour of uninfected dogs as compared to a solvent control [75]. Conversely, blood seeking females were significantly more attracted than males when exposed to the odour of L. infantum infected dogs [75]. Similarly, female Lu. (Lu.) longipalpis were significantly more attracted to Golden Hamsters (Mesocricetus auratus) experimentally infected by L. infantum, than to uninfected individuals [117]. Finally, the attractiveness of infected vs non-infected dogs to sand flies was also assessed for Ph. perniciosus and Ph. perfiliewi under experimental and field conditions, demonstrating that male and female insects were significantly more attracted to dogs infected by L. infantum than to non-infected [76]. Nevertheless, the VOCs profile of the animals in the above study was not assessed. Overall, this scientific evidence supports the concept that the parasite may modify the profile of VOCs on the host to make it more attractive to sand fly vectors.
Conclusions
The olfactory cues of arthropod vectors have been mainly studied for mosquitoes and ticks, with the identification of attractive and repellent VOCs emanated by their vertebrate hosts, and in a lesser extent for phlebotomine sand flies in which up to date, only attractive VOCs have been assessed (Table 1). The potential of these compounds to influence the behaviour of these arthropods have been assessed in several studies, being proved as a key factor in the host choice by ticks, mosquitoes, and sand flies. These investigations are of great importance to better understand the host preference of some vectors by assessing the VOCs production profile, which differ not only according with the different host species, but also within hosts of the same species, affecting the individual susceptibility to arthropod bites [128, 129]. In addition, such studies are also useful for improving strategies of monitoring and control, particularly under the current global warming scenario, which also affects arthropod chemical ecology [130].
The differences in the production of VOCs in infected vs non-infected hosts, indicate that vector-borne pathogens such as P. falciparum and L. infantum may modify the profile of these compounds in infected hosts to make them more attractive for blood seeking vectors, which consequently increases the odds of transmission of VBDs. Such intriguing ecological hypothesis remains less explored for ticks and tick-borne pathogens. Under the above circumstances, the identification of VOCs biomarkers produced by infected host cells could be a potential tool for the diagnosis of vector borne pathogens. Nevertheless, the complexity of the skin microenvironment of vertebrate hosts should be considered when evaluating the VOCs production. For example, infections by skin dwelling microorganisms (e.g., bacteria, dermatophytes, mites), may influence the composition of VOCs on the host, potentially increasing the attraction to arthropod vectors [131, 132]. The above may be also true for non-infectious diseases such as cancer, which are also known to alter the VOCs profile of the hosts [28]. However, whether the conditions above increase the risk of infection by vector borne pathogens should be further investigated.
Volatile organic compounds may also be useful for improving the mass rearing of arthropod vectors by using compounds that mimic the odour of the hosts, which could be applied in artificial membranes to increase the acceptability of arthropods to artificial feeding systems; therefore, avoiding the use of live animals in the mass rearing of blood feeding arthropods. Finally, another future perspective for the use of VOCs is the potential integration between compounds emitted by hosts and vector pheromones, which may be used as a strategy for the monitoring of arthropod vectors through the increase in the attractiveness of trapping methods. All the applications above could be potentially useful towards reducing the impact of VBDs of medical and veterinary relevance.
Availability of data and materials
Not applicable.
Abbreviations
- VOCs:
-
Volatile organic compounds
- VBDs:
-
Vector-borne diseases
- CO2 :
-
Carbon dioxide
- SPME:
-
Solid-phase micro extraction
- DCSE:
-
Direct-contact sorptive extraction
- GC:
-
Gas chromatography
- NMR:
-
Nuclear magnetic resonance
- MS:
-
Mass spectrometry
- EAG:
-
Electroantennography
- EAD:
-
Electroantennographic detection
- HMBPP:
-
(E)-4-hydroxy-3-methyl-but-2-enyl pyrophosphate
References
Otranto D, Dantas-Torres F. Canine and feline vector-borne diseases in Italy: current situation and perspectives. Parasit Vectors. 2010;3:2. https://doi.org/10.1186/1756-3305-3-2.
Di Giovanni F, Wilke ABB, Beier JC, Pombi M, Mendoza-Roldan JA, Desneux N, et al. Parasitic strategies of arthropods of medical and veterinary importance. Entomol Generalis. 2021;41:511–22. https://doi.org/10.1127/entomologia/2021/1155.
Bezerra-Santos MA, Dantas-Torres F, Benelli G, Otranto D. Emerging parasites and vectors in a rapidly changing world: from ecology to management. Acta Trop. 2023;238:106746. https://doi.org/10.1016/j.actatropica.2022.106746.
Anderson JF, Magnarelli LA. Biology of ticks. Infect Dis Clin North Am. 2008;22:195–215. https://doi.org/10.1016/j.idc.2007.12.006.
Benelli G, Romano D, Rocchigiani G, Caselli A, Mancianti F, Canale A, et al. Behavioral asymmetries in ticks–lateralized questing of Ixodes ricinus to a mechatronic apparatus delivering host-borne cues. Acta Trop. 2018;178:176–81. https://doi.org/10.1016/j.actatropica.2017.11.024.
Leal B, Zamora E, Fuentes A, Thomas DB, Dearth RK. Questing by tick larvae (Acari: Ixodidae): a review of the influences that affect off-host survival. Ann Entomol Soc Am. 2020;113:425–38. https://doi.org/10.1093/aesa/saaa013.
Bray DP, Hamilton JG. Host odor synergizes attraction of virgin female Lutzomyia longipalpis (Diptera: Psychodidae). J Med Entomol. 2007;44:779–87. https://doi.org/10.1603/0022-2585(2007)44[779:hosaov]2.0.co;2.
Nigam Y, Ward RD. The effect of male sandfly pheromone and host factors as attractants for female Lutzomyia longipalpis (Diptera: Psychodidae). Physiol Entomol. 1991;16:305–12. https://doi.org/10.1111/j.1365-3032.1991.tb00569.x.
Kelly DW, Dye C. Pheromones, kairomones and the aggregation dynamics of the sandfly Lutzomyia longipalpis. Anim Behav. 1997;53:721–31. https://doi.org/10.1006/anbe.1996.0309.
Spiegel CN, Dias DB, Araki AS, Hamilton JG, Brazil RP, Jones TM. The Lutzomyia longipalpis complex: a brief natural history of aggregation-sex pheromone communication. Parasit Vectors. 2016;9:580. https://doi.org/10.1186/s13071-016-1866-x.
Poldy J. Volatile cues influence host-choice in arthropod pests. Animals. 2020;10:1984. https://doi.org/10.3390/ani10111984.
Alavez-Rosas D, Vargas-Abasolo R, Albores-Flores CI, Meneses-Arias MG, Gutiérrez-Cabrera AE, Benelli G, et al. Chemical ecology of triatomines: current knowledge and implications for Chagas disease vector management. J Pest Sci. 2024;97:507–20. https://doi.org/10.1007/s10340-023-01678-6.
Carapito Â, Roque ACA, Carvalho F, Pinto J, de Guedes Pinho P. Exploiting volatile fingerprints for bladder cancer diagnosis: a scoping review of metabolomics and sensor-based approaches. Talanta. 2024;268:125296. https://doi.org/10.1016/j.talanta.2023.125296.
de Oliveira Filho JG, Sarria ALF, Ferreira LL, Caulfield JC, Powers SJ, Pickett JA, et al. Quantification of brown dog tick repellents, 2-hexanone and benzaldehyde, and release from tick-resistant beagles, Canis lupus familiaris. J Chromatogr B Analyt Technol Biomed Life Sci. 2016;1022:64–9. https://doi.org/10.1016/j.jchromb.2016.03.014.
Hassaballa IB, Matoke-Muhia D, Masiga DK, Sole CL, Torto B, Tchouassi DP. Behavioural responses of Phlebotomus duboscqi to plant-derived volatile organic compounds. Med Vet Entomol. 2021;35:625–32. https://doi.org/10.1111/mve.12541.
Wooding M, Rohwer ER, Naudé Y. Chemical profiling of the human skin surface for malaria vector control via a non-invasive sorptive sampler with GC×GC-TOFMS. Anal Bioanal Chem. 2020;412:5759–77. https://doi.org/10.1007/s00216-020-02799-y.
Zhang MZ, Wang J, Du LF, He PJ, Jia N. The impact of volatiles on tick-host interaction and vector competence. Curr Opin Insect Sci. 2024;62:101162. https://doi.org/10.1016/j.cois.2024.101162.
Hassaballa IB, Torto B, Sole CL, Tchouassi DP. Exploring the influence of different habitats and their volatile chemistry in modulating sand fly population structure in a leishmaniasis endemic foci Kenya. PLoS Negl Trop Dis. 2021;15:e0009062. https://doi.org/10.1371/journal.pntd.0009062.
Khan Z, Bohman B, Ignell R, Hill SR. Odour-mediated oviposition site selection in Aedes aegypti depends on aquatic stage and density. Parasit Vectors. 2023;16:264. https://doi.org/10.1186/s13071-023-05867-1.
Dormont L, Mulatier M, Carrasco D, Cohuet A. Mosquito attractants. J Chem Ecol. 2021;47:351–93. https://doi.org/10.1007/s10886-021-01261-2.
Michalet S, Minard G, Chevalier W, Meiffren G, Saucereau Y, Van Tran V, et al. Identification of human skin bacteria attractive to the Asian tiger mosquito. Environ Microbiol. 2019;21:4662–74. https://doi.org/10.1111/1462-2920.14793.
Isberg E, Ignell R. Cattle-derived unsaturated aldehydes repel biting midges and mosquitoes. J Chem Ecol. 2022;48:359–69. https://doi.org/10.1007/s10886-021-01347-x.
Takabayashi J, Shiojiri K. Multifunctionality of herbivory-induced plant volatiles in chemical communication in tritrophic interactions. Curr Opin Insect Sci. 2019;32:110–7. https://doi.org/10.1016/j.cois.2019.01.003.
Razo-Belman R, Ozuna C. Volatile organic compounds: a review of their current applications as pest biocontrol and disease management. Horticulturae. 2023;9:441. https://doi.org/10.3390/horticulturae9040441.
Busula AO, Takken W, de Boer JG, Mukabana WR, Verhulst NO. Variation in host preferences of malaria mosquitoes is mediated by skin bacterial volatiles. Med Vet Entomol. 2017;31:320–6. https://doi.org/10.1111/mve.12242.
Sethi S, Nanda R, Chakraborty T. Clinical application of volatile organic compound analysis for detecting infectious diseases. Clin Microbiol Rev. 2013;26:462–75. https://doi.org/10.1128/CMR.00020-13.
Rodríguez-Hernández P, Rodríguez-Estévez V, Arce L, Gómez-Laguna J. Application of volatilome analysis to the diagnosis of Mycobacteria infection in livestock. Front Vet Sci. 2021;8:635155. https://doi.org/10.3389/fvets.2021.635155.
Oxner M, Trang A, Mehta J, Forsyth C, Swanson B, Keshavarzian A, et al. The versatility and diagnostic potential of VOC profiling for non infectious diseases. BME Front. 2023;4:0002. https://doi.org/10.34133/bmef.0002.
Kaplan I. Attracting carnivorous arthropods with plant volatiles: the future of biocontrol or playing with fire? Biol Control. 2012;60:77–89. https://doi.org/10.1016/j.biocontrol.2011.10.017.
Giunti G, Benelli G, Palmeri V, Canale A. Bactrocera oleae-induced olive VOCs routing mate searching in Psyttalia concolor males: impact of associative learning. Bull Entomol Res. 2018;108:40–7. https://doi.org/10.1017/S0007485317000451.
Svobodová M, Sádlová J, Chang KP, Volf P. Short report: distribution and feeding preference of the sand flies Phlebotomus sergenti and P. papatasi in a cutaneous leishmaniasis focus in Sanliurfa Turkey. Am J Trop Med Hyg. 2003;68:6–9.
Allan BF, Goessling LS, Storch GA, Thach RE. Blood meal analysis to identify reservoir hosts for Amblyomma americanum ticks. Emerg Infect Dis. 2010;16:433–40. https://doi.org/10.3201/eid1603.090911.
Wodecka B, Rymaszewska A, Skotarczak B. Host and pathogen DNA identification in blood meals of nymphal Ixodes ricinus ticks from forest parks and rural forests of Poland. Exp Appl Acarol. 2014;62:543–55. https://doi.org/10.1007/s10493-013-9763-x.
Kang DS, Martinez R, Hosein A, Shui Feng R, James L, Lovin DD, et al. Identification of host blood meals of mosquitoes (Diptera: Culicidae) collected at the Aripo savannas scientific reserve in Trinidad West Indies. J Med Entomol. 2019;56:1734–8. https://doi.org/10.1093/jme/tjz113.
Hlavackova K, Dvorak V, Chaskopoulou A, Volf P, Halada P. A novel MALDI-TOF MS-based method for blood meal identification in insect vectors: a proof of concept study on phlebotomine sand flies. PLoS Negl Trop Dis. 2019;13:e0007669. https://doi.org/10.1371/journal.pntd.0007669.
Messahel NE, Benallal KE, Halada P, Lafri I, Manseur H, Hakem A, et al. Identification of blood source preferences and Leishmania infection in sand flies (Diptera: Psychodidae) in north-eastern Algeria. Vet Parasitol Reg Stud Reports. 2022;31:100729. https://doi.org/10.1016/j.vprsr.2022.100729.
Katusi GC, Hermy MRG, Makayula SM, Ignell R, Govella NJ, Hill SR, et al. Seasonal variation in abundance and blood meal sources of primary and secondary malaria vectors within Kilombero Valley Southern Tanzania. Parasit Vectors. 2022;15:479. https://doi.org/10.1186/s13071-022-05586-z.
Remadi L, Farjallah D, Chargui N, Belgacem S, Baba H, Zrieq R, et al. Blood meal analysis and molecular detection of mammalian Leishmania DNA in wild-caught Sergentomyia spp. from Tunisia and Saudi Arabia. Parasitol Res. 2023;122:2181–91. https://doi.org/10.1007/s00436-023-07919-y.
Pittman W, Pollock NB, Taylor EN. Effect of host lizard anemia on host choice and feeding rate of larval western black-legged ticks (Ixodes pacificus). Exp Appl Acarol. 2013;61:471–9. https://doi.org/10.1007/s10493-013-9709-3.
Sadlova J, Vojtkova B, Becvar T, Lestinova T, Spitzova T, Bates P, et al. Host competence of the African rodents Arvicanthis neumanni, A. niloticus and Mastomys natalensis for Leishmania donovani from Ethiopia and L. (Mundinia) sp. from Ghana. Int J Parasitol Par Wildlf. 2020;11:40–5. https://doi.org/10.1016/j.ijppaw.2019.12.002.
Mendoza-Roldan JA, Zatelli A, Latrofa MS, Iatta R, Bezerra-Santos MA, Annoscia G, et al. Leishmania (Sauroleishmania) tarentolae isolation and sympatric occurrence with Leishmania (Leishmania) infantum in geckoes, dogs and sand flies. PLoS Negl Trop Dis. 2022;16:e0010650. https://doi.org/10.1371/journal.pntd.0010650.
Giraldo D, McMeniman CJ. Quantification of Anopheles gambiae olfactory preferences under semi-field conditions. Cold Spring Harb Protoc. 2023. https://doi.org/10.1101/pdb.prot108304.
Barbosa-Cornelio R, Cantor F, Coy-Barrera E, Rodríguez D. Tools in the investigation of volatile semiochemicals on insects: from sampling to statistical analysis. Insects. 2019;10:241. https://doi.org/10.3390/insects10080241.
Takken W, Knols BG. Odor-mediated behavior of Afrotropical malaria mosquitoes. Annu Rev Entomol. 1999;44:131–57. https://doi.org/10.1146/annurev.ento.44.1.131.
Smallegange RC, Qiu YT, van Loon JJ, Takken W. Synergism between ammonia, lactic acid and carboxylic acids as kairomones in the host-seeking behaviour of the malaria mosquito Anopheles gambiae sensu stricto (Diptera: Culicidae). Chemic Senses. 2005;30:145–52. https://doi.org/10.1093/chemse/bji010.
Ríos-Delgado SM, Rodríguez-Ramírez AD, Cruz-López L, Escobar-Pérez LA, Aburto-Juárez Mde L, Torres-Estrada JL. Respuesta de Anopheles albimanus a compuestos volátiles de casas del sur de Chiapas, México [Behavioral response of Anopheles albimanus to volatile compounds collected inside houses from the south of Chiapas, Mexico]. Salud Publica Mex. 2008;50:367–74. https://doi.org/10.1590/s0036-36342008000500009.
Smallegange RC, Qiu YT, Bukovinszkine-Kiss G, Van Loon JJA, Takken W. The effect of aliphatic carboxylic acids on olfaction-based host-seeking of the malaria mosquito Anopheles gambiae sensu stricto. J Chem Ecol. 2009;35:933–43. https://doi.org/10.1007/s10886-009-9668-7.
Verhulst NO, Beijleveld H, Knols BG, Takken W, Schraa G, Bouwmeester HJ, et al. Cultured skin microbiota attracts malaria mosquitoes. Malar J. 2009;8:302. https://doi.org/10.1186/1475-2875-8-302.
Carey AF, Wang G, Su CY, Zwiebel LJ, Carlson JR. Odorant reception in the malaria mosquito Anopheles gambiae. Nature. 2010;464:66–71. https://doi.org/10.1038/nature08834.
Verhulst NO, Mbadi PA, Kiss GB, Mukabana WR, van Loon JJ, Takken W, et al. Improvement of a synthetic lure for Anopheles gambiae using compounds produced by human skin microbiota. Malar J. 2011;10:28. https://doi.org/10.1186/1475-2875-10-28.
Smallegange RC, Bukovinszkine-Kiss G, Otieno B, Mbadi PA, Takken W, Mukabana WR, et al. Identification of candidate volatiles that affect the behavioural response of the malaria mosquito Anopheles gambiae sensu stricto to an active kairomone blend: laboratory and semi-field assays. Physiol Entomol. 2012;37:60–71. https://doi.org/10.1111/j.1365-3032.2011.00827.x.
Tchouassi DP, Sang R, Sole CL, Bastos AD, Mithoefer K, Torto B. Sheep skin odor improves trap captures of mosquito vectors of Rift Valley fever. PLoS Negl Trop Dis. 2012;6:e1879. https://doi.org/10.1371/journal.pntd.0001879.
Torres Estrada JL, Ríos Delgado SM, Takken W. Indoor volatiles of primary school classrooms in Tapachula, Chiapas, Mexico, are attractants to Aedes aegypti females. J Am Mosq Control Assoc. 2013;29:297–300. https://doi.org/10.2987/12-6278.1.
Verhulst NO, Beijleveld H, Qiu YT, Maliepaard C, Verduyn W, Haasnoot GW, et al. Relation between HLA genes, human skin volatiles and attractiveness of humans to malaria mosquitoes. Infect Genet Evol. 2013;18:87–93. https://doi.org/10.1016/j.meegid.2013.05.009.
Jaleta KT, Hill SR, Birgersson G, Tekie H, Ignell R. Chicken volatiles repel host-seeking malaria mosquitoes. Malar J. 2016;15:354. https://doi.org/10.1186/s12936-016-1386-3.
Verhulst NO, Weldegergis BT, Menger D, Takken W. Attractiveness of volatiles from different body parts to the malaria mosquito Anopheles coluzzii is affected by deodorant compounds. Sci Rep. 2016;6:27141. https://doi.org/10.1038/srep27141.
Kemibala EE, Mafra-Neto A, Saroli J, Silva R, Philbert A, Ng’habi K, et al. Is Anopheles gambiae attraction to floral and human skin-based odours and their combination modulated by previous blood meal experience? Malar J. 2020;19:318. https://doi.org/10.1186/s12936-020-03395-2.
Kythreoti G, Sdralia N, Tsitoura P, Papachristos DP, Michaelakis A, Karras V, et al. Volatile allosteric antagonists of mosquito odorant receptors inhibit human-host attraction. J Biol Chem. 2021;296:100172. https://doi.org/10.1074/jbc.RA120.016557.
Omolo MO, Ndiege IO, Hassanali A. Semiochemical signatures associated with differential attraction of Anopheles gambiae to human feet. PLoS ONE. 2021;16:e0260149. https://doi.org/10.1371/journal.pone.0260149.
Giraldo D, Rankin-Turner S, Corver A, Tauxe GM, Gao AL, Jackson DM, et al. Human scent guides mosquito thermotaxis and host selection under naturalistic conditions. Curr Biol. 2023;33:2367-2382.e7. https://doi.org/10.1016/j.cub.2023.04.050.
Osterkamp J, Wahl U, Schmalfuss G, Haas W. Host-odour recognition in two tick species is coded in a blend of vertebrate volatiles. J Comp Physiol A. 1999;185:59–67. https://doi.org/10.1007/s003590050366.
Bunnell T, Hanisch K, Hardege JD, Breithaupt T. The fecal odor of sick hedgehogs (Erinaceus europaeus) mediates olfactory attraction of the tick Ixodes hexagonus. J Chem Ecol. 2011;37:340–7. https://doi.org/10.1007/s10886-011-9936-1.
Borges LM, de Oliveira Filho JG, Ferreira LL, Louly CC, Pickett JA, Birkett MA. Identification of non-host semiochemicals for the brown dog tick, Rhipicephalus sanguineus sensu lato (Acari: Ixodidae), from tick-resistant beagles, Canis lupus familiaris. Ticks Tick Borne Dis. 2015;6:676–82. https://doi.org/10.1016/j.ttbdis.2015.05.014.
Kariuki MW, Hassanali A, Ng’ang’a MM. Characterisation of cattle anal odour constituents associated with the repellency of Rhipicephalus appendiculatus. Exp Appl Acarol. 2018;76:221–7. https://doi.org/10.1007/s10493-018-0304-5.
Faraone N, Light M, Scott C, MacPherson S, Hillier NK. Chemosensory and behavioural responses of Ixodes scapularis to natural products: role of chemosensory organs in volatile detection. Insects. 2020;11:502. https://doi.org/10.3390/insects11080502.
Zeringóta V, Pereira-Junior RA, Sarria ALF, Henrique ACC, Birkett MA, Borges LMF. Identification of a non-host semiochemical from miniature pinscher, Canis lupus familiaris, that repels Rhipicephalus sanguineus sensu lato (Acari: Ixodidae). Ticks Tick Borne Dis. 2021;12:101582. https://doi.org/10.1016/j.ttbdis.2020.101582.
López-López N, Rojas JC, Cruz-López L, Ulloa-García A, Malo EA. Dog hair volatiles attract Rhipicephalus sanguineus sensu lato (Acari: Ixodidae) females. J Med Entomol. 2023;60:432–42. https://doi.org/10.1093/jme/tjad019.
Rebollar-Tellez EA, Hamilton JGC, Ward RD. Response of female Lutzomyia longipalpis to host odour kairomones from human skin. Physiol Entomol. 1999;24:220–6. https://doi.org/10.1046/j.1365-3032.1999.00133.x.
Andrade AJ, Andrade MR, Dias ES, Pinto MC, Eiras ÁE. Are light traps baited with kairomones effective in the capture of Lutzomyia longipalpis and Lutzomyia intermedia? An evaluation of synthetic human odor as an attractant for phlebotomine sand flies (Diptera: Psychodidae: Phlebotominae). Mem Instit Oswaldo Cruz. 2008;103:337–43. https://doi.org/10.1590/S0074-02762008000400004.
Pinto MC, Bray DP, Eiras AE, Carvalheira HP, Puertas CP. Attraction of the cutaneous leishmaniasis vector Nyssomyia neivai (Diptera: Psychodidae) to host odour components in a wind tunnel. Parasit Vectors. 2012;5:210. https://doi.org/10.1186/1756-3305-5-210.
Machado VE, Corrêa AG, Goulart TM, Silva FB, Ortiz DG, Pinto MC. Attraction of the sand fly Nyssomyia neivai (Diptera: Psychodidae) to chemical compounds in a wind tunnel. Parasit Vectors. 2015;8:147. https://doi.org/10.1186/s13071-015-0748-y.
Tavares DDS, Salgado VR, Miranda JC, Mesquita PRR, Rodrigues FM, Barral-Netto M, et al. Attraction of phlebotomine sandflies to volatiles from skin odors of individuals residing in an endemic area of tegumentary leishmaniasis. PLoS ONE. 2018;13:e0203989. https://doi.org/10.1371/journal.pone.0203989.
Magalhães-Junior JT, Barrouin-Melo SM, Corrêa AG, da Rocha Silva FB, Machado VE, Govone JS, et al. A laboratory evaluation of alcohols as attractants for the sandfly Lutzomyia longipalpis (Diptera:Psychodidae). Parasit Vectors. 2014;7:60. https://doi.org/10.1186/1756-3305-7-60.
Magalhães-Junior JT, Oliva-Filho AA, Novais HO, Mesquita PRR, Rodrigues FM, Pinto MC, et al. Attraction of the sandfly Lutzomyia longipalpis to possible biomarker compounds from dogs infected with Leishmania infantum. Med Vet Entomol. 2019;33:322–5. https://doi.org/10.1111/mve.12357.
Staniek ME, Hamilton JGC. Odour of domestic dogs infected with Leishmania infantum is attractive to female but not male sand flies: evidence for parasite manipulation. PLoS Pathog. 2021;17:e1009354. https://doi.org/10.1371/journal.ppat.1009354.
Chelbi I, Maghraoui K, Zhioua S, Cherni S, Labidi I, Satoskar A, et al. Enhanced attraction of sand fly vectors of Leishmania infantum to dogs infected with zoonotic visceral leishmaniasis. PLoS Negl Trop Dis. 2021;15:e0009647. https://doi.org/10.1371/journal.pntd.0009647.
Wachira BM, Kabaka JM, Mireji PO, Okoth SO, Nganga MM, Changasi R, et al. Characterization of a composite with enhanced attraction to savannah tsetse flies from constituents or analogues of tsetse refractory waterbuck (Kobus defassa) body odor. PLoS Negl Trop Dis. 2021;15:e0009474. https://doi.org/10.1371/journal.pntd.0009474.
Mireji PO, Mang’era CM, Bwana BK, Hassanali A. Perspectives on odor-based control of tsetse flies in Africa. Front Physiol. 2022;13:831618. https://doi.org/10.3389/fphys.2022.831618.
Franklinos LHV, Jones KE, Redding DW, Abubakar I. The effect of global change on mosquito-borne disease. Lancet Infect Dis. 2019;19:e302–12. https://doi.org/10.1016/S1473-3099(19)30161-6.
Benelli G. Pathogens manipulating tick behaviour—through a glass, darkly. Pathogens. 2020;9:664. https://doi.org/10.3390/pathogens9080664.
Giunti G, Becker N, Benelli G. Invasive mosquito vectors in Europe: from bioecology to surveillance and management. Acta Trop. 2023;239:106832. https://doi.org/10.1016/j.actatropica.2023.106832.
Braks MAH, Meijerink J, Takken W. The response of the malaria mosquito, Anopheles gambiae, to two components of human sweat, ammonia and l-lactic acid, in an olfactometer. Physiol Entomol. 2001;26:142–8. https://doi.org/10.1046/j.1365-3032.2001.00227.x.
Dekker T, Steib B, Cardé RT, Geier M. L-lactic acid: a human-signifying host cue for the anthropophilic mosquito Anopheles gambiae. Med Vet Entomol. 2002;16:91–8. https://doi.org/10.1046/j.0269-283x.2002.00345.x.
Smallegange RC, Knols BG, Takken W. Effectiveness of synthetic versus natural human volatiles as attractants for Anopheles gambiae (Diptera: Culicidae) sensu stricto. J Med Entomol. 2010;47:338–44. https://doi.org/10.1603/me09015.
Logan JG, Birkett MA, Clark SJ, Powers S, Seal NJ, Wadhams LJ, et al. Identification of human-derived volatile chemicals that interfere with attraction of Aedes aegypti mosquitoes. J Chem Ecol. 2008;34:308–22. https://doi.org/10.1007/s10886-008-9436-0.
Syed Z, Leal WS. Acute olfactory response of Culex mosquitoes to a human- and bird-derived attractant. Proc Natl Acad Sci U S A. 2009;106:18803–8. https://doi.org/10.1073/pnas.0906932106.
de Boer JG, Robinson A, Powers SJ, Burgers SLGE, Caulfield JC, Birkett MA, et al. Odours of Plasmodium falciparum-infected participants influence mosquito-host interactions. Sci Rep. 2017;7:9283. https://doi.org/10.1038/s41598-017-08978-9.
Wynne NE, Lorenzo MG, Vinauger C. Mechanism and plasticity of vectors’ host-seeking behavior. Curr Opin Insect Sci. 2020;40:1–5. https://doi.org/10.1016/j.cois.2020.02.001.
Allan SA, Bernier UR, Kline DL. Attraction of mosquitoes to volatiles associated with blood. J Vector Ecol. 2006;31:71–8.
Geier M, Boeckh J. A new Y-tube olfactometer for mosquitoes to measure the attractiveness of host odours. Entomol Exp Appl. 1999;1999:9–19. https://doi.org/10.1046/j.1570-7458.1999.00519.x.
Healy TP, Copland MJ. Human sweat and 2-oxopentanoic acid elicit a landing response from Anopheles gambiae. Med Vet Entomol. 2000;14:195–200. https://doi.org/10.1046/j.1365-2915.2000.00238.x.
van Loon JJ, Smallegange RC, Bukovinszkiné-Kiss G, Jacobs F, De Rijk M, Mukabana WR, et al. Mosquito attraction: crucial role of carbon dioxide in formulation of a five-component blend of human-derived volatiles. J Chem Ecol. 2015;41:567–73. https://doi.org/10.1007/s10886-015-0587-5.
Eneh LK, Fillinger U, Borg Karlson AK, Kuttuva Rajarao G, Lindh J. Anopheles arabiensis oviposition site selection in response to habitat persistence and associated physicochemical parameters, bacteria and volatile profiles. Med Vet Entomol. 2019;33:56–67. https://doi.org/10.1111/mve.12336.
Schoelitsz B, Mwingira V, Mboera LEG, Beijleveld H, Koenraadt CJM, Spitzen J, et al. Chemical mediation of oviposition by Anopheles mosquitoes: a push-pull system driven by volatiles associated with larval stages. J Chem Ecol. 2020;46:397–409. https://doi.org/10.1007/s10886-020-01175-5.
Elliott RM, Brennan B. Emerging phleboviruses. Curr Opin Virol. 2014;5:50–7. https://doi.org/10.1016/j.coviro.2014.01.011.
Maroli M, Feliciangeli MD, Bichaud L, Charrel RN, Gradoni L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med Vet Entomol. 2013;27:123–47. https://doi.org/10.1111/j.1365-2915.2012.01034.x.
Cecílio P, Cordeiro-da-Silva A, Oliveira F. Sand flies: basic information on the vectors of leishmaniasis and their interactions with Leishmania parasites. Commun Biol. 2022;5:305. https://doi.org/10.1038/s42003-022-03240-z.
Maia C, Dantas-Torres F, Campino L. Parasite biology: the reservoir hosts. In: Bruschi F, Gradoni L, editors. The leishmaniases: old neglected tropical diseases. Cham: Springer; 2018. https://doi.org/10.1007/978-3-319-72386-0_4.
González E, Molina R, Iriso A, Ruiz S, Aldea I, Tello A, et al. Opportunistic feeding behaviour and Leishmania infantum detection in Phlebotomus perniciosus females collected in the human leishmaniasis focus of Madrid, Spain (2012–2018). PLoS Negl Trop Dis. 2021;15:e0009240. https://doi.org/10.1371/journal.pntd.0009240.
Lozano-Sardaneta YN, Jiménez-Girón EI, Rodríguez-Rojas JJ, Sánchez-Montes S, Álvarez-Castillo L, Sánchez-Cordero V, et al. Species diversity and blood meal sources of phlebotomine sand flies (Diptera: Psychodidae) from Los Tuxtlas, Veracruz. Mexico Acta Trop. 2021;216:105831. https://doi.org/10.1016/j.actatropica.2021.105831.
Calzolari M, Romeo G, Bergamini F, Dottori M, Rugna G, Carra E. Host preference and Leishmania infantum natural infection of the sand fly Phlebotomus perfiliewi in northern Italy. Acta Trop. 2022;226:106246. https://doi.org/10.1016/j.actatropica.2021.106246.
Hamilton JG, Ramsoondar TM. Attraction of Lutzomyia longipalpis to human skin odours. Med Vet Entomol. 1994;8:375–80. https://doi.org/10.1111/j.1365-2915.1994.tb00101.x.
Dougherty MJ, Guerin PM, Ward RD, Hamilton JGC. Behavioural and electrophysiological responses of the phlebotomine sandfly Lutzomyia longipalpis (Diptera: Psychodidae) when exposed to canid host odour kairomones. Physiol Entomol. 1999;24:251–62. https://doi.org/10.1046/j.1365-3032.1999.00139.x.
Kakumanu ML, Marayati BF, Schal C, Apperson CS, Wasserberg G, Ponnusamy L. Oviposition-site selection of Phlebotomus papatasi (Diptera: Psychodidae) sand flies: attraction to bacterial isolates from an attractive rearing medium. J Med Entomol. 2021;58:518–27. https://doi.org/10.1093/jme/tjaa198.
Dantas-Torres F, Chomel BB, Otranto D. Ticks and tick-borne diseases: a one health perspective. Trends Parasitol. 2012;28:437–46. https://doi.org/10.1016/j.pt.2012.07.003.
Jongejan F, Uilenberg G. The global importance of ticks. Parasitol. 2004;129:S3-14. https://doi.org/10.1017/s0031182004005967.
Otranto D. Arthropod-borne pathogens of dogs and cats: from pathways and times of transmission to disease control. Vet Parasitol. 2018;251:68–77. https://doi.org/10.1016/j.vetpar.2017.12.021.
Ferreira LL, de Oliveira Filho JG, de Oliveira SF, Lacerda Ferraz AL, Mascarin GM. Attract or repel Amblyomma sculptum ticks: screening of semiochemicals. Vet Parasitol. 2020;278:109036. https://doi.org/10.1016/j.vetpar.2020.109036.
Ferreira LL, Sarria ALF, de Oliveira Filho JG, de Silva FO, Powers SJ, Caulfield JC, et al. Identification of a non-host semiochemical from tick-resistant donkeys (Equus asinus) against Amblyomma sculptum ticks. Ticks Tick Borne Dis. 2019;10:621–7. https://doi.org/10.1016/j.ttbdis.2019.02.006.
Louly CC, Soares SF, Silveira DN, Neto OJ, Silva AC, Borges LM. Differences in the susceptibility of two breeds of dogs, English cocker spaniel and beagle, to Rhipicephalus sanguineus (Acari: Ixodidae). Int J Acarol. 2009;35:25–32. https://doi.org/10.1080/01647950802655251.
Louly CC, Soares SF, da Nóbrega SD, Guimarães MS, Borges LM. Differences in the behavior of Rhipicephalus sanguineus tested against resistant and susceptible dogs. Exp Appl Acarol. 2010;51:353–62. https://doi.org/10.1007/s10493-009-9334-3.
Kelly M, Su CY, Schaber C, Crowley JR, Hsu FF, Carlson JR, et al. Malaria parasites produce volatile mosquito attractants. MBio. 2015;6:e00235-e315. https://doi.org/10.1128/mBio.00235-15.
Correa R, Coronado LM, Garrido AC, Durant-Archibold AA, Spadafora C. Volatile organic compounds associated with Plasmodium falciparum infection in vitro. Parasit Vectors. 2017;10:215. https://doi.org/10.1186/s13071-017-2157-x.
Debebe Y, Hill SR, Birgersson G, Tekie H, Ignell R. Plasmodium falciparum gametocyte-induced volatiles enhance attraction of Anopheles mosquitoes in the field. Malar J. 2020;19:327. https://doi.org/10.1186/s12936-020-03378-3.
Sanford T, Shutler D. Manipulation by Plasmodium parasites of Anopheles mosquito behavior and human odors. Acta Parasitol. 2022;67:1463–70. https://doi.org/10.1007/s11686-022-00621-6.
Magalhães-Junior JT, Mesquita PR, Oliveira WF, Oliveira FS, Franke CR, Rodrigues FM, et al. Identification of biomarkers in the hair of dogs: new diagnostic possibilities in the study and control of visceral leishmaniasis. Anal Bioanal Chem. 2014;406:6691–700. https://doi.org/10.1007/s00216-014-8103-2.
Nevatte TM, Ward RD, Sedda L, Hamilton JGC. After infection with Leishmania infantum, Golden Hamsters (Mesocricetus auratus) become more attractive to female sand flies (Lutzomyia longipalpis). Sci Rep. 2017;7:6104. https://doi.org/10.1038/s41598-017-06313-w.
Cator LJ, Lynch PA, Read AF, Thomas MB. Do malaria parasites manipulate mosquitoes? Trends Parasitol. 2012;28:466–70. https://doi.org/10.1016/j.pt.2012.08.004.
Cator L. Malaria altering host attractiveness and mosquito feeding. Trends Parasitol. 2017;33:338–9. https://doi.org/10.1016/j.pt.2017.03.003.
Busula AO, Verhulst NO, Bousema T, Takken W, de Boer JG. Mechanisms of Plasmodium-enhanced attraction of mosquito vectors. Trends Parasitol. 2017;33:961–73. https://doi.org/10.1016/j.pt.2017.08.010.
Emami SN, Lindberg BG, Hua S, Hill SR, Mozuraitis R, Lehmann P, et al. A key malaria metabolite modulates vector blood seeking, feeding, and susceptibility to infection. Science. 2017;355:1076–80. https://doi.org/10.1126/science.aah4563.
Emami SN, Hajkazemian M, Mozūraitis R. Can Plasmodium’s tricks for enhancing its transmission be turned against the parasite? New hopes for vector control. Pathog Glob Health. 2019;113:325–35. https://doi.org/10.1080/20477724.2019.1703398.
Schaber CL, Katta N, Bollinger LB, Mwale M, Mlotha-Mitole R, Trehan I, et al. Breathprinting reveals malaria-associated biomarkers and mosquito attractants. J Infect Dis. 2018;217:1553–60. https://doi.org/10.1093/infdis/jiy072.
Miller JJ, Odom John AR. The malaria metabolite HMBPP does not trigger erythrocyte terpene release. ACS Infect Dis. 2020;6:2567–72. https://doi.org/10.1021/acsinfecdis.0c00548.
Robinson A, Busula AO, Voets MA, Beshir KB, Caulfield JC, Powers SJ, et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc Natl Acad Sci U S A. 2018;115:E4209–18. https://doi.org/10.1073/pnas.1721610115.
De Moraes CM, Stanczyk NM, Betz HS, Pulido H, Sim DG, Read AF, et al. Malaria-induced changes in host odors enhance mosquito attraction. Proc Natl Acad Sci U S A. 2014;111:11079–84. https://doi.org/10.1073/pnas.1405617111.
De Moraes CM, Wanjiku C, Stanczyk NM, Pulido H, Sims JW, Betz HS, et al. Volatile biomarkers of symptomatic and asymptomatic malaria infection in humans. Proc Natl Acad Sci U S A. 2018;115:5780–5. https://doi.org/10.1073/pnas.1801512115.
Smallegange RC, Verhulst NO, Takken W. Sweaty skin: an invitation to bite? Trends Parasitol. 2011;27:143–8. https://doi.org/10.1016/j.pt.2010.12.009.
Ellwanger JH, Cardoso JDC, Chies JAB. Variability in human attractiveness to mosquitoes. Curr Res Parasitol Vector Borne Dis. 2021;1:100058. https://doi.org/10.1016/j.crpvbd.2021.100058.
Wilke ABB, Benelli G, Beier JC. Anthropogenic changes and associated impacts on vector-borne diseases. Trends Parasitol. 2021;37:1027–30. https://doi.org/10.1016/j.pt.2021.09.013.
Zhang H, Zhu Y, Liu Z, Peng Y, Peng W, Tong L, et al. A volatile from the skin microbiota of flavivirus-infected hosts promotes mosquito attractiveness. Cell. 2022;185:2510-2522.e16. https://doi.org/10.1016/j.cell.2022.05.016.
Kim D, Crippen TL, Dhungel L, Delclos PJ, Tomberlin JK, Jordan HR. Behavioral interplay between mosquito and mycolactone produced by Mycobacterium ulcerans and bacterial gene expression induced by mosquito proximity. PLoS ONE. 2023;18:e0289768. https://doi.org/10.1371/journal.pone.0289768.
Acknowledgements
Not applicable.
Funding
This research was supported by EU funding within the MUR PNRR Extended Partnership initiative on Emerging Infectious Diseases (Project no. PE00000007, INF-ACT).
Author information
Authors and Affiliations
Contributions
Marcos Antonio Bezerra-Santos: Investigation, formal analysis, data curation, writing—original draft, writing—review and editing. Giovanni Benelli: Investigation, formal analysis, writing—review and editing. Giacinto Salvatore Germinara: Investigation, formal analysis, writing—review and editing. Petr Volf: Investigation, formal analysis, writing—review and editing. Domenico Otranto: Investigation, formal analysis, data curation, writing—review and editing. All authors contributed to the study and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
None.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Bezerra-Santos, M.A., Benelli, G., Germinara, G.S. et al. Smelly interactions: host-borne volatile organic compounds triggering behavioural responses in mosquitoes, sand flies, and ticks. Parasites Vectors 17, 227 (2024). https://doi.org/10.1186/s13071-024-06299-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-024-06299-1