
Abstract
Background
Trypanosomatid parasites are widely distributed in nature and can have a monoxenous or dixenous life-cycle. These parasites thrive in a wide number of insect orders, some of which have an important economic and environmental value, such as bees. The objective of this study was to develop a robust and sensitive real-time quantitative PCR (qPCR) assay for detecting trypanosomatid parasites in any type of parasitized insect sample.
Methods
A TaqMan qPCR assay based on a trypanosomatid-conserved region of the α-tubulin gene was standardized and evaluated. The limits of detection, sensitivity and versatility of the α-tubulin TaqMan assay were tested and validated using field samples of honeybee workers, wild bees, bumblebees and grasshoppers, as well as in the human infective trypanosomatid Leishmania major.
Results
The assay showed a detection limit of 1 parasite equivalent/µl and successfully detected trypanosomatids in 10 different hosts belonging to the insect orders Hymenoptera and Orthoptera. The methodology was also tested using honeybee samples from four apiaries (n = 224 worker honeybees) located in the Alpujarra region (Granada, Spain). Trypanosomatids were detected in 2.7% of the honeybees, with an intra-colony prevalence of 0% to 13%. Parasite loads in the four different classes of insects ranged from 40.6 up to 1.1 × 108 cell equivalents per host.
Conclusions
These results show that the α-tubulin TaqMan qPCR assay described here is a versatile diagnostic tool for the accurate detection and quantification of trypanosomatids in a wide range of environmental settings.
Graphical Abstract

Similar content being viewed by others

Background
Trypanosomatids (Euglenozoa: Kinetoplastea: Trypanosomatidae) are a widely distributed family of protozoan parasites that can infect numerous orders of insects [1, 2]. Parasites of the family Trypanosomatidae are classified on a nontaxonomic basis into two groups based on the number of hosts needed to complete development, with dixenous trypanosomatids (5 genera) having a life-cycle that requires infection of both an insect vector and a mammalian host and monoxenous trypanosomatids (19 genera) requiring only a singular, particular host, mainly insects in the orders Diptera, Hemiptera and Hymenoptera [1]. Monoxenous trypanosomatids have developed numerous strategies for transmission, such as cannibalism, predation, necrophagy, injection with saliva and fecal, oral or transovarial route [1, 3,4,5], and they can be found in adult-stage insects but also in larvae, pupae and or insect eggs. These parasites show diverse locations for thriving and proliferating within their insect hosts, such as the digestive tract, particularly the gut (foregut, midgut or hindgut), but also the Malpighian tubes and the hemolymph [3, 6, 7]. The wide diversity of typing units (TUs) found in trypanosomatids (> 300 so far) makes them one of the most successful groups of parasites in nature [4].
In this regard, several studies have demonstrated the continuous presence and dissemination of trypanosomatid parasites in insects. For example, a study on the presence of trypanosomatids in Austrian mosquitoes revealed that 12 out of 19 species harbored these parasites with a prevalence ranging from 16% to 61% [8]. In other studies, trypanosomatids were identified in eight species of Drosophila analyzed in southwest Ohio (USA), with a prevalence of 1–17% [9], and the prevalence of trypanosomatids on insects in the island of Curaçao and Papua New Guinea was 38% and 15%, respectively [10, 11]. In addition, the presence of monoxenous parasites has been identified in 4.3% of tsetse flies collected in the Dzanga-Sangha Protected Area (Central African Republic) [12]. Within this broad diversity of hosts, hymenopterans—and particularly bees—are a group of insects in which the presence of trypanosomatid parasites has been widely described. Bees are major pollinators involved in the maintenance of all terrestrial ecosystems, and they are currently experiencing a massive decline in numbers that is related to a wide variety of stressors, including habitat loss, climate change, pesticides and parasites [13].
Four species of trypanosomatid parasites have been identified to date (Lotmaria passim, Crithidia mellificae, C. bombi and C. acanthocephali) in bumblebees, wasps or honeybees [14,15,16]. These four parasite species have been found in hymenopteran samples across four continents, indicating the wide distribution of trypanosomatid flagellates across highly different ecological settings [17,18,19,20,21,22,23]. Trypanosomatid parasites have been associated with winter colony losses [21], reduced bumblebee colony fitness [24, 25] and/or reduced honeybee lifespan under experimental conditions [26], suggesting that they are a significant health threat to these insects.
To obtain a precise picture of the circulating levels of these parasites in nature, novel targets and molecular amplification assays are needed. The use of TaqMan probes versus SYBR green tracking dyes offers several advantages, such as higher specificity and sensitivity, which increases the chances for target detection. This detection sensitivity could be further increased by the use of multicopy gene families or of repeated DNA fragments as primers and probe targets [27, 28]. Sensitive and polyvalent multispecies detection assays are achieved using conserved regions that could capture the diversity of a particular group of organisms. The spliced leader RNA (SL-RNA), which consists of a conserved sequence of 39 bp added to the 5′ end of trypanosomatid pre-messenger RNA (mRNA) molecules, and the 28S ribosomal RNA (rRNA) locus have been used as targets for TaqMan real-time quantitative PCR (qPCR) assays aimed at detecting trypanosomatids in different blood samples, with limits of detection of 100 parasites per milliliter of blood [29, 30]. An alternative to these regions is the alpha/beta (α/β) tubulin array, a conserved multicopy trypanosomatid locus essential for the formation of the microtubular cytoskeleton. This α/β heterodimer is arranged in tandem repeats of gene copies varying in number between nine copies found in the Leishmania mexicana genome [31], 10 copies in Leishmania tarentolae [32], 12 copies in Leishmania amazonensis or Leptomonas pyrrhocoris [33, 34] and up to 19 copies in Trypanosoma brucei [35]. Therefore, this locus could be a good target for universal detection and quantification of trypanosomatid parasites. To date, honeybee trypanosomatid infections have been quantified using SYBR-green qPCR assays using as targets the small subunit rRNA (SSU rRNA) locus [6], the cytochrome b gene [36] or the internal transcribed spacer 1 (ITS1) [37]. However, no TaqMan qPCR assay has been developed yet for detection of trypanosomatid infections in insects. Therefore, the development of such an assay for diagnosis in bees or other insect hosts could be a powerful analytical tool.
Here we report our development of a novel TaqMan qPCR assay based on the amplification of the conserved α-tubulin gene fragment. The limits of detection, sensitivity and versatility of the α-tubulin TaqMan assay were tested and validated using field samples from honeybee workers, wild bees, bumblebees and grasshoppers, and the usefulness of the assay was demonstrated.
Methods
Lotmaria passim culture and field sampling
The trypanosomatid L. passim C1 strain was cultured in modified BHT media as previously described [38]. Genomic DNA (gDNA) was isolated from these cultures during the mid-log phase of growth (5 × 106 cells/ml) using a standard phenol/chloroform/isoamyl protocol (see section Genomic DNA) and then used for generating qPCR standard curves.
During the field sampling, a total of 224 honeybees (approx. 28 bees/hive) from four apiaries (coded as 1–4) located in the Alpujarra region (south of Granada, Spain) were sampled in September 2018. Two hives (coded as A and B) were sampled in each apiary (Additional file 1: Table S1, Figures S1, S2). An additional 50 worker honeybees were collected from rosemary (Rosmarinus officinalis) flowers in the city center of Granada (37°08′48.3″N 3°36′28.2″W). Finally, 43 samples of wild bees, bumblebees and grasshoppers were collected during autumn 2021 and spring 2022 at different locations of Granada and Jaen provinces (Andalucía, Spain) as indicated in Additional file 1: Table S2. All insect samples were immediately frozen at − 20 °C before DNA extraction.
Genomic DNA
Genomic DNA was extracted by crushing the abdomens of adult bees and larvae and incubating the samples overnight in lysis buffer and proteinase K at 55 °C. The gDNA was then purified using a standard phenol/chloroform/isoamyl protocol and treated with RNase A. The concentration and purity of the extracted DNA were determined on a Nanodrop 2000 spectrometer (Thermo Fisher Scientific, Waltham, MA, USA).
α-Tubulin TaqMan qPCR
The primers and probes used for the TaqMan assay were designed to amplify a conserved region of the α-tubulin gene of trypanosomatid parasites (Additional file 1: Table S3, Figure S1A). Representative sequences of the α-tubulin gene from 23 trypanosomatid species were obtained from the TritrypDB database and aligned using ClustalW [39]. ePrimer3 software was used to design primers in the conserved 3′ end of the α-tubulin open reading frame (amplicon length: 193 bp), and the PrimerQuest™ Tool (Integrated DNA Technologies, Inc., Coralville, IA, USA) was used to design the TaqMan probe (Additional file 1: Table S3). BLAST analysis of the α-tubulin locus against organisms of the order Insecta resulted in no potential cross-reactions.
To check the versatility of the α-tubulin locus to amplify trypanosomatids, we performed a conventional PCR using the primers 198F/199R. Each reaction mixture contained 20 mM Tris-HCl, 3 mM MgCl2, 1 mM dNTPs, 0.2 µM of each primer (198F/119R), 3% kb Extender and Platinum® and 2 U of Taq DNA Polymerase (Thermo Fisher Scientific). The PCR amplification protocol consisted of an initial denaturation at 95° for 4 min, followed by 39 cycles at 95 °C for 30 s, 60 °C for 30 s and 72 °C for 20 s, with a final extension at 72 °C for 5 min. gDNA from L. passim cultures was used as the positive control, and autoclaved Milli-Q water was used as the negative control.
The qPCR reactions were performed using the TaqMan™ Fast Universal PCR Master Mix (1×), no AmpErase™ UNG (Applied Biosystems, Thermo Fisher Scientific) in a total reaction volume of 10 μl containing 900 nM of primers (198F-199R), 250 nM of α-tubulin probe and an average of 500 ng of DNA sample. The cycling conditions consisted of 95 °C for 5 min, followed by 40 cycles of 95 °C for 10 s and 60 °C for 25 s, in a CFX96 Real-Time System Thermocycler (Bio-Rad Laboratories, Hercules, CA, USA). The 18S gene was used as the qPCR internal control, using the oligonucleotides and conditions previously described, and reactions were performed in triplicate per sample.
Quantification of trypanosomatid loads
Genomic DNA from “pseudo-infected” bees was used to construct the standard curves for qPCR quantification. This artificial sample was generated mixing non-infected bee gDNA with tenfold decreasing concentrations of L. passim gDNA. The corresponding copy number of α-tubulin/µl in the starting concentration was calculated using the following equation [40].
where nα-tubulin is the number of copies of α-tubulin in the diploid genome of L. passim (copies); CgDNA is the concentration of L. passim gDNA (ng/µl); ConstA is the Avogadro constant (6.022 × 1023 pb/mol); LgDNA is the size of the diploid genome of L. passim SF (PRA-403) strain (64 Mb [41]; and Mpb is the weight of a double-stranded base pair (average: 6.6 × 1011 ng/mol).
The standard curve was generated by tenfold dilutions of 2.24 × 107 down to 2.24 copies/µl of α-tubulin of “pseudo-infected” bee gDNA. Each point of the standard curve was obtained from the mean value of three replicates and was considered acceptable only when all the replicates were positive. The cycle threshold (Ct) values of positive samples were obtained and copies were extrapolated from standard curves in each qPCR. To obtain the number of cell equivalents per sample, the copies/µl obtained in the unknown samples were divided by 24, which is the median number of copies of the α-tubulin gene in the diploid genome of the closest Lotmaria passim related trypanosomatid Leptomonas pyrrhocoris [34], and finally multiplied by the total sample volume in which the sample was resuspended. The analytical sensitivity was taken from the last dilution of the standard curve in which the triplicated parasite DNA was successfully measured.
Sequencing
To determine the trypanosomatid species in the positive samples, the SSU rRNA gene fragment was amplified using the primers S762 and S763 according to Maslov et al. [42]. The sequences obtained have been deposited in the Genbank database under the accession numbers OP805899-OP805907.
Results
The trypanosome α-tubulin region is a pan-specific molecular target in different insect hosts
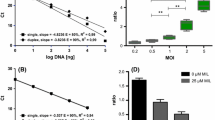
The primers and probes were designed to target a conserved fragment of 193 bp of the α-tubulin gene, as shown in the Clustal analysis of 23 homolog genes in trypanosomatids (Fig. 1a, b). This region was tested and successfully amplified by conventional PCR giving a single amplicon using gDNA from L. passim cultures (data not shown).

Trypanosomatid α-tubulin amplification in multiple insect hosts. a Illustration of the locus of trypanosomatid alpha/beta (α/β) tubulin tandem repeated arrays. As highlighted in dark blue, the 193-bp amplicon is located at the 3´end of each α-tubulin repeat, being the number of repeats variable across trypanosomatid species. b Clustal analysis of the 3′ end of the α-tubulin open reading frame showing conserved blocks of homology (in blue) that were used for designing primers and TaqMan probes. c Representative 2% agarose gel electrophoresis of the 193 α-tubulin trypanosomatid region amplified in samples of bumblebees, wild bees and grasshoppers. Numbers refer to different samples. The negative control (C-) and the positive control (C+) show negative and positive samples, respectively. F, Forward; FAM, dye label; M, molecular size marker; MGB, minor groove binder; R, reverse
To assay the performance of the 193-bp α-tubulin region as target for the PCR in various host species, a total of 43 samples comprising 27 wild bee, nine bumblebee and seven grasshopper species were tested for the presence of trypanosomatid parasites (Additional file 1: Table S2; Fig. 1c). The α-tubulin fragment was amplified in 21 out of 43 samples (48.8% of the examined samples), being found in 11 out of the 20 insect species analyzed (Additional file 1: Table S2). The positivity of the PCR was 42.8% (12/28) in solitary bees, 87.5% (7/8) in bumblebees and 28% (2/7) in grasshoppers (Additional file 1: Table S2). Of the positive samples, C. bombi (percentage of identity = 100%) from five different Bombus terrestris samples and Lotmaria passim (percentage of identity = 99.8%) from a bee of genus Halictus were successfully amplified and sequenced using a fragment of the SSU rRNA [42]. These data support the use of the 193-bp α-tubulin gene locus as a pan-specific molecular target for trypanosomatid identification.
Analytical sensitivity of the α-tubulin TaqMan assay
Once the capacity of the α-tubulin TaqMan assay to amplify the 193-bp α-tubulin region in multiple insect hosts was verified, we used this region as a molecular target in “pseudo-infected” bee gDNA spiked with decreased concentrations of Lotmaria passim or Leishmania major DNA (1:10 dilution), to ascertain the efficiency and sensitivity of the TaqMan qPCR assay in different trypanosomatid species. The efficiency of the standard curve using L. passim gDNA was 99.7%, with R2 = 0.999 (y = − 3.328x + 39.395) and the last detected Ct of 35.72 ± 0.1 (Additional file 1: Figure S3A). The analytical sensitivity of the assay resulted in a limit of detection (LOD) of 24 α-tubulin copies/µl (= 1 cell equivalent/µl). The efficiency of the standard curve using L. major gDNA was 94.2%, with R2 = 0.996 (y = − 3.469x + 42.472) and the last detected Ct of 33.8 ± 0.1 (Additional file 1: Figure S3B). The analytical sensitivity of the assay resulted in a LOD of 240 α-tubulin copies/µl (= 10 cell equivalent/µL).
Trypanosomatid infection levels in field-collected insect samples.
To determine the analytical power of the TaqMan assay in different insect samples, we determined trypanosomatid infection levels (cell equivalents/bee) in honeybees and other pollinators. Honeybee samples (n = 224 samples) from four apiaries located in the south of Granada Province and in flowering plants in Granada city were collected, and trypanosomatid loads were quantified by qPCR. Trypanosomatids were detected in 2.68% (6/224) of the honeybee samples examined. Of the four apiaries analyzed, two apiaries (apiaries 1 and 2) were infected with trypanosomatids, with a positivity percentage ranging from 4.7% (1/21; colony 2A) to 13.3% (4/30; colony 1A). In comparison, trypanosomatid prevalence in honeybees feeding on flowering plants in Granada city was 4% (2/50). The quantification of the parasitemia varied from 40.6 to 7.7 × 107 trypanosomatids per honeybee, which shows the disparity in infection levels found in nature (Fig. 2). The two positive bees analyzed from flowering plants in Granada city showed a high parasite loads of 2.3 × 107 and 1.5 × 107 parasites per bee, respectively (Fig. 2). The amplification and sequencing of the SSU rRNA [42] was only possible in three of the honeybee samples analyzed that tested positive for trypanosomatids, revealing the uniform presence of Lotmaria passim with a percentage of identity of 99.57% and 99.71% respectively.

Trypanosomatid yields in the insect samples analyzed with the α-tubulin TaqMan assay. Data are presented as a violin plot showing the density of cell equivalents per insect. The red circles represent 8 infected samples of the western honey bee Apis mellifera, the yellow squares represent 4 infected samples of the buff-tailed bumblebee Bombus terrestris, the blue cross represents an infected sample of Halictus spp. and the blue triangle represents an infected white-banded digger bee Amegilla quadrifasciata. The blue dotted line represents 1 × 104 cell equivalents per insect [Log4(10)] and 1 × 106 cell equivalents per insect [Log6(10)]
To ascertain the capacity to measure trypanosomatid yields in other insect species, a selected group of insect hosts was chosen for this study. The quantification resulted in successful quantifications with variable yields ranging from 1.8 × 103 parasites per sample for Amegilla quadrifasciata to the highest parasite loads in Bombus terrestris of 7.1 × 107 parasites per insect (Fig. 2).
Discussion
Trypanosomatid parasites are one of the most extended groups of parasites, with at least 24 genera, showing a high plasticity for thriving in insect hosts in many different ecological scenarios [1]. Determination of the prevalence of trypanosomatids in nature requires sensitive molecular tools for evaluating the ecology and distribution of this group of parasites. In the present study, we found trypanosomatid infection in 11 out of the 20 insect species examined based on amplification of the α-tubulin loci in a standard PCR, with an overall positivity of 48.8%. These results demonstrate the potential of this qPCR assay for analyzing trypanosomatid biodiversity in insect hosts. Among the insects found competent to harbor trypanosomatids, we found both solitary bees (genus Anthophora, Amegilla spp. and Lasioglossum immunitum) and social bees (Halictus spp., Apis mellifera and B. terrestris) and also non-hymenopteran insects such as grasshoppers (Calliptamus italicus). Although there is a wide variety in the functional traits of the insects studied, such as body size, phenology, nesting location, sociality and/or diet, which could influence pathogen prevalence [43], our data support earlier findings that trypanosomatid infection [38, 43,44,45] can occur in both social and solitary species. For example, the α-tubulin TaqMan assay described herein detected for the first time trypanosomatid infection in bees belonging to the genus Amegilla, namely in a wild solitary bee also known as the white-banded Digger Bee. This medium-sized solitary bee usually looks for sunny slopes in sandy areas in various habitats and feeds on plants of the Verbenaceae, Lamiaceae and Boraginaceae families. Bees belonging to genus Amegilla have a wide distribution and can be found throughout Spain [46]. We also detected trypanosomatid infection in the grasshopper C. italicus. To our knowledge, only one study has shown the capacity of Leishmania hertigi to infect individuals of the order Orthoptera (Schistocerca gregaria), thereby inducing agglutination reactions in the hemolymph and further demonstrating that trypanosomes are capable of inducing defense reactions in this host [47].
Multicopy or repetitive gene sequences are generally used to increase the sensitivity of PCR assays [27, 28]. In the present study, the α-tubulin TaqMan assay had a detection limit of 24 copies of α-tubulin/µl, which represents 1 parasite equivalent/µl. Molecular methods for detecting multiple trypanosomatid species in insect samples have been developed using conserved regions of small and large subunits of rRNA genes [42], ITS sequences flanking the 5.8S rRNA gene [48], the SL-region or the 28S–large subunit (LSU) of the rRNA [29, 49]. Trypanosome nuclear genomes also contain an array of alternating α- and β-tubulin genes that can carry up to 19 copies per haploid genome in T. brucei [35]. The region between the α- and β-tubulin locus has been successfully used to design nested PCR for evaluating trypanosome infections in cattle [50]. However, however, there has been no report of a TaqMan qPCR assay for trypanosomatid detection and quantification based on this region. Thus far, TaqMan assays have been successfully applied for the diagnosis and quantification of specific trypanosomatid species in clinical samples. For example, infections with T. cruzi have been determined in infected human blood samples using kinetoplast DNA (kDNA) and nuclear satellite DNA targets with detection limits of 0.23 and 0.70 parasite equivalents/ml, respectively [51]. Leishmania species have been detected with a sensitivity of three copies of parasite per reaction using the mitochondrial cytochrome c oxidase I (COI; 10–20 copies in each mitochondrion) or the amino acid permease 3 (AAP3) (2 copies in the parasite genome) genes [52]. To date, few attempts have been made to obtain multispecies identification using TaqMan qPCR. For example, in two studies, the 28S, 18S and 5S rRNA targets showed a limit of 1000 to 100 trypanosomes per milliliter of blood in a diverse set of trypanosomatids; however, the assay was only tested in animal blood samples and was not validated for the detection of monoxenous trypanosomatid parasites in insect samples [29, 30]. The application of the α-tubulin TaqMan assay described in this article represents a reliable sensitive assay which would be an effective alternative method in that it showed a good dynamic range of detection in L. passim and L. major down to between 1 and 10 cell equivalents/µl, thereby allowing for the detection of low levels of circulating monoxenous and dixenous parasites in nature.
The quantitative measurement of parasite loads has important implications not only for assessing host health but also for evaluating changing host behavioral patterns, such as differences in secondary sex and male choice in birds or differences in plastic transmission between hosts and insect vectors. The effects exerted by trypanosomatids once inside their hosts can be many and diverse, from asymptomatic to symptomatic effects, depending on the species, the host and other factors, such as the presence of co-infections with other pathogens or physicochemical changes in the environment of the host [4]. For example, Trypanosoma brucei triggers the innate immune response and modifies feeding behavior in tsetse flies [53, 54], Leishmania spp. facilitates morphological changes in the stomodeal valve of sand fly hosts [55] and Blastocrithidia gerrides generates male skating endurance in water striders [56]. Since bees are threatened by different parasitic pests[13], the quantitative measurement of trypanosomatid yields could be an indicator of colony health status. In the present study we found two separate groups of parasite loads of insect hosts with parasite numbers < 1 × 104 parasites and > 1 × 106 trypanosomatids per insect, with the yields particularly high in three of the four samples of bumblebees. These data show the variability of infection intensities in the hosts, possibly reflecting different transmission capacities. In addition, our study shows the presence of L. passim in Apis mellifera and Halictus spp. The capacity for sharing hosts has been postulated as a potential driver of parasite spillover or spillback between wild bees and managed bees and is an important parasite trait that could influence parasite disease emergence [57]. Although the precise symptoms and pathology of trypanosomatids in bees are still unclear, it has been reported that honey bees Osmia cornuta and Apis mellifera experimentally infected with 2.5 × 104 cells/bee and 4 × 104 cells/bee of C. mellificae and L. passim showed increased mortality rates compared to uninoculated controls [26, 45], with parasite loads shown to range from 2 × 104 parasites/bee up to 1 × 106 parasites/bee using SYBR green qPCR [58, 59]. These data indicate that it is essential to monitor trypanosomatids in order to know the possible risks to infected honeybee colonies and to implement possible control measures in parasitized apiaries.
In conclusion, the use of highly sensitive and quantitative methods for trypanosomatid detection are crucial for detecting, controlling and preventing the spreading of trypanosomes and could help in determining the competence of insect hosts for transmitting such parasites and/or the impact of biotic and/or abiotic factors in pathogen dynamics. Based on the results of the present study, the α-tubulin TaqMan qPCR assay could be a useful analytical tool to ascertain the infection status in apiaries and other natural and experimental settings, allowing for the detection of trypanosomatid parasites in numerous niches and hosts in nature.
Availability of data and materials
All data analyzed during this study are included in this published article.
Change history
17 March 2023
This article has been corrected since original publication; please see the linked erratum for further details.
16 March 2023
A Correction to this paper has been published: https://doi.org/10.1186/s13071-023-05723-2
References
Frolov AO, Kostygov AY, Yurchenko V. Development of monoxenous trypanosomatids and phytomonads in insects. Trends Parasitol. 2021;37:538–51.
Maslov DA, Opperdoes FR, Kostygov AY, Hashimi H, Lukeš J, Yurchenko V. Recent advances in trypanosomatid research: genome organization, expression, metabolism, taxonomy and evolution. Parasitology. 2019;146:1–27.
de Almeida Dias F, da Costa Vasconcellos LR, Attias M, Souto-Padrón TC, Lopes AH. Transovum transmission of trypanosomatid cysts in the milkweed bug, Oncopeltus fasciatus. PLoS ONE. 2014;9:e108746.
Lukeš J, Butenko A, Hashimi H, Maslov DA, Votýpka J, Yurchenko V. Trypanosomatids are much more than just trypanosomes: clues from the expanded family tree. Trends Parasitol. 2018;34:466–80.
Maslov DA, Votýpka J, Yurchenko V, Lukeš J. Diversity and phylogeny of insect trypanosomatids: all that is hidden shall be revealed. Trends Parasitol. 2013;29:43–52.
Arismendi N, Castro MP, Vargas M, Zapata C, Riveros G. The trypanosome Lotmaria passim prevails in honey bees of different ages and stages of development. J Apic Res. 2020;61:63–9.
Hamilton PT, Votýpka J, Dostálová A, Yurchenko V, Bird NH, Lukeš J, et al. Infection dynamics and immune response in a newly described Drosophila -trypanosomatid association. mBio. 2015;6:e01356-15.
Schoener E, Uebleis SS, Cuk C, Nawratil M, Obwaller AG, Zechmeister T, et al. Trypanosomatid parasites in Austrian mosquitoes. PLoS ONE. 2018;13:e0196052.
Ebbert MA, Burkholder JJ, Marlowe JL. Trypanosomatid prevalence and host habitat choice in Woodland Drosophila. J Invertebr Pathol. 2001;77:27–32.
Králová J, Grybchuk-Ieremenko A, Votýpka J, Novotný V, Kment P, Lukeš J, et al. Insect trypanosomatids in Papua New Guinea: high endemism and diversity. Int J Parasitol. 2019;49:1075–86.
Votýpka J, Kment P, Kriegová E, Vermeij MJA, Keeling PJ, Yurchenko V, et al. High prevalence and endemism of trypanosomatids on a small Caribbean island. J Eukaryot Microbiol. 2019;66:600–7.
Votýpka J, Petrželková KJ, Brzoňová J, Jirků M, Modrý D, Lukeš J. How monoxenous trypanosomatids revealed hidden feeding habits of their tsetse fly hosts. Folia Parasitol (Praha). 2021;68:2021.019. https://doi.org/10.14411/fp.2021.019.
Goulson D, Nicholls E, Botias C, Rotheray EL. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 2015;347:1255957–1255957.
Bartolomé C, Jabal-Uriel C, Buendía-Abad M, Benito M, Ornosa C, De la Rúa P, et al. Wide diversity of parasites in Bombus terrestris (Linnaeus, 1758) revealed by a high-throughput sequencing approach. Environ Microbiol. 2021;23:478–83.
Gabín-García LB, Bartolomé C, Guerra-Tort C, Rojas-Nossa SV, Llovo J, Maside X. Identification of pathogens in the invasive hornet Vespa velutina and in native Hymenoptera (Apidae, Vespidae) from SW-Europe. Sci Rep. 2021;11:11233.
Schwarz RS, Bauchan GR, Murphy CA, Ravoet J, de Graaf DC, Evans JD. Characterization of two species of trypanosomatidae from the honey bee Apis mellifera: Crithidia mellificae Langridge and McGhee, and Lotmaria passim n. gen., n. sp. J Eukaryot Microbiol. 2015;62:567–83.
Arismendi N, Bruna A, Zapata N, Vargas M. PCR-specific detection of recently described Lotmaria passim (Trypanosomatidae) in Chilean apiaries. J Invertebr Pathol. 2016;134:1–5.
Castelli L, Branchiccela B, Invernizzi C, Tomasco I, Basualdo M, Rodriguez M, et al. Detection of Lotmaria passim in Africanized and European honey bees from Uruguay Argentina and Chile. J Invertebr Pathol. 2019;160:95–7. https://doi.org/10.1016/j.jip.2018.11.004.
Cepero A, Ravoet J, Gómez-Moracho T, Bernal J, Del Nozal MJ, Bartolomé C, et al. Holistic screening of collapsing honey bee colonies in Spain: a case study. BMC Res Notes. 2014;7:649.
Pislak Ocepek M, Toplak I, Zajc U, Bevk D. The pathogens spillover and incidence correlation in bumblebees and honeybees in Slovenia. Pathogens. 2021;10:884.
Ravoet J, Maharramov J, Meeus I, De Smet L, Wenseleers T, Smagghe G, et al. Comprehensive bee pathogen screening in belgium reveals Crithidia mellificae as a new contributory factor to winter mortality. PLoS ONE. 2013;8:e72443.
Regan T, Barnett MW, Laetsch DR, Bush SJ, Wragg D, Budge GE, et al. Characterisation of the British honey bee metagenome. Nat Commun. 2018;9:4995.
Ribani A, Utzeri VJ, Taurisano V, Galuppi R, Fontanesi L. Analysis of honey environmental DNA indicates that the honey bee (Apis mellifera L.) trypanosome parasite Lotmaria passim is widespread in the apiaries of the North of Italy. J Invertebr Pathol. 2021;184:107628.
Meeus I, Brown MJF, De Graaf DC, Smagghe G. Effects of invasive parasites on bumble bee declines: parasites and bumble bee declines. Conserv Biol. 2011;25:662–71.
Schlüns H, Sadd BM, Schmid-Hempel P, Crozier RH. Infection with the trypanosome Crithidia bombi and expression of immune-related genes in the bumblebee Bombus terrestris. Dev Comp Immunol. 2010;34:705–9.
Gómez-Moracho T, Buendía-Abad M, Benito M, García-Palencia P, Barrios L, Bartolomé C, et al. Experimental evidence of harmful effects of Crithidia mellificae and Lotmaria passim on honey bees. Int J Parasitol. 2020;50:1117–24.
Edwards SG, O’Callaghan J, Dobson ADW. PCR-based detection and quantification of mycotoxigenic fungi. Mycol Res. 2002;106:1005–25.
González-Salgado A, Patiño B, Gil-Serna J, Vázquez C, González-Jaén MT. Specific detection of Aspergillus carbonarius by SYBR Green and TaqMan quantitative PCR assays based on the multicopy ITS2 region of the rRNA gene. FEMS Microbiol Lett. 2009;295:57–66.
González-Andrade P, Camara M, Ilboudo H, Bucheton B, Jamonneau V, Deborggraeve S. Diagnosis of trypanosomatid infections. J Mol Diagn. 2014;16:400–4.
Medkour H, Varloud M, Davoust B, Mediannikov O. New molecular approach for the detection of Kinetoplastida parasites of medical and veterinary interest. Microorganisms. 2020;8:356.
Sádlová J, Podešvová L, Bečvář T, Bianchi C, Gerasimov ES, Saura A, et al. Catalase impairs Leishmania mexicana development and virulence. Virulence. 2021;12:852–67.
Goto Y, Kuroki A, Suzuki K, Yamagishi J. Draft genome sequence of Leishmania tarentolae parrot Tar II, obtained by single-molecule real-time sequencing. Microbiol Resour Announc. 2020;9:e00050-e120.
Ramírez CA, Requena JM, Puerta CJ. Alpha tubulin genes from Leishmania braziliensis: genomic organization, gene structure and insights on their expression. BMC Genomics. 2013;14:454.
Flegontov P, Butenko A, Firsov S, Kraeva N, Eliáš M, Field MC, et al. Genome of Leptomonas pyrrhocoris: a high-quality reference for monoxenous trypanosomatids and new insights into evolution of Leishmania. Sci Rep. 2016;6:23704.
Ersfeld K, Asbeck K, Gull K. Direct visualisation of individual gene organisation in Trypanosoma brucei by high-resolution in situ hybridisation. Chromosoma. 1998;107:237–40.
Vejnovic B, Stevanovic J, Schwarz RS, Aleksic N, Mirilovic M, Jovanovic NM, et al. Quantitative PCR assessment of Lotmaria passim in Apis mellifera colonies co-infected naturally with Nosema ceranae. J Invertebr Pathol. 2018;151:76–81.
Ravoet J, Schwarz RS, Descamps T, Yañez O, Tozkar CO, Martin-Hernandez R, et al. Differential diagnosis of the honey bee trypanosomatids Crithidia mellificae and Lotmaria passim. J Invertebr Pathol. 2015;130:21–7.
Buendía-Abad M, Higes M, Martín-Hernández R, Barrios L, Meana A, Fernández Fernández A, et al. Workflow of Lotmaria passim isolation: experimental infection with a low-passage strain causes higher honeybee mortality rates than the PRA-403 reference strain. Int J Parasitol Parasites Wildl. 2021;14:68–74.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23:2947–8.
Brankatschk R, Bodenhausen N, Zeyer J, Bürgmann H. Simple absolute quantification method correcting for quantitative PCR efficiency variations for microbial community samples. Appl Environ Microbiol. 2012;78:4481–9.
Runckel C, DeRisi J, Flenniken ML. A draft genome of the honey bee trypanosomatid parasite Crithidia mellificae. PLoS ONE. 2014;9:e95057.
Maslov DA, Lukeš J, Jirku M, Simpson L. Phylogeny of trypanosomes as inferred from the small and large subunit rRNAs: implications for the evolution of parasitism in the trypanosomatid protozoa. Mol Biochem Parasitol. 1996;75:197–205.
Figueroa LL, Compton S, Grab H, McArt SH. Functional traits linked to pathogen prevalence in wild bee communities. Sci Rep. 2021;11:7529.
Schmid-Hempel P, Wilfert L, Schmid-Hempel R. Pollinator diseases: the Bombus-Crithidia system. In: Wilson K, Fenton A, Tompkins D, editors. Wildlife disease ecology. 1st ed. Cambridge: Cambridge University Press; 2019. p. 3–31.
Strobl V, Yañez O, Straub L, Albrecht M, Neumann P. Trypanosomatid parasites infecting managed honeybees and wild solitary bees. Int J Parasitol. 2019;49:605–13.
Molina C, Bartomeus I. Guía de campo de las abejas de España. 1a edición. Almenara: Tundra; 2019.
Ingram GA, East J, Molyneux DH. Naturally occurring agglutinins against trypanosomatid flagellates in the haemolymph of insects. Parasitology. 1984;89:435–51.
Gaithuma AK, Yamagishi J, Martinelli A, Hayashida K, Kawai N, Marsela M, et al. A single test approach for accurate and sensitive detection and taxonomic characterization of Trypanosomes by comprehensive analysis of internal transcribed spacer 1 amplicons. PLoS Negl Trop Dis. 2019;13:e0006842.
Medkour H, Varloud M, Davoust B, Mediannikov O. New molecular approach for the detection of Kinetoplastida parasites of medical and veterinary interest. Microorganisms. 2020;8:356.
Ofori JA, Bakari SM, Bah S, Kolugu MK, Aning GK, Awandare GA, et al. A longitudinal two-year survey of the prevalence of trypanosomes in domestic cattle in Ghana by massively parallel sequencing of barcoded amplicons. PLoS Negl Trop Dis. 2022;16:e0010300.
Ramírez JC, Cura CI, da Cruz MO, Lages-Silva E, Juiz N, Velázquez E, et al. Analytical validation of quantitative real-time PCR methods for quantification of Trypanosoma cruzi DNA in blood samples from Chagas disease patients. J Mol Diagn. 2015;17:605–15.
Fotouhi-Ardakani R, Ghafari SM, Ready PD, Parvizi P. Developing, modifying, and validating a TaqMan real-time PCR technique for accurate identification of Leishmania Parasites causing most Leishmaniasis in Iran. Front Cell Infect Microbiol. 2021;11:731595.
Geiger A, Ponton F, Simo G. Adult blood-feeding tsetse flies, trypanosomes, microbiota and the fluctuating environment in sub-Saharan Africa. ISME J. 2015;9:1496–507.
Van Den Abbeele J, Caljon G, De Ridder K, De Baetselier P, Coosemans M. Trypanosoma brucei Modifies the Tsetse salivary composition, altering the fly feeding behavior that favors parasite transmission. PLoS Pathog. 2010;6:e1000926.
Volf P, Hajmova M, Sadlova J, Votypka J. Blocked stomodeal valve of the insect vector: similar mechanism of transmission in two trypanosomatid models. Int J Parasitol. 2004;34:1221–7.
Arnqvist G, Mäki M. Infection rates and pathogenicity of trypanosomatid gut parasites in the water strider Gerris odontogaster (Zett) (Heteroptera: Gerridae). Oecologia. 1990;84:194–8.
Graystock P, Blane EJ, McFrederick QS, Goulson D, Hughes WOH. Do managed bees drive parasite spread and emergence in wild bees? Int J Parasitol Parasites Wildl. 2016;5:64–75.
Stevanovic J, Schwarz RS, Vejnovic B, Evans JD, Irwin RE, Glavinic U, et al. Species-specific diagnostics of Apis mellifera trypanosomatids: a nine-year survey (2007–2015) for trypanosomatids and microsporidians in Serbian honey bees. J Invertebr Pathol. 2016;139:6–11.
Xu G, Palmer-Young E, Skyrm K, Daly T, Sylvia M, Averill A, et al. Triplex real-time PCR for detection of Crithidia mellificae and Lotmaria passim in honey bees. Parasitol Res. 2018;117:623–8.
Acknowledgements
We truly appreciate all the support and ideas given by Professor Antonio Osuna Carrillo de Albornoz for the completion of this paper and the members of the CTS-183 research team.
Funding
This work was supported by the Spanish Programme for Knowledge Generation and Scientific and Technological Strengthening of the R+D+I System: Proyecto generación del Conocimiento 2018 (PGC2018- 098929-A-I00) and proyecto PID2021.126938OB.I00 financiado por MCIN/ AEI /10.13039/501100011033 y por FEDER Una manera de hacer Europa.
Author information
Authors and Affiliations
Contributions
OBG, JSP, FJO and LMDP designed the experiment. LMDP, AVM, MZ and FJO collected the samples. OBG, JCDP, JSP and LMDP developed the assays. LMDP and TGM prepared the figures. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Location of the samples. Sampling was performed in 4 locations of Alpujarra region, south of Granada (Andalusia region, Spain). The figures were designed using mapchart software (https://www.mapchart.net/index.html) and Google maps (https://www.google.es/maps/?hl=es). Figure S2. Experimental design and sampling. Honeybee samples were collected from 4 apiaries (coded as 1-4) at Capileira, Torvizcón and Las Barreras, Alpujarra region locations situated at the Alpujarra region (south of Granada, Spain). Two random hives in each apiary (coded as A and B) were sampled. Figure S3. The analytical performance of the α-tub TaqMan assay in different trypanosomatid species was measured using standard amplification curves and linear regression curves. The efficiency and limits of detection were obtained using 7 serial fold dilutions of bee gDNA spiked with 2.4 × 107 copies/ul of L. passim down to 2.4 copies/µl of L. passim (A) or L. major (B) α-tubulin. Table S1. Number of honeybees collected from each hive at the different apiaries in Granada. Table S2. Wildbees, bumblebees and grasshoppers analyzed for the presence of trypanosomatid parasites. Table S3. Primers and probes sequences for qPCR assay to detect trypanosomatid parasites and insect DNA as an internal control.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Barranco-Gómez, O., De Paula, J.C., Parada, J.S. et al. Development of a TaqMan qPCR assay for trypanosomatid multi-species detection and quantification in insects. Parasites Vectors 16, 69 (2023). https://doi.org/10.1186/s13071-023-05687-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05687-3