
Abstract
Background
With the largest cattle population in Africa and vast swathes of fertile lands infested by tsetse flies, trypanosomosis is a major challenge for Ethiopian farmers. Managing the problem strategically and rationally requires comprehensive and detailed information on disease and vector distribution at the national level. To this end, the National Institute for Control and Eradication of Tsetse and Trypanosomosis (NICETT) developed a national atlas of tsetse and African animal trypanosomosis (AAT) for Ethiopia.
Methods
This first edition of the atlas focused on the tsetse-infested areas in western Ethiopia. Data were collected between 2010 and 2019 in the framework of national surveillance and control activities. Over 88,000 animals, mostly cattle, were tested with the buffy-coat technique (BCT). Odour-enhanced traps were deployed in approximately 14,500 locations for the entomological surveys. Animal- and trap-level data were geo-referenced, harmonized and centralized in a single database.
Results
AAT occurrence was confirmed in 86% of the districts surveyed (107/124). An overall prevalence of 4.8% was detected by BCT in cattle. The mean packed cell volume (PCV) of positive animals was 22.4, compared to 26.1 of the negative. Trypanosoma congolense was responsible for 61.9% of infections, T. vivax for 35.9% and T. brucei for 1.7%. Four tsetse species were found to have a wide geographic distribution. The highest apparent density (AD) was reported for Glossina pallidipes in the Southern Nations, Nationalities and People's Region (SNNPR) (3.57 flies/trap/day). Glossina tachinoides was the most abundant in Amhara (AD 2.39), Benishangul-Gumuz (2.38), Gambela (1.16) and Oromia (0.94) regions. Glossina fuscipes fuscipes and G. morsitans submorsitans were detected at lower densities (0.19 and 0.42 respectively). Only one specimen of G. longipennis was captured.
Conclusions
The atlas establishes a reference for the distribution of tsetse and AAT in Ethiopia. It also provides crucial evidence to plan surveillance and monitor control activities at the national level. Future work on the atlas will focus on the inclusion of data collected by other stakeholders, the broadening of the coverage to tsetse-free areas and continuous updates. The extension of the atlas to data on control activities is also envisaged.
Graphical Abstract

Similar content being viewed by others

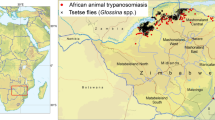

Background
Ethiopia has the largest livestock population in Africa, with an estimated 70.3 million cattle, 42.9 million sheep, 52.5 million goats, 11.3 million equines (2.1 horses, 8.9 donkeys and 0.3 mules) and 7.3 million camels [1]. Livestock resources contribute 45% of the agricultural gross domestic product (GDP) and 19% of the total GDP [2]. Further development of the livestock sector is one of the key targets of the Growth and Transformation Plan (GTP), the development strategy aiming to project Ethiopia to middle-income country status by 2025. However, the livestock sector falls far short of achieving its full potential. High disease prevalence, inadequate feed supply, poor genetic resources and poor marketing are the main bottlenecks for the development of the livestock sector in Ethiopia [3].
African animal trypanosomosis (AAT), also known as ‘nagana’, is one of the major livestock diseases constraining agricultural production in sub-Saharan Africa [4, 5]. AAT is caused by unicellular parasitic protozoa called trypanosomes, which are transmitted by the bite of hematophagous tsetse flies (Genus: Glossina). Among the many existing species of trypanosomes, Trypanosoma vivax, T. congolense and T. brucei have a particular economic relevance in livestock, and in particular in cattle [6]. Tsetse flies also transmit human African trypanosomosis (HAT), which is caused by two subspecies of T. brucei (i.e. T. brucei gambiense and T. brucei rhodesiense) [7]. In addition to tsetse flies, which are the sole cyclical or biological vectors of trypanosomosis, AAT can also be mechanically transmitted by other blood-sucking arthropods such as Tabanids and Stomoxys [8]. Notably, the mechanical mode of transmission enabled T. vivax to spread beyond sub-Saharan Africa and to become endemic also in Latin America [9].
Ethiopia is geographically located at the northeastern limit of the African tsetse belt [10]. Bioclimatic factors restrict tsetse distribution to the western part of the country, with a maximum longitude of 38 degrees East and a maximum latitude of 12° North [11, 12]. The lowlands bordering Sudan and South Sudan provide tsetse with favourable environmental conditions, but the flies have also progressively spread into the long, often steep river valleys carved into the massifs and plateaux of central Ethiopia [13, 14]. Further eastward spread is constrained by the low temperatures characterizing the Ethiopian highlands, but also by the semidesertic climate of the eastern lowlands. In the 1970s, the altitude of 1600 m was assumed to be the breeding limit for tsetse in Ethiopia [11], but in the following decades flies have also been captured at altitudes that are close to 2000 m [14]. Based on the shifting altitude limits, and depending on whether breeding areas only or areas of dispersal are considered, the area of tsetse infestation in Ethiopia has been variously estimated over the years between 66,000 km2 and 220,000 km2 [13]. However, in the absence of recent and comprehensive data on tsetse occurrence, these estimates should be considered as ballpark figures.
At the time of developing the atlas described in this article, which includes data for the 10-year period 2010–2019, five regional states were affected by tsetse flies in Ethiopia. These were, from north to south and anti-clockwise, Amhara, Benishangul-Gumuz, Oromia, Gambela and the Southern Nations, Nationalities and People's Region (SNNPR). After 2019, additional regional states were established in Ethiopia, but in this paper all results and maps are based on the regional divisions that were in effect in the period of data collection between 2010 and 2019.
Within the five regions mentioned above, four river basins or hydrological systems influence tsetse distribution in the country: the Abay (Blue Nile)/Didesa, Baro/Akobo, Gibe/Omo and Rift Valley (Fig. 1).

River basins influencing the geographic distribution of tsetse flies in western Ethiopia: Abay (Blue Nile)/Didesa, Baro/Akobo, Gibe/Omo and Rift Valley
Six species of tsetse flies were historically recorded in Ethiopia: Glossina pallidipes and G. morsitans submorsitans of the savannah/morsitans group, G. fuscipes fuscipes and G. tachinoides of the riverine/palpalis group and G. longipennis and G. brevipalpis of the forest/fusca group [10, 15]. Of these, only the four species of the savannah and riverine groups have high economic importance and wide geographic distribution in the country [12, 13].
Compared to tsetse flies, AAT has a broader area of occurrence [13, 16]. This is especially the case for T. vivax, which is also reported from tsetse-free regional states such as Tigray and Afar [17,18,19], and it is believed to occur throughout the country [13]. In Ethiopia, the presence of T. vivax beyond the tsetse belt is mainly ascribed to mechanical transmission by non-tsetse vectors [20] and possibly also to the movement of animals between tsetse-free and tsetse-infested areas. Similar epizootic patterns are observed for T. vivax in several African countries [21, 22].
The severity of the AAT problem in Ethiopia is difficult to overstate. In particular, as of the 1970s, resettlement programmes moved large numbers of farmers into tsetse-infested areas, where they had to contend with the trypanosomosis challenge ever since [13]. The extremely high reliance of Ethiopian agriculture on draught-oxen also meant that, in the resettlement areas, trypanosomosis critically constrained crop production. Today, smallholders in tsetse-infested areas widely consider AAT as the major animal disease they have to grapple with. This has been well documented in SNNPR [23,24,25,26], western Oromia [25, 27, 28], western Amhara [29], Benishangul-Gumuz [30] and Gambela [25]. Beyond the tsetse belt, a severe AAT problem is also reported by farmers in Tigray and in the tsetse-free areas of Amhara, including an important problem of both curative and prophylactic drug misuse and drug resistance [18, 20, 31, 32]. Indeed, the use of drugs is often the only trypanosomosis control tool available to farmers [33], and drug usage can be extremely high. Six treatments per year are frequent [34], and up to 20 treatments in a year was reported for high-value oxen [35]. As opposed to AAT, HAT is not a major disease in Ethiopia [36,37,38,39,40].
Because of the persisting challenge of tsetse and AAT, in 2013 the government of Ethiopia established the National Institute for Control and Eradication of Tsetse and Trypanosomosis (NICETT), a specialized national structure under the Ministry of Agriculture. NICETT’s mandate included the coordination of activities against tsetse and trypanosomosis at the national level, and its mission was to make affected areas free of the problem. Its ultimate goal was to increase the production and productivity of livestock and crops and to enhance food security and self-sufficiency of the affected populations. The institute was endowed with core staffing (approximately 300 members) and technical capacities. The overall government funding for NICETT was approximately 2.4 M USD per year (period 2018–2019), which was proof of Ethiopia’s commitment to the progressive control of trypanosomosis [41]. NICETT infrastructure included a central office and a tsetse mass-rearing facility in Kaliti (Addis Ababa) [42]. Furthermore, four regional offices were strategically placed in the tsetse-infested areas. These offices are located in Finote Selam (Amhara), Asossa (Benishangul-Gumuz), Bedelle (Oromia) and Arba Minch (SNNPR) (Fig. 2).

Animal Health Institute in Ethiopia, including the head office in Sebeta and its five Centres in Finote Selam (Amhara), Asossa (Benishangul-Gumuz), Kaliti (Addis Ababa), Bedelle (Oromia) and Arba Minch (SNNPR)
One of NICETT’s main responsibilities was to collect field data on the occurrence of tsetse and trypanosomosis. This information is crucial to target control activities at the national level and to assess their impact. Entomological and parasitological surveys were carried out both to generate a baseline before the start of control operations and for monitoring purposes during and after interventions. Over the years, these surveys have produced a vast amount of data. However, the lack of a centralized information system has severely hindered data analysis. To address this gap, NICETT developed a national atlas of tsetse and AAT. The initiative was technically supported by the Food and Agriculture Organization of the United Nations (FAO) through two projects [43, 44], which were implemented in the framework of the Programme Against African Trypanosomosis (PAAT) [45].
In 2022, NICETT was merged with the National Animal Health Diagnostic and Investigation Centre (NAHDIC) to create the Animal Health Institute (AHI). AHI was established with three goals: (1) animal health related research and diagnosis; (2) tsetse and trypanosomosis control and prevention; (3) animal health advisory and training service. The merger aimed to enhance the research, control, diagnostic and coordination capacity of NICETT, since NAHDIC was a long-standing animal health diagnostic and referral centre for Ethiopia and for the eastern Africa region. All mandates, roles and functions of NICETT were transferred to AHI (regulation number 503/2022). After the merger, NICETT central office was moved to Sebeta, and the four regional offices and the Kaliti mass-rearing facility were converted into AHI centres. Two additional AHI centres are planned to be established in Gambela and the recently established South West Ethiopia Peoples’ Region.
Methods
The FAO continental atlas of tsetse and AAT provided the blueprint for the development of a national atlas in Ethiopia [12, 16]. National atlases previously developed in other countries (i.e. Sudan, Mali, Kenya, Zimbabwe and Burkina Faso) also provided additional methodological references [21, 46,47,48,49].
Input data
The data on tsetse and AAT occurrence presented in this paper were collected by NICETT between 2010 and 2019 in the tsetse-infested areas of western Ethiopia. Data were collected either before the start of control activities (i.e. baseline data) or for monitoring purposes during or after interventions.
African animal trypanosomosis data
NICETT carried out trypanosomosis surveillance through its regional branch offices. The diagnostic method of choice was the buffy-coat technique (BCT) [50], and all AAT data presented in this paper are based on this parasitological test. A clinical diagnosis of AAT is often made by regional veterinary laboratories, district veterinary services and veterinary extension workers. However, these actors rarely use confirmatory tests and therefore they did not provide relevant information for the atlas. Universities, academic and research institutions also collect data on AAT in the context of research [16]. However, these data are not normally transmitted to the central veterinary authorities, and therefore they were not included in this first edition of the national atlas. Community-based animal health workers and livestock keepers make tentative diagnoses for the purpose of local treatment animals with trypanocides.
In Ethiopia, trypanosomosis surveillance normally focuses on bovines because of the heavy economic burden of the disease in these animals [51]. As a result, > 99.6% of the animals tested in the period 2010–2019 were cattle (the remaining 0.4% being sheep, goats and equines).
During surveys, the field teams usually perform site selection together with the district technical teams. Animals were randomly selected, and data on individual animals were recorded in hard copy datasheets. These include the owner’s name, administrative units [i.e. regional state, zone, district (woreda) and peasant association (kebele)], position (GPS-measured altitude, latitude and longitude), date of the survey, animal species, breed (‘local’ or ‘cross’), colour, sex, age, body condition (‘good’, ‘medium’ or ‘poor), packed cell volume (PCV) and the detected trypanosome species (i.e. T. vivax, T. congolense, T. brucei and mixed-infections thereof). The hard copy datasheets generated by field teams were subsequently entered into digital spreadsheets. These were first assembled by regional offices and then transmitted to the central office on a monthly, quarterly or annual basis.
Tsetse fly data
Tsetse surveys were carried out by NICETT’s regional offices with technical support from animal health extension workers at the district and peasant association level. Regional laboratories occasionally participate in the entomological surveys, especially in the event of AAT outbreaks. Local communities are also engaged, and they support field staff by transporting materials, opening access roads, clearing trapping sites and looking after the traps and targets.
Several types of traps were used in the study period, including NGU/NG2G [52], monopyramidal [53], bipyramidal [54], monoconical [55] and biconical [56]; sticky panels were also used [57]. Fly attractants like acetone, octenol, phenol, 3-week-old cattle urine or a combination thereof are used to enhance attractiveness for tsetse. Traps were deployed in suitable habitats for the flies, and their individual location was geo-referenced with GPS. Grease was smeared at the bottom of the trap poles to prevent ants hunting the captured flies, and vegetation was cleared for visibility in a radius of 2 m around the trap. As a rule, traps were maintained in position for 2 or 3 days (i.e. 48 or 72 h) before being removed. Flies were then collected from the cages and counted. Their species and sex were identified and recorded.
Field teams registered data from individual traps using standard recording sheets [58]. Recorded information includes the administrative units, GPS-measured geographic coordinates, trap type, vegetation type, odour attractant, start and end date of trapping, number of tsetse flies captured (disaggregated by species and sex) and number of other biting flies captured.
The atlas development process
Capacity development provided by FAO was a key first step for the development of the atlas. The training focused on data collation, cleaning, harmonization, geo-referencing and handling. Another important enabling factor was the standardization of the field data recording formats, which were initially different across the centres. In terms of data flow, the field teams routinely capture data on paper recording sheets, which are subsequently entered into digital spreadsheets. The digital files are then transferred to the national office via email on a monthly, quarterly, semestral or yearly basis.
For the development of the atlas, special efforts were made systematically to collate existing data from decentralized centres and the head office. A digital repository was developed to centralize all input data. The repository is first organized by type of data (tsetse or AAT) and then by region, year, district and month of collection for easy data tracking, extraction and handling.
Following the data assembling in the repository, major efforts were devoted to data cleaning, harmonization and geo-referencing involving the head office, the centres and supported by FAO. Dates recorded in the Ethiopian calendar were converted to the Gregorian calendar. GPS-based geographic coordinates were harmonized to the GIS-ready standard ‘decimal degrees’. Potential data entry errors in the coordinates (i.e. outliers) were systematically checked in GIS and corrected if needed. Data were also checked for possible duplications.
The database
The atlas database is divided into two components: one for entomological data and one for AAT data. Each component is stored in a separate Microsoft Excel file. In both files, each record is linked to the respective source file in the data repository.
Tsetse database
For the tsetse database, each row represents a single trap. The related information includes the region, zone, district (woreda), peasant association (kebele), village (locality), trap identifier (trap ID), trap type, altitude, latitude, longitude, vegetation type, odour attractant, start and end date of trapping, duration of trapping, number of tsetse flies captured, tsetse species, tsetse sex, number of other biting flies captured (tabanids and Stomoxys) and the source data file. When a single trap captured more than one tsetse species, separate records are entered for each tsetse species. The abundance of flies, or AD, is also recorded, and it is expressed as number of flies/trap/day.
AAT database
For the AAT database, each row represents a single tested animal. The related information includes region, zone, district (woreda), peasant association (kebele), village (locality), altitude, latitude, longitude, date of the survey, animal identifier (ID), animal species, breed, colour, sex, age, body condition, PCV, the detected trypanosome species, the diagnostic method, sampling method and the source data file name and the path thereof for easy tracking.
Results
Figure 3 summarizes the atlas by showing the reported occurrence of tsetse flies and AAT for the period 2010–2019. Surveyed locations where tsetse and AAT were not detected are also included in the figure.

Reported geographic distribution of tsetse flies (genus Glossina) and African animal trypanosomosis in Ethiopia. Data collection period: 2010–2019
Figure 3 shows that the atlas provides a good coverage of the tsetse-infested areas in Ethiopia. Also, as field surveys normally address both tsetse and AAT at the same time, there is a close correspondence between the data coverage of the two components.
Tsetse flies
A total of 16,865 trapping events were assembled in the atlas. These trapping events originated from 14,498 different locations, for a total intensity of 45,820 trap days. The AD, as measured by the average number of flies captured per trap per day, was 3.1 flies/trap/day. The results of these surveys, disaggregated by region, are summarized in Table 1, while results disaggregated by zone and by district are in Additional file 1: S1 and Additional file 2: S2, respectively. The geographic distribution of the different tsetse species in western Ethiopia is shown in Fig. 4, while higher resolution maps are available in Additional file 3: S3.

Presence (coloured circles) and absence (surveyed but not detected, grey circles) of tsetse fly species in Ethiopia. Data collection period: 2010–2019
The envelope of tsetse distribution in Ethiopia is confirmed to extend from the westernmost part of the country in Gambela to 38.2° East and from the southernmost tip of SNNPR to 12.0° North. Tsetse are also shown to penetrate deeply into the valleys of the Gibe/Omo, Baro/Akobo and Abay/Didesa River systems.
Four species of tsetse flies are confirmed to have a broad geographic distribution in Ethiopia, i.e. G. pallidipes, G. morsitans submorsitans, G. fuscipes fuscipes and G. tachinoides. In some areas in the Oromia region, all four species were found in the same locations. Glossina longipennis was also captured, albeit only as a single specimen in the Gibe River basin. Glossina brevipalpis was not detected in this study period.
Glossina pallidipes was found to have the highest AD. In particular, in the SNNPR, G. pallidipes was detected at an average apparent density of 3.57 flies/trap/day. In addition to being broadly distributed across the SNNPR, G. pallidipes was also abundant in western Oromia (AD 0.81) and detected in small areas in Gambela. The species was not detected in Benishangul-Gumuz and Amhara. Looking at the river basins and hydrological systems, G. pallidipes was found in the Rift Valley, Gibe/Omo and Baro/Akobo as well as in the upper reaches of the Didesa River. For G. morsitans submorsitans, the other species of the savannah group present in Ethiopia, the broadest distribution and highest densities were detected in Benishangul-Gumuz (AD 0.84) and western Oromia (AD 0.73). The species was also present at lower densities in Gambela (AD 0.02) and Amhara (AD 0.02), while it was not detected in SNNPR. In terms of river basins, G. morsitans submorsitans was found in the Baro/Akobo, Abay/Didesa and upstream parts of the Gibe/Omo.
As to the tsetse species of the riverine group, G. fuscipes fuscipes was detected in western Oromia (AD 0.28), Gambela (AD 0.28) and SNNPR (AD 0.17). Its distribution is centred around the Baro/Akobo basin, but it extends further east and south into the Didesa and Gibe-Omo. For G. tachinoides, apparent densities were at their highest in Amhara (AD 2.39) and Benishangul-Gumuz (AD 2.38), but the species was also widely distributed in western Oromia (AD 0.94) and present in Gambela (AD 1.16).
African animal trypanosomosis
For the period 2010–2019, data on the testing of 88,331 animals were assembled in the national atlas. Since 99.6% of these were bovines (88,003), from here on results are presented for cattle only. The results disaggregated by region are summarized in Table 2, while the geographic distribution of the different trypanosome species is shown in Fig. 5. Results disaggregated by zone and by district are in Additional file 4: S4 and Additional file 5: S5, respectively.

Presence (coloured squares) and absence (surveyed but not detected, grey squares) of T. vivax, T. congolense and T. brucei in cattle as determined with the buffy-coat technique (BCT). Data collection period: 2010–2019
A total of 4241 animals were found positive for trypanosomal infection, for an overall prevalence of 4.82%. These figures, and those provided in Table 2, Additional file 4: S4 and Additional file 5: S5, include mixed infections with more than one species of trypanosome. With the exception of Gambela, where testing was limited, parasite rates were remarkably similar in the different regions. AAT was confirmed in all but one of the 34 surveyed zones and in 86% of the surveyed districts (107/124). The impact of AAT on the haematocrit was sizable, with the mean PCV of affected animals being 22.4 compared to 26.1 of negative animals.
Close to two thirds of all infections were caused by T. congolense (2685, 61.9%) and more than a third by T. vivax (1559, 35.9%). Despite this difference, the two species seem to have a similar geographic distribution in western Ethiopia, both having been detected across the surveyed areas (Fig. 5). Indeed, both T. vivax and T. congolense were found in virtually all surveyed zones (32/34). The remaining small number of infections (74, 1.7%) were caused by T. brucei, which displayed a patchier pattern of detection (12/34 study zones).
Database completeness
Entomological and epidemiological data without a geographical reference, and in particular without geographical coordinates, have not been included in the database yet, but kept separately for further checking and later incorporation. Therefore, data completeness was assessed only for fully georeferenced records. For both the tsetse and AAT databases, 100% completeness was achieved for administrative units (region, zone, district and peasant associations), while 46.39% and 9.2% village names were complete, respectively. For the tsetse database, tsetse species, trap ID, trap type, attractants used, survey period and trapping duration were 100% complete; high levels of completeness were also achieved for vegetation (91.21%), altitude (96.41%), tsetse intervention activity (71.64%) and data source files (95.31%). For the AAT database, the sampling method, data source file, date of survey, animal species and breed were 100% complete, whereas other elements like altitude, color, sex, age, body condition and PCV were 92.5%, 30.23%, 99.99%, 97.74%, 98.57% and 99.88% complete, respectively.
Discussion
This first edition of the national atlas of Ethiopia provides a large scale and up-to-date reference on the distribution of tsetse flies and AAT across the tsetse-infested areas of the country. However, despite the vast amount of data collected and the large geographical coverage, the atlas is still affected by a number of gaps and limitations. We discuss the main ones in the following section. We then look at the findings in relation to the existing literature, with a focus on publications on tsetse and AAT occurrence in specific locations of Ethiopia [12, 16].
Present limitations and gaps of the atlas
The lack of data on the occurrence of AAT in tsetse-free areas is arguably the main limitation of this first edition of the atlas. Tsetse-free areas in Ethiopia include the three northernmost and easternmost regions (i.e. Tigray, Afar, Somali, Harari regions and Dire Dawa city administration) as well as central, eastern and southern Oromia and large parts of Amhara. As documented in several countries, AAT and especially T. vivax can occur at large distances from the tsetse belt [21, 22]. Trypanosoma vivax is known to be widespread in Ethiopia [13, 59], and while the burden of AAT in tsetse-free areas is bound to be lower than within the tsetse belt, its impact should not be underestimated.
Another major limitation of the atlas is that, to date, it does not incorporate data collected by universities and other academic or educational institutions. In the framework of the continental atlas of tsetse and AAT, FAO is in the process of mapping data from scientific publications for the whole of Africa [12, 16, 22], and Fig. 6 shows these data overlaid onto the national atlas in Ethiopia. All data extracted by FAO from publications are available for national authorities in Ethiopia to complement their national atlas; however, the raw data behind the papers are not available at FAO level. In particular, the information that can be extracted from the papers lacks the trap- and animal-level data upon which hinges the national atlas in Ethiopia. A proper inclusion of the published information into the national atlas will require the engagement of academic institutions, with a view towards including the raw data that underpin scientific publications.

The time coverage of 10 years is arguably another limitation of this first edition of the atlas. In fact, other existing national atlases cover periods of 15–20 years or more [21, 46,47,48,49]. A time frame longer than 10 years would allow temporal trends to be better captured [48], and it would also help fill some of the existing geographical gaps.
Looking at the AAT component of the atlas, one weakness is its total reliance on BCT as a diagnostic method. BCT is an effective technique when AAT diagnosis is geared towards control, as it is the case in the surveillance activities upon which the national atlas is based. In fact, BCT-positive animals are normally those showing more overt clinical symptoms and reduced PCV, and as such they represent good targets for treatment [60]. However, the use of more sensitive diagnostic tools such as serological or molecular methods would enhance knowledge of exposure to and diversity of pathogenic trypanosomes [61].
Another limitation of the national AAT database is the very limited data on host species other than cattle (< 0.5%). This is not a minor gap in a country with very high livestock numbers and diversity and where trypanosomosis is well known to affect also small ruminants [62], donkeys [63] and other livestock species [17].
Finally, the national atlas for Ethiopia lacks data on the occurrence of T. evansi, an important pathogenic, non-tsetse-transmitted trypanosome. Trypanosoma evansi causes the disease called ‘surra’ [64], which has a broad global distribution [65]. In Africa, surra mainly affects camels, and in Ethiopia the disease is present in the eastern, northern and southern parts of the country [17, 19, 66]. The inclusion of T. evansi infections in the national atlas would give a more complete picture of vector-borne animal trypanosomoses in the country.
Occurrence of tsetse species
In this section we discuss our findings on the geographic distribution of tsetse species in Ethiopia in relation to the available literature.
Glossina pallidipes is the dominant tsetse species in many countries in eastern Africa [47, 67]. In western Ethiopia, our study found G. pallidipes to have the highest densities of all species and a broad distribution in two areas (i.e. SNNPR and western Oromia). The species was also captured in one area in Gambela, despite the relatively low sampling effort in this region, thus corroborating previous reports [68]. The absence of detection of G. pallidipes in Benishangul-Gumuz and Amhara is in line with historical and current knowledge [10, 12].
Regarding G. morsitans submorsitans, its continental distribution stretches from Senegal to Ethiopia. In many countries its populations have been shrinking because of land cover changes and the reduction of wildlife hosts [46, 69], and the species is now mainly confined to protected areas [22]. By contrast, our findings show that G. morsitans submorsitans maintains a fairly broad distribution in western Ethiopia, having been detected in all but one study regions (i.e. SNNPR). Furthermore, even in the SNNPR the species could still be present, having been reported from the Omo valley as recently as in 2015 [12, 70].
The populations of G. fuscipes fuscipes in Ethiopia are believed to be separated from the main belt in central Africa [10]. However, the species is reported across the border with Sudan [21], while recent information from South Sudan is very limited [71]. In our study the highest densities of G. fuscipes fuscipes were detected in Gambela, followed by Oromia and SNNPR. The absence of detection of this species in the Benishangul-Gumuz and Amhara regions, as well as in the Rift Valley basin, is consistent with current knowledge [12].
Even more markedly than for G. fuscipes fuscipes, the populations of G. tachinoides in Ethiopia are separated from the species’ main continental belt, which stretches from Guinea to the Central African Republic [10]. In our study, G. tachinoides was found to be the main species at the northern limit of the tsetse distribution in Ethiopia, and in particular in the Abay River basin in Amhara and Benishangul-Gumuz regions. The species is also widely distributed in Oromia and Gambela, while the absence of detection of G. tachinoides in SNNPR is consistent with current knowledge [12].
Glossina longipennis is known to be distributed in fairly fragmented populations in eastern Africa [10, 15], including recent reports from Kenya [12, 47] and Tanzania [67]. The species was also known to be present in Ethiopia [13], although it did not emerge in recent literature reviews [12]. In our study, the finding of one G. longipennis in the Gibe/Omo basin is consistent with past records [13], and it indicates that the species is likely to be persisting in Ethiopia at very low densities.
Finally, G. brevipalpis was known to be patchily distributed from South Africa to Ethiopia [10, 15], and recent reports confirm its presence from South Africa to Kenya [47, 67, 72, 73]. However, our study, consistently with recent literature reviews [12], did not find evidence of its persistence in Ethiopia.
Occurrence of African animal trypanosomosis
The national atlas shows that AAT is widespread in the tsetse-infested regions of Ethiopia. The overall prevalence of 4.8% is in line with estimates from meta-analysis of published data [74], with the advantage of being based on a much larger and up-to-date dataset.
When compared with large-scale investigations of similar size, scope and diagnostic method, the observed prevalence for bovine trypanosomosis in western Ethiopia is shown to be higher than in tsetse-infested areas in Kenya (1.8% [47]), but lower than in Burkina Faso (6% [49]) and Mali (7% [46]). It is also lower than Nigeria, where a literature review yielded an 8.6% prevalence [22]. Regarding the relative proportion of T. vivax and T. congolense, the ratio between these two species that we found in Ethiopia (i.e. 0.58) was similar to that in Mali (i.e. 0.76) and Burkina Faso (i.e. 0.9), where T. congolense was also predominant. By contrast, T. vivax was relatively more prevalent in Kenya (ratio 1.14). As to T. brucei, its very low prevalence in Ethiopia is similar to the very low levels found in other countries [22, 46, 47, 49].
When interpreting the enzootic situation of AAT in western Ethiopia, it is important to note that it is influenced by past and ongoing control activities. A detailed discussion of past and present interventions carried out over the years is beyond the scope of this paper. However, notably, more than two-thirds of the tsetse-infested areas in Ethiopia are presently estimated to be at some level of control. Insecticide-treated cattle (ITC) is the most widely used control method [75], because of its cost-effectiveness in areas where a sufficient number of cattle are present [76]. Other tsetse-control methods deployed in Ethiopia include insecticide-treated targets [75], ground spraying, the sterile insect technique [42] and the sequential aerosol technique [77, 78]. Regarding the direct control of AAT through trypanocidal drugs, diminazene aceturate and isometamidium chloride are the most widely used compounds [34]. NICETT is active in the procurement, distribution and administration of trypanocides, but other stakeholders such as regional authorities and NGOs are also involved. Worryingly, aberrant use and poor quality of trypanocidal drugs are widespread in Ethiopia, and they pose a serious risk for drug resistance [34, 79, 80].
Conclusions
AHI will continue to enhance and regularly update the national atlas of tsetse and AAT, and it will also make efforts to gather and include data collected by universities and other learning and research institutions. Future work should also tackle the issue of disease control data. Indeed, a large amount of information is available at AHI on the application of different tsetse and AAT control tools. However, as was the case for epizootic and entomological data before the atlas was developed, a system to centralize and map control data is lacking. When the latter is developed, it will be possible to combine epizootic, entomological and control data and thereby assess and monitor the progress in the control of AAT at the national level through the application of progressive control pathway (PCP) approach [41]. A number of these activities are being implemented in the framework of the recently launched project ‘COntrolling and progressively Minimizing the Burden of Animal Trypanosomosis’ (COMBAT) [81].
Availability of data and materials
Relevant data are within the manuscript and its Supporting Information files. Point-level data, which are not within the manuscript and its Supporting Information files, are the property of the Government of Ethiopia. Requests for data can be addressed to: Tesfaye Rufael Chibssa, Director General, Animal Health Institute, PO Box: 04 Sebeta, Ethiopia, Phone: + 251 11 338 0898, Email: director@ahi.gov.et; info@ahi.gov.et. Requests will be evaluated on the basis of the national data policy.
Abbreviations
- AAT:
-
African animal trypanosomosis
- AHI:
-
Animal Health Institute
- BCT:
-
Buffy-coat technique
- FAO:
-
Food and Agriculture Organization of the United Nations
- GPS:
-
Global Positioning System
- HAT:
-
Human African trypanosomosis
- HQ:
-
Head Quarters
- ITC:
-
Insecticide-treated cattle
- ITT:
-
Insecticide-treated targets
- NAHDIC:
-
National Animal Health Diagnostic and Investigation Centre
- NICETT:
-
National Institute for Control and Eradication of Tsetse and Trypanosomosis
- PCP:
-
Progressive control pathway
- PCV:
-
Packed cell volume
- PAAT:
-
Programme against African Trypanosomosis
- SNNPR:
-
Southern Nations, Nationalities and People's Region
- WHO:
-
World Health Organization
References
CSA: Agricultural Sample Survey 2020/21 [2013 E.C.] - Volume II - Report on Livestock and Livestock Characteristics. Addis Ababa, Ethiopia: Central Statistical Agency; 2021.
MoA. Guideline on import and export of animals and animal genetic resources. Addis Ababa: Ministry of Agriculture; 2012.
MoA. Animal Health Year Book (2009/10). Addis Ababa: Ministry of Agriculture; 2010. p. 63.
Shaw A, Wint GRW, Cecchi G, Mattioli RC, Robinson T. Intervening against bovine trypanosomosis in eastern Africa: mapping the costs and benefits, vol. 11. Rome: Food and Agriculture Organization of the United Nations; 2017.
Swallow BM. Impacts of trypanosomiasis on African agriculture. Vol. 2. Rome, Italy: Food and Agriculture Organization of the United Nations (FAO); 2000. https://www.fao.org/3/X4755EN/x4755en.pdf.
Taylor K, Authié EML. Pathogenesis of animal trypanosomiasis. In: Maudlin I, Holmes PH, Miles MA, editors. The trypanosomiases. Wallingford: CABI Publishing; 2004. p. 331–53.
Büscher P, Cecchi G, Jamonneau V, Priotto G. Human African trypanosomiasis. Lancet. 2017;390:2397–409.
Desquesnes M, Dia ML. Mechanical transmission of Trypanosoma vivax in cattle by the African tabanid Atylotus fuscipes. Vet Parasitol. 2004;119:9–19.
Gonzatti MI, González-Baradat B, Aso PM, Reyna-Bello A. Trypanosoma (Duttonella) vivax and typanosomosis in Latin America: secadera/huequera/cacho hueco. Trypanosomes and Trypanosomiasis. Cham: Springer; 2014. p. 261–85.
Ford J, Katondo KM. The distribution of tsetse flies in Africa (3 maps). Nairobi, Kenya: Organization of African Unity/Cook, Hammond & Kell; 1977.
Langridge WP. A tsetse and trypanosomiasis survey of Ethiopia. London: Ministry of Overseas Development; 1976.
Cecchi G, Paone M, Argiles Herrero R, Vreysen MJ, Mattioli RC. Developing a continental atlas of the distribution and trypanosomal infection of tsetse flies (Glossina species). Parasit Vectors. 2015;8:284.
Abebe G. Trypanosomosis in Ethiopia. Ethiop J Biol Sci. 2005;4:75–121.
Slingenbergh J. Tsetse control and agricultural development in Ethiopia. World Animal Rev. 1992;70:30–6.
Moloo SK. The distribution of Glossina species in Africa and their natural hosts. Insect Sci Appl. 1993;14:511–28.
Cecchi G, Paone M, Feldmann U, Vreysen MJB, Diall O, Mattioli RC. Assembling a geospatial database of tsetse-transmitted animal trypanosomosis for Africa. Parasit Vectors. 2014;7:39.
Birhanu H, Fikru R, Said M, Kidane W, Gebrehiwot T, Hagos A, et al. Epidemiology of Trypanosoma evansi and Trypanosoma vivax in domestic animals from selected districts of Tigray and Afar regions, Northern Ethiopia. Parasites Vectors. 2015;8:1–11.
Hadush B, Afera B, Aregawi G, Awol N, Mulu A, Woldu K. Preliminary study on mechanically transmitted bovine trypanosomosis and management of trypanocidal drugs in selected peasant associations of Tigray. 2015.
Fikru R, Andualem Y, Getachew T, Menten J, Hasker E, Merga B, et al. Trypanosome infection in dromedary camels in Eastern Ethiopia: prevalence, relative performance of diagnostic tools and host related risk factors. Vet Parasitol. 2015;211:175–81.
Cherenet T, Sani RA, Speybroeck N, Panandam JM, Nadzr S, Van den Bossche P. A comparative longitudinal study of bovine trypanosomiasis in tsetse-free and tsetse-infested zones of the Amhara Region, northwest Ethiopia. Vet Parasitol. 2006;140:251–8.
Ahmed SK, Rahman AH, Hassan MA, Salih SEM, Paone M, Cecchi G. An atlas of tsetse and bovine trypanosomosis in Sudan. Parasit Vectors. 2016;9:194.
de Gier J, Cecchi G, Paone M, Dede P, Zhao W. The continental atlas of tsetse and African animal trypanosomosis in Nigeria. Acta Trop. 2020;204:105328.
Berhe T, Bayleyegn G, Assefa F, Girma S. Prevalence of bovine trypanosomosis and its vector density in Tembro District, South Eastern Ethiopia. J Animal Sci Adv. 2015;5:1333–43.
Gechere G, Terefe G, Belihu K. Impact of tsetse and trypanosomiasis control on cattle herd composition and calf growth and mortality at Arbaminch District (Southern Rift Valley, Ethiopia). Trop Anim Health Prod. 2012;44:1745–50.
Stein J, Ayalew W, Rege E, Mulatu W, Lemecha H, Tadesse Y, et al. Trypanosomosis and phenotypic features of four indigenous cattle breeds in an Ethiopian field study. Vet Parasitol. 2011;178:40–7.
Mekuria S, Ashenafi H, Kebede N, Eyasu T, Terefe G. The impact of Gibe III hydroelectric dam on the situation of livestock diseases with particular emphasis on bovine trypanosomosis in Southern Ethiopia. Int J Vet Sci Res. 2020;6:104–13.
Kassaye B, Tsegaye D. Prevalence of bovine trypanosomosis, Tsets e Density and farmers perceptions on the Impact of Control Program in Kelle m Wollega Zone, Western Oromia, Ethiopia. J Veterinar Sci Technol. 2016;7:295.
Denu TA, Asfaw Y, HailuTolossa Y. Bovine trypanosomosis in three districts of Southwest Oromia. Ethiopia Ethiopian Veterinary Journal. 2012;16:23–39.
Dagnachew S, Sangwan AK, Abébé G. Epidemiology of bovine trypanosomosis in the Abay (Blue Nile) basin areas of Northwest Ethiopia. Rev Elev Med Vet Pays Trop. 2005;58:151.
Tesfaye D, Speybroeck N, De Deken R, Thys E. Economic burden of bovine trypanosomosis in three villages of Metekel zone, Northwest Ethiopia. Trop Anim Health Prod. 2012;44:873–9.
Wyohannes D, Kebede E, Abebe G. Study on the assessment of drug resistance on Trypanosoma vivax in Tselemti woreda, Tigray, Ethiopia. Ethiop Vet J. 2010;14:15–30.
Sinshaw A, Abebe G, Desquesnes M, Yoni W. Biting flies and Trypanosoma vivax infection in three highland districts bordering lake Tana, Ethiopia. Vet Parasitol. 2006;142:35–46.
Efrem DB, Yacob HT, Hagos AT, Basu AK. Bovine trypanosomosis in Gimbi district of Western Oromia, Ethiopia. Animal Biol. 2010;60:123–31.
Tekle T, Terefe G, Cherenet T, Ashenafi H, Akoda K, Teko-Agbo A, et al. Aberrant use and poor quality of trypanocides: a risk for drug resistance in south western Ethiopia. BMC Vet Res. 2018;14:4.
Moti Y, De Deken R, Thys E, Van Den Abbeele J, Duchateau L, Delespaux V. PCR and microsatellite analysis of diminazene aceturate resistance of bovine trypanosomes correlated to knowledge, attitude and practice of livestock keepers in South-Western Ethiopia. Acta Trop. 2015;146:45–52.
WHO: Control and surveillance of African trypanosomiasis. In: Technical Report Series. Geneva: World Health Organization; 1998.
Hide G. History of sleeping sickness in East Africa. Clin Microbiol Rev. 1999;12:112–25.
WHO: Control and surveillance of human African trypanosomiasis. In: Technical Report Series. Geneva: World Health Organization; 2013.
Franco JR, Cecchi G, Paone M, Diarra A, Grout L, Kadima Ebeja A, et al. The elimination of human African trypanosomiasis: achievements in relation to WHO road map targets for 2020. PLoS Negl Trop Dis. 2022;16:e0010047.
Simarro PP, Cecchi G, Franco JR, Paone M, Fèvre EM, Diarra A, et al. Estimating and mapping the population at risk of sleeping sickness. PLoS Negl Trop Dis. 2012;6:e1859.
Diall O, Cecchi G, Wanda G, Argiles-Herrero R, Vreysen MJB, Cattoli G, et al. Developing a progressive control pathway for African animal trypanosomosis. Trends Parasitol. 2017;33:499–509.
Alemu T, Kapitano B, Mekonnen S, Aboset G, Kiflom M, Bancha B, et al. Area-wide control of tsetse and trypanosomosis: Ethiopian experience in the Southern Rift Valley. Area-wide control of insect pests. Cham: Springer; 2007. p. 325–35.
FAO: Improving food security in sub-Saharan Africa by supporting the progressive reduction of tsetse-transmitted trypanosomosis in the framework of NEPAD (Phase 2) - Project findings and recommendations. Rome: Food and Agriculture Organization of the United Nations; 2019, 43. https://www.fao.org/3/CA3271EN/ca3271en.pdf.
FAO: Developing capacities for the control of tsetse and trypanosomosis in Ethiopia – Project Profile. Food and Agriculture Organization of the United Nations; 2022. https://www.fao.org/3/cb9186en/cb9186en.pdf.
Hursey BS. The programme against African trypanosomiasis: aims, objectives and achievements. Trends Parasitol. 2001;17:2–3.
Diarra B, Diarra M, Diall O, Bass B, Sanogo Y, Coulibaly E, et al. A national atlas of tsetse and African animal trypanosomosis in Mali. Parasit Vectors. 2019;12:466.
Ngari NN, Gamba DO, Olet PA, Zhao W, Paone M, Cecchi G. Developing a national atlas to support the progressive control of tsetse-transmitted animal trypanosomosis in Kenya. Parasit Vectors. 2020;13:1–12.
Shereni W, Neves L, Argilés R, Nyakupinda L, Cecchi G. An atlas of tsetse and animal African trypanosomiasis in Zimbabwe. Parasit Vectors. 2021;14:50.
Percoma L, Rayaissé JB, Gimonneau G, Bengaly Z, Pooda SH, Pagabeleguem S, et al. An atlas to support the progressive control of tsetse-transmitted animal trypanosomosis in Burkina Faso. Parasit Vectors. 2022;15:1–13.
Murray M, Murray PK, McIntyre WIM. An improved parasitological technique for the diagnosis of African trypanosomiasis. Trans R Soc Trop Med Hyg. 1977;71:325–6.
Shaw A, Cecchi G, Wint GRW, Mattioli RC, Robinson T. Mapping the economic benefits to livestock keepers of intervening against bovine trypanosomosis in Eastern Africa. Prev Vet Med. 2014;113:197–210.
Brightwell R, Dransfield RD, Kyorku C, Golder TK, Tarimo SA, Mungai D. A new trap for Glossina pallidipes. Tropical Pest Management. 1987;33:151–9.
Gouteux J, Lancien J. Le piège pyramidal à tsetse (Diptera: Glossinidae) pour la capture et la lutte. Essais comparatifs et déscription de nouveaux systèmes de capture [The pyramidal tsetse (Diptera: Glossinidae) trap for capture and control. Comparative tests and description of new capture systems]. Trop Med Parasitol. 1986;37:61–6.
Gouteux J-P, Le Gall F. Piège bipyramidal à tsé-tsé pour la protection de l’élevage en République centrafricaine. World Anim Rev. 1992;37:43.
Lancien J. Description du piège monoconique utilisé pour l’élimination des glossines en République Populaire du Congo. Cah ORSTOM, sér Ent Méd Parasitol. 1981;19:235–8.
Challier A, Laveissiere C. Un nouveau piège pour la capture des glossines (Glossina: Diptera: Muscidae). Description et essais sur le terrain [A new trap for capturing tsetse (Glossina: Diptera: Muscidae). Description and field trials]. Cahiers ORSTOM: Série Entomologie Médicale et Parasitologie. 1973;11:251–62.
Vreysen MJB, Khamis IS, Vloedt AMVV. Evaluation of sticky panels to monitor populations of Glossina austeni (Diptera: Glossinidae) on Unguja island of Zanzibar. Bull Entomol Res. 1996;86:289–96.
Leak SGA, Ejigu D, Vreysen MJB. Collection of entomological baseline data for tsetse area-wide integrated pest management programmes. Vol. 1. Rome, Italy: Food and Agriculture Organization of the United Nations; 2008. https://www.fao.org/3/i0535e/i0535e.pdf.
Fikru R, Goddeeris BM, Delespaux V, Moti Y, Tadesse A, Bekana M, et al. Widespread occurrence of Trypanosoma vivax in bovines of tsetse- as well as non-tsetse-infested regions of Ethiopia: a reason for concern? Vet Parasitol. 2012;190:355–61.
Moti Y, Fikru R, Büscher P, Van Den Abbeele J, Duchateau L, Delespaux V. Detection of African animal trypanosomes: the haematocrit centrifugation technique compared to PCR with samples stored on filter paper or in DNA protecting buffer. Vet Parasitol. 2014;203:253–8.
Oie. Chapter 2.4.17. Trypanosomosis (Tsetse-transmitted). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. vol. 1: World Organisation for animal Health (OIE); 2013.
Dinka H, Abebe G. Small ruminants trypanosomosis in the southwest of Ethiopia. Small Rumin Res. 2005;57:239–43.
Abebe R, Wolde A. A cross-sectional study of trypanosomosis and its vectors in donkeys and mules in Northwest Ethiopia. Parasitol Res. 2010;106:911–6.
Desquesnes M, Holzmuller P, Lai D-H, Dargantes A, Lun Z-R, Jittaplapong S. Trypanosoma evansi and surra: a review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. BioMed Res Int. 2013;2013:194176.
Aregawi WG, Agga GE, Abdi RD, Büscher P. Systematic review and meta-analysis on the global distribution, host range, and prevalence of Trypanosoma evansi. Parasit Vectors. 2019;12:67.
Olani A, Habtamu Y, Wegayehu T, Anberber M. Prevalence of camel trypanosomosis (surra) and associated risk factors in Borena zone, southern Ethiopia. Parasitol Res. 2016;115:1141–7.
Daffa J, Byamungu M, Nsengwa G, Mwambembe E, Mleche W. Tsetse distribution in Tanzania: 2012 status. Tanzania Vet J. 2013;28:1–11.
Kedir M, Lelisa K, Damena D. Bovine trypanosomosis and tsetse fly vectors in Abobo and Gambela districts, Southwestern Ethiopia. J Vet Sci Technol. 2016;7:2.
Adam Y, Marcotty T, Cecchi G, Mahama C, Solano P, Bengaly Z, et al. Bovine trypanosomosis in the Upper West Region of Ghana: entomological, parasitological, and serological cross-sectional surveys. Res Vet Sci. 2012;92:462–8.
Terefe E, Haile A, Mulatu W, Dessie T, Mwai O. Phenotypic characteristics and trypanosome prevalence of Mursi cattle breed in the Bodi and Mursi districts of South Omo Zone, southwest Ethiopia. Trop Anim Health Prod. 2015;47:485–93.
Lukaw YS, Mohammed Y, Abdelrahman M, Jaja L, Ochi E. Preliminary study on population structure, infection rate and habitats of Glossina fuscipes fuscipes (Diptera: Glossinidae) in Western Equatoria State South Sudan. IJISET-Int J Innov Sci Eng Technol. 2016;3:226–35.
De Beer CJ, Venter GJ, Green KK, Esterhuizen J, De Klerk DG, Ntshangase J, et al. An update of the tsetse fly (Diptera: Glossinidae) distribution and African animal trypanosomosis prevalence in north-eastern KwaZulu-Natal, South Africa. Onderstepoort J Vet Res. 2016;83:1–10.
Mulandane FC, Snyman LP, Brito DR, Bouyer J, Fafetine J, Van Den Abbeele J, et al. Evaluation of the relative roles of the Tabanidae and Glossinidae in the transmission of trypanosomosis in drug resistance hotspots in Mozambique. Parasit Vectors. 2020;13:1–16.
Leta S, Alemayehu G, Seyoum Z, Bezie M. Prevalence of bovine trypanosomosis in Ethiopia: a meta-analysis. Parasit Vectors. 2016;9:1–9.
Bekele J, Asmare K, Abebe G, Ayelet G, Gelaye E. Evaluation of Deltamethrin applications in the control of tsetse and trypanosomosis in the southern rift valley areas of Ethiopia. Vet Parasitol. 2010;168:177–84.
Shaw A, Wint GRW, Cecchi G, Torr SJ, Mattioli RC, Robinson TP. Mapping the benefit-cost ratios of interventions against bovine trypanosomosis in Eastern Africa. Prev Vet Med. 2015;122:406–16.
Adam Y, Cecchi G, Kgori PM, Marcotty T, Mahama CI, Abavana M, et al. The sequential aerosol technique: a major component in an integrated strategy of intervention against riverine tsetse in Ghana. PLoS Negl Trop Dis. 2013;7:e2135.
Kgori PM, Modo S, Torr SJ. The use of aerial spraying to eliminate tsetse from the Okavango Delta of Botswana. Acta Trop. 2006;99:184–99.
Degneh E, Ashenafi H, Kassa T, Kebede N, Shibeshi W, Asres K, et al. Trypanocidal drug resistance: a threat to animal health and production in Gidami district of Kellem Wollega Zone, Oromia Regional State. West Ethiop Prev Vet Med. 2019;168:103–7.
Dagnachew S, Tsegaye B, Awukew A, Tilahun M, Ashenafi H, Rowan T, et al. Prevalence of bovine trypanosomosis and assessment of trypanocidal drug resistance in tsetse infested and non-tsetse infested areas of Northwest Ethiopia. Parasite Epidemiol Control. 2017;2:40–9.
Boulangé A, Lejon V, Berthier D, Thévenon S, Gimonneau G, Desquesnes M, et al. The COMBAT project: controlling and progressively minimizing the burden of vector-borne animal trypanosomosis in Africa [version 2; peer review: 3 approved]. Open Res Eur. 2022;2:67.
Acknowledgements
The national atlas of tsetse and AAT in Ethiopia is an initiative of the Government of Ethiopia, and it was developed through the National Institute for Control and Eradication of Tsetse and Trypanosomosis (NICETT). We gratefully acknowledge all NICETT staff that were involved in the collection, recording and transmission of the data that were used for the development of the atlas. FAO provided technical assistance to the initiative in the framework of the Programme against African Trypanosomosis (PAAT) (www.fao.org/paat). In particular, technical assistance was provided within the two-phase FAO project ‘Improving food security in sub-Saharan Africa by supporting the progressive reduction of tsetse-transmitted trypanosomosis in the framework of the NEPAD’ (codes GTFS/RAF/474/ITA and GCP/RAF/502/ITA) and the FAO Technical Cooperation Project ‘Developing national implementation capacities for the control of tsetse and animal African trypanosomosis in Ethiopia’ (code TCP/ETH/3702).
Disclaimers
The boundaries and names shown and the designations used on the maps presented in this paper do not imply the expression of any opinion whatsoever on the part of FAO concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement. The views expressed in this paper are those of the authors and do not necessarily reflect the views of FAO.
Funding
Human and financial resources to develop the national atlas of tsetse and AAT in Ethiopia were provided by the Government of Ethiopia, through NICETT. FAO also provided funding, in particular thanks to the financial support of the Government of Italy (project ‘Improving food security in sub-Saharan Africa by supporting the progressive reduction of tsetse-transmitted trypanosomosis in the framework of the NEPAD’, codes GTFS/RAF/474/ITA and GCP/RAF/502/ITA) and the FAO Technical Cooperation Project ‘Developing national implementation capacities for the control of Tsetse and African animal trypanosomosis in Ethiopia, code TCP/ETH/3702).
Author information
Authors and Affiliations
Contributions
Conceptualization: GC, BK, TG. Data curation: BK, TG, MP, GC, DA, AB, ZW, TK, AS, ED, AK, NA. Formal analysis: GC, BK, MP, TG. Funding acquisition: DB, GC, WZ. Investigation: DA, AB, ZW, TK, AS, ED, AK, NA. Methodology: GC, MP. Project administration: DB, BK, WZ. Supervision: DB, WZ. Validation: BK, TG, MP, GC. Visualization: GC. Writing—original draft: GC, BK, TG. Writing—review and editing: TG, BK, DB, DA, AB, ZW, TK, AS, ED, AK, NA, WZ, MP, GC. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: S1.
Apparent density of tsetse flies in Ethiopia by zone. Data collection period: 2010–2019.
Additional file 2: S2.
Apparent density of tsetse flies in Ethiopia by district (woreda). Data collection period: 2010–2019.
Additional file 3: S3.
Presence and absence (surveyed but not detected) of tsetse fly species in Ethiopia. Data collection period: 2010–2019.
Additional file 4: S4.
Prevalence of bovine trypanosomosis in Ethiopia by zone. Data collection period: 2010–2019.
Additional file 5: S5.
Prevalence of bovine trypanosomosis in Ethiopia by district (woreda). Data collection period: 2010–2019.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gebre, T., Kapitano, B., Beyene, D. et al. The national atlas of tsetse flies and African animal trypanosomosis in Ethiopia. Parasites Vectors 15, 491 (2022). https://doi.org/10.1186/s13071-022-05617-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05617-9