
Abstract
Background
The canine heartworm Dirofilaria immitis, a filarioid nematode of dogs and other carnivores, is widespread in the USA and the world. Over 20 different mosquito species serve as intermediate hosts of D. immitis, but their contribution to transmission varies according to factors like host feeding patterns, geographic locations and climatic conditions. The yellow fever mosquito, Aedes aegypti, is a competent vector of D. immitis but is often dismissed as a vector of veterinary relevance given its anthropophilic feeding behavior. We evaluated the prevalence of D. immitis in pet dogs along the USA-Mexico border and assessed whether Ae. aegypti in the area are naturally infected with heartworm and are potentially acting as a vector.
Methods
A total of 200 whole blood samples collected from pet dogs in the Lower Rio Grande Valley in south Texas from 2016 to 2019 were included in this study. Canine serum samples for D. immitis were tested using the DiroCHEK® Canine Heartworm Antigen Test Kit pre- and post-immune complex dissociations (ICD) and blood samples were tested using high-resolution melt (HRM) quantitative PCR (qPCR) and a probe-based qPCR. Additionally, mosquito specimens were collected and identified, and Ae. aegypti heads, abdomens and pools were tested using conventional PCR (cPCR) and HRM qPCR.
Results
Overall, heartworm prevalence in dogs aged > 6 months was 40.8% (64/157) when the results from all testing modalities were considered. Heartworm antigen was detected in 33.5% and 40.7% of the dogs using DiroCHEK® pre- and post-ICD, respectively. By molecular screening, 20.1% of dogs tested positive with probe-based qPCR, while only one tested positive with HRM qPCR. Of the Ae. aegypti abdomens from blood-fed Ae. aeygpti tested, 20 (21.7%) from mosquitoes that fed on dogs and four (7%) from those that fed on humans tested positive for heartworm. Among Ae. aegypti heads from blood-fed Ae. aeygpti, two (1.1%) were positive based on cPCR and four (2.5%) were positive based on HRM qPCR. No D. immitis DNA was detected in the 208 pools of whole bodies (358 individuals) of Ae. aegypti gravid females.
Conclusions
Our study highlights a high prevalence of heartworm in dogs in south Texas and provides evidence that Ae. aegypti could be contributing to heartworm transmission in canine populations in this region.
Graphical Abstract

Similar content being viewed by others


Background
Aedes aegypti is an important arbovirus vector relevant to public health on the global scale, but it has not been implicated to date as being important to veterinary health due to its anthropophilic nature [1]. However, with high levels of non-human feeding reported in south Texas [2], there is the potential for Ae. aegypti to transmit animal pathogens, such as Dirofilaria immitis, the agent of dog heartworm, or allow for zoonotic bridge transmission of agents from animals to humans.
There are 23 known mosquito species or species complexes that are competent vectors of D. immitis in the USA [3]. The main mosquito species implicated in D. immitis transmission are: (i) competent as biological vectors based on laboratory infection studies; (ii) mammalophilic and utilize dogs or other canines as hosts; and (iii) distributed in areas of high heartworm prevalence [3]. Both field and laboratory evidence incriminate Aedes vexans, A. trivittatus, A. punctipennis and Culex quinquefasciatus as important vectors [3, 4]. Some mosquito species are highly competent in laboratory settings but are considered less important in nature due to infrequent feeding on dogs. Aedes aegypti falls into this category, as it is used as a competent model of D. immitis transmission in the laboratory [5,6,7,7] but is considered to be a highly anthropophilic species in most contexts [1]. Studies in the state of Florida (USA) and in Argentina have documented field-collected Ae. aegypti infected with D. immitis [8, 9], with two of 22 pools testing positive in Argentina and 17% prevalence in Florida. However, wild populations of this mosquito are rarely subjected to molecular screening for the presence of D. immitis DNA. Our recent study documented Ae. aegypti feeding more frequently on dogs than humans in south Texas [2], demonstrating the potential for high contact with canine reservoirs of D. immitis in some contexts. In this area, Ae. aegypti meets the three criteria outlined above, implicating that it is potentially an important vector of D. immitis in south Texas, but no studies have been conducted on mosquito vectors of heartworm in the region.
Dirofilaria immitis is a filarioid nematode that needs both a mosquito vector and a vertebrate host to complete its life-cycle. Microfilariae enter the mosquito during feeding on an infected dog and reach the head as third-stage larvae (L3) where they can be transmitted to new individuals [3]. Upon being bitten by an infected mosquito, the L3, the infective stage of D. immitis, migrate and may develop into the adult stage in dogs and cats, which often develop clinical disease. Disease severity varies, with severe disease being characterized by exercise intolerance, coughing, hemoptysis, tachypnea, dyspnea and syncope, heart failure or death [10]. Humans can also be infected with D. immitis via a mosquito bite, but infection is usually asymptomatic [10]. Canine heartworm disease, caused by D. immitis, is one of the most ubiquitous parasitic diseases of dogs in the USA with more than 100,000 cases each year [11, 12]. The cost of treatment for dogs in the USA is estimated to be US$75 million annually [12]. Disease in companion animals can be prevented through chemoprophylaxis and is recommended for dogs, cats and ferrets [13]. In the USA, canine heartworm disease is most prevalent in the southern states where mosquito vectors are present year-round [14,15].
From 2013 to 2016, canine heartworm incidence increased 17.9% in the southeastern USA, which is more than in other regions of the USA [15]. Several studies have investigated wild canids and shelter dogs and D. immitis in Texas [16,17,18], although no studies, to our knowledge, have presented results for owned dogs in the Lower Rio Grande Valley (LRGV). Many dogs in low-income communities in the LRGV live a stray lifestyle, making them unique when compared to other owned dogs in the state, whose canine heartworm prevalence was reported at 5.5% in 2008 [19] and incidence as 3.19% in 2016 [15]. The objectives of this study were to evaluate the prevalence of canine heartworm in pet dogs in low- and middle-income communities along the USA-Mexico border and to assess whether Ae. aegypti in the area is infected with heartworms, including disseminated infections that may support their role as a biological vector.
Methods
Study area
Samples of dog blood and mosquitoes were collected in the LRGV in south Texas along the USA-Mexico border in low- and middle-income communities, labeled communities A-V, located in Hidalgo and Cameron counties (Fig. 1). Of these communities, four (I, J, V, K) were classified as middle-income (annual household income: US$30,000–40,000) and the remainder were classified as low-income (annual household income: US$15,000–29,999) [20]. Most of these study sites are unincorporated neighborhoods called colonias, which are predominantly Hispanic, low-income communities where one or more city infrastructure or service is missing (e.g. poor water sanitation, lack of drainage, lack of trash collection) [21]. This lack of infrastructure can lead to a more suitable habitat for Ae. aegypti larvae. A detailed description of these neighborhoods has been reported previously [2, 20]. This area has a subtropical climate with a cold and dry season from November to February and a rainy season from April to October that peaks in September [22]. The annual temperature and rainfall in the LRGV ranges from 17.4 °C to 28.7 °C and 609 mm of rain, respectively, and past mosquito surveillance in these study sites documents year-round occurrence of adult female Ae. aegypti [20].

Map of sampling sites in Hidalgo and Cameron counties, Texas, USA
Dog sampling and diagnostics
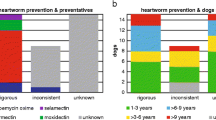
Samples of dog blood were collected via jugular or cephalic venipuncture from 340 pet dogs in eight colonias in the LRGV from 2016 to 2019 (Fig. 1) as previously described [23]. Of these 340 samples, 200 with adequate serum volume were selected for further testing after processing for a study focused on canine Chagas disease [23]. Of these 200 samples, extracted DNA or serum was depleted in 51 samples before all molecular testing was completed, leaving only 149 dogs that were tested using all molecular methods. Dog owners gave informed consent to sampling, and samples of 3–5 ml (from dogs > 2.3 kg) or 0.5–2 ml (from dogs < 2.3 kg) of blood were collected and divided into an EDTA tube and a serum collection tube. Age (< 6 months, 6–12 months, 1–5 years, > 5 years), breed group (herding, hound, nonsporting, other, sporting, terrier, toy, working) and sex of the dogs were recorded. All samples came from dogs aged at least 6 weeks. Rabies vaccination (RABVAC 1; Boehringer Ingelheim Vetmedica, Boehringer Ingelheim, Ingelheim, Germany) was administered to all dogs aged ≥ 3 months as incentive for participation and as a public health protective measure. Canine sampling and vaccination protocols were approved by the Institutional Animal Care and Use Committee at Texas A&M University (AUP# 20180460 and #2015-0289 CA). Blood was separated into serum, plasma and buffy coat by centrifuging at 5488 relative centrifugal field (RCF) for 8 min and DNA was extracted from the buffy coat using the E.Z.N.A. kit (Omega Bio-Tek, Norcross, GA, USA).
Serum samples were tested using the DiroCHEK® Heartworm Antigen Test Kit (Synbiotics Corporation, Zoetis, Kalamazoo, MI, USA) pre- and post-heat treatment, according to the manufacturer’s instructions. For immune complex dissociations (ICD) by heat treatment, 500 µl of serum was heated to 103 °C for 10 min in a dry heat block [24, 25], followed by centrifugation; the resulting supernatant was used as a template in the antigen test. Positive or no antigen detected (NAD) results for each sample were assessed by visual color change according to the manufacturer’s recommendation. In addition to a visual color change, optical density readings from a spectrophotometer (Epoch; BioTek Instruments Inc., Winooski, VT, USA) at 490 nm were obtained, as previously described [26].
A high-resolution melt (HRM) qPCR was used to test the extracted buffy coat DNA for D. immitis DNA. Previously identified primers for a 115-bp section of the mitochondrial 12S ribosomal RNA gene found in filarial worms capable of detecting multiple species [27] were used, in conjunction with a previous HRM qPCR protocol [28]. Each conventional PCR (cPCR) and HRM quantitative PCR (qPCR) plate contained ≥ 4 negative controls consisting of nuclease-free H2O and ≥ 1 positive controls of D. immitis DNA provided by the Texas A&M Parasitology Diagnostic Laboratory. We also used a simplex probe-based qPCR that targeted a 166-bp fragment of the cytochrome oxidase c subunit (cox1) gene of D. immitis based on a previously validated protocol [29] with the following modifications [16]: (i) oligonucleotides (Fil.COI.749-F, Fil.COI.914-R) and the FAM reporter probe (D.immi.COI.777-P) were synthesized by Invitrogen (Thermo Fisher Scientific, Waltham, MA, USA; (ii) qPCR reactions had a final volume of 20 µl containing 3.5 µl nuclease-free water, 0.5 µl of 20 µM probe, 0.5 µl of 50 µM forward and reverse primers, 10 µl of TaqMan® Multiplex Master Mix (Applied Biosystems, Thermo Fisher Scientific) and 5 µl of DNA template. The qPCR was performed on a QuantStudio 3 Real-Time PCR system (Applied Biosystems, Thermo Fisher Scientific ) using TaqMan® universal-cycling conditions, as follows: a hold stage at 95 °C for 20 s, and 40 cycles of a two-step PCR stage at 95 °C for 1 s followed by 60 °C for 20 s. All qPCR runs included a positive and negative control. The positive control consisted of DNA extracted from an adult D. immitis provided by the Texas A&M Parasitology Diagnostic Laboratory. The non-template negative control consisted of nuclease-free water.
Mosquito sampling and diagnostics
Mosquito sample collection took place each month between September 2016 and December 2018 using baited BG-Sentinel 2 traps (BGS2; Biogents AG, Regensburg, Germany) and CDC Autocidal Gravid Ovitraps (AGO) (SpringStar, Inc., Woodinville, WA, USA and CDC Dengue Branch, Puerto Rico) [2, 20]. Briefly, BGS2 were baited with BG Lures outside of homes once per week for 24 h. AGOs were placed both inside and outside of homes and were checked once per week. Not all trap types were placed in all communities. Trap placement was determined by the willingness of residents to participate in the study. Mosquitoes were then identified to the species level using a dichotomous key [30] and were separated by sex. Female blood-fed mosquitoes were placed individually in nuclease-free 1.5-ml micro-centrifuge tubes and unfed females, males and gravid females each were pooled in separate tubes. Samples were stored at − 20 °C or − 80 °C until further processing.
Species identifications based on morphology made under a dissecting microscope were confirmed with a molecular protocol based on a PCR using primers for the cox1 gene followed by Sanger sequencing [31]. Methods for DNA extraction and bloodmeal host identification have been previously described [2]. All female Ae. aegypti mosquitoes (n = 184) with identified bloodmeal hosts and with sufficient DNA were used. Briefly, mosquitoes were washed with 10% bleach to remove exogenous DNA, abdomens or heads were homogenized in 200 μl of lysis solution and DNA was extracted using the Kingfisher™ Flex Purification System and the MagMax Core Nucleic Acid Purification Kit (Thermo Fisher Scientific) per the manufacturer’s instructions.
We used two molecular screening methods to detect D. immitis DNA in mosquito samples: cPCR with Sanger sequencing and HRM qPCR. cPCR with Sanger sequencing was used to identify the presence filarial DNA as belonging to D. immitis, and HRM qPCR was used to detect small amounts of filarial DNA present in samples, with a positive read from one modality indicating a positive sample. Mosquito abdomens and heads were tested separately to distinguish D. immitis in the DNA of the bloodmeal (blood-engorged abdomen) versus disseminated L3 in the mouthpart tissues [3]. We used primers designed by Rishniw et al. [32] for the cox1 gene that are specific to D. immitis for both types of PCR. cPCR followed by Sanger sequencing was used on DNA extracted from blood-engorged abdomens and heads. Heads only and pools of whole-bodied gravid females were tested with HRM qPCR given that the species identification results had already been confirmed using Sanger sequencing when testing the abdomens.
Statistical analyses
Overall prevalence of D. immitis in dogs was calculated by categorizing as positive any dog that either tested positive by serum antigen and/or tested PCR-positive. Two prevalence values are reported, one for dogs of all ages and one for dogs of aged > 6 months. This distinction was made because the life-cycle of the parasite makes it difficult to detect infections in young dogs [25, 33] and because most epidemiology studies report prevalence only in dogs aged > 6 months. Dogs that tested positive by at least one modality were considered to be positive for analysis purposes. Unless otherwise mentioned, all values reported included dogs of all ages. We used a Z-test for two independent proportions to compare differences in prevalence between female and male dogs and between mosquitoes that fed on dogs to mosquitoes that fed on other vertebrates. For comparison of prevalence among breed group and age groups, we used the Chi-square goodness of fit tests for more than two categories.
Results
Heartworm prevalence in dogs
A total of 200 dog serum samples were tested using the DiroCHEK® Heartworm Antigen Test Kit. A high prevalence of D. immitis infection was found in sampled dogs of all ages: 33.5% (67/200) in pre-heat-treated samples and 40.7% (81/199) in post-heat-treated samples (Table 1). Only one sample tested positive pre-ICD and NAD post-ICD whereas 14 samples tested negative pre-ICD and positive post-ICD. Prevalence for heartworm in different communities ranged from 7.1% (1/14; community T) to 80% (4/5; community M). Using the HRM qPCR to assess 158 of the 200 dog blood samples (i.e. those samples that still had a sufficient quantity of DNA), we found only one (0.06%) dog positive for D. immitis DNA. Using the probe-based qPCR to assess the same 149 dog samples for D. immitis DNA, we detected 30 (20.1%) dogs that tested positive. Of these 30 dog samples with evidence of D. immitis DNA in the blood (buffy coat), 27 also tested positive for heartworm infection by DiroCHEK® post-ICD treatment. Combining all diagnostic modalities, 39.3% (70/178) of dogs of all ages were positive for heartworm, and 40.8% (64/157) of dogs aged > 6 months were positive.
In total, 178 dogs were tested using all modalities, of which 96 were male and 82 were female. There was no significant difference in prevalence of D. immitis between sexes, with 67.7% (64/96) of males testing positive and 52.4% (43/82) of females testing positive (χ2 = 3.71, df = 1, P = 0.054) when dogs of all ages were considered. The number of positive dogs varied by age group (χ2 = 8.34, df = 3, P = 0.040), with dogs aged 6–12 months showing the highest level of prevalence followed by dogs aged between 1 and 5 years. However, the number of positive dogs did not significantly vary by breed group (χ2 = 9.94 df = 7, P = 0.192) (Table 2). All analyses, with the exception of overall prevalence in dogs aged ≥ 6 months, included dogs of all ages (i.e. aged ≥ 6 weeks).
D. immitis DNA in Ae. aegypti
We used cPCR with Sanger sequencing to test abdomens of blood-fed Ae. aeygpti (n = 184) with the blood host identified for D. immitis presence; 25 (13.6%) of these abdomens from mosquitoes with identified vertebrate bloodmeal tested positive for D. immitis DNA. Dirofilaria immitis was more commonly found (χ2 = 9.07, df = 1, P = 0.003) in the abdomens of mosquitoes that had fed on dogs (21.7%; 20/92) than in those that fed on cats, sheep, human and other vertebrates (5.4%; 5/92) (Table 3). The heads of four mosquitoes tested positive for D. immitis DNA, including three heads from mosquitoes that fed on dogs and one head from a mosquito that fed on a cat. No D. immitis DNA was found in the 208 pools of gravid females (358 individuals). The obtained 25 sequences had 97–100% similarity to the reference D. immitis sequence (GenBank Accession numbers: OP681144, OP681145 ).
Discussion
Our findings suggest that there is a high prevalence of canine heartworm infection in dogs from south Texas along the USA-Mexico border, with an infection prevalence of 40.8% in dogs from diverse communities, most of which consist of low-income colonias. Texas has been indicated to be an area of high canine heartworm incidence [34]. In pet dogs that have regular access to veterinary care, 23,489 positive cases were reported in 2016, 28,320 cases in 2017, 31,252 cases in 2018 and 35,656 cases in 2019 [13]. These data confirm a previously established trend of increasing prevalence of canine heartworm infection in Texas, with an increase of 17.23% from 2013 to 2016 [15]. Shelter dogs in the same area (i.e. Edinburg, TX) were documented in a previous study to have a lower prevalence of D. immitis than our samples, with 19/33 (20.9%) dogs being positive [35]; however, the diagnostic test used was a commercial, point-of-care enzyme-linked immunosorbent assay (ELISA) kit and did not include ICD by heat treatment, which may explain the discordance. In general, shelter dogs tend to have higher canine heartworm prevalence than owned dogs [36, 37]. However, shelter dogs having a lower incidence of infection than owned dogs from middle- and low-income communities may be explained by the lower socioeconomic status (SES) of sampled colonias, as low SES has been linked with higher prevalence of D. immitis in the state of North Carolina (USA) [38], or by other variables, such as sampling year or location. Moreover, owned dogs in the sampled area of the present study generally are kept outside and have a stray lifestyle. Therefore, it is likely that many of these owned dogs were not receiving veterinary care or macrocyclic-lactone heartworm preventives as two-thirds of dogs in the USA are not receiving preventative care [15]. Even for dogs receiving preventive care, failures of heartworm preventative measures have been reported in endemic states, such as Texas [39]. These same communities also have many stray dogs which likely have even higher heartworm prevalence rates than the population of dogs sampled in this study.
The American Heartworm Society recommends both antigen testing and microfilariae testing of all canine samples [33], but microfilariae tests are not routine procedure. Consistent with our own previous results [16, 42], antigen and microfilaria testing by qPCR in the present study gave different estimates of prevalence. Antigen tests work well at diagnosing infection with higher numbers of female D. immitis worms but are less sensitive for male-only infection with ≤ 3 females [40]. Moreover, antigen tests are more effective at identifying female worms that are > 6 months of age, making infection difficult to diagnosis in dogs aged < 6 months as it takes 50–58 days post-infection for L3 to develop into adults [41]. qPCR is a reliable and specific diagnostic tool for D. immitis [42] and the use of multiple diagnostic tests has been shown to improve diagnostic performance [29, 43, 44]. Two earlier studies found that qPCR detected a higher prevalence rate than antigen testing alone [29, 45], similar to our results as three dogs in the present study were considered to be D. immitis microfilaria positive and NAD before and after ICD. However, to our knowledge, the sensitivity of probe-based qPCR and HRM qPCR for the detection of nematodes have not been compared, making it difficult to comment on the differences between the two types of qPCR used in the present study. The increased detection of antigen post-ICD was expected given the study population, which comprised dogs from an endemic area, many of which had no history of heartworm prevention. Also, antigen tests are recommended for dogs aged > 6–7 months [25, 33]. In the present study, six dogs aged < 6 months tested positive in the post-ICD treatment, which is not surprising since heat treatment allows for earlier detection of infection [46]. Accordingly, we present our results both including and excluding dogs aged < 6 months. We saw a trend of highest infection prevalence in dogs aged 6–12 months, with a decrease of prevalence of infection after 1 year of age. This decrease could be explained by the potential fatality of dogs with untreated infection, with older dogs likely to have been infected longer and therefore have more advanced disease.
We found that of the Ae. aegypti abdomens with bloodmeal identified from Ae. aegypti that fed on dogs, 21.7% contained D. immitis DNA, suggesting that Ae. aegypti in these communities are frequently exposed to microfilaremic dogs. Some mosquitoes that fed on other vertebrates also tested positive for D. immitis, which may be explained by infections in other hosts (e.g. cats) and/or by a previous feeding on dogs that was not the most recent bloodmeal (i.e. mixed bloodmeal from > 1 vertebrate species). We also found four Ae. aegypti heads positive for D. immitis DNA out of the 157 tested, a rare observation which has only been documented for wild populations in the state of Florida (USA) [8]. L3, the infective stage of D. immitis, move to the mosquito head and enter the salivary gland; saliva, hemolymph and L3 move to the skin then enter the host body through the mosquito bite wound [47]. Therefore, the detection of D. immitis DNA in mosquito heads represents the presence of L3, providing evidence of the suitability of the Ae. aegypti as a biological vector. The prior findings of Ae. aegypti being a competent vector of D. immitis based on laboratory studies [6, 7] combined with the results presented in this study suggest that Ae. aegypti likely contributes to canine heartworm transmission in south Texas. Indeed, potential transmission has been assumed of a mosquito species if a positive result was found in a competent species [38]. In their study, Palmer et al. noted that large worm burdens in Ae. aegypti can lead to destruction of the excretory system and therefore mortality, a phenomenon that may be occurring in Ae. aegypti in the LRGV as no gravid females tested positive for D. immitis DNA [48]. As the relative role of different mosquito species in the transmission of D. immitis to dogs is rarely measured, little is known on the role of Ae. aegypti contribution to transmission. In an earlier study, Paras et al. concluded that a mosquito can be considered to be a relevant vector based on its abundance and infection prevalence, reporting Ae. albopictus as an important vector based on a prevalence of approximately 15% calculated based on pool testing [49]. Still, a lower prevalence (e.g. 0.30% in Aedes albopictus [38], 0.7% in Culix erraticus [50], 2.3% in Ae. vexans [51]) has been reported for accepted heartworm vectors. Given that we found a prevalence of 13.6% in abdomens and 2.5% in heads, it reasonable to conclude that Ae. aegypti has a sufficiently high prevalence to be a vector of D. immitis.
While our previous study documented that these populations of Ae. aegypti fed 50% of the time on dogs, we also documented 31% feeding on humans and 12% feeding on cats [2]. This result suggests the potential for Ae. aegypti to also contribute as a bridge vector species, allowing spill-over transmission of D. immitis from dogs to humans. Human infections with D. immitis have been reported in North and South America, Europe, Asia and Australia, although most cases are asymptomatic [10]. When symptoms do occur, they include cough, chest pain, fever, hemoptysis and pleural effusion [10]. Infection is difficult to diagnose and may be underreported [52]. In the USA, fewer than 120 cases have been reported since 1941 [2], with the highest reported incidence in humans occurring in the southeastern region, with 23 reported cases as of 2005 [52]. At least one case of D. immitis infection in a human patient has been reported in Texas [53]. Infection in dogs is highest in the southeast USA [19] and is increasing in prevalence in this region [15]. It should be noted that this region overlaps with the establishment of Ae. aegypti in many regions of the southern USA [11, 19, 54]
Conclusions
In the present study, the prevalence of D. immitis infection in owned dogs in low- and middle-income communities in the LRGV, Texas, was high (40.8%). As prevalence estimates varied based on the diagnostic modality used, our results further support the use of integrated immunodiagnostic and molecular tests to determine heartworm prevalence. The finding of D. immitis DNA in the heads of Ae. aegypti provides evidence that this mosquito could be contributing to heartworm transmission in canine populations and potential spill-over to human populations. However, further study is needed to inform the relative role of Ae. aegypti and other mosquito species in transmitting D. immitis in the region.
Availability of data and materials
Data is available upon request.
Abbreviations
- cPCR:
-
Conventional PCR
- HRM:
-
High-resolution melt
- ICD:
-
Immune complex dissociation
- LRGV:
-
Lower Rio Grande Valley
- NAD:
-
No antigen detected
- qPCR:
-
Quantitative PCR
References
Takken W, Verhulst NO. Host preferences of blood-feeding mosquitoes. Ann Rev Entomol. 2013;58:433–53.
Olson MF, Ndeffo-Mbah ML, Juarez JG, Garcia-Luna S, Martin E, Borucki MK, et al. High rate of non-human feeding by Aedes aegypti reduces Zika virus transmission in South Texas. Viruses. 2020;12:453.
Ledesma N, Harrington L. Mosquito vectors of dog heartworm in the United States: vector status and factors influencing transmission efficiency. Top Companion Anim Med. 2011;26:178–85.
Tiawsirisup S, Nithiuthai S. Vector competence of Aedes aegypti (L.) and Culex quinquefasciatus (Say) for Dirofilaria immitis (Leidy). Southeast Asian J Trop Med Public Health. 2006;37:110–4.
Serrão ML, Labarthe N, Lourenço-de-Oliveira R. Vectorial competence of Aedes aegypti (Linnaeus 1762) Rio de Janeiro strain, to Dirofilaria immitis (Leidy 1856). Mem Inst Oswaldo Cruz. 2001;96:593–8.
Ledesema N, Harrington L. Fine-scale temperature fluctuation and modulations of Dirofilaria immitis larval development in Aedes aegypti. Vet Parasitol. 2015;209:93–100.
Hendrix CM, Brunner CJ, Bellamy LK. Natural transmission of Dirofilaria immitis by Aedes aegypti. J Am Mosq Control Assoc. 1986;2:48–51.
Ledesma NA, Kaufman PE, Xue RD, Leyen C, Macapagal MJ, Winokur OC, et al. Entomological and sociobehavioral components of heartworm (Dirofilaria immitis) infection in two Florida communities with high or low prevalence of dogs with heartworm infection. J Am Vet Med Assoc. 2019;254:93–103.
Vezzani D, Mesplet M, Eiras DF, Fontanarrosa MF, Schnittger L. PCR detection of Dirofilaria immitis in Aedes aegypti and Culex pipiens from urban temperate Argentina. Parasitol Res. 2011;108:985–9.
Lee ACY, Montgomery SP, Theis JH, Blagburn BL, Eberhard ML. Public health issues concerning the widespread distribution of canine heartworm disease. Trends Parasitol. 2010;26:168–73.
Bowman DD, Liu Y, McMahan CS, Nordone SK, Yabsley MJ, Lund RB. Forecasting United States heartworm Dirofilaria immitis prevalence in dogs. Parasit Vectors. 2016;9:1–12.
Self SCW, Liu Y, Nordone SK, Yabsley MJ, Walden HS, Lund RB, et al. Canine vector-borne disease: mapping and the accuracy of forecasting using big data from the veterinary community. Anim Health Res Rev. 2019;20:47–60.
Companion Animal Parasite Council (CAPC). 2022. https://capcvet.org/ Accessed 15 Jan 2022.
Wang D, Bowman DD, Brown HE, Harrington LC, Kaufman PE, Nelsen CT, et al. Factors influencing US canine heartworm (Dirofilaria immitis) prevalence. Parasit Vectors. 2014;7:264.
Drake J, Wiseman S. Increasing incidence of Dirofilaria immitis in dogs in USA with focus on the southeast region 2013–2016. Parasit Vectors. 2018;11:39.
Sobotyk C, Nguyen N, Negrón V, Varner A, Saleh MN, Hilton C, et al. Detection of Dirofilaria immitis via integrated serological and molecular analyses in coyotes from Texas, United States. Int J Parasitol Parasites Wildl. 2022;18:20–4.
Custer JW, Pence DB. Dirofilariases in wild canids from the gulf coastal prairies of Texas and Louisiana USA. Vet Parasitol. 1981;8:71–82.
Paras KL, Little SE, Reichard MV, Reiskind MH. Detection of Dirofilaria immitis and Ehrlichia species in coyotes (Canis latrans), from rural Oklahoma and Texas. Vector Born Zoonotic Dis. 2012;12:7.
Bowman D, Little SE, Lorentzen L, Shields J, Sullivan MP, Carlin EP. Prevalence and geographic distribution of Dirofilaria immitis, Borrelia burgdorferi, Ehrlichia canis, and Anaplasma phagocytophilum in dogs in the United States: results of a national clinic-based serologic study. Vet Parasitol. 2009;160:138–48.
Martin E, Medeiros MCI, Carbajal E, Valdez E, Juarez JG, Garcia-Luna S, et al. Surveillance of Aedes aegypti indoors and outdoors using Autocidal Gravid Ovitraps in South Texas during local transmission of Zika virus, 2016 to 2018. Acta Trop. 2019;192:129–37.
Hargrove WL, Juárez-Carillo PM, Korc M. Healthy vinton: a health impact assessment focused on water and sanitation in a small rural town on the US-Mexico border. Int J Environ Res Public Health. 2015;12:3864–88.
National Oceanic and Atmospheric Administration (NOAA). National Weather Service: Climate Prediction Center. Local climatological data, McAllen Miller Int Airport, TX. https://w2.weather.gov/climate/xmacis.php?wfo=bro. Accessed 20 July 2022.
Curtis-Robles R, Zecca IB, Roman-Cruz V, Carbajal ES, Auckland LD, Flores I, et al. Trypanosoma cruzi (agent of Chagas disease) in sympatric human and dog populations in “colonias” of the Lower Rio Grande Valley of Texas. Am J Trop Med. 2017;96:805–14.
Little SE, Raymond MR, Thomas JE, Gruntmeir J, Hostetler JA, Meinkoth JH, et al. Heat treatement prior to testing allows detections of antigen of Dirofilaria immitis in feline serum. Parasit Vectors. 2014;7:1.
Little S, Saleh M, Wohltjen M, Nagamori Y. Prime detection of Dirofilaria immitis: understanding the influence of blocked antigen on heartworm test performance. Parasit Vectors. 2018;11:186.
Sobotyk C, Savadelis MD, Verocai GG. Detection and cross-reaction of Dirofilaria repens using a commercial heartworm antigen test kit. Vet Parasitol. 2021;289:109302.
Wongkamchai S, Monkong N, Mahannol P, Taweethavonsawat P, Loymak S, Foongladda S. Rapid detection and identification of Brugia malayi, B. pahangi, and Dirofilaria immitis by high-resolution melting assay. Vector Borne Zoonotic Dis. 2013;13:31–6.
Rojas A, Rojas D, Montenegro VM, Baneth G. Detection of Dirofilaria immitis and other arthropod-borne filarioids by an HRM real-time qPCR, blood-concentrating techniques and a serological assay in dogs from Costa Rica. Parasit Vectors. 2015;8:170.
Laidoudi Y, Davoust B, Varloud M, Niang EHA, Fenollar F, et al. Development of a multiplex qPCR-based approach for the diagnosis of Dirofilaria immitis, D. repens and Acanthocheilonema reconditum. Parasit Vectors. 2020;13:9.
Fox M. Illustrated key to common mosquitoes of Louisiana. In: Mosquito control training manual. Baton Rouge: Louisiana Mosquito Control Association; 2007. p. 86–150.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
Rishniw M, Barr SC, Simpson KW, Frongillo MF, Franz M, Dominguez Alpizar JL. Discrimination between six species of canine microfilariae by a single polymerase chain reaction. Vet Parasitol. 2006;135:303–14.
American Heartworm Society. Our vision: a world without heartworms. 2022. https://www.heartwormsociety.org/ Accessed 15 Jan 2022.
Brown HE, Harrington LC, Kaufman PE, McKay T, Bowman DD, Nelsen CT, et al. Key factors influencing canine heartworm, Dirofilaria immitis, in the United States. Parasit Vectors. 2012;5:245.
Hodo CL, Rodriguez JY, Curtis-Robles R, Zecca IB, Snowden KF, Cummings KJ, et al. Repeated cross-sectional study of Trypanosoma cruzi in shelter dogs in Texas, in the context of Dirofilaria immitis and tick-borne pathogen prevalence. J Vet Intern Med. 2019;33:158–66.
Donnett U, Hubbard K, Woodruff K, Varela-Stokes A. Prevalence of canine heartworm infection in Mississippi animal shelters. Vet Parasitol. 2018;259:68–73.
Tzipory N, Crawford PC, Levy JK. Prevalence of Dirofilaria immitis, Ehrlichia canis, and Borrelia burgdorferi in pet dogs, racing greyhounds, and shelter dogs in Florida. Vet Parasitol. 2010;171:136–9.
Spence Beaulieu MR, Federico JL, Reiskind MH. Mosquito diversity and dog heartworm prevalence in suburban areas. Parasit Vectors. 2020;13:12.
Hampshire VA. Evaluation of efficacy of heartworm preventive products at the FDA. Vet Parasitol. 2005;133:191–5.
Courtney CH, Zeng QY. Comparison of heartworm antigen test kit performance in dogs having low heartworm burdens. Vet Parasitol. 2001;96:317–22.
Bowman D, Atkins C. Heartworm biology, treatment, and control. Vet Clin Small Anim. 2009;39:1127–58.
Negron V, Saleh MN, Sobotyk C, Luksovsky JL, Harvey TV, Verocai GG. Probe-based qPCR as an alternative to modified Knott’s when screening dogs for heartworm (Dirofilaria immitis) infection in combination with antigen detection tests. Parasit Vectors. 2022;15:306.
Lane JN, Litster A, Little SE, Rodriguez JY, Mwacalimba KK, Sundstrom KD, et al. Optimizing heartworm diagnosis in dogs using multiple test combinations. Parasit Vectors. 2021;14:1495.
Panetta JL, Davies Calvani NE, Orr B, Gianfranco Nicoletti A, Ward MP, Slapeta J. Multiple diagnostic tests demonstrate an increased risk of canine heartworm disease in northern Queensland, Australia. Parasit Vectors. 2021;14:393.
Alho AM, Landum M, Ferreira C, Meireles J, Goncalves L, de Carvalho M, et al. Prevalence and seasonal variations of canine dirofilariosis in Portugal. Vet Parasitaol. 2014;206:99–105.
Carmichael J, McCall S, DiCosty U, Mansour A, Roycroft L. Evaluation of Dirofilaria immitis antigen detection comparing heated and unheated serum in dogs with experimental heartworm infections. Parasit Vectors. 2017;10:486.
Taylor AER. The Development of Dirofilaria immitis in the mosquito Aedes aegypti. J Helminthol. 1960;34:27–38.
Palmer CA, Wittorck DD, Christensen BM. Ultrastructure of Malpighian tubules of Aedes aegypti infected with Dirofilaria immitis. J Invertebr Pathol. 1986;48:310–7.
Paras KL, O’Brien VA, Reiskind MH. Comparison of the vector potential of different mosquito species for the transmission of heartworm, Dirofilaria immitis, in rural and urban areas in and surrounding Stillwater, Oklahoma, USA. Med Vet Entomol. 2014;28:60–7.
Mckay T, Bianco T, Rhoes L, Bernett S. Prevalence of Dirofilaria immitis (Nematoda: Filarioidea) in mosquitoes from northeast Arkansas, the United States. J Med Entomol. 2013;50:871–8.
Huang S, Smith DJ, Molaei G, Andreadis TG, Larsen SE, Lucchesi EF. Prevalence of Dirofilaria immitis (Spirurida: Onchocercidae) infection in Aedes, Cluex, and Culiseta mosquitoes from North San Joaguin Valley, CA. J Med Entomol. 2013;50:1315–23.
Theis JH. Public health aspects of dirofilariasis in the United States. Vet Parasitol. 2005;133:157–80.
Skidmore JP, Dooley PD, DeWitt C. Human extrapulmonary dirofilariasis in Texas. South Med J. 2000;93:1009–10.
Khan SU, Ogden NH, Fazil AA, Gachon PH, Dueymes GU, et al. Current and projected distributions of Aedes aegypti and Ae. Albopictus in Canada and the US. Environ Health Perspect. 2020;128:057007.
Acknowledgements
We thank all dog owners, community members and promotoras who made this study possible. We thank Jose G. Juarez, Selene Garcia-Luna, Estelle Martin, Ester Carbajal, Edwin Valdez and Justin Bejcek who contributed to dog and mosquito sample collection. We thank Lisa Auckland, Keswick Killets and Valery Roman-Cruz for help processing samples. We also thank Jose G. Juarez for help with map development, and Wendy Tang who assisted with laboratory diagnostics.
Funding
This study was funded by the American Kennel Club Canine Health Foundation Grant No. 02821-A and Texas A and M AgriLife Research.
Author information
Authors and Affiliations
Contributions
Project administration: SH, GV and GH. Investigation: NS, IZ, CS, MS, SL and MO. Formal analysis: NS. Visualization: NS. Writing—original draft preparation: NS. Writing—review and editing: NS, CS, MS, SH, GV and GH. Supervision: GH. All authors have read and approved of the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Canine sampling and vaccination protocols were approved by the Institutional Committee on Animal Use and Care at Texas A&M University (AUP# 20180460 and #2015-0289 CA). The ARRIVE guidelines applicable to observational field-based studies were followed (https://arriveguidelines.org/).
Consent for publication
Not applicable.
Competing interests
The authors have no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Scavo, N.A., Zecca, I.B., Sobotyk, C. et al. High prevalence of canine heartworm, Dirofilaria immitis, in pet dogs in south Texas, USA, with evidence of Aedes aegypti mosquitoes contributing to transmission. Parasites Vectors 15, 407 (2022). https://doi.org/10.1186/s13071-022-05514-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05514-1