
Abstract
Background
Vector control is the main intervention used to control arboviral diseases transmitted by Aedes mosquitoes because there are no effective vaccines or treatments for most of them. Control of Aedes mosquitoes relies heavily on the use of insecticides, the effectiveness of which may be impacted by resistance. In addition, rational insecticide application requires detailed knowledge of vector distribution, dynamics, resting, and feeding behaviours, which are poorly understood for Aedes mosquitoes in Africa. This study investigated the spatiotemporal distribution and insecticide resistance status of Aedes aegypti across ecological extremes of Ghana.
Methods
Immature mosquitoes were sampled from containers in and around human dwellings at seven study sites in urban, suburban, and rural areas of Ghana. Adult Aedes mosquitoes were sampled indoors and outdoors using Biogents BG-Sentinel 2 mosquito traps, human landing catches, and Prokopack aspiration. Distributions of immature and adult Aedes mosquitoes were determined indoors and outdoors during dry and rainy seasons at all sites. The phenotypic resistance status of Aedes mosquitoes to insecticides was determined using World Health Organization susceptibility bioassays. The host blood meal source was determined by polymerase chain reaction.
Results
A total of 16,711 immature Aedes were sampled, with over 70% found in car tyres. Significantly more breeding containers had Aedes immatures during the rainy season (11,856; 70.95%) compared to the dry season (4855; 29.05%). A total of 1895 adult Aedes mosquitos were collected, including Aedes aegypti (97.8%), Aedes africanus (2.1%) and Aedes luteocephalus (0.1%). Indoor sampling of adult Aedes yielded a total of 381 (20.1%) and outdoor sampling a total of 1514 (79.9%) mosquitoes (z = − 5.427, P = 0.0000) over the entire sampling period. Aedes aegypti populations were resistant to dichlorodiphenyltrichloroethane at all study sites. Vectors showed suspected resistance to bendiocarb (96–97%), permethrin (90–96%) and deltamethrin (91–96%), and were susceptible to the organophosphate for all study sites. Blood meal analysis showed that the Aedes mosquitoes were mostly anthropophilic, with a human blood index of 0.9 (i.e. humans, 90%; human and dog, 5%; dog and cow, 5%).
Conclusions
Aedes mosquitoes were found at high densities in all ecological zones of Ghana. Resistance of Aedes spp. to pyrethroids and carbamates may limit the efficacy of vector control programmes and thus requires careful monitoring.
Graphical abstract

Similar content being viewed by others

Background
Aedes aegypti and Aedes albopictus are the most important vectors of several arboviruses, notably yellow fever, dengue, chikungunya, and Zika virus [1]. The importance of Aedes mosquitoes in sub-Saharan Africa has increased recently because of outbreaks of arboviral diseases in multiple countries there [2]. In West Africa, within the last 5 years, there have been outbreaks of dengue in Burkina Faso [3,4,5], Cote d’Ivoire [6, 7], and Senegal [8], yellow fever in Cote d’Ivoire [9,10,11] and Nigeria [12,13,14,15,16], and recent confirmed cases of dengue and outbreaks of yellow fever in Ghana [17,18,19,20]. Therefore, the risk of dengue, yellow fever and chikungunya outbreaks in Ghana appears to be high.
Aedes aegypti are highly anthropophilic and, throughout most of the world, typically endophilic [21]. Immature stages develop preferentially in artificial containers, usually in close proximity to humans [22,23,24,25]. In sub-Saharan Africa, two morphological subspecies (ecotypes) have been acknowledged: domestic Aedes aegypti aegypti and sylvan Aedes aegypti formosus. The presence/absence of white abdominal scale patterns [26] is used to differentiate the ecotypes, but at present, clear genetic boundaries appear to be absent, probably as a result of recent and historical gene flow [27, 28]. Aedes aegypti formosus more frequently breed away from domestic environments, and feed readily on animals (zoophagy), so are less likely to be a threat to humans in the urban environments where Aedes aegypti aegypti populations thrive [29]. However, urbanisation of the sylvatic environment could lead to contact between Ae. aegypti formosus and humans, and this typically sylvatic species might adapt to new urban environments and hosts; it is also probable that introgression through hybridization of urban and sylvatic subspecies of Ae. aegypti may lead to variation, potentially increasing the role of Ae. aegypti formosus as vectors [28, 30]. The consistency of bionomic traits across ecozones remains poorly investigated. However, measures of abundance and distribution of Aedes would give more reliable insights for both risk and mitigation strategies for infestations [31]. Several species of Aedes, including Aedes africanus, readily feed on animals (both domestic and wild), as well as humans, hence their potential importance as bridge vectors and for zoonotic transmission [32]. Identification of the source of vector blood meals is critical to understanding the degree of human–vector interaction (i.e. anthropophily) [33, 34], which is a crucial parameter in the estimation of the capacity of a vector to transmit a disease [35].
Seasonal variations in population density are expected for Aedes, with lower abundances in dry seasons, rising with increasing temperatures, and potentially greater breeding site availability in the rainy season [36,37,38]. However, human activities involving water storage and the disposal of potential water-holding containers greatly influence the breeding of Aedes in individual households and may lead to the provision of breeding sites year-round [39]. Key factors that may influence Aedes productivity in different container types include the frequency of water replenishment, the availability of food for the larvae, the degree of sunlight exposure [40], and container coverings [41]. The adaptation of these vectors to urban domestic habitats has led to their exploitation of a range of artificial containers and their capacity to exploit potential breeding water situated indoors or outdoors [42, 43].
Currently, and despite frequent concerns regarding the efficacy of the methods used for their deployment [44], insecticidal interventions are the main tool used to control Aedes-borne arboviral infections, since vaccines for these are either unavailable, ineffective, or in limited supply [45,46,47]. To ensure that efficacy is maximised, correct insecticide choice is crucial, and requires surveillance to determine the susceptibility of target populations, alongside locating the adults and immatures that are to be targeted [48]. Sustained effectiveness must also be considered: geographical variation in susceptibility may rapidly lead to the spread of insecticide resistance and require revision regarding the most suitable insecticide for use.
Another important parameter when considering how to target vector control, especially for insecticidal spray deployment, is whether mosquitoes tend to rest indoors (endophily) or outdoors (exophily) after blood-feeding [49]. Insecticide-based intervention directed at the adult resting population is a relevant approach for Aedes control and disease prevention. Targeted indoor residual spraying on Aedes resting locations can provide a significant protective effect against arboviral transmission, and this method also has the potential to control pyrethroid-resistant Aedes mosquitoes, as other classes of insecticides (non-pyrethroids) are available for residual application [50].
This study aimed to characterize the breeding habitats, seasonal abundance, and resting behaviour of Ae. aegypti, and their insecticide susceptibility, in rural, suburban, and urban sites in different ecological zones of Ghana. In addition to identifying targets and options for control, the results will also aid the development of a surveillance system for Aedes as vectors of arboviruses for the planning of disease control in Ghana [51, 52].
Methods
Study sites
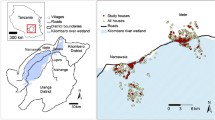
This study was carried out in seven sites comprising rural, suburban, and urban locations within the three major ecological zones of Ghana, i.e. coastal savannah, forest, and Sahel savannah, across wet and dry seasons, between May 2017 and May 2018. The selection of the study sites was based on both ecological zone and population (urban, suburban, and rural), since variations in species, abundance, and susceptibility status of Aedes mosquitoes could be influenced by these parameters. Residents in the rural study sites store water for domestic use in diverse containers because the water supply system is unreliable. Water supply in the rural study sites is mainly from harvested rainwater, wells, and boreholes, and these also supplement the irregular piped water supply system characteristic of our urban and suburban study sites. Furthermore, it is known that Ae. aegypti is adapted to urban settings while Ae. albopictus and other zoonotic species are found in the sylvatic zone. One of the sites has a port where car tyres are imported, thus may be a channel for the importation of species of Aedes from the Americas previously unknown in Ghana. The sample sites are shown in Fig. 1.

Map of Ghana showing the study sites
In the coastal savannah zone of southern Ghana the study sites were as follows: Ada Foah (suburban site; 5°47′N, 0°38′E), a tourist town; Accra (urban site; 5°33′0″N, 0°12′0″W), the capital of Ghana and its most populous city; and Tema (urban site; 5°40′0″N, 0°0′0″E), a port city where the importation of tyres from Asia and the Americas might facilitate the invasion of Aedes genotypes or species previously unknown in Ghana. The coastal savannah has a tropical savannah climate, with an annual mean temperature of 26.5 °C and average annual precipitation of 787 mm (Table 1).
The urban site within the forest zone was Konongo (06°37′00″N, 001°13′00″ W), a town located in Asante-Akim central district in the middle of Ghana. In the forest zone, there is a high possibility that sylvan Aedes mosquitoes, which can serve as bridge vectors, might be present. The forest zone has a tropical rainforest climate, with an annual average temperature of 26.4 °C and annual average precipitation of 1399.5 mm (Table 1).
The sites in the Sahel savannah ecological zone were Larabanga, Navrongo, and Paga. Larabanga (rural site; 9°5′0″N, 1°49′0″W) is a village situated close to Mole national park, which harbours monkeys that could serve as reservoirs for arboviruses, and has experienced yellow fever outbreaks [53]. Navrongo (urban site; 10°53′5″N, 01°05′25″W) is a town close to the border between Ghana and Burkina Faso; an outbreak of dengue fever was relatively recently reported in Burkina Faso, in the period between 2016 and 2019 [54]. The last site was Paga (suburban site; 10°59′32″N, 01°06′48″W), a small town located on the border of Burkina Faso and 166 km south of Ouagadougou, where an outbreak of dengue fever was recently reported [54] (Table 1).
In both the coastal savannah and forest area there is generally a bimodal pattern of rainfall, with the long rainy season from April to June, and a short rainy season from October through November. Rainfall in the Sahel savannah is unimodal, with the rainy season between May and November and the dry season from December to April. Sampling was done during the rainy season from April through June 2017 in the coastal savannah and forest zone, from May through June 2018 in the Sahel savannah, during the dry season from January through March 2018 in the coastal savannah and forested savannah, and from December 2017 through January 2018 in the Sahel savannah.
Distribution of immature Aedes mosquitoes
Exhaustive entomological surveys were carried out at each of the study sites. Water-holding containers in and around human dwellings were inspected for immature Aedes in the dry and rainy seasons, and those positive for Aedes immatures recorded. Larval habitats were sampled once per season. Every possible Aedes breeding container was inspected for the presence of Aedes immatures at each site. Because it was difficult to sample most of the containers in which the Aedes bred by dipping, all Aedes immatures encountered at each breeding habitat were collected to determine the density of vectors. All pupae and larvae (first to fourth instars) from positive containers (air conditioner saucers, car tyres, drums, tanks, buckets, and discarded containers) were collected using pipettes and ladles [39, 55], counted, and recorded on field data forms. Water from large containers was first sieved and larval samples placed in a white plastic tray with some of the water in which they were pipetted. Mosquito samples were placed immediately in labelled specimen bowls filled with water from the container from which they had been collected, and transported to the insectary. Immature mosquitoes were reared in the insectary in large white plastic trays at an average temperature of 28.15 ± 1.8 °C (± SD) and relative humidity of 80.9 ± 6.3%, and larvae were fed on TetraMin Baby fish food (Tetra Werke, Melle, Germany).
Adult female Aedes mosquitoes that emerged from the collected larvae were used for the World Health Organization (WHO) susceptibility bioassays [56] and later identified morphologically using standard taxonomic keys [57]. Coordinates of all collection points were recorded using a GPSMAP 60CSx Geographical Positioning System (GPS) instrument (Garmin International, Olathe, KS).
Characterization of Aedes breeding habitats and relative abundance
For each entomological survey, the habitat type, its household location (indoors or outdoors), and its physical characteristics were recorded. Six container types were classified based on their use and material: car tyres, air conditioner saucers, discarded containers, drums, tanks, and buckets. Air conditioner saucers are small (1–2 L) plastic containers positioned below the outlet of air conditioners to collect water. Discarded containers were defined as 50- to 100-L-capacity containers, which included broken jars, bottles, small plastic food containers, tins, plates, cans, cooking pots, and broken pots made of plastic or metal. Drums were defined as 100- to 500-L-capacity plastic water storage containers. Tanks were defined as 100- to 500-L-capacity water storage containers made of metal or concrete. Buckets comprised 10- to 25-L water storage containers made of metal or plastic. It is notable that pipeborne water was absent in Larabanga and Paga, with the consequence that households tend to have long-term water storage in tanks, drums, buckets, and pots, especially during the dry season, which may serve as potential Aedes larval breeding habitats (Fig. 2).

Habitat types encountered during the larval survey and sampling: a air conditioner saucer, b car tyres, c buckets, d tank, e discarded container, and f drum
Distribution of adult Aedes mosquitoes
The spatial distribution of adult Aedes mosquitoes was determined by sampling inside households and outdoors. In the Sahel savannah zone, Aedes mosquitoes were sampled indoors and outdoors using three methods that employed one of the following: BG-Sentinel 2 traps (BG traps; Biogents, Regensburg, Germany); human landing catches (HLC); or Prokopack Aspirators (PPK) (Hock, Gainesville, FL) [58]. The relative trapping efficiency of the three sampling methods, i.e. using BG traps, HLC, or PPK, was compared for the Sahel savannah zone to determine the most efficient vector sampling tool for future surveillance. Due to logistical challenges, in the coastal and forest ecological zones, only two of the methods were employed, the one which used BG traps and the one which used HLC.
The GPS coordinates of all collection points were recorded. Two cross-sectional surveys were undertaken, one in the dry season (December 2017–March 2018) and one in the rainy season (April–June 2017 and May–June 2018).
Adult mosquito collection using BG traps, HLC and PPK
BG traps were set both indoors [living room(s) and bedrooms] and outdoors (open but secure verandas, granaries, or under a shed/tree where people sit to chat, about 5 m from the house) from 5:00 to 8:00 a.m. and 3:00–7:00 p.m. The BG traps were baited with CO2 which was produced by either BG-Lures and/or from a mixture of 17.5 g yeast [Angel Yeast (Egypt)] and 250 g sugar in 1 L of water [59]. The mosquito collection net of the BG trap was changed at 1-h intervals. Mosquitoes trapped within the collection net were placed in a cooler box containing ice and then transported to the insectary.
The HLC method was also used to sample host-seeking adult Aedes mosquitoes. On each day, two trained volunteers were positioned indoors and two were positioned outdoors to catch Aedes mosquitoes. The collected Aedes were placed in labelled paper cups, which were placed in cool boxes with ice packs and transported to the insectary for identification and further processing.
Prokopack aspiration was employed at the three sites in the Sahel savannah area: Larabanga, Navrongo, and Paga. Sixteen houses were randomly selected for Aedes collection per site. Aedes mosquitoes were sampled indoors and outdoors. Aedes caught within the Prokopack plastic collection cups were labelled and placed in a cooler box containing an ice pack and transported to the insectary for identification. The heights at which the mosquitoes were caught by PPK while resting were recorded using a tape measure, to determine whether there was heterogeneity in resting height among sites.
Sixteen houses were randomly selected for each sampling method at each site. Sampling using each type of sampling tool was done on 4 different days during each season. On each of the sampling days, the houses used for sampling had not previously been used for this purpose. Mosquito sample collection using PPK and HLC was done hourly from 5:00 to 8:00 a.m. and then from 3:00 to 7:00 p.m.
Meetings were held at each study site with chiefs and residents to introduce the research study to the public. All participants in this study were adults (> 18 years old). Written and signed consent was obtained from all of the adults who volunteered to participate in HLC before they were trained and the mosquito sampling began. A copy of the signed consent form was given to each of the HLC volunteers and another copy kept in a locked cabinet with restricted access in the offices of the Department of Medical Microbiology, University of Ghana Medical School. Verbal and written consent was obtained from household heads to sample mosquitoes in their houses and compounds. All volunteers were remunerated at the end of the study.
Insecticide susceptibility tests
Aedes larvae were collected from natural breeding sites or from oviposition traps that were set in each site. Oviposition traps were made from used car tyres that had been cut into three parts which could hold about 3 L of water each. Oviposition traps were set for the collection of Aedes immatures during time points when it was difficult to obtain sufficient numbers for the WHO susceptibility bioassays. Collected larvae were brought to the insectary at the Department of Medical Microbiology, University of Ghana, and were raised to adults under standard conditions (25 ± 2 °C, 80% ± 4% relative humidity, 12-h:12-h light/dark cycle). Batches of 20–25 non-blood-fed 3- to 5-day-old females were used for the susceptibility bioassays. Four replicates and two controls were used for each insecticide tested using the standard WHO susceptibility bioassay procedure [56].
The WHO test papers were impregnated with a pyrethroid (0.05% deltamethrin or 0.75% permethrin), an organochloride [4% dichlorodiphenyltrichloroethane (DDT)], an organophosphate (5% malathion), or a carbamate (0.1% bendiocarb) insecticide. These pre-impregnated papers are supplied with diagnostic concentrations based on those required for Anopheles. As the concentrations of permethrin and malathion of the test papers are three times and approximately six times, respectively, the diagnostic concentrations required for Aedes mosquitoes, this was a limitation of the study. However, these test papers are far more commonly used for the assessment of the susceptibility of Aedes to these insecticides than those custom produced at the recommended concentrations [60]. The knockdown time was reported every 10 min during the 60-min exposure period. Mortality was recorded after the 24-h recovery period. ‘Resistant’ mosquitoes were defined as mosquitoes that survived for 24 h after the end of the bioassay, and ‘susceptible’ mosquitoes as those that were knocked down or died during the 60-min exposure time, or that died within the 24-h recovery period.
Data analysis
Descriptive analysis was performed to compare larval and adult abundance between different populations (urban, suburban and rural), indoors and outdoors, and seasons.
The abundances of Aedes larvae and adults were compared among the seasons, indoor and outdoor study sites (ecozones), and sampling methods (adults). For all sites, a Kruskal–Wallis test was used to compare the abundance of adult mosquitoes for HLC and BG traps, and a Wilcoxon rank-sum test was used to test for associations between continuous and categorical variables. Nested generalized linear mixed models with sites nested within ecological zones were used to model the effect of ecozone, season, population (urban, suburban and rural), and sampling methods on larval and adult abundance. A regression analysis was done to test trap efficacy. Probability values less than 0.05 were interpreted as statistically significant.
Human blood index was calculated as the proportion of positive human blood specimens per total number of specimens tested.
Insecticide susceptibility was classified using the following WHO criteria [56]: 98–100% mortality, the test population is considered susceptible; 90–97% mortality, possible resistance of the test population (which requires confirmation); below 90% mortality, the test population is considered resistant. Knockdown and mortality rates were compared between sites using Chi-square. Statistical analysis was performed using Stata 16 (StataCorp, College Station, TX).
Results
Larval breeding habitats and their productivity
A total of 81 positive breeding habitats were identified during the study period across the seven sites (only positive breeding habitats were recorded). Generalized linear model analysis revealed a significant interaction effect between ecozone and population on abundance. Compared to the other sites, the chance of sampling Aedes larvae was higher in the forest zone [unadjusted (unadj.) B = − 204.12 (− 306.01 to − 102.24), P = 0.000]. The abundance of Aedes larvae was higher in suburban areas [unadj. B = − 138.01 (− 224.77 to − 51.26), P = 0.002] than in urban areas (Table 2).
There were significantly more positive habitats during the rainy season than during the dry season (n = 50 vs n = 31; df = 5, χ2 = 19.44, P = 0.001; Table 3). Within the seven sites sampled, 78 (96.3%) of the larval breeding habitats were located outdoors and three (3.7%) indoors (all in Larabanga) (Table 3), with larval abundances of 16,426 (98.3%) and 285 (1.7%), respectively (n = 78 vs n = 3; z = − 0.138, P = 0.8903).
A total of 16,711 Aedes immatures were collected over the entire sampling period, of which 12,348 (73.9%) were from car tyres, 3138 (18.8%) from discarded containers, 730 (4.4%) from air conditioner saucers, 230 (1.4%) from buckets, 210 (1.3%) from tanks, and 55 (0.3%) from drums (χ2 = 1.020, df = 5, P = 0.96; Table 4). For all the different sites, car tyres had the highest proportion, 71.3% (8453), of immatures during the rainy season. The same observation was made during the dry season, with the highest abundance of Aedes immatures, 3895 (80.2%) (χ2 = 2.106, df = 2, P = 0.3490; Table 4), in car tyres.
Regarding the different ecological zones, significantly more Aedes immatures were collected from the coastal savannah (9819; 58.8%), followed by the Sahel savannah (5794; 34.7%), and then the forest zone (1098; 6.6%) (χ2 = 16.071, df = 2, P = 0.0003). A higher proportion of immature Aedes was collected in urban areas, with an abundance of 10,876 (65.1%) (Tema, 4338; Accra, 3670; Navrongo, 1770; Konongo, 1098), followed by the suburban areas, with a total of 3890 (23.3%) (Paga, 2079; Ada Foah, 1811), and then rural areas, with a total abundance of 1945 (11.6%) (Larabanga, 1945) (χ2 = 10.040, df = 2, P = 0.0066). We found more Aedes immatures outdoors (16,426; 98.3%) than indoors (285; 1.7%) (z = − 0.138, P = 0.8903).
Seasonal distribution of adult Aedes mosquitoes
A total of 1895 adult Aedes mosquitoes were collected from the study sites. Generalized linear model analysis revealed a significant interaction effect between outdoor collection, ecozone and population (urban, suburban, and rural) on abundance. The chance of sampling adult Aedes mosquitoes increased outdoors [adj. B = 1.49 (1.0271–1.9602), P = 0.000]. There was a significant difference between adult Aedes mosquito abundance in suburban sites [adj. B = − 1.49 (− 2.0433 to − 0.9320), P = 0.000] and urban sites (Table 5). Adult Aedes were more abundant during the rainy season (1257; 66.3%) compared to the dry season (638; 33.7%) (z = − 1.433, P = 0.1519). Across the different ecological zones, the abundance of Aedes was high in the coastal savannah (955; 50.4%) [Accra (urban), 718; Tema (urban), 161; Ada Foah (suburban), 76], followed by the Sahel savannah 837 (44.2%) [Navrongo (urban), 577; Paga (suburban), 173; Larabanga (rural), 87], and the forest zone (103; 5.4%) [Konongo (urban), 103] (\(\chi\)2 = 0.359, df = 2, P = 0.835). The urban sites had the highest abundances of Aedes mosquitoes (1559, 82.3%) (Accra, 718; Tema, 161; Konongo, 103; Navrongo, 577) followed by the suburban sites (249; 13.1%) (Ada Foah, 76; Paga, 173), and then the rural site (87; 4.6%) (Larabanga, 87) (\(\chi\)2 = 20.147, df = 2, P = 0.0001).
At the different sites during the dry season, the highest abundance of Aedes mosquitoes was found in Accra (178; 27.9%) (HLC, 163; BG trap, 15), followed by Navrongo (173; (27.1%) (HLC, 157; BG trap, 16), Tema (108; 16.9%) (HLC, 102; BG trap, 6), Konongo (103; 16.1%) (HLC, 88; BG trap, 15), Ada (60; 9.4%) (HLC, 50; BG trap, 10), Larabanga (15; 2.4%) (HLC, 0; BG trap, 15), and then Paga (1; 0.2%) (HLC, 1; BG trap, 0) (\(\chi\)2 = 20.500, df = 6, P = 0.0023).
During the rainy season, the highest abundance of Aedes mosquitoes was found in Accra (540; 43.0%) (HLC, 499; BG trap, 41), followed by Navrongo (404; 32.1%) (HLC, 354; BG trap, 50), Paga (172; 13.7%) (HLC, 168; BG trap, 4), Larabanga (72; 5.7%) (HLC, 54; BG trap, 18), Tema (53; 4.2%) (HLC, 31; BG trap, 22), Ada Foah (16; 1.3%) (HLC, 0; BG trap, 16), and then Konongo (0, 0%) (HLC, 0; BG trap, 0), (\(\chi\)2 = 132.896, df = 6, P = 0.0001).
Indoor and outdoor abundance of adult Aedes populations
Overall mosquito abundance was highest outdoors as compared to indoors over the entire sampling period. Indoor sampling yielded a total of 381 (20.1%) and outdoor sampling a total of 1514 (79.9%) adult Aedes over the entire sampling period (z = − 5.427, P = 0.0000). During the rainy season, a higher proportion of Aedes mosquitoes was captured outdoors (77.8%; 978) than indoors (22.2%; 279) (z = − 2.989, P = 0.0028). Similarly, a greater number of Aedes mosquitoes were captured outdoors (536; 84%) than indoors (102; 16%) during the dry season (z = − 5.021, P = 0.0000; Fig. 3).

Seasonal distribution of adult Aedes mosquitoes captured indoors (IN) and outdoors (OUT). BG BG-Sentinel 2 trap, HLC human landing catches
Comparison of trap efficiency for Aedes mosquito sampling
A total of 1140 Aedes mosquitoes were collected by HLC, BG traps, and PPK in Larabanga, Navrongo, and Paga during the experiment. Overall, the abundance of adult Aedes mosquitoes was 2.4 times higher for HLC (734) compared to PPK (303), and 7.1 times higher for HLC compared to BG traps (103). There was a significant difference between the abundance of Aedes mosquitoes for HLC and BG traps [P = 0.000, 95% confidence interval (CI) = − 2.180248 to − 0.7245137], but no significant difference between HLC and PPK (P = 0.350, 95% CI = − 0.4820909 to 1.359284; Table 6).
Generalized linear model analysis revealed a significant interaction effect between outdoor collection, study site and sampling method on abundance. More adult Aedes mosquitoes were collected outdoors [adj. B = 0.87 (0.22, 1.52), P = 0.009]. Adult Aedes mosquitoes were more abundant in Navrongo [adj. B = 0.83 (0.07, 1.58), P = 0.032], and BG traps were the least efficient means of collecting Aedes mosquitoes [adj. B = − 1.39 (− 2.14, − 0.64), P < 0.001; Table 7].
Resting height of Aedes mosquitoes
The maximum height at which Aedes mosquitoes were caught resting was 5 m and the lowest 1 m. The mean preferred resting height of the caught Aedes mosquitoes ranged from 1.8 to 2.0 m indoors and 1.3–2.8 m outdoors (\(\chi\) 2 = 1.408, df = 2, P = 0.4945). No mosquito was caught resting indoors in Navrongo (Table 8).
Aedes species composition at the study sites
Morphological identification of all collected adult Aedes showed that Aedes aegypti (1854; 97.8%) was the most abundant species present at all sites followed by Aedes africanus (40; 2.1%) and Aedes luteocephalus (1; 0.01%) (Table 9). All 11,506 Aedes mosquitoes that emerged from the larvae collected from the sites and reared in the insectary were identified morphologically as Aedes aegypti (Table 9).
Blood meal analysis
Blood meal analysis was carried out on blood-fed mosquitoes that were sampled using BG traps and PPK in Larabanga, Navrongo, and Paga. PCR amplification of DNA segments of 20 of 44 blood-fed mosquitoes showed that 18 (90%) had taken a human blood meal, one (5%) had fed on a human and a cow, and one (5%) had taken blood meals from a dog and a goat.
Phenotypic resistance of Aedes to insecticides
Phenotypic test results showed that Aedes mosquito populations from all study sites were resistant to DDT (range 0–88%). The highest level of DDT resistance was seen in Tema, where none of the mosquitoes died on exposure to this insecticide. The vectors showed resistance to permethrin in Tema (21%), Accra (40.0%) and Larabanga (89%), and suspected resistance in Navrongo (90%), Paga (96%) and Konongo (90%) (\(\chi\) 2 = 1.331, df = 12, P = 0.0001). Aedes mosquitoes showed resistance to deltamethrin in Tema (68%) and suspected resistance in Accra (91.3%), Ada Foah (94%), Konongo (94%), Larabanga (93%), Navrongo (96%) and Paga (93%) (\(\chi\) 2 = 560.000, df = 6, P = 0.0001). Aedes mosquitoes were resistant to bendiocarb in Larabanga (81%), showed suspected resistance in Tema (95.0%), Konongo (96%), Navrongo (96%) and Paga (97%), and were susceptible to bendiocarb in Accra and Ada Foah (\(\chi\) 2 = 1.331, df = 12, P = 0.0001). Aedes mosquitoes were susceptible to the organophosphate (malathion) at all sites (Fig. 4).

Phenotypic resistance status of Aedes mosquitoes to different insecticides. DDT Dichlorodiphenyltrichloroethane
Discussion
Seasonal variation in population density is common for Aedes mosquitoes due to their sensitivity to changes in temperature and moisture [53, 61]. This study found a significantly higher abundance of Aedes immatures during the rainy season. The rains may have resulted in an increase in aquatic habitats for Aedes spp. that breed in car tyres and the other types of suitable containers that were encountered in this study [62], and thus an increase in the abundance of Aedes immatures due to an increase in the rate of oviposition. This finding was similar to that of Ngugi et al. [39], who found a higher abundance of immatures during the rainy season in Kenya. However, Appawu et al. [53] showed an increase in the abundance of Aedes immatures during the dry season in Ghana.
Car tyres, buckets, tanks, drums, discarded containers, and air conditioner saucers that had collected water were the main breeding sites and supported the development of Aedes immatures in all or some of the study sites. The distribution of Aedes immatures between container types varied between the dry and rainy seasons. In all, only car tyres could be considered key breeding habitats in both seasons at all sites, and over 70% of the Aedes immatures were collected from these during the study period. The abundance of Aedes immatures in car tyres found here is consistent with the findings of a study conducted in the Central African Republic [63], where used car tyres were the most heavily colonized and productive larval habitats for Aedes in both early and late wet seasons. Car tyres should therefore be targeted for vector control to eliminate most Aedes immatures. Habitats that were non-productive were not included in this study. Larval indices (container index, house index, and Breteau index) were not calculated because we did not record habitats that did not have larvae in them, which was a major limitation of this study.
The WHO susceptibility bioassays showed that Aedes mosquito populations from all of the study sites were resistant to DDT. This finding is similar to that of a previous study done in Accra, Ghana, which showed that Ae. aegypti were resistant to DDT [64]. Deltamethrin, permethrin, and bendiocarb resistance were also recorded in the present study. As these are some of the most widely used insecticides for the control of Aedes spp. [65, 66], their use in Ghana could negatively affect the efficacy of vector control efforts there. Deltamethrin and permethrin resistance could result from the widespread use of pyrethroids for the impregnation of bed nets and for indoor residual spraying against malaria vectors. Pyrethroid resistance has been recently reported in an Aedes population in Ghana [64]. Cross-resistance between pyrethroids and DDT is also known to occur [29]. A previous study in Ghana [64] found Aedes to be susceptible to permethrin. Other studies have also reported pyrethroid resistance in Aedes spp. in Africa and Asia [67,68,69].
In this study, Ae. aegypti was the predominant species in all of the study areas for both adult and larval sampling. The high number of Ae. aegypti caught from a geographically wide range of sites in different ecological zones may be explained by the behaviour of this species, which is highly anthropophilic and associated with human habitations, as reported by many studies undertaken in Central Africa and West Africa [64, 68, 70]. The high abundance of Ae. aegypti observed in the present study implies that this vector of yellow fever and dengue is well established in Ghana, which could increase the potential for the transmission of arboviral diseases across the country because outbreaks of these are more severe in the absence of effective vector control.
This calls for constant vector monitoring in Ghana to prevent outbreaks of arboviral diseases. The zoonotic Aedes species Ae. africanus and Ae. luteocephalus, which are involved in sylvatic cycles between non-human primates, were also found in this study, which suggests that they may act as bridge vectors and carry disease between sylvan and domestic environments, as both are vectors of yellow fever in Ghana [64]. In addition, Hanley et al. [71] reported that Ae. africanus is the primary vector of sylvatic yellow fever virus in the rainforests of Central Africa, extending outward along the riverine forests there, and Diallo et al. [72] detected yellow fever in the zoonotic species Ae. luteocephalus in Senegal. Suzuki et al. [64] found only one Ae. albopictus in Accra, Ghana, and in our current study we did not find any individuals of this species in any of the study sites. The absence of this species from our samples suggests that environmental factors in Ghana may not be favourable for its establishment and proliferation. The dominance of its sister species, Ae. aegypti, may result from favourable environmental conditions for its proliferation.
The present study showed that Ae. aegypti rests at an average height of 1.8–2.0 m indoors and 1.3–2.8 m outdoors. These observations are quite different from those of studies undertaken in Iquitos, Peru [58] and Acapulco, Mexico [73], which showed that of 56 and 626 Ae. aegypti collected indoors, respectively, 82% rested at a height of less than 1.5 m. Resting height may have major implications for the efficacy of indoor residual spraying due to the exophilic behaviours of some Aedes mosquitoes. Insecticide pressure indoors through indoor residual spraying may also trigger exophagy and thus outdoor transmission of arboviruses.
Conclusions
The results of this study indicate that breeding habitats of Ae. aegypti in Ghana are abundant outdoors and are diverse across the country. Car tyres were the most productive containers for Aedes, as > 70% of the Aedes larvae were collected from these. Thus, targeting tyres in source reduction efforts may be a cost-effective means of reducing the risk of arboviral disease transmission in Ghana. Resistance of Ae. aegypti to pyrethroids and carbamates requires careful monitoring as it could limit the efficacy of vector control programmes. Management strategies for vector control that take into account insecticide resistance are thus urgently needed for Ghana.
Availability of data and materials
All datasets generated and/or analysed during this study are available on request.
Abbreviations
- BG:
-
BG-Sentinel 2
- DDT:
-
Dichlorodiphenyltrichloroethane
- GPS:
-
Geographical Positioning System
- HLC:
-
Human landing catch
- PPK:
-
Prokopack Aspirator
- WHO:
-
World Health Organization
References
Farraudiere L, Sonor F, Crico S, Etienne M, Mousson L, Hamel R, et al. First detection of dengue and chikungunya viruses in natural populations of Aedes aegypti in Martinique during the 2013–2015 concomitant outbreak. Rev Panam Salud Publica. 2017;41:e63.
Braack L, Gouveia de Almeida AP, Cornel AJ, Swanepoel R, de Jager C. Mosquito-borne arboviruses of African origin: review of key viruses and vectors. Parasit Vectors. 2018;11:29.
Tarnagda Z, Cisse A, Bicaba BW, Diagbouga S, Sagna T, Ilboudo AK, et al. Dengue fever in Burkina Faso, 2016. Emerg Infect Dis. 2018;24:170–2.
Lee JS, Mogasale V, Lim JK, Ly S, Lee KS, Sorn S, et al. A multi-country study of the economic burden of dengue fever based on patient-specific field surveys in Burkina Faso, Kenya, and Cambodia. PLoS Negl Trop Dis. 2019;13:e0007164.
Ouattara LPE, Sangare I, Namountougou M, Hien A, Ouari A, Soma DD, et al. Surveys of arboviruses vectors in four cities stretching along a railway transect of Burkina Faso: risk transmission and insecticide susceptibility status of potential vectors. Front Vet Sci. 2019;6:140.
Suzuki T, Kutsuna S, Taniguchi S, Tajima S, Maeki T, Kato F, et al. Dengue virus exported from Cote d’Ivoire to Japan, June 2017. Emerg Infect Dis. 2017;23:1758–60.
Fofana D, Beugre JMV, Yao-Acapovi GL, Lendzele SS. Risk of dengue transmission in Cocody (Abidjan, Ivory Coast). J Parasitol Res. 2019;2019:4914137.
NEWS DESK. Senegal declares end of dengue epidemic. Outbreak News Today. 2018. http://outbreaknewstoday.com/senegal-declares-end-dengue-epidemic-48679/. Assessed 30 Mar 2020.
Kone AB, Konan YL, Coulibaly ZI, Fofana D, Guindo-Coulibaly N, Diallo M, et al. Entomological evaluation of the risk of urban outbreak of yellow fever in 2008 in Abidjan, Cote d'Ivoire. Med Sante Trop. 2013;23:66–71.
Konan YL, Coulibaly ZI, Allali KB, Tétchi SM, Koné AB, Coulibaly D, et al. Gestion de l’épidémie de fièvre jaune en 2010 à Séguéla (Côte d’Ivoire): intérêt d’une investigation pluridisciplinaire. Santé Publique. 2014;26:859–67.
Zahouli JBZ, Koudou BG, Muller P, Malone D, Tano Y, Utzinger J. Urbanization is a main driver for the larval ecology of Aedes mosquitoes in arbovirus-endemic settings in south-eastern Cote d’Ivoire. PLoS Negl Trop Dis. 2017;11:e0005751.
Adogo LY, Ogoh MO. Review article: yellow fever in Nigeria: a review of the current situation. Afr J Clin Exp Microbiol. 2019;21:1.
Ajogbasile FV, Oguzie JU, Oluniyi PE, Eromon PE, Uwanibe JN, Mehta SB et al. Real-time metagenomic analysis of undiagnosed fever cases unveils a yellow fever outbreak in Edo State, Nigeria. Sci Rep. 2020;10:3180.
World Health Organisation a. Yellow fever—Nigeria. WHO. 2021. https://www.who.int/csr/don/24-november-2020-yellow-fever-nigeria/en/. Assessed 24 Nov 2020.
World Health Organisation b. Yellow fever—Senegal. WHO. 2021. https://www.who.int/csr/don/29-december-2020-yellow-fever-senegal/en/. Assessed 29 Dec 2020.
World Health Organisation c. Yellow fever—Guinea. WHO. 2021. https://www.who.int/csr/don/23-december-2020-yellow-fever-guinea/en/. Assessed 23 Dec 2020.
Stoler J, Delimini RK, Bonney JH, Oduro AR, Owusu-Agyei S, Fobil JN, et al. Evidence of recent dengue exposure among malaria parasite-positive children in three urban centers in Ghana. Am J Trop Med Hyg. 2015;92:497–500.
Amoako N, Duodu S, Dennis FE, Bonney JHK, Asante KP, Ameh J, et al. Detection of dengue virus among children with suspected malaria, Accra. Ghana Emerg Infect Dis. 2018;24:1544–7.
Bonney JHK, Hayashi T, Dadzie S, Agbosu E, Pratt D, Nyarko S, et al. Molecular detection of dengue virus in patients suspected of Ebola virus disease in Ghana. PLoS ONE. 2018;13:e0208907.
Manu SK, Bonney JHK, Pratt D, Abdulai FN, Agbosu EE, Frimpong PO, et al. Arbovirus circulation among febrile patients at the greater Accra Regional Hospital, Ghana. BMC Res Notes. 2019;12:332.
Scott TW, Takken W. Feeding strategies of anthropophilic mosquitoes result in increased risk of pathogen transmission. Trends Parasitol. 2012;28:114–21.
Lounibos LP. Invasions by insect vectors of human disease. Annu Rev Entomol. 2002;47:233–66.
Brown JE, McBride CS, Johnson P, Ritchie S, Paupy C, Bossin H, et al. Worldwide patterns of genetic differentiation imply multiple “domestications” of Aedes aegypti, a major vector of human diseases. Proc Biol Sci. 2011;278:2446–54.
Powell JR, Tabachnick WJ. History of domestication and spread of Aedes aegypti—a review. Mem Inst Oswaldo Cruz. 2013;108:11–7.
Brown JE, Evans BR, Zheng W, Obas V, Barrera-Martinez L, Egizi A, et al. Human impacts have shaped historical and recent evolution in Aedes aegypti, the dengue and yellow fever mosquito. Evolution. 2014;68:514–25.
Mattingly PF. Taxonomy of Aedes aegypti and related species. Bull World Health Organ. 1967;36:552–4.
Huber K, Ba Y, Dia I, Mathiot C, Sall AA, Diallo M. Aedes aegypti in Senegal: genetic diversity and genetic structure of domestic and sylvatic populations. Am J Trop Med Hyg. 2008;79:218–29.
Rose NH, Sylla M, Badolo A, Lutomiah J, Ayala D, Aribodor OB, et al. Climate and urbanization drive mosquito preference for humans. Curr Biol. 2020;30:3570–9.
Weetman D, Kamgang B, Badolo A, Moyes CL, Shearer FM, Coulibaly M, et al. Aedes mosquitoes and Aedes-borne arboviruses in Africa: current and future threats. Int J Environ Res Public Health. 2018;15: 220.
Weaver SC, Reisen WK. Present and future arboviral threats. Antiviral Res. 2010;85:328–45.
Wat’Senga Tezzo F, Fasine S, Manzambi Zola E, Marquetti MDC, Binene Mbuka G, Ilombe G, et al. High Aedes spp. larval indices in Kinshasa, Democratic Republic of Congo. Parasit Vectors. 2021;14:92.
Diallo M, Sall AA, Moncayo AC, Ba Y, Fernandez Z, Ortiz D, et al. Potential role of sylvatic and domestic African mosquito species in dengue emergence. Am J Trop Med Hyg. 2005;73:445–9.
Richards SL, Ponnusamy L, Unnasch TR, Hassan HK, Apperson CS. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in relation to availability of human and domestic animals in suburban landscapes of central North Carolina. J Med Entomol. 2006;43:543–51.
Gyawali N, Taylor-Robinson AW, Bradbury RS, Huggins DW, Hugo LE, Lowry K, et al. Identification of the source of blood meals in mosquitoes collected from north-eastern Australia. Parasit Vectors. 2019;12:198.
Kramer LD, Ciota AT. Dissecting vectorial capacity for mosquito-borne viruses. Curr Opin Virol. 2015;15:112–8.
Schultz GW. Seasonal abundance of dengue vectors in Manila, Republic of the Philippines. Southeast Asian J Trop Med Public Health. 1993;24:369–75.
Barrera R, Amador M, Mackay AJ. Population dynamics of Aedes aegypti and dengue as influenced by weather and human behavior in San Juan, Puerto Rico. PLOS Negl Trop Dis. 2011;5:e1378.
Duncombe J, Espino F, Marollano K, Velazco A, Ritchie SA, Hu W, et al. Characterising the spatial dynamics of sympatric Aedes aegypti and Aedes albopictus populations in the Philippines. Geospat Health. 2013;8:255.
Ngugi HN, Mutuku FM, Ndenga BA, Musunzaji PS, Mbakaya JO, Aswani P, et al. Characterization and productivity profiles of Aedes aegypti (L.) breeding habitats across rural and urban landscapes in western and coastal Kenya. Parasit Vectors. 2017;10:331.
Ibarra AMS, Ryan SJ, Beltran E, Mejia R, Silva M, Munoz A. Dengue vector dynamics (Aedes aegypti) influenced by climate and social factors in Ecuador: implications for targeted control. PLoS ONE. 2013;8:e78263.
Kittayapong P, Strickman D. Distribution of container-inhabiting Aedes larvae (Diptera: Culicidae) at a dengue focus in Thailand. J Med Entomol. 1993;30:601–6.
Wan Norafikah O, Wasi N, Sabtu N, Sai’e S, Heah K, Abdul N, et al. Distribution of Aedes mosquitoes in three selected localities in Malaysia. Sains Malays. 2012;41:1309–13.
Dom NC, Ahmad AH, Ismail R. Habitat characterization of Aedes sp. breeding in urban hotspot area. Procedia Soc Behav Sci. 2013;85:100–9.
Bowman LR, Donegan S, McCall PJ. Is dengue vector control deficient in effectiveness or evidence? Systematic review and meta-analysis. PLOS Negl Trop Dis. 2016;10:e0004551.
Londono-Renteria B, Troupin A, Colpitts TM. Arbovirosis and potential transmission blocking vaccines. Parasit Vectors. 2016;9:516.
Marchi S, Trombetta CM, Montomoli E. Emerging and re-emerging arboviral diseases as a global health problem. Public Health—emerging and re-emerging issues: IntechOpen; 2018. https://www.intechopen.com/chapters/61770.
Achee NL, Grieco JP, Vatandoost H, Seixas G, Pinto J, Ching-Ng L, et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl Trop Dis. 2019;13:e0006822.
Owusu HF, Jancaryova D, Malone D, Muller P. Comparability between insecticide resistance bioassays for mosquito vectors: time to review current methodology. Parasit Vectors. 2015;8:357.
Chadee DD. Resting behaviour of Aedes aegypti in Trinidad: with evidence for the re-introduction of indoor residual spraying (IRS) for dengue control. Parasit Vectors. 2013;6:255.
Vazquez-Prokopec GM, Medina-Barreiro A, Che-Mendoza A, Dzul-Manzanilla F, Correa-Morales F, Guillermo-May G, et al. Deltamethrin resistance in Aedes aegypti results in treatment failure in Merida, Mexico. PLoS Negl Trop Dis. 2017;11(6):e0005656.
Messina JP, Kraemer MU, Brady OJ, Pigott DM, Shearer FM, Weiss DJ, et al. Mapping global environmental suitability for Zika virus. eLife. 2016;5:e15272
Namountougou M, Soma DD, Balboné M, Kaboré DA, Kientega M, Hien A, et al. Monitoring insecticide susceptibility in Aedes Aegypti populations from the two biggest cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: implication of metabolic resistance. Trop Med Infect Dis. 2020;5:84.
Appawu M, Dadzie S, Abdul H, Asmah H, Boakye D, Wilson M, et al. Surveillance of viral haemorrhagic fevers in Ghana: entomological assessment of the risk of transmission in the northern regions. Ghana Med J. 2006;40:137–41.
Badolo A, Sombié A, Pignatelli PM, Sanon A, Yaméogo F, Wangrawa DW, et al. Insecticide resistance levels and mechanisms in Aedes aegypti populations in and around Ouagadougou, Burkina Faso. PLoS Negl Trop Dis. 2019;13:e0007439.
Agha SB, Tchouassi DP, Bastos ADS, Sang R. Assessment of risk of dengue and yellow fever virus transmission in three major Kenyan cities based on Stegomyia indices. PLoS Negl Trop Dis. 2017;11:e0005858.
World Health Organisation. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. WHO. 2016. https://apps.who.int/iris/bitstream/handle/10665/250677/9789241511575-eng.pdf?sequence=1. Assessed 15 Jan 2017.
Huang Y-M. The subgenus Stegomyia of Aedes in the Afrotropical region with keys to the species (Diptera: Culicidae). Zootaxa. 2004;700:1–120.
Vazquez-Prokopec GM, Galvin WA, Kelly R, Kitron U. A new, cost-effective, battery-powered aspirator for adult mosquito collections. J Med Entomol. 2009;46:1256–9.
Ndenga BA, Mutuku FM, Ngugi HN, Mbakaya JO, Aswani P, Musunzaji PS, et al. Characteristics of Aedes aegypti adult mosquitoes in rural and urban areas of western and coastal Kenya. PLoS ONE. 2017;12:e0189971.
Moyes CL, Vontas J, Martins AJ, Ng LC, Koou SY, Dusfour I, et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLOS Negl Trop Dis. 2017;11:e0005625.
Reinhold J, Lazzari C, Lahondère C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: a review. Insects. 2018;9:158.
Li CF, Lim TW, Han LL, Fang R. Rainfall, abundance of Aedes aegypti and dengue infection in Selangor, Malaysia. Southeast Asian J Trop Med Public Health. 1985;16:560–8.
Kamgang B, Ngoagouni C, Manirakiza A, Nakouné E, Paupy C, Kazanji M. Temporal patterns of abundance of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) and mitochondrial DNA analysis of Ae. albopictus in the Central African Republic. PLoS Negl Trop Dis. 2013;7:e2590.
Suzuki T, Osei JH, Sasaki A, Adimazoya M, Appawu M, Boakye D, et al. Risk of transmission of viral haemorrhagic fevers and the insecticide susceptibility status of Aedes aegypti (Linnaeus) in some sites in Accra. Ghana Ghana Med J. 2016;50:136–41.
Manjarres-Suarez A, Olivero-Verbel J. Chemical control of Aedes aegypti: a historical perspective. Rev Costarricense Salud Pública. 2013;22:68–75.
Amelia-Yap ZH, Chen CD, Sofian-Azirun M, Low VL. Pyrethroid resistance in the dengue vector Aedes aegypti in Southeast Asia: present situation and prospects for management. Parasit Vectors. 2018;11:332.
Nawaz S, Tahir HM, Asif Mahmood M, Summer M, Ali S, Ali A, et al. Current status of pyrethroids resistance in Aedes aegypti (Culicidae: Diptera) in Lahore District, Pakistan: a novel mechanistic insight. J Med Entomol. 2021;58:2432–8.
Kamgang B, Yougang AP, Tchoupo M, Riveron JM, Wondji C. Temporal distribution and insecticide resistance profile of two major arbovirus vectors Aedes aegypti and Aedes albopictus in Yaoundé, the capital city of Cameroon. Parasit Vectors. 2017;10:469.
Demok S, Endersby-Harshman N, Vinit R, Timinao L, Robinson LJ, Susapu M, et al. Insecticide resistance status of Aedes aegypti and Aedes albopictus mosquitoes in Papua New Guinea. Parasit Vectors. 2019;12:333
Leal SDV, Fernandes Varela IB, Lopes Gonçalves AAL, Sousa Monteiro DD, Ramos de Sousa CM, Lima Mendonça ML, et al. Abundance and updated distribution of Aedes aegypti (Diptera: Culicidae) in Cabo Verde Archipelago: a neglected threat to Public Health. Int J Environ Res Public Health. 2020;17:1291.
Hanley KA, Monath TP, Weaver SC, Rossi SL, Richman RL, Vasilakis N. Fever versus fever: the role of host and vector susceptibility and interspecific competition in shaping the current and future distributions of the sylvatic cycles of dengue virus and yellow fever virus. Infect Genet Evol. 2013;19:292–311.
Diallo D, Sall AA, Diagne CT, Faye O, Hanley KA, Buenemann M, et al. Patterns of a sylvatic yellow fever virus amplification in southeastern Senegal, 2010. Am J Trop Med Hyg. 2014;90:1003–13.
Dzul-Manzanilla F, Ibarra-López J, Bibiano Marín W, Martini-Jaimes A, Leyva JT, Correa-Morales F, et al. Indoor resting behavior of Aedes aegypti (Diptera: Culicidae) in Acapulco, Mexico. J Med Entomol. 2016;54:501–4.
Acknowledgements
We thank the residents of the study sites for their support during our study. Our sincere gratitude goes to Dr Victor Asoala, Stephen Kantum, and Bernice Ama Baako; all of the staff of the Entomology Department of the Navrongo Health Research Centre, Ghana; Mr Sylvester Coleman of the President Malaria Initiative, Tamale; and Mr Isaac K. Sraku and Prince Ashong-Pappoe of the Medical Microbiology Department, University of Ghana for field and laboratory assistance.
Funding
This study was supported by grants from the National Institute of Health (D43 TW 011513).
Author information
Authors and Affiliations
Contributions
CMO-A, JAAM, DW, and YAA were responsible for the study design, supervised the data collection, and contributed to the writing of the manuscript. CMO-A performed the data collection and analysis, laboratory work, and drafted the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Ethics and Protocol Review Committee of the College of Health Sciences of the University of Ghana (protocol identification number CHS-Et/M.9-P1.5/2017–2018). Meetings were held at each study site with the chiefs and the residents to introduce the research study to the public. All study participants were adults (at least 18 years old). Written and signed consent was obtained from all the adults who volunteered to participate in HLC sampling before they were trained and the sampling of mosquitoes commenced. A copy of their signed consent form was given to each of the HLC volunteers and another copy kept in a locked cabinet with restricted access in the offices of the Department of Medical Microbiology, University of Ghana Medical School. Verbal consent was obtained from household heads to sample mosquitoes in their houses and compounds. Ethical approval for the use of volunteers for mosquito sampling and the protocols involved was sought from the Ethics and Protocol Review Committee of the College of Health Sciences of the University of Ghana (protocol identification number CHS-Et/M.9-P1.5/2017–2018). Permission to carry out the study at the various study sites was sought from community leaders, and verbal consent was sought from the heads of households before houses were entered for inspection and sampling.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Owusu-Asenso, C.M., Mingle, J.A.A., Weetman, D. et al. Spatiotemporal distribution and insecticide resistance status of Aedes aegypti in Ghana. Parasites Vectors 15, 61 (2022). https://doi.org/10.1186/s13071-022-05179-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05179-w