
Abstract
Background
Microsporidia is a large group of eukaryotic obligate intracellular spore-forming parasites, of which 17 species can cause microsporidiosis in humans. Most human-infecting microsporidians belong to the genera Enterocytozoon and Encephalitozoon. To date, only five microsporidian species, including Encephalitozoon-like, have been found in hard ticks (Ixodidae) using microscopic methods, but no sequence data are available for them. Furthermore, no widespread screening for microsporidian-infected ticks based on DNA analysis has been carried out to date. Thus, in this study, we applied a recently developed DNA metabarcoding method for efficient microsporidian DNA identification to assess the role of ticks as potential vectors of microsporidian species causing diseases in humans.
Methods
In total, 1070 (493 juvenile and 577 adult) unfed host-seeking Ixodes ricinus ticks collected at urban parks in the city of Poznan, Poland, and 94 engorged tick females fed on dogs and cats were screened for microsporidian DNA. Microsporidians were detected by PCR amplification and sequencing of the hypervariable V5 region of 18S rRNA gene (18S profiling) using the microsporidian-specific primer set. Tick species were identified morphologically and confirmed by amplification and sequencing of the shortened fragment of cytochrome c oxidase subunit I gene (mini-COI).
Results
All collected ticks were unambiguously assigned to I. ricinus. Potentially zoonotic Encephalitozoon intestinalis was identified in three fed ticks (3.2%) collected from three different dogs. In eight unfed host-seeking ticks (0.8%), including three males (1.1%), two females (0.7%) and three nymphs (0.7%), the new microsporidian sequence representing a species belonging to the genus Endoreticulatus was identified.
Conclusions
The lack of zoonotic microsporidians in host-seeking ticks suggests that I. ricinus is not involved in transmission of human-infecting microsporidians. Moreover, a very low occurrence of the other microsporidian species in both fed and host-seeking ticks implies that mechanisms exist to defend ticks against infection with these parasites.
Graphical abstract

Similar content being viewed by others

Background
Microsporidia is a group of obligate, intracellular spore-forming eukaryotic parasites. The phylum contains more than 1500 described species belonging to at least 200 genera that infect virtually all animal phyla, including protists [1, 2]. To date, 17 species belonging to eight genera and one holding genus (Microsporidium) were found to cause a wide range of clinical human diseases [3]. In immunocompromised patients, microsporidian infections cause various clinical manifestations, ranging from localized to disseminated, depending on the causative species and host immune status [4]. Until the AIDS pandemic, microsporidiosis was rarely recognized in humans; however, the prevalence of microsporidian infections in case of large infection outbreaks or in HIV-positive patients can reach almost 70% [5]. This shows that, as opportunistic parasites, microsporidians can pose a serious threat, especially when they co-occur with another pathogenic agent. The most common microsporidium infecting humans is Enterocytozoon bieneusi, followed by three of four Encephalitozoon species, including E. cuniculi, E. hellem and E. intestinalis [6,7,8]. In addition, spores of E. bieneusi and Encephalitozoon spp. have been detected in feces of companion animals, including dogs, cats and horses (for review see [6]).
Ticks (Ixodida) are obligate hematophagous arthropods that transmit a wide variety of zoonotic pathogens [9,10,11]. Among them, the castor bean tick (Ixodes ricinus) is the major vector of causative agents of viral, bacterial and protozoan zoonotic diseases in Europe [12]. Moreover, urban green areas may act as pathogen exchange hot spots because of increasing populations of potential reservoirs of tick-borne pathogens and the exposure of humans and domesticated animals to the infected ticks [13, 14].
Knowledge about microsporidian infections in ticks is sparse. To date, only five microsporidian species have been found in these hosts (Table 1). Among them, the Encephalitozoon-like species was detected in fed females of Anocentor nitens and Amblyomma cajennense (Ixodidae) [15, 16]. Screening of microsporidians in other hard ticks, i.e., Dermacentor marginatus, D. reticulatus, I. ricinus and I. persulcatus, revealed the presence of three species belonging to the genus Nosema [17,18,19,20,21,22,23,24]. Another species of the genus Nosema was found in the only examined representative of soft ticks, Ornithodoros parkei (Argasidae) [25]. However, all microsporidians found to date in ticks were detected morphologically; none of those species was confirmed by DNA sequencing.
Over the past years, molecular data have proven their usefulness in many fields of biology, including parasitology. Amplification of marker genes in combination with DNA sequencing is widely used for the rapid screening of multiple samples to detect specific infection agents. In addition, major advances in large-scale DNA sequencing have enabled the development of DNA metabarcoding approaches for biodiversity profiling in which different species are detected simultaneously using group-specific or universal PCR primers [26, 27]. Such approaches have already been applied also to the detection of microsporidian biodiversity [28,29,30]. In this article, we used the DNA metabarcoding approach to examine the prevalence and diversity of microsporidians in I. ricinus to assess the role of ticks as potential vectors of microsporidian species causing diseases in humans.
Methods
Tick sampling
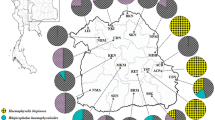
In total, 1164 ticks were tested in this study. The sample of host-seeking ticks included 493 juveniles (30 larvae, 463 nymphs) and 577 adults (291 females, 286 males) that were collected from May to June 2017 and April to May 2018 by dragging and flagging at six recreational parks in the city of Poznan, Poland (Fig. 1 and Additional file 1: Table S1). Dragging was conducted by passing the cotton drag (1 m2) over the ground level vegetation, while flagging was done by waving over (1 m2) and through the higher vegetation (ca. 50–100 cm). Collection sessions were performed between 10:00 am and 7:00 pm. Additionally, 94 engorged females removed from pet animals were provided by veterinary clinics in 2013 and 2016 (Fig. 1; Additional file 2: Table S2).

Sampling places of unfed host-seeking ticks (red circles) and feeding (green circles) ticks in the city of Poznan (blue dot on the Europe map). The size of circle corresponds to the number of analyzed ticks. Numeric value inside the circles: number of microsporidian-positive ticks/number of analyzed ticks. For sampling place characteristics, see Additional file 1: Table S1 and Additional file 2: Table S2
All ticks were preserved in 96% ethanol until DNA extraction. Tick species was initially identified by morphological traits and confirmed by DNA barcoding using cytochrome c oxidase subunit I (COI) amplification and NGS sequencing as described below.
DNA extraction
Engorged ticks were placed separately in 2-ml tubes containing Lysing Matrix A (MP Biomedicals, USA). After the addition of 360 μl of ATL lysis buffer (Qiagen, Germany), samples were homogenized twice for 30 s at 6.0 ms−1 using a FastPrep-24 homogenizer (MP Biomedicals, USA). Proteinase K (Bio Basic, Canada) was added to the samples to the final concentration of 0.2 mg ml−1 and then samples were incubated for 48 h at 56 °C with shaking. Subsequently, 100 μl of the lysate was used to extract total genomic DNA using the DNeasy Blood and Tissue Kit (Qiagen) according to the manufacturer’s instructions.
From unfed host-seeking ticks, total genomic DNA was extracted using ammonium hydroxide method [31]. Individual ticks were crushed using a metal stylus in 10 μl of 0.7 M ammonium hydroxide solution (POCH S.A., Poland) and then the next 90 μl of the solution was added. Samples were incubated for 20 min at 99 °C, and then the tubes were opened and left under the same conditions to concentrate the lysate to about 50 μl. Next, the samples were centrifuged for 5 min at 14.100 ×g, and the supernatant was collected to fresh tubes. DNA extracts were additionally purified from PCR inhibitors using CentriPure Gel Filtration Plates filled with Zoladex (Biotech GmbH, Germany), according to the manufacturer’s protocol.
DNA isolates from adults were analyzed individually, while those from nymphs and larvae were pooled into groups of three individuals. The samples included in the microsporidian-positive pool were re-analyzed separately in a second round of PCR amplification and NGS sequencing. Negative controls from blank DNA extractions and PCR reagents also were amplified and sequenced.
Amplification of mini-COI and 18S rDNA for NGS sequencing
DNA barcode covering 322 bp from the 5′ end of the COI gene (mini-COI) was amplified using bcdF01 (CATTTTCHACTAAYCATAARGATATTGG) [32] and bcdR06 (GGDGGRTAHACAGTYCAHCCNGT) [29] primers tailed at 5′ ends with dual-indexed adapters (forward tail: CCATCTCATCCCTGCGTGTCTCCGACTCAG-index-GAT, reverse tail: CCTCTCTATGGGCAGTCGGTGAT-index) for sequencing using the Ion Torrent system (Life Technologies, USA). PCR amplification was performed in a 5-µl volume containing Hot FIREPol DNA Polymerase (Solis BioDyne, Estonia), 0.25 µM of each tailed primer and 1 µl of template DNA. Amplification program was as follows: 12 min at 95 °C, followed by 35 cycles of 15 s at 95 °C, 30 s at 50 °C and 45 s at 72 °C, with a final extension step at 72 °C for 5 min.
The hypervariable V5 region of 18S rRNA gene (18S rDNA) was amplified in two technical replications using the microsporidian-specific primer set CM-V5F (GATTAGANACCNNNGTAGTTC) and CM-V5R (TAANCAGCACAMTCCACTC) [29]. PCR reactions were done in a total volume of 10 µl containing Hot FIREPol DNA Polymerase, 0.25 µM of each tailed primer and 1 µl of template DNA. Amplification program was as follows: 12 min at 95 °C, followed by 40 cycles of 15 s at 95 °C, 30 s at 50 °C and 30 s at 72 °C, with a final extension step at 72 °C for 5 min.
Library construction and NGS sequencing
The 18S rDNA and mini-COI libraries were prepared separately. For each PCR reaction, 3 µl was electrophoresed on a 2% agarose gel to check amplification efficiency. Then, the amplicons were pooled and purified using 2% E-Gel SizeSelect II Agarose Gels system (Invitrogen, USA), according to the manufacturer’s protocol. DNA concentration and fragment length distribution of the libraries were established using High Sensitivity D1000 Screen Tape assay on 2200 Tape Station system (Agilent Technologies, USA). For the emulsion PCR, the 18S rDNA and mini-COI libraries were pooled in the 10:1 ratio. Emulsion PCR and sequencing were carried out using the Ion Torrent One Touch System II, the Ion 520 and Ion 530 Kit-OT2, Ion 530 chip and the S5 system (Life Technologies) according to the manufacturer's instructions.
Read processing and data analysis
Raw sequence data were pre-filtered by Ion Torrent Suite software version 5.10.1 (Life Technologies) to remove polyclonal and low-quality sequences. Further bioinformatic analysis was conducted using fastq data and custom workflow. Sequence reads < 180-bp were removed from the dataset. Leading and trailing low-quality bases were removed using Trimmomatic version 0.39 [33]. Fastx-Toolkit [34] was used to extract sequences with the minimum of 50% of bases with a quality score ≥ 25. Quality filtered sequences were separated into individual combinations of indexes in Geneious R11.1.5. Chimeras were removed using the default settings in UCHIME2 version 4.2.40 [35]. Next, the sequences were trimmed at 5′ and 3′ ends to exclude PCR primers.
Operational taxonomic unit (OTU) clustering was done in USEARCH version 11.0.667 [36]. Singletons (< 10 reads) were removed, and then OTUs were clustered from the sequences whose abundance exceeded a threshold of ten counts using the cluster_otus algorithm [37]. The OTU consensus sequences were compared to GenBank (www.ncbi.nlm.nih.gov) using BLASTN [38] optimized for highly similar sequences (megablast algorithm) [39]. The OTU consensus sequences were compared to GenBank using 97% identity threshold to determine tick species and 100% identities for identification of microsporidian species.
Amplification and Sanger sequencing of the large fragment of 18S rDNA
A large fragment of the 18S rRNA gene, covering V1-V5 hypervariable regions, was amplified using V1F (CACCAGGTTGATTCTGCCTGAC) [40] and CM-V5R (TAANCAGCACAMTCCACTC) [29] primers. PCR reactions were prepared in two technical replicates, each in a total volume of 10 µl containing Hot FIREPol DNA Polymerase, 0.25 µM of each primer and 1 µl of template DNA. Amplification program was as follows: 12 min at 95 °C, followed by 35 cycles of 15 s at 95 °C, 1 min at 60 °C and 1 min at 72 °C, with a final extension step at 72 °C for 10 min. After amplification, technical replications were pooled, and 5 μl was analyzed by electrophoresis on 1.5% agarose gel stained with GelRed (Biotium, USA). Samples containing visible bands were purified with Escherichia coli exonuclease I and FastAP Alkaline Phosphatase (Thermo Scientific, USA) and sequenced using BigDye Terminator v3.1 Cycle Sequencing Kit and ABI Prism 3130XL Genetic Analyzer (Applied Biosystems, USA), following the manufacturer’s instructions. Sequence chromatograms were checked for accuracy in Geneious R11.1.5 (Biomatters Ltd.).
Phylogenetic analysis
For phylogenetic analysis, we used all published 18S rRNA gene sequences assigned to the genus Endoreticulatus. According to Trzebny et al. [29], the genus Cystosporogenes was used as a close outgroup and a chytrid Chytridiopsis typographi (Chytridiopsida) was used to root the tree (Table 2). Sequences were aligned using the L–INS–i algorithm in MAFFT version 7.388 [58, 59] as implemented in Geneious R11.1.5. The final alignment consisted of 1436 nucleotide positions. The best fit model of DNA evolution (GTR + I + G) was chosen by PartitionFinder2 [60]. Phylogenetic trees were reconstructed using maximum likelihood in Garli version 2.0 [61] and Bayesian inference in MrBayes version 3.2.6 [62]. Each BI run of four independent chains was performed in 2 × 10,000,000 generations, and the trees were sampled every 1000 generations. The final consensus tree was generated after discarding the burn-in fraction of 25% of initial trees; the average standard deviation of split frequencies dropped below 0.003. Bootstrap support for the ML tree was calculated by using 10,000 data replicates as implemented in Garli. The tree was edited in FigTree version 1.4.4 [63] and further in Corel Draw X4.
Statistical analysis
The chi-square test with Yates’ correction [64, 65] was used for statistical comparison between the microsporidian prevalence in feeding ticks and the ticks’ host as well as tick collection areas. The level of statistical significance was established at P < 0.05.
The statistical significances between microsporidian infection and developmental stage, year and collection area of unfed host-seeking ticks were analyzed using the chi-square test with Yates’ correction [64, 65], and differences were considered significant if P < 0.05.
Results
Based on mini-COI sequence data, all analyzed ticks were unambiguously assigned to I. ricinus. The representative sequences are available in GenBank under accession nos. MZ970304–MZ970324.
Three feeding females (3.2%) collected from different dogs out for walks in the Rusalka area (Fig. 1; Additional file 2: Table S2) were positive for microsporidian 18S rDNA and shared the same DNA sequence (MT911430–MT911432), which was identical to sequences representing E. intestinalis found in humans (e.g. KM058742). However, the correlations between the microsporidian-positive ticks and both the tick host (χ2Yest = 0.156, df = 1, P = 0.693) and tick collection areas (χ2Yest = 0.443, df = 2, P = 0.801) were not statistically significant.
Microsporidian DNA was also detected in eight unfed host-seeking ticks (0.8%), including three males (1.1%), two females (0.7%) and three nymphs (0.7%) (Fig. 1; Additional file 1: Table S1). In this case also there was no correlation between tick developmental stage and microsporidian infections (χ2Yest = 0.452, df = 3, PYest = 0.929). All microsporidian-positive samples shared the same 18S rDNA sequence (MT911422–MT911429) most similar to the group of sequences which originated from Endoreticulatus spp. (e.g. KY615713, LC467310, JN792450, AY009115). Phylogenetic analysis based on the longer fragment of the 18S rRNA gene (Fig. 2) revealed that the new sequence, named Endoreticulatus sp. PL02, is sister to the sequence derived from Endoreticulatus sp. found in Eilema complana (Lepidoptera) collected in Bulgaria [51].

Phylogenetic relationships of Endoreticulatus 18S rDNA sequences inferred from Bayesian inference and maximum likelihood analyses. Values near branches show Bayesian posterior probabilities (PP) and bootstrap support values (BS) (PP/BS). Black circles: maximally supported; empty circles: supported > 0.95 PP and > 75% BS. The sequence found in this study is in bold. The uncultured human pathogenic microsporidium (JN619406.1) is in red
None of the analyzed host-seeking ticks (n = 1070) were positive for any microsporidian species causing human microsporidiosis.
Endoreticulatus sp. PL02 was detected mainly in ticks collected near the Rusalka Lake: in three males and two females in 2017 and in two nymphs in 2018, while one infected nymph was identified in Morasko area in 2017. However, no correlation was found between Endoreticulatus sp. PL02 infection and the year of tick collection (χ2Yest = 0.221, df = 1, PYest = 0.638) and the study area (χ2Yest = 4.02, df = 5, PYest = 0.543).
Discussion
Our data suggest that I. ricinus is not involved in transmission of zoonotic microsporidian species, because none of the analyzed host-seeking ticks was positive for any human pathogenic species. Encephalitozoon intestinalis was detected only in feeding ticks removed from dogs. There are numerous reports about the occurrence of human pathogenic microsporidians in dogs [66,67,68,69]. Similarly, Encephalitozoon-like microsporidians were found in Amblyomma and Anocentor ticks that had been collected from horses and then fed on rabbits [15, 16]. These parasites were observed in the midgut epithelial cells and acinus of the salivary gland in different phases of the host’s development, which suggests that this microsporidium may infect ticks. The authors suggested that the rabbit’s or horse’s blood meals were the source of infection [15, 16]. Presumably, in our study, the presence of E. intestinalis in engorged ticks resulted from their earlier feeding on dogs infected by microsporidians. Moreover, the lack of zoonotic microsporidians in unfed host-seeking nymphs and adults suggests that E. intestinalis is not transmitted transstadially.
Results of the phylogenetic analysis showed that Endoreticulatus sp. PL02 forms a clade with Endoreticulatus sp. recorded in an E. complana moth [51]. Both sequences were nested in the strongly supported clade grouping Endoreticulatus sequences found in insects (Fig. 2 and Table 2). In addition, our phylogenetic tree suggests that two sequences previously described as closely related to the Endoreticulatus, the Microsporidium sp. clones Chula Myositis 1 and Chula Keratitis 1, which caused microsporidian myositis or stromal keratitis in humans, probably represent a different genus, so far unknown from the complete 18S rRNA gene (Fig. 2) [70, 71].
Our data show that both the diversity and prevalence of microsporidians in I. ricinus are very low. Among unfed host-seeking ticks we noticed only one type of microsporidian 18S rDNA sequence. Considering that we detected Endoreticulatus sp. PL02 based on molecular analyses, the comparison with N. slovaca [20, 23], Unikaryon (Nosema) ixodis [17, 24] and Nosema-like [22] previously noticed in I. ricinus is unfeasible. Thus, further morphological and ultramicroscopy studies are needed to confirm that the Endoreticulatus sp. PL02 sequence found in this study represents a different species not yet recorded in ticks.
Low prevalence of microsporidians in I. ricinus found in this study corresponds well with the previous reports (Table 1). In the most extensive survey on the prevalence of microsporidians in ticks [18], none of about 6200 I. ricinus individuals were positive for these parasites, and only N. slovaca was found in 0.2% of about 3000 D. reticulatus ticks. Similar low prevalence was found in D. reticulatus (0.1%) and I. ricinus (0.5%) sampled in Hungary [20] and in I. persulcatus (1.6%) collected in the vicinity of St. Petersburg [21]. Additionally, the intensity of infection in I. persulcatus from St. Petersburg was very low and amounted to one spore per smear [21].
The presence of Endoreticulatus sp. PL02 in unfed host-seeking adults and nymphs suggests vertical transovarial transmission of this microsporidian species. Horizontal transmission is most frequently reported in Microsporidia [72,73,74,75]. Microsporidians transmitted vertically are typically less virulent than horizontally transmitted species and show higher specificity to the host. However, mortality of early developmental stages of hosts who acquired the parasite as a result of transovarial transmission is generally higher than for infections caused by species acquired during the host life span [60,61,62]. This mortality results from destruction of various host tissues and subsequent depletion of essential energy reserves necessary for pupation [72, 76,77,78]. Previous studies have shown that Endoreticulatus usually is transmitted horizontally and infects the midgut only; however, horizontal transmission also can occur in this genus [43, 76,77,78]. In Lymantria dispar (Lepidoptera), for example, it has been shown that E. schubergi was transmitted to offspring from females and males via the egg chorion [78]. Although our results suggest that Endoreticulatus sp. PL02 might spread vertically, the determination of the exact pathway of transovarial transmission of this species and its potential impact on I. ricinus nymph and larval mortality requires further detailed studies.
Ticks are exposed to microsporidians that infect the hosts they feed on. Consequently, these parasites also can infect the epithelial cells of the ticks’ midgut [15, 16]. However, in our study, the microsporidian species known to infect vertebrates were not observed in unfed host-seeking juvenile and adult ticks. Therefore, ticks must have evolved some mechanisms to overcome this infection. The low prevalence of microsporidians in ticks also suggests that ticks developed some unknown defense mechanisms against microsporidian infection. Some of them may involve the immune defense pathway to maintain the pathogens at the level, which does not impair their fitness and development [79,80,81,82]. The pro‐phenoloxidase (proPO) cascade has an important role in melanization reaction, which is the first immune response against pathogens [83, 84]. Cell-surface proteins of pathogens activate the conversion of proPO into active phenoloxidase (PO) [84]. Phenoloxidase activity is used to determine the host’s immune defense ability; higher PO activity levels mean higher resistance to pathogens [85,86,87,88,89]. Sokolova et al. [90] showed that during the infection of Gryllus bimaculatus by Paranosema grylli, the activity of PO expressed as the number of positively stained hemocytes was suppressed from 40–50% to 10–20%. Nevertheless, N. grylli does not suppress cellular reactions such as clamp formation and phagocytosis of liberated spores. In addition, Tokarev et al. [91] noticed that in Locusta migratoria and G. bimaculatus infected by P. locustae and P. grylli, respectively, microsporidians produce enlarged or malformed spores. These types of spore modifications were a result of abnormal sporogony and were noticed in the melanized sites, which indicates that melanization contributes to the abnormal development of microsporidians. Invertebrate immune systems are very heterogeneous [92]; therefore, assessment of how microsporidians influence the cellular immune response of ticks should be tested. These studies should also consider other factors, including tick gut microbiome composition and other pathogens that may co-occur with microsporidians.
Conclusions
Our data suggest that I. ricinus is not involved in transmission of zoonotic microsporidian species. Moreover, we noticed a very low prevalence of these parasites in the tested ticks. Although we found E. intestinalis in engorged tick females, which indicates that ticks can acquire zoonotic microsporidians with their food, none of unfed tested ticks were positive for this microsporidian DNA. Moreover, a very low occurrence of the other microsporidian species in both fed and host-seeking ticks implies that mechanisms exist to defend ticks against infection with these parasites. The only microsporidian species found in unfed host-seeking ticks, Endoreticulatus sp. PL02, probably use I. ricinus as a specific host. Further studies are needed to determine the host specificity of Endoreticulatus sp. PL02 and potential defense mechanisms developed by ticks against microsporidian parasites.
Availability of data and materials
Sequences generated in this study are available in GenBank under accession nos. MT911422–MT911432 and MZ970304–MZ970324. Additional details are available in Additional file 1: Table S1 and Additional file 2: Table S2. Sequence alignments and the remaining data that support the findings of this study are available from the corresponding author upon request.
Abbreviations
- 18S rDNA:
-
DNA coding small subunit ribosomal RNA gene
- BS:
-
Bootstrap support values
- COI:
-
Cytochrome c oxidase subunit I gene
- CP:
-
Cytadela park
- ML:
-
Area around Malta Lake
- MO:
-
Morasko
- OTU:
-
Operational taxonomic unit
- PO:
-
Phenoloxidase
- PP:
-
Bayesian posterior probabilities
- proPO:
-
Pro‐phenoloxidase
- RL:
-
Area around Rusalka Lake
- SP:
-
Solacki park
- TP:
-
Tysiaclecia park
References
Vávra J, Lukeš J. Microsporidia and ‘the art of living together.’ In: Rollinson D, editor. Adv parasitol. Cambridge: Academic Press; 2013. p. 253–319.
Becnel JJ, Takvorian PM, Cali A. Checklist of available generic names for microsporidia with type species and type hosts. In: Weiss LM, Becnel JJ, editors. Microsporidia. Chichester: Wiley; 2014. p. 671–86.
Han B, Weiss LM. Microsporidia: obligate intracellular pathogens within the fungal kingdom. Microbiol Spectr. 2017. https://doi.org/10.1128/microbiolspec.FUNK-0018-2016.
Didier ES, Weiss LM. Microsporidiosis: not just in AIDS patients. Curr Opin Infect Dis. 2011;24:490–5.
Nkinin SW, Asonganyi T, Didier ES, Kaneshiro ES. Microsporidian infection is prevalent in healthy people in Cameroon. J Clin Microbiol. 2007;45:2841–6.
Fayer R, Santin-Duran M. Epidemiology of microsporidia in human infections. In: Weiss LM, Becnel JJ, editors. Microsporidia pathogen opportunity. 1st ed. Hoboken: Wiley; 2014. p. 135–64.
Franzen C, Müller A. Microsporidiosis: human diseases and diagnosis. Microbes Infect. 2001;3:389–400.
Stentiford GD, Becnel JJ, Weiss LM, Keeling PJ, Didier ES, Williams BAP, et al. Microsporidia—emergent pathogens in the global food chain. Trends Parasitol. 2016;32:336–48.
Gern L, Rouvinez E, Toutoungi LN, Godfroid E. Transmission cycles of Borrelia burgdorferi sensu lato involving Ixodes ricinus and/or I. hexagonus ticks and the European hedgehog, Erinaceus europaeus, in suburban and urban areas in Switzerland. Folia Parasitol (Praha). 1977;44:309–14.
Pfäffle M, Littwin N, Muders SV, Petney TN. The ecology of tick-borne diseases. Int J Parasitol. 2013;43:1059–77.
Millán J, Proboste T, de Fernández Mera IG, Chirife AD, de Fuente la J, Altet L. Molecular detection of vector-borne pathogens in wild and domestic carnivores and their ticks at the human-wildlife interface. Ticks Tick Borne Dis. 2016;7:284–90.
Földvári G. Life cycle and ecology of Ixodes ricinus: the roots of public health importance. In: Braks MAH, van Wieren SE, Takken W, Sprong H, editors. Ecology and prevention of Lyme borreliosis. Wageningen: Wageningen Academic Publishers; 2016. p. 31–40.
Rizzoli A, Silaghi C, Obiegala A, Rudolf I, Hubálek Z, Földvári G, et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: new hazards and relevance for public health. Front Public Health. 2014;2:1–26.
Oechslin CP, Heutschi D, Lenz N, Tischhauser W, Péter O, Rais O, et al. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasit Vectors. 2017. https://doi.org/10.1186/s13071-017-2500-2.
Ribeiro MFB, Guimarães AM. Encephalitozoon-like microsporidia in the ticks Amblyomma cajennense and Anocentor nitens (Acari: Ixodidae). J Med Entomol. 1998;35:1029–33.
Ribeiro MFB, Passos LMF. Natural co-infection of Babesia caballi and Encephalitozoon-like microsporidia in the tick Anocentor nitens (Acari: Ixodidae). J Invertebr Pathol. 2006;93:183–5.
Weiser J, Rehacek J, Zizka Z, Ciampor F, Kocianova E. Nosema slovaca Weiser et Rehacek, 1975 and Unikaryon ixodis (Weiser, 1957) comb. n. in ixodid ticks. Acta Parasitol. 1999;44:99–107.
Rehácek J, Weiser J. Natural infection of the tick Dermacentor reticulatus (Fabr.) with the microsporidian Nosema slovaca Weiser et Rehácek in Slovakia. Folia Parasitol (Praha). 1978;25:165–70.
Weiser J, Rehacek J. A microsporidian from Dermacentor reticulatus. II Inernationales Arbeitskolloquium uber Naturherde von Infekt Zentraleuropa. 1976;487–9.
Rehacek J, Kovacova E, Kocianova E. Isolation of Nosema slovaca (Microsporidiae) from Dermacentor reticulatus ticks (Acari: Ixodidae) collected in Hungary. Exp Appl Acarol. 1996;20:57–60.
Tokarev YS, Dubinina H, Alekseev A. Microsporidian infection in a natural population of Ixodes persulcatus schulze (Acarina: Ixodidae) in North-Western Russia. Bul Acad Stiint Mold. 2007;302:66.
Tokarev YS, Movile AA. A first record of microsporidia in the ixodid tick Ixodes ricinus L. (Ixodidae) in the territory of the CIS, Republic Moldova. Parazitologiia. 2004;1970:388–405.
Weiser J, Rehacek J. Nosema slovaca sp. n.: a second microsporidian of the tick Ixodes ricinus. J Invertebr Pathol. 1975;26:411.
Weiser J. Parasites of some blood sucking arthropods. Českoslov Parasitol. 1957;4:355–8.
Krinsky WL. Nosema parkei sp. n., a Microsporidian from the Argasid Tick, Ornithodoros parkei Cooley. J Protozool. 1970;24:52–6.
Weiss LM, Vossbrinck CR. Microsporidiosis: molecular and diagnostic aspects. Adv Parasitol. 1998;40:351–95.
Shehab AY, Moneer EA, Allam AF, Khalil SS, Tolba MM. Intestinal Microsporidia infection in leukemic children: microscopic and molecular detection. Acta Parasitol. 2021;66:346–53.
Williams BAP, Hamilton KM, Jones MD, Bass D. Group-specific environmental sequencing reveals high levels of ecological heterogeneity across the microsporidian radiation. Environ Microbiol Rep. 2018;10:328–36.
Trzebny A, Slodkowicz-Kowalska A, Becnel JJ, Sanscrainte N, Dabert M. A new method of metabarcoding Microsporidia and their hosts reveals high levels of microsporidian infections in mosquitoes (Culicidae). Mol Ecol Resour. 2020;20:1486–504.
Dubuffet A, Chauvet M, Moné A, Debroas D, Lepère C. A phylogenetic framework to investigate the microsporidian communities through metabarcoding and its application to lake ecosystems. Environ Microbiol. 2021;23:4344–59.
Rijpkema S, Bruinink H. Detection of Borrelia burgdorferi sensu lato by PCR in questing Ixodes ricinus larvae from the Dutch North Sea island of Ameland. Exp Appl Acarol. 1996;20:381–5.
Dabert M, Witalinski W, Kazmierski A, Olszanowski Z, Dabert J. Molecular phylogeny of acariform mites (Acari, Arachnida): strong conflict between phylogenetic signal and long-branch attraction artifacts. Mol Phylogenet Evol. 2010;56:222–41.
Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–20.
Hannon GJ. FASTX-Toolkit. 2010. http://hannonlab.cshl.edu. Accessed Aug 2020.
Edgar RC. UCHIME2: improved chimera prediction for amplicon sequencing. bioRxiv. 2016. https://doi.org/10.1101/074252.
Edgar RC. Search and clustering orders of magnitude faster than BLAST. Bioinformatics. 2010;26:2460–1.
Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods. 2013;10:996–8.
Zhang Z, Schwartz S, Wagner L, Miller W. A greedy algorithm for aligning DNA sequences. J Comput Biol. 2000;7:203–14.
Morgulis A, Coulouris G, Raytselis Y, Madden TL, Agarwala R, Schäffer AA. Database indexing for production MegaBLAST searches. Bioinformatics. 2008;24:1757–64.
Zhu X, Wittner M, Tanowitz HB, Kotler D, Cali A, Weiss LM, et al. Small subunit rRNA sequence of Enterocytozoon bieneusi and its potential diagnostic role with use of the polymerase chain reaction. J Infect Dis. 1993;168:1570–5.
Zhang L, Du Y, Wang YJ. A new species of Endoreticulatus (Protozoa, Microspore) from the larva of silkworm, Bombyx mori L. J Southwest Agric Univ. 1995;17:363–7.
Malone LA. A new pathogen, Microsporidium itiiti n. sp. (Microsporidia) from the Argentine stem weevil, Listronotis bonariensis (Coleoptera: Curculionidae). J Protozool. 1985;32:531–5.
Pilarska DK, Radek R, Huang W-F, Takov DI, Linde A, Solter LF. Review of the genus Endoreticulatus (Microsporidia, Encephalitozoonidae) with description of a new species isolated from the grasshopper Poecilimon thoracicus (Orthoptera: Tettigoniidae) and transfer of Microsporidium itiiti Malone to the genus. J Invertebr Pathol. 2015;124:23–30.
Zwölfer W. Die Pebrie des Schwammspinner und Goldafters, eine neue wirtschaftlich bedeutungsvolle Infektionskrankheit. Zeitschrift Für Angew Entomol. 1927;12:498–500.
Cali A, El Gary M. Ultrastructural study of the development of Pleistophora schubergi Zwölfer, 1927 (Protozoa, Microsporida) in larvae of the spruce budworm, Choristoneura fumiferana and its subsequent taxonomic change to the genus Endoreticulatus. J Protozool. 1991;38:271–8.
Wang CY, Solter LF, T’sui WH, Wang CH. An Endoreticulatus species from Ocinara lida (Lepidoptera: Bombycidae) in Taiwan. J Invertebr Pathol. 2005;89:123–35.
Hoch G, Verucchi S, Schopf A. Microsporidian pathogens of the oak processionary moth, Thaumetopoea processionea (L.) (Lep., Thaumetopoeidae), in eastern Austria’s oak forests. Mitt Dtsch Ges Allg Angew Ent. 2008;16:225–8.
Malysh JM, Kononchuk AG, Frolov AN. Detection of microsporidia infecting beet webworm Loxostege sticticalis (Pyraloidea: Crambidae) in European part of Russia in 2006–2008. Plant Prot News. 2019. https://doi.org/10.31993/2308-6459-2019-2(100)-45-51.
Qiu H, Lu X, Li M, He X, He X. Phylogenetic analysis of complete rRNA gene sequence of Endoreticulatus sp. Shengzhou from the silkworm Bombyx mori in Zhejiang of China. J Anim Vet Adv. 2012;11:1056–62.
Xu X, Shen Z, Zhu F, Tao H, Tang X, Xu L. Phylogenetic characterization of a microsporidium (Endoreticulatus sp. Zhenjiang) isolated from the silkworm, bombyx mori. Parasitol Res. 2012;110:815–9.
Pilarska D, Takov D, Hyliš M, Radek R, Fiala I, Solter L, et al. Natural occurrence of microsporidia infecting Lepidoptera in Bulgaria. Acta Parasitol. 2017;62:858–69.
Pilarska D, Linde A, Solter L, McManus M, Takov D. New data on the biology of the microsporidium Endoreticulatus schubergi infecting the browntail moth Euproctis chrysorhoea (Lepidopter: Lymantriidae). Acta Zool Bulg. 2002;54:55–62.
Kleespies RG, Vossbrinck CR, Lange M, Jehle JA. Morphological and molecular investigations of a microsporidium infecting the European grape vine moth, lobesia botrana den. et Schiff., and its taxonomic determination as Cystosporogenes legeri nov. comb. J Invertebr Pathol. 2003;83:240–8.
Canning EU, Curry A, Cheney SA, Lafranchi-Tristem NJ, Ebert D, Rifardt D, et al. Flabelliforma montana (Phylum Microsporidia) from Phlebotomus ariasi (Diptera, Psychodidae): ultrastructural observations and phylogenetic relationships. Eur J Protistol. 2001;37:207–21.
Kyei-Poku G, Gauthier D, Schwarz R, van Frankenhuyzen K. Morphology, molecular characteristics and prevalence of a Cystosporogenes species (Microsporidia) isolated from Agrilus anxius (Coleoptera: Buprestidae). J Invertebr Pathol. 2011;107:1–10.
Solter LF, Pilarska DK, McManus ML, Zúbrik M, Patočka J, Huang WF, et al. Host specificity of microsporidia pathogenic to the gypsy moth, Lymantria dispar (L.): field studies in Slovakia. J Invertebr Pathol. 2010;105:1–10.
Corsaro D, Wylezich C, Venditti D, Michel R, Walochnik J, Wegensteiner R. Filling gaps in the microsporidian tree: rDNA phylogeny of Chytridiopsis typographi (Microsporidia: Chytridiopsida). Parasitol Res. 2019;118:169–80.
Katoh K, Misawa K, Kuma K, Miyata T. MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002;30:3059–66.
Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:772–80.
Lanfear R, Calcott B, Ho SYW, Guindon S. PartitionFinder: combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol Biol Evol. 2012;29:1695–701.
Zwickl DM. Genetic algorithm approaches for the phylogenetic analysis of large biological sequence datasets under the maximum likelihood criterion. Austin: The University of Texas at Austin; 2006.
Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Höhna S, et al. Mrbayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst Biol. 2012;61:539–42.
Rambaut A. FigTree v. 1.4.4. 2018. https://github.com/rambaut/figtree. Accessed Aug 2020.
Pearson K. On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Philos Mag Ser. 1900;5(50):157–75.
Yates F. Contingency tables involving small numbers and the χ 2 test. Suppl J R Stat Soc. 1934;1:217.
Duzlu O, Yildirim A, Onder Z, Ciloglu A, Yetismis G, Inci A. Prevalence and genotyping of microsporidian parasites in dogs in Turkey: zoonotic concerns. J Eukaryot Microbiol. 2019;66:771–7.
Piekarska J, Kicia M, Wesołowska M, Kopacz Ż, Gorczykowski M, Szczepankiewicz B, et al. Zoonotic microsporidia in dogs and cats in Poland. Vet Parasitol. 2017;246:108–11.
Piekarska J, Kicia M, Wesolowska M, Kopacz Z, Gorczykowski M, Sobieraj B, et al. Human-pathogenic microsporidia in household dogs and cats in Wroclaw (Poland). Ann Parasitol. 2016;62:2016.
Xu H, Jin Y, Wu W, Li P, Wang L, Li N, et al. Genotypes of Cryptosporidium spp., Enterocytozoon bieneusi and Giardia duodenalis in dogs and cats in Shanghai, China. Parasit Vectors. 2016;9:1–9.
Suankratay C, Thiansukhon E, Nilaratanakul V, Putaporntip C, Jongwutiwes S. Disseminated infection caused by novel species of Microsporidium, Thailand. Emerg Infect Dis. 2012;18:302–4.
Pariyakanok L, Satitpitakul V, Putaporntip C, Jongwutiwes S. Femtosecond laser-assisted anterior lamellar keratoplasty in stromal keratitis caused by an Endoreticulatus-like microsporidia. Cornea. 2015;34:588–91.
Solter LF. Epizootiology of Microsporidiosis in invertebrate hosts. In: Weiss LM, Becnel JJ, editors. Microsporidia pathogen opportunity. First Edition. Hoboken: Wiley; 2014. p. 165–94.
Andreadis TG. Microsporidian parasites of mosquitoes. J Am Mosq Control Assoc. 2007;23:3–29.
Dunn AM, Smith JE. Microsporidian life cycles and diversity: the relationship between virulence and transmission. Microbes Infect. 2001;3:381–8.
Dunn AM, Terry RS, Smith JE. Transovarial transmission in the Microsporidia. Adv Parasitol. 2001;48:57–100.
Pilarska DK, Solter LF, Kereselidze M, Linde A, Hoch G. Microsporidian infections in Lymantria dispar larvae: interactions and effects of multiple species infections on pathogen horizontal transmission. J Invertebr Pathol. 2006;93:105–13.
Goertz D, Hoch G. Horizontal transmission pathways of terrestrial microsporidia: a quantitative comparison of three pathogens infecting different organs in Lymantria dispar L. (Lep.: Lymantriidae) larvae. Biol Control. 2008;44:196–206.
Goertz D, Hoch G. Vertical transmission and overwintering of microsporidia in the gypsy moth, Lymantria dispar. J Invertebr Pathol. 2008;99:43–8.
Hajdušek O, Šíma R, Ayllón N, Jalovecká M, Perner J, de la Fuente J, et al. Interaction of the tick immune system with transmitted pathogens. Front Cell Infect Microbiol. 2013;4:1–15.
Gulia-Nuss M, Nuss AB, Meyer JM, Sonenshine DE, Roe RM, Waterhouse RM, et al. Genomic insights into the Ixodes scapularis tick vector of Lyme disease. Nat Commun. 2016;7:10507.
Smith AA, Pal U. Immunity-related genes in Ixodes scapularis-perspectives from genome information. Front Cell Infect Microbiol. 2014;4:1–12.
Kotsyfakis M, Schwarz A, Erhart J, Ribeiro JMC. Tissue- and time-dependent transcription in Ixodes ricinus salivary glands and midguts when blood feeding on the vertebrate host. Sci Rep. 2015;5:1–10.
Gillespie JP, Kanost MR, Trenczek T. Biological mediators of insect immunity. Annu Rev Entomol. 1997;23:159–66.
Soderhall K, Cerenius L. Role of the prophenoloxidase-activating system in invertebrate immunity. Curr Opin Immunol. 1998;10:23–8.
Cotter SC, Wilson K. Heritability of immune function in the caterpillar Spodoptera littoralis. Heredity (Edinb). 2002;88:229–34.
Nigam Y, Maudlin I, Welburn S, Ratcliffe NA. Detection of phenoloxidase activity in the Hemolymph of Tsetse flies, refractory and susceptible to infection with Trypanosoma brucei rhodesiense. J Invertebr Pathol. 1997;69:279–81.
Reeson AF, Wilson K, Gunn A, Hails RS, Goulson D. Baculovirus resistance in the noctuid Spodoptera exempta is phenotypically plastic and responds to population density. Proc R Soc B Biol Sci. 1998;265:1787–91.
Hagen HE, Grunewald J, Ham PJ. Induction of the prophenoloxidase-activating system of Simulium (Diptera: Simuliidae) following Onchocerca (Nematoda: Filarioidea) infection. Parasitology. 1994;109:649–55.
Gomes SAO, Feder D, Thomas NES, Garcia ES, Azambuja P. Rhodnius prolixus infected with Trypanosoma rangeli in vivo and in vitro experiments. J Invertebr Pathol. 1999;73:289–93.
Sokolova JY, Tokarev YS, Lozinskaia YL, Glupov VV. A morphofunctional analysis of the hemocytes in the cricket Gryllus bimaculatus (Orthoptera: Gryllidae) normally and in acute microsporidiosis due to Nosema grylli. Parazitologiia. 2000;34:408–19.
Tokarev YS, Sokolova YY, Entzeroth R. Microsporidia-insect host interactions: teratoid sporogony at the sites of host tissue melanization. J Invertebr Pathol. 2007;94:70–3.
Loker ES, Adema CM, Zhang SM, Kepler TB. Invertebrate immune systems – not homogeneous, not simple, not well understood. Immunol Rev. 2004;198:10–24.
Acknowledgements
We thank Urszula Sobczynska, Lukasz Broda, Weronika Pawlik and Aneta Lys, Adam Mickiewicz University in Poznan, for their assistance in collecting ticks for this study. This research was supported in part by PLGrid Infrastructure.
Funding
This study was supported by grant no. 2020/37/N/NZ8/01735 from the National Science Centre of Poland and by the project Passport to the Future—Interdisciplinary doctoral studies at the Faculty of Biology, AMU, POWR.03.02.00-00-I006/17. The authors received financial support to cover the costs of Open Access from the Initiative of Excellence-Research University (05/IDUB/2019/94) at Adam Mickiewicz University, Poznan, Poland.
Author information
Authors and Affiliations
Contributions
All authors designed the research; JL provided and morphologically analyzed ticks; AT performed sequencing and data analysis; AT, MD co-wrote the first draft of the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Characteristics of the questing ticks analyzed in this study (see Fig. 1 in the text).
Additional file 2: Table S2.
Characteristics of feeding ticks analyzed in this study (see Fig. 1 in the text).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Trzebny, A., Liberska, J., Slodkowicz-Kowalska, A. et al. Metabarcoding reveals low prevalence of microsporidian infections in castor bean tick (Ixodes ricinus). Parasites Vectors 15, 26 (2022). https://doi.org/10.1186/s13071-022-05150-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-022-05150-9