
Abstract
Background
Trichomonas vaginalis is the causative agent of trichomoniasis, which is one of the most common sexually transmitted diseases worldwide. Trichomoniasis has a high incidence and prevalence and is associated with serious complications such as HIV transmission and acquisition, pelvic inflammatory disease and preterm birth. Although trichomoniasis is treated with oral metronidazole (MTZ), the number of strains resistant to this drug is increasing (2.5–9.6%), leading to treatment failure. Therefore, there is an urgent need to find alternative drugs to combat this disease.
Methods
Herein, we report the in vitro and in silico analysis of 12 furanyl N-acylhydrazone derivatives (PFUR 4, a-k) against Trichomonas vaginalis. Trichomonas vaginalis ATCC 30236 isolate was treated with seven concentrations of these compounds to determine the minimum inhibitory concentration (MIC) and 50% inhibitory concentration (IC50). In addition, compounds that displayed anti-T. vaginalis activity were analyzed using thiobarbituric acid reactive substances (TBARS) assay and molecular docking. Cytotoxicity analysis was also performed in CHO-K1 cells.
Results
The compounds PFUR 4a and 4b, at 6.25 µM, induced complete parasite death after 24 h of exposure with IC50 of 1.69 µM and 1.98 µM, respectively. The results showed that lipid peroxidation is not involved in parasite death. Molecular docking studies predicted strong interactions of PFUR 4a and 4b with T. vaginalis enzymes, purine nucleoside phosphorylase, and lactate dehydrogenase, while only PFUR 4b interacted in silico with thioredoxin reductase and methionine gamma-lyase. PFUR 4a and 4b led to a growth inhibition (< 20%) in CHO-K1 cells that was comparable to the drug of choice, with a promising selectivity index (> 7.4).
Conclusions
Our results showed that PFUR 4a and 4b are promising molecules that can be used for the development of new trichomonacidal agents for T. vaginalis.

Similar content being viewed by others
Background
Trichomonas vaginalis is an anaerobic, flagellated protozoan parasite, which infects the urogenital tract of humans causing trichomoniasis, the most prevalent non-viral sexually transmitted infection worldwide [1]. Trichomoniasis has an estimated incidence rate of 276 million cases per year [1]. In Brazil, the prevalence varies between 2.5% and 20% in women [1,2,3,4,5,6]. Trichomonas vaginalis has a greater impact on public health as it is associated with increased risk of preterm delivery, pelvic inflammatory disease [7] and HIV acquisition and transmission [8, 9]. It is important to note that non-adherence and/or noncompliance to treatment for trichomoniasis, which happens frequently, helps in maintaining the chain of infection [8].
For more than five decades, in most countries, the treatment for T. vaginalis has been restricted to 5-nitroimidazoles, with metronidazole (MTZ) and tinidazole (TNZ) being the only drugs approved by the U.S. Food and Drug Administration Agency [9, 10]. The activity of MTZ depends on the reduction of its nitro group by hydrogenosome metabolism and by flavin enzyme thioredoxin reductase (TrxR) [11]; however, the increasing number of T. vaginalis isolates resistant to 5-nitroimidazoles is a major concern [12,13,14] representing 2.5–9.6% of clinical cases [12, 13, 15]. Nonetheless, inhibition of TrxR and other enzymes such as lactate dehydrogenase (LDH), methionine gamma-lyase (MGL), and purine nucleoside phosphorylase (PNP) also seem to be a rational approach for chemotherapy against T. vaginalis due to their importance on protozoan survival [16,17,18].
In this context, there is a need to identify new molecules with potential trichomonacidal activity as an alternative to 5-nitroimidazoles. N-acylhydrazone derivatives have been found to have several biological properties, such as antimalarial [19], antitumor [20], antifungal [21] and antibacterial [22] activities, exhibiting good pharmacological potential. In addition, furan and related derivatives represent an important class of heterocyclic compounds that have been reported to exhibit a wide range of biological activities, such as antibacterial, antiviral, anti-inflammatory, antifungal, antitumor, etc. [23,24,25].
Considering the need for new trichomonacidal compounds, a series of furanyl N-acylhydrazone derivatives (PFUR) was designed by molecular hybridization technique combining both pharmacophoric groups, i.e. the N-acylhydrazone and furan moieties in the molecules. Therefore, this study was aimed at e the in vitro activity and selectivity of 12 synthesized PFUR derivatives against T. vaginalis and at identifying their likely binding to T. vaginalis proteins in silico.
Methods
Chemicals
The synthesis of furanyl N-acylhydrazone derivatives was carried out as described by Cardoso et al. [26] and the synthesized samples were kindly provided by Dr Marcus Vinicius Nora de Souza from the Instituto de Tecnologia em Fármacos-Far-Manguinhos, Fiocruz, Brazil. Briefly, the synthesis was carried out by the reaction between 2-(H2NNHCO-furan), which were generated from methyl furan-2-carboxylate, and arylaldehydes resulting in the formation of 12 derivatives, which are as follows: PFUR 4 (precursor), PFUR 4a, PFUR 4b, PFUR 4c, PFUR 4d, PFUR 4e, PFUR 4f, PFUR 4g, PFUR 4h, PFUR 4i, PFUR 4j and PFUR 4k (Fig. 1).

Chemical structures of PFUR 4 and 4a-k synthesized by the reaction of 2-(H2NNHCO-furan) and aryl aldehydes (EtOH, RCHO, r.t., 1–72 h, 40–97%)
Trichomonas vaginalis culture
The isolate T. vaginalis Donne ATCC 30236 that is susceptible to MTZ was axenically grown in pre-warmed trypticase-yeast extract-maltose (TYM) medium without agar (pH 6.0), supplemented with 10% sterile bovine serum (SBS) (previously inactivated at 56 °C for 30 min) with 3% streptomycin (5 mg/ml) and incubated at 37 °C under microaerobic conditions [27]. The motility and morphology of the culture were analyzed under light microscopy at 400× magnification. A trypan blue (0.4%) exclusion assay was performed to ensure that minimum viability of 95% and logarithmic growth phase had been achieved before proceeding to antiparasitic and biochemical assays.
Anti-T. vaginalis assay
Drug susceptibility assays were performed to analyze the antiparasitic potential of 12 PFUR compounds against T. vaginalis as previously described [28, 29]. All compounds were diluted in dimethylsulfoxide (DMSO). For the assay, 96-well microtiter plates were seeded with 2.6 × 105 trophozoites/ml of TYM (initial density), then PFUR drugs were added into the wells at a concentration of 100 µM, and the plates were incubated at 37 °C with 5% CO2 for 24 h. Following incubation, an aliquot of trophozoites and trypan blue (0.4%) (1:1, v/v) was counted in the Neubauer chamber to determine trophozoite motility, morphology and viability through trypan blue dye exclusion. Three controls were used in each assay: a negative control containing only trophozoites; a 0.6% DMSO control (vehicle for solubilization); and a positive control containing MTZ at 100 µM (Sigma-Aldrich, St. Louis, Missouri, USA). Only the compounds that reduced the viability of parasites by 100% were used in the subsequent experiments.
The minimum inhibitory concentration (MIC) and 50% inhibitory concentration (IC50) values against T. vaginalis were established in the same conditions described previously with trophozoites exposed for 24 h to PFUR concentrations ranging from 1.5625 µM to 100 µM. The MIC was confirmed by transferring the culture solutions contained in 96-well plates from MIC and from concentrations directly below and above, as well as controls, to tubes containing 1.5 ml of fresh TYM medium, with SBS and antibiotics. Tubes were then re-incubated at 37 °C with 5% CO2 for 96 h and trophozoites were counted in the Neubauer chamber every 24 h to confirm MIC. The IC50 was calculated using GraphPad Prism 7.0 software.
A kinetic growth curve was established to understand the time required for the antiparasitic activity of compounds on T. vaginalis cultures. 96-well plates were prepared as previously described, compounds were added at MIC, and the plates were incubated at 37 °C with 5% CO2 for 96 h. The viability of trophozoites was observed under light microscopy and the growth analysis was performed at 1, 6, 12, 24, 48, 72 and 96 h using trypan blue (0.4%) dye exclusion and motility and morphology analysis. All assays were performed in three independent set-ups in triplicate and the results were expressed as a percentage of viable trophozoites compared with the negative control.
Thiobarbituric acid reactive substances (TBARS) assay
The treatment and control groups were evaluated for the formation of TBARS in an acid-heating reaction, and malondialdehyde (MDA) was chosen as the biomarker for lipid peroxidation. The assay was performed as described by Ohkawa et al. [30] with modifications. For this, 96-well plates were seeded with T. vaginalis (2.6 × 105 trophozoites/ml), the compounds (at MIC) and controls were added, and plates were incubated at 37 °C with 5% CO2 for 24 h. After incubation, aliquots of T. vaginalis control and treatment groups were incubated with 0.8% TBA, acetic acid/HCl (pH 3.4), and 8.1% SDS (sodium dodecyl sulfate) at 95 °C for 2 h. The absorbance of the samples was measured at 532 nm and the results were expressed as nmol MDA/105 trophozoites.
Molecular docking
Molecular docking of PFUR compounds was performed using crystal structure of T. vaginalis purine nucleoside phosphorylase (TvPNP; PDB 1Z36), lactate dehydrogenase (TvLDH; PDB 5A1T), and methionine gamma-lyase (TvMGL; PDB 1E5E), which were retrieved from RSCB protein data bank (http://www.rcsb.org/). The structure of T. vaginalis thioredoxin reductase (TvTrxR) was constructed using the server SWISS-MODEL [31] based on the UniProt entry A2DSU2. Target protein structures were prepared by removing ligands, ions, and water with CHIMERA 1.5.3 [32] software. Non-polar hydrogen atoms were merged, Gasteiger atomic charges were assigned, the atom type of protein structures was specified, and the molecular docking simulation was performed using Autodock Vina 1.1.2 software [33]. The binding sites were defined by the co-complexed ligands in the crystal structure and the grid box was set up according to the corresponding residues. PFUR 4a, PFUR 4b, and MTZ were built and optimized in the software Avogadro 1.1.1 [34]. Followed by structural optimization, the ligands were prepared for docking by merging non-polar hydrogen atoms, defining rotatable bonds and assigning partial atomic charge using AutoDock Tools version 1.5.6 [35]. Docking poses of the investigated compounds were visualized using Accelrys Discovery Studio 3.5.
Cell culture
Chinese Hamster Ovary (CHO-K1) cells obtained from the Rio de Janeiro Cell Bank (PABCAM, Federal University of Rio de Janeiro, RJ, Brazil) were grown as a monolayer in Dulbecco’s Modified Eagle’s Medium (DMEM) (Vitrocell Embriolife, São Paulo, Brazil), supplemented with 10% fetal bovine serum (Vitrocell Embriolife), 1% l-glutamine, and 1% penicillin/streptomycin at 37 °C in an atmosphere of 95% humidified air and 5% CO2.
Cytotoxicity assay
The cytotoxicity assay was performed following a method previously described [28, 36]. CHO-K1 cell line viability and proliferation were determined by measuring the formation of water-insoluble formazan as a result of 3-(4,5-dimethythiazol-2-yl)–2,5-diphenyl tetrazolium bromide (MTT) reduction. In 96-well plates, 2 × 104 cells were seeded per well and grown at 37 °C, 5% CO2 for 24 h prior to the cell viability assay. After that, the cells were treated with compounds at 1.5625, 3.125, 6.25 and 12.5 μM for another 24 h at 37 °C, 5% CO2. In addition, 0.6% DMSO and MTZ at 100 μM were added as controls. After incubation, MTT (Sigma-Aldrich) at 5 mg/ml was added per well and the plates were re-incubated for 3 h at 37 °C, 5% CO2. Subsequently, DMSO was added to solubilize formazan crystals. The absorbance was measured on a microplate reader at a wavelength of 492 nm. The inhibition of cell growth was determined according to the formula: Cell proliferation inhibition (%) = (1 − Abs492 treated cells/Abs492 control cells) × 100. The 50% cytotoxic concentration (CC50) was calculated using GraphPad Prism 7.0 software. The assay was performed at least three independent times in triplicate. The selectivity index (SI) was calculated through the ratio CC50/IC50.
Statistical analysis
Statistical analysis was carried out by two-way analysis of variance (ANOVA) for the kinetic growth curve and for all other assays by one-way ANOVA, both followed by Tukey’s post-hoc test for multiple comparisons between groups. All tests were performed using GraphPad Prism 7.03 software, considering a probability value of P < 0.05. Data are expressed as the mean ± standard deviation (SD).
Results
Anti-T. vaginalis assay
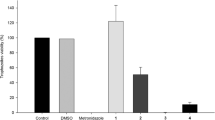
The screening for trichomonacidal activity showed the potential of two compounds from the PFUR group, 4a and 4b, as both reduced the trophozoites viability by 100% at 100 µM after 24 h (ANOVA: F(14, 30) = 57.32, P < 0.0001). Moreover, PFUR 4h had no effect on trophozoites growth, PFUR 4d and 4f were able to reduce growth by 26.3% and 43.1%, respectively; however, such reduction was not significant when compared with MTZ, as for other compounds (PFUR 4, 4c, 4e, 4g, 4i-k) viability reduction rates varied from 5.5% to 18% and were not statistically different from the negative control group (ANOVA: F(14, 30) = 57.32, P < 0.0001) (Fig. 2). Unsurprisingly, 0.6% DMSO and negative controls induced no reduction in viability, exhibited negative trypan blue (0.4%) staining, and showed good motility and morphology, whereas treatment with MTZ completely diminished viability and exhibited positive trypan blue staining and negative motility.

In vitro trichomonacidal activity of PFUR 4 and 4a-k at 100 µM against Trichomonas vaginalis ATCC 30236 isolate, confirmed by the trypan blue (0.4%) assay after 24 h of exposure: Control (untreated trophozoites), 0.6% DMSO (vehicle for solubilization), MTZ (metronidazole at 100 µM). Viability of 100% in control corresponds to 2.6 × 105 trophozoites/ml. Data are presented as the mean ± standard deviation of at least three independent experiments. Different letters show a significant difference at P < 0.05
Considering the full viability reduction caused by PFUR 4a and 4b, these compounds were chosen for antiparasitic and biochemical tests and molecular docking. The tests showed that PFUR 4a (ANOVA: F(9, 20) = 317.1, P < 0.0001) and PFUR 4b (ANOVA: F(9, 20) = 799.5, P < 0.0001) were able to completely reduce trophozoites viability at 6.25 µM (Fig. 3a, b), which was determined as MIC. IC50 was statistically found to be 1.69 µM and 1.98 µM for PFUR 4a and PFUR 4b, respectively, after 24 h exposure (Table 1).

Anti-Trichomonas vaginalis assay. MIC and IC50 for the antiparasitic activity of PFUR 4a (a) and PFUR 4b (b) against Trichomonas vaginalis ATCC 30236 after exposure to 1.5625, 3.125, 6.25, 12.5, 25, 50 and 100 µM concentrations for 24 h. Kinetic growth curves of Trichomonas vaginalis ATCC 30236 after 1, 6, 12, 24, 48, 72 and 96 h of treatment with PFUR 4a (c) and PFUR 4b (d) at 6.25 µM. Growth was completely inhibited after 24 h. Control (untreated trophozoites), 0.6% DMSO (vehicle for solubilization), MTZ (metronidazole at 100 µM). Viability of 100% in control corresponds to 2.6 × 105 trophozoites/ml. Data are presented as the mean ± standard deviation of at least three independent experiments. Different letters show a significant difference at P < 0.05
The data obtained from kinetic growth curve showed that PFUR 4a at 6.25 µM was able to reduce trophozoite growth by 36.8% at 12 h and 98.7% at 24 h; conversely, the negative control maintained full growth at all exposure times and MTZ completely inhibited growth at 12 h exposure (Two-way ANOVA: F(12, 36) = 299.2, P < 0.0001) (Fig. 3c). As for PFUR 4b at 6.25 µM, growth reduction was 79.9% at 12 h and 99.7% at 24 h, whereas for the negative control and MTZ, the profile was the same as described above (Two-way ANOVA: F(12, 36) = 401.1, P < 0.0001) (Fig. 3d). For both compounds, there were no viable trophozoites after 24 h.
TBARS assay
Data analysis demonstrated that treatment with 0.6% DMSO did not increase lipid peroxidation levels when compared with the control group (untreated trophozoites). The results also showed that MTZ, PFUR 4a, and PFUR 4b treatments were unable to significantly induce lipid peroxidation levels than those of the control (ANOVA: F(4, 25) = 4.901, P= 0.0047) (Fig. 4).

Lipid peroxidation levels measured through thiobarbituric acid reactive substances assay with malondialdehyde (MDA) as a biomarker, after 24 h of exposure to PFUR 4a and 4b, both at 6.25 µM. Control (untreated trophozoites), DMSO (vehicle for solubilization), MTZ (metronidazole at 100 µM). Data are presented as the mean ± standard deviation of at least three independent experiments
Molecular docking
Next, a molecular docking study was carried out with the two most active compounds, PFUR 4a and 4b, to investigate their possible mechanism of action through their in silico predicted interaction with proteins that play a role in T. vaginalis survival.
We noticed that PFUR 4a interacts in silico with TvPNP with binding free energy (ΔGbind) of − 6.4 kcal/mol and four residues may be involved in hydrogen bonds with PFUR 4a (Fig. 5a, b, Table 2). As shown in Fig. 5c and d, PFUR 4a could possibly form eight conventional hydrogen bonds and one carbon-hydrogen bond (SER240: 2.29 Å) with active site residues of the enzyme TvLDH, alongside other non-covalent interaction that yield a docking score of − 5.8 kcal/mol (Table 2). We also performed docking studies of PFUR 4a with enzymes TvTrxR and TvMGL; however, no favorable binding mode was found (data not shown).

Representation of 2D projection and predicted binding mode of PFUR 4a with the T. vaginalis enzymes TvPNP (a, b) and TvLDH (c, d). The distance (Å) of the hydrogen bonds between specific residues and PFUR 4a is shown in green
In the case of PFUR 4b, Fig. 6a and b show that it may form four conventional hydrogen bonds and one carbon-hydrogen bond (GLY89) with the active site residues of the enzyme TvPNP, alongside other non-covalent interactions that yield a docking score of − 6.8 kcal/mol (Table 2). PFUR 4b has a significant binding score of − 6.0 kcal/mol with TvTrxR as shown in Fig. 6c and d and it could be involved in six hydrogen bonds that might help in maintaining the binding mode (Table 2).

Representation of 2D projection and predicted binding mode of PFUR 4b with the T. vaginalis enzymes TvPNP (a, b), TvTrxR (c, d), TvLDH (e, f) and TvMGL (g, h). The distance (Å) of the hydrogen bonds between specific residues and PFUR 4b is shown in green
It is worth mentioning that PFUR 4b might potentially form seven hydrogen bonds with residues in the active site of TvLDH (Fig. 6e, f, and Table 2). This binding mode is complemented with hydrophobic interactions and Van der Waals forces, generating a ΔGbind of − 5.9 kcal/mol (Table 2). Furthermore, a likely binding mode of PFUR 4b in the active site of TvMGL is shown in Fig. 6g and h (docking score of − 5.7 kcal/mol). Hydrogen bonds might be formed between PFUR 4b and three residues of TvMGL, and other chemical interactions may help in maintaining the binding mode of the compound (Table 2).
Considering that MTZ is the reference drug for T. vaginalis treatment, we also investigated its possible interaction with the abovementioned enzymes. In silico analysis showed MTZ possibly binds to the active site of TvPNP (ΔGbind of − 5.0 kcal/mol; Fig. 7a, b), TvLDH (ΔGbind of − 5.2 kcal/mol; Fig. 7c, d), and TvMGL (ΔGbind of − 4.2 kcal/mol; Fig. 7e, f). The possible hydrogen bonds and other interactions of MTZ with the enzymes can be seen in Table 2.

Representation of 2D projection and predicted binding mode of metronidazole (MTZ) with the T. vaginalis enzymes TvPNP (a, b), TvLDH (c, d) and TvMGL (e, f). The distance (Å) of the hydrogen bonds between specific residues and MTZ is shown in green
Cytotoxic assay
The MTT assay showed that PFUR 4a induced 18.6% of growth inhibition at 6.25 µM and only 4.4% at 3.125 µM, none of the concentrations tested for this compound were significantly different from MTZ (ANOVA: F(5, 12) = 15.98, P < 0.0001), which caused 8.5% of inhibition (Fig. 8a). For PFUR 4b, the results demonstrated 8.8% of growth inhibition at 6.25 µM and 4.9% inhibition at 3.125 µM, once again none of the concentrations tested were significantly different from MTZ (ANOVA: F(5, 12) = 9.427, P= 0.0008) (Fig. 8b). As expected, 0.6% DMSO did not alter cell growth and 1.5625 µM concentrations of both compounds also had no effect on cells. The CC50 for both compounds was higher than the highest concentration tested (12.5 µM) (Table 1) because less than 50% of cell growth was inhibited at this concentration (Fig. 8a, b).

Cytotoxicity effect of PFUR 4a (a) and PFUR 4b (b) at 1.5625, 3.125, 6.25 and 12.5 µM on CHO-K1 cells through MTT assay after 24 h of exposure; 0.6% DMSO (vehicle for solubilization) and MTZ (metronidazole at 100 µM). Data are presented as the mean ± standard deviation of at least three independent experiments. Different letters show a significant difference between treatments at P < 0.05
Discussion
Trichomoniasis has a significant impact on public health due to its high incidence and prevalence worldwide [1]. The infection is associated with HIV acquisition and transmission, a higher risk of preterm delivery, and a higher prevalence of HSV-1 and HSV-2 [37,38,39,40], aggravating its effect on people’s health. In addition, with an increase in treatment failures resulting in recurrent infections, there is an obvious need to develop new alternatives for treating trichomoniasis. Our data showed two furanyl N-acylhydrazone derivatives, PFUR4a and 4b, presented in vitro antiprotozoal activity against T. vaginalis (Fig. 2) in promising concentrations and with a considerably good selectivity towards the parasite. Our in silico and in vitro analyses indicated that these two compounds are promising treatment alternatives, which may act slightly differently from the reference drug, and should be further explored.
In this context, we highlight that both PFUR 4a and 4b, which demonstrated a high trichomonacidal activity, presented a MIC value of 6.25 µM (Fig. 3a, b) and IC50 of 1.69 µM and 1.98 µM, respectively (Table 1). Moreover, a kinetic growth analysis showed that PFUR 4a and 4b, both at 6.25 µM (MIC), were able to fully reduce T. vaginalis viability to a level that was not significantly different from MTZ after a 24 h treatment (Fig. 3c, d). Similar kinetic growth inhibition profiles have been demonstrated in previous studies [28, 36]. The furanyl N-acylhydrazone derivatives tested in this study were previously investigated for their antitubercular potential and PFUR 4b exhibited moderate activity against Mycobacterium tuberculosis with a MIC of 100.3 µM [26]. When compared, these results suggest that these compounds might be more active/selective against protozoan species.
Other N-acylhydrazone derivatives have been reported as antiparasitic agents against Trypanosoma cruzi in vitro and in vivo in promising concentrations and also showed low cytotoxicity against mammalian cells in both cases [41], consequently demonstrating a great level of selectivity towards protozoan species. However, to the best of authors’ knowledge there are no prior reports of compounds containing the N-acylhydrazone moiety being active against T. vaginalis, or a species of the genus Trichomonas. As for the furan moiety, previous studies have shown that compounds containing this pharmacophoric group are active against T. vaginalis and Giardia lamblia [42, 43]. These studies substantiate the antiprotozoan potential of furanyl N-acylhydrazone derivatives demonstrated in our results.
The compounds PFUR 4a and 4b are quite similar in structure, the former having a 5-nitrofuran-2-yl radical and the latter a 5-nitrothiopen-2-yl radical, and both compounds are distinguished from the other 10 compounds by having this nitro (NO2) substituent in their structure. Similarly, MTZ also has a nitro group in its structure and this group plays an essential role in the production of toxic intermediates after its reduction and MTZ activation [44]. Considering these similarities, it has been speculated that nitro groups of PFUR 4a and 4b might play a role in altering the redox equilibrium in hydrogenosomes leading to the trichomonacidal activity reported here. Similarly, a previous study on N-acylhydrazones containing 4-hydroxy-3-methoxyphenyl showed antiparasitic activity against Plasmodium falciparum demonstrating that the addition of a nitro group significantly increased the compound’s activity, indicating that nitro groups could either induce or enhance antiparasitic activity against protozoan species [19]. In addition, the structural resemblance of compounds PFUR 4a and 4b could help elucidate the similar results observed in the in vitro antiparasitic and biochemical assays performed here.
Moreover, the TBARS assay demonstrated that T. vaginalis trophozoites exposed to PFUR 4a and 4b for 24 h did not present a level of lipid peroxidation significantly higher than the one observed for the control group (Fig. 4). Considering that lipid peroxidation is a chain of reactions mediated by free radicals that results in an oxidative deterioration of polyunsaturated lipids, which are abundant in components of biological membranes [45], these results indicate that membrane damage is not a significant factor to be considered as a mechanism by which these compounds induce T. vaginalis death.
Docking studies were performed to gain insight into the binding mode of compounds, PFUR 4a and PFUR 4b, with enzymes required for T. vaginalis survival. PFUR 4a presented a satisfactory binding mode with TvPNP and TvLDH, while PFUR 4b interacted in silico with TvPNP, TvTrxR, TvLDH and TvMGL (Figs. 5, 6 and Table 2). The results indicate that these enzymes could be somehow involved in these compounds’ action against T. vaginalis.
Trichomonas vaginalis PNP is a well-known potential target for chemotherapy [46, 47] and both compounds interact in silico with key residues in the active site of the enzyme, which are relevant for the inhibition of its activity, indicating they may act as TvPNP inhibitors. Based on the metabolic deficiency of T. vaginalis towards the lack of de novo synthesis of purine nucleosides and the requirement of purine salvage to maintain their purine nucleotide pools in order to survive [16], modulation of this pathway may be involved in the mechanism of action of PFUR 4a and PFUR 4b against T. vaginalis, highlighting the significance of this predicted interaction. Furthermore, based on the significance of TvLDH for parasite survival and in silico interactions described, impairment of energy metabolism could also explain the reduction in viability of the flagellated parasitic protozoan T. vaginalis by PFUR 4a and PFUR 4b.
Likewise, in silico interaction of PFUR 4b with key residues in the active site of TvTrxR and TvMGL could indicate a possible modulation of both enzymes, probably through their inhibition, by this compound. Considering the exclusivity of in silico interaction of PFUR 4b with TvTrxR and TvMGL when compared to PFUR 4a, we may hypothesize that modulation of antioxidant defense (through TvTrxR) and the content of sulfur-containing amino acids (via TvMGL) could account for the faster onset of action of PFUR 4b when compared to PFUR 4b (Fig. 3c, d). It is worth mentioning that TvMGL is a potential drug target for anti-T. vaginalis chemotherapy since it is not found in mammals, and the structure of TvTrxR is very different from human TrxR [18], highlighting the biological application of PFUR 4b. Nonetheless, more studies must be conducted to confirm this hypothesis and improve our understanding of the mechanism of action for PFUR 4a and PFUR 4b.
In order to establish a comparison between the putative effects of PFUR 4a and 4b and the known activity of MTZ, we also analyzed the possible binding mode of MTZ with the enzymes investigated here. Interestingly, the in silico interactions of TvPNP, TvLDH, and TvMGL were stronger with PFUR 4a and 4b than with MTZ (Fig. 7 and Table 2). In T. vaginalis, among the actions described [11, 44, 48], MTZ is said to disrupt the DNA helical structure inhibiting the synthesis of proteins that result in cell death [44, 48]; therefore, inhibiting the enzymes investigated here may not be required for the action of MTZ. This indicates that the mechanism of action of PFUR 4a and 4b might be somewhat different from the mechanism of action of MTZ, highlighting that PFUR 4a and 4b may act via modulation of energy metabolism, antioxidant defenses, and the content of sulfur-containing amino acids. Nonetheless, more studies are needed to support this hypothesis.
The cytotoxicity assay performed in CHO-K1 cells showed that at their MIC (6.25 µM), both compounds exhibited no cytotoxic effect on non-tumor/normal cells (Fig. 8a, b) according to the standards set by the International Organization for Standardization (ISO) at ISO 10993-5:2009, which states that a decrease of cell viability by more than 30% is considered a cytotoxic effect [49, 50]. Therefore, since PFUR 4a and PFUR 4b reduced viability by 18.6% and 8.8% at their MIC, respectively, this inhibition of growth does not represent a cytotoxic effect. In addition, levels of cell growth inhibition are significantly the same as the one found for MTZ, demonstrating that these compounds do not affect cells more than the commercial drugs used for trichomoniasis. Lastly, both compounds showed a promising SI (Table 1), which can be much better than currently presented considering that the higher concentration tested (12.5 µM) inhibited less than 20% of the mammalian cells. Altogether, these results emphasize the potential application of these molecules.
Finally, the need for new and effective molecules for treating trichomoniasis is emerging exponentially and all data gathered here show that furanyl N-acylhydrazone derivatives, PFUR 4a and 4b, have a highly significant antiparasitic activity against T. vaginalis with a great level of selectivity for the parasite over non-tumor/normal cells and are, therefore, promising molecules for studies on new trichomonacidal agents.
Conclusions
Our study demonstrates that furanyl N-acylhydrazone derivatives PFUR 4a and PFUR 4b have a potent anti-T. vaginalis activity in vitro, which is statistically similar to the reference drug. At low concentrations, these compounds show good and promising selectivity towards T. vaginalis. Moreover, molecular docking analyses point to the modulation of enzymes TvPNP, TvLDH, TvTrxR, and TvMGL by these compounds. In conclusion, our data characterized PFUR 4a and PFUR 4b as promising trichomonacidal agents and support further studies not only with these compounds but also other N-acylhydrazone derivatives, as alternatives for treating trichomoniasis.
Availability of data and materials
Data supporting the conclusions of this article are provided within the article. The datasets used and/or analyzed during the present study are available from the corresponding author upon reasonable request.
Abbreviations
- PFUR:
-
Furanyl N-acylhydrazone derivatives
- STI:
-
Sexually transmitted infection
- ATCC:
-
American type culture collection
- MIC:
-
Minimum inhibitory concentration
- IC50 :
-
50% inhibitory concentration
- CC50 :
-
50% cell inhibitory concentration
- WHO:
-
World Health Organization
- TvPNP:
-
T. vaginalis purine nucleoside phosphorylase
- TvLDH:
-
T. vaginalis lactate dehydrogenase
- TvMGL:
-
T. vaginalis methionine gamma-lyase
- TvTrxR:
-
T. vaginalis thioredoxin reductase
- MTZ:
-
Metronidazole
- DMSO:
-
Dimethylsulfoxide
- TBARS:
-
Thiobarbituric acid reactive substances
- MDA:
-
Malondialdehyde
- TBA:
-
Thiobarbituric acid
- CHO-K1:
-
Chinese hamster ovary cells
- MTT:
-
3-(4,5-dimethythiazol-2-yl)–2,5-diphenyl tetrazolium bromide
- SI:
-
Selective index
References
WHO. Global incidence and prevalence of selected curable sexually transmitted infections-2008. Geneva: World Health Organization; 2012. https://www.who.int/reproductivehealth/publications/rtis/stisestimates/en/. Accessed 31 Aug 2019.
Grama DF, da Casarotti LS, de Morato MGVA, Silva LS, Mendonça DF, Limongi JE, et al. Prevalence of Trichomonas vaginalis and risk factors in women treated at public health units in Brazil: a transversal study. Trans R Soc Trop Med Hyg. 2013;107:584–91.
Rocha DAP, Filho RAAB, Mariño JM, dos Santos CMB. “Hidden” sexually transmitted infections among women in primary care health services, Amazonas, Brazil. Int J STD AIDS. 2014;25:878–86.
Bruni MP, Lopes S, Stauffert D, Caetano C, Filho C, Siqueira L, et al. Trichomonas vaginalis infection among women attending frequency, risk factors and clinical signs. DST-J bras Doenças Sex Transm. 2015;27:86–91.
Ambrozio CL, Nagel AS, Jesk S, Bragança GCM, Borsuk S, Villela MM. Trichomonas vaginalis prevalence and risk factors for women in southern Brazil. Rev Inst Med Trop Sao Paulo. 2016;58:1–5.
Dos Gatti FA, Ceolan E, Greco FSR, Santos PC, Klafke GB, de Oliveira GR, et al. The prevalence of trichomoniasis and associated factors among women treated at a university hospital in southern Brazil. PLoS ONE. 2017;12:e0173604.
Poole DN, McClelland RS. Global epidemiology of Trichomonas vaginalis. Sex Transm Infect. 2013;89:418–22.
Menezes CB, Frasson AP, Tasca T. Trichomoniasis—are we giving the deserved attention to the most common non-viral sexually transmitted disease worldwide? Microb Cell. 2016;3:404–18.
Meites E, Gaydos CA, Hobbs MM, Kissinger P, Nyirjesy P, Schwebke JR, et al. A review of evidence-based care of symptomatic trichomoniasis and asymptomatic Trichomonas vaginalis infections. Clin Infect Dis. 2015;61:S837–48.
FDA. FDA approved drug products. https://www.accessdata.fda.gov/scripts/cder/daf/. Accessed 31 Aug 2019.
Leitsch D, Kolarich D, Binder M, Stadlmann J, Altmann F, Duchêne M. Trichomonas vaginalis: metronidazole and other nitroimidazole drugs are reduced by the flavin enzyme thioredoxin reductase and disrupt the cellular redox system Implications for nitroimidazole toxicity and resistance. Mol Microbiol. 2009;72:518–36.
Schwebke JR, Barrientes FJ. Prevalence of Trichomonas vaginalis isolates with resistance to metronidazole and tinidazole. Antimicrob Agents Chemother. 2006;50:4209–10.
Kirkcaldy RD, Augostini P, Asbel LE, Bernstein KT, Kerani RP, Mettenbrink CJ, et al. Trichomonas vaginalis antimicrobial drug resistance in 6 US cities, STD surveillance network, 2009–2010. Emerg Infect Dis. 2012;18:939–43.
Workowski KA, Bolan GA. Sexually transmitted diseases treatment guidelines, 2015. MMWR recommended reports. Atlanta: Center for Disease Control; 2015.
Das S, Huengsberg M, Shahmanesh M. Treatment failure of vaginal trichomoniasis in clinical practice. Int J STD AIDS. 2005;16:284–6.
Munagala NR, Wang CC. Adenosine is the primary precursor of all purine nucleotides in Trichomonas vaginalis. Mol Biochem Parasitol. 2003;127:143–9.
Sato D, Nozaki T. Methionine gamma-lyase: the unique reaction mechanism, physiological roles, and therapeutic applications against infectious diseases and cancers. IUBMB Life. 2009;61:1019–28.
Setzer M, Byler K, Ogungbe I, Setzer W. Natural products as new treatment options for trichomoniasis: a molecular docking investigation. Sci Pharm. 2017;85:5.
Dos Santos Filho JM, de Queiroz E Silva DMA, Macedo TS, Teixeira HMP, Moreira DRM, Challal S, et al. Conjugation of N-acylhydrazone and 1,2,4-oxadiazole leads to the identification of active antimalarial agents. Bioorganic Med Chem. 2016;24:5693–701.
Do Amaral DN, Cavalcanti BC, Bezerra DP, Ferreira PMP, De Castro RP, Sabino JR, et al. Docking, synthesis and antiproliferative activity of N-acylhydrazone derivatives designed as combretastatin A4 analogues. PLoS ONE. 2014;9:e85380.
Cachiba TH, Carvalho BD, Carvalho DT, Cusinato M, Prado CG, Dias ALT. Síntese e avaliação preliminar da atividade antibacteriana e antifúngica de derivados N-acilidrazônicos. Quim Nova. 2012;35:1566–9.
Zhang H, Kunadia A, Lin Y, Fondell JD, Seidel D, Fan H. Identification of a strong and specific antichlamydial N-acylhydrazone. PLoS ONE. 2017;12:e0185783.
Lukevits E, Demicheva L. Biological activity of furan derivatives (review). Chem Heterocycl Compd. 1993;29:243–67.
Banerjee R, Kumar HKS, Banerjee M. Medicinal significance of furan derivatives: a review. Int J Rev Life Sci. 2012;2:7–16.
Keay BA, Dibble PW. In: Katritzky AR, Rees CW, Scriven EFV, editors. In: Comprehensive heterocyclic chemistry II, a review of literature 1982–1995: the structure, reactions, synthesis, and uses of heterocyclic compounds. 1st ed. Oxford: Pergamon Press; 1996. p. 395–436.
de Faria Cardoso LN, Nogueira TCM, Kaiser CR, Wardell JL, Wardell SMSV, de Souza MVN. Synthesis and anti-tubercular activity of thienyl and furanyl derivatives. Mediterr J Chem. 2016;5:356–66.
Diamond LS. The establishment of various trichomonads of animals and man in axenic cultures. J Parasitol. 1957;43:488–90.
Sena-Lopes Â, das Neves RN, Bezerra FSB, de Oliveira Silva MT, Nobre PC, Perin G, et al. Antiparasitic activity of 1,3-dioxolanes containing tellurium in Trichomonas vaginalis. Biomed Pharmacother. 2017;89:284–7.
Rigo GV, Petro-silveira B, Devereux M, Mccann M, Luis A, Tasca T. Anti-Trichomonas vaginalis activity of metallodrugs and synergistic effect with metronidazole. Parasitology. 2019;146:1179–83.
Ohkawa H, Ohishi N, Yagi K. Assay for lipid peroxides in animal tissues thiobarbituric acid reaction. Anal Biochem. 1979;95:351–8.
Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018;46:W296–303.
Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera? A visualization system for exploratory research and analysis. J Comput Chem. 2004;25:1605–12.
Trott O, Olson AJ. AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem. 2010;31:455–61.
Hanwell MD, Curtis DE, Lonie DC, Vandermeersch T, Zurek E, Hutchison GR. Avogadro: an advanced semantic chemical editor, visualization, and analysis platform. J Cheminform. 2012;4:17.
Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, et al. AutoDock4 and AutoDockTools4: automated docking with selective receptor flexibility. J Comput Chem. 2009;30:2785–91.
da Carapina Silva C, Pacheco BS, das Neves RN, Dié Alves MS, Sena-Lopes A, Moura Â, et al. Antiparasitic activity of synthetic curcumin monocarbonyl analogues against Trichomonas vaginalis. Biomed Pharmacother. 2019;111:367–77.
McClelland RS, Sangaré L, Hassan WM, Lavreys L, Mandaliya K, Kiarie J, et al. Infection with Trichomonas vaginalis increases the risk of HIV-1 acquisition. J Infect Dis. 2007;195:698–702.
Van Der Pol B, Kwok C, Pierre-Louis B, Rinaldi A, Salata RA, Chen P-L, et al. Trichomonas vaginalis infection and human immunodeficiency virus acquisition in African women. J Infect Dis. 2008;197:548–54.
Allsworth JE, Ratner JA, Peipert JF. Trichomoniasis and other sexually transmitted infections: results from the 2001–2004 national health and nutrition examination surveys. Sex Transm Dis. 2009;36:738–44.
Kissinger P. Trichomonas vaginalis: a review of epidemiologic, clinical and treatment issues. BMC Infect Dis. 2015;15:307.
dos Santos Filho JM, Moreira DRM, de Simone CA, Ferreira RS, McKerrow JH, Meira CS, et al. Optimization of anti-Trypanosoma cruzi oxadiazoles leads to identification of compounds with efficacy in infected mice. Bioorg Med Chem. 2012;20:6423–33.
Meneses-Marcel A, Rivera-Borroto OM, Marrero-Ponce Y, Montero A, Machado Tugores Y, Escario JA, et al. New antitrichomonal drug-like chemicals selected by bond (edge)-based TOMOCOMD-CARDD descriptors. J Biomol Screen. 2008;13:785–94.
Navin P, Sarvil P, Amit P, Divyesh P, Dhansukh R, Moo-Puc R, et al. Synthesis and biological evaluation of newer 1,3,4-oxadiazoles incorporated with benzothiazepine and benzodiazepine moieties. Zeitschr Naturforschung C. 2017;72:133–46.
Kulda J. Trichomonads, hydrogenosomes and drug resistance. Int J Parasitol. 1999;29:199–212.
Grotto D, Maria LS, Valentini J, Paniz C, Schmitt G, Garcia SC, et al. Importance of the lipid peroxidation biomarkers and methodological aspects for malondialdehyde quantification. Quim Nova. 2009;32:169–74.
Zang Y, Wang W-H, Wu S-W, Ealick SE, Wang CC. Identification of a subversive substrate of Trichomonas vaginalis purine nucleoside phosphorylase and the crystal structure of the enzyme-substrate complex. J Biol Chem. 2005;280:22318–25.
Rinaldo-Matthis A, Wing C, Ghanem M, Deng H, Wu P, Gupta A, et al. Inhibition and structure of Trichomonas vaginalis purine nucleoside phosphorylase with picomolar transition state analogues. Biochemistry. 2007;46:659–68.
Edwards DI. Nitroimidazole drugs-action and resistance mechanisms I. Mechanism of action. J Antimicrob Chemother. 1993;31:9–20.
ISO-International Organazition for Standardization. ISO 10993-5:2009-Biological evaluation of medical devices. Part 5: tests for in vitro cytotoxicity. Switzerland: International Organazition for Standardization; 2009.
Jung O, Smeets R, Hartjen P, Schnettler R, Feyerabend F, Klein M, et al. Improved in vitro test procedure for full assessment of the cytocompatibility of degradable magnesium based on ISO 10993-5/-12. Int J Mol Sci. 2019;20:255.
Acknowledgments
Not applicable.
Funding
This study was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brazil (CAPES)-Finance Code 001-Masters Scholarship (MSDA, Grant Number 1693097).
Author information
Authors and Affiliations
Contributions
MSDA performed (or participated in the execution of) all experiments, conducted data analysis, and wrote and edited the manuscript. ASL and SB conceived the study and provided expertise for the experimental work. RNN performed the anti-Trichomonas vaginalis assay. NVS performed the cytotoxicity assay. MD performed the TBARS assay. AMC performed the molecular docking. TCMN and MVNS synthesized the furanyl N-acylhydrazone derivatives. LS, FKS, and TC provided expertise for the experimental work. MSDA, RNN, ASL, MD, AMC, NVS, TCMN, MVNS, LS, FKS, TC and SB reviewed and contributed to final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Alves, M.S.D., das Neves, R.N., Sena-Lopes, Â. et al. Antiparasitic activity of furanyl N-acylhydrazone derivatives against Trichomonas vaginalis: in vitro and in silico analyses. Parasites Vectors 13, 59 (2020). https://doi.org/10.1186/s13071-020-3923-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-3923-8