
Abstract
Polycystic ovarian syndrome (PCOS) is a disorder with a foundation of neuroendocrine dysfunction, characterized by increased gonadotropin-releasing hormone (GnRH) pulsatility, which is antagonized by dopamine. The dopamine receptor 2 (DRD2), encoded by the DRD2 gene, has been shown to mediate dopamine’s inhibition of GnRH neuron excitability through pre- and post-synaptic interactions in murine models. Further, DRD2 is known to mediate prolactin (PRL) inhibition by dopamine, and high blood level of PRL have been found in more than one third of women with PCOS. We recently identified PRL as a gene contributing to PCOS risk and reported DRD2 conferring risk for type 2 diabetes and depression, which can both coexist with PCOS. Given DRD2 mediating dopamine’s action on neuroendocrine profiles and association with metabolic-mental states related to PCOS, polymorphisms in DRD2 may predispose to development of PCOS. Therefore, we aimed to investigate whether DRD2 variants are in linkage to and/or linkage disequilibrium (i.e., linkage and association) with PCOS in Italian families. In 212 Italian families, we tested 22 variants within the DRD2 gene for linkage and linkage disequilibrium with PCOS. We identified five novel variants significantly linked to the risk of PCOS. This is the first study to identify DRD2 as a risk gene in PCOS, however, functional studies are needed to confirm these results.
Similar content being viewed by others


Introduction
Polycystic ovarian syndrome (PCOS) is a common endocrinopathy of reproductive age women with a worldwide prevalence of 5–10% [1]. It is classically defined by oligo-/anovulation, hyperandrogenism, and formation of peripheral antral follicles in the ovaries, and it is associated with infertility, insulin resistance, type 2 diabetes, and depression [2,3,4,5,6]. PCOS is a disorder with a foundation of neuroendocrine dysfunction, characterized by increased gonadotropin-releasing hormone (GnRH) pulsatility and luteinizing hormone (LH) and follicle-stimulating hormone (FSH) ratio [7]. The pathogenesis of PCOS, however, is multifaceted and heterogenous, rooted in complex environmental and genetic etiology not yet fully defined, although clustering and susceptibility loci demonstrate a genetic component [8,9,10].
Reproductive function is primarily driven by the hypothalamic-pituitary-gonadal (HPG) axis [11]. Pulsatile GnRH secretion by the hypothalamus induces LH and FSH release by the anterior pituitary [11]. The HPG axis is regulated by numerous hormones, peptides, and neurotransmitters [11]. Aberrations in these regulating components thereby may induce endocrinopathies such as PCOS. Ovarian thecal and stromal cell hyperplasia and hyperandrogenism has been suggested to be a consequence of LH surplus sustained by rapid GnRH pulse frequency [12]. Of note, dopamine is antagonistic to GnRH release and pulsatility [13,14,15]. Also, prolactin (PRL) blood levels if elevated can suppress GnRH, and women with PCOS have been found having high PRL blood levels [16]. Dopamine levels are higher in patients with PCOS [17], and dopamine infusions have been shown to induce a significant reduction in LH levels in humans, and bromocriptine, a dopamine agonist, has demonstrated efficacy in reducing LH:FSH ratios and restoring ovarian function in women with PCOS [18, 19]. Dopamine exerts its effects through dopamine receptors expressed in the brain and peripheral tissues [20].
The dopamine receptor 2 (DRD2), encoded by the DRD2 gene, has been shown to mediate dopamine’s inhibition of GnRH neuron excitability through pre- and post-synaptic interactions in murine models [14] and to suppress PRL secretion [21]. Thus, if DRD2 carries genetic variants affecting its function, it can contribute to higher PRL levels inhibiting the reproductive cycles and/or contribute to GnRH abnormal pulsatility. Studies in ewe models have also suggested that DRD2 affects hypothalamic GnRH gene expression, and DRD2 specific antagonists stimulate GnRH and LH pulsatility [22]. Related to this, lowered brain dopamine levels and reduced DRD2 expression have been found in PCOS rat models [23]. DRD2 may also play a role in metabolic phenotypes associated with PCOS. Studies have demonstrated that reduced DRD2 activation is involved in development of insulin resistance in obese mice, and pharmaceutical agonism of DRD2 alleviates insulin resistance in animals [24, 25]. Importantly, insulin resistance is considered an important associated feature of hyperandrogenic anovulation in PCOS [26, 27]. Reduction in dopaminergic tone, as well as DRD2 signaling and regulation is also implicated in follicles of PCOS ovaries from rat PCOS models [28].
Given DRD2-mediated dopamine inhibition of PRL [21], and the elevated PRL blood levels found in PCOS [16], we studied the PRL pathway in PCOS and recently identified the prolactin receptor (PRLR) gene as contributing to PCOS risk [29]. Of note, we also reported DRD2 [30] and PRL-variants [31] conferring risk for type 2 diabetes and depression, which can both coexist with PCOS [6, 32]. Given dopamine’s action through DRD2 in neuroendocrine profiles and association with metabolic-mental states related to PCOS, polymorphisms in DRD2 may predispose to development of PCOS. Therefore, we aimed to investigate whether DRD2 variants confer risk to PCOS in Italian families.
Materials and methods
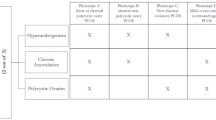
Among 212 Italian families with rich type 2 diabetes (T2D) family history, phenotyped for PCOS per Rotterdam diagnostic criteria [33]. cases were selected if at least two of the following were present: chronic anovulation or oligomenorrhea, clinical or biochemical hyperandrogenism, and/or polycystic ovaries [33]. We amplified 22 microarray-based single nucleotide polymorphisms (SNPs) located within the DRD2 gene (Supplementary Table 1). We excluded genotyping and Mendelian errors using PLINK [34]. We then analyzed the 22 SNPs for 2-point parametric-linkage to and linkage-disequilibrium (LD, i.e., linkage and association) with PCOS across the following models: dominant completely penetrant (D1), dominant incompletely penetrant (D2), recessive completely penetrant (R1) and recessive incompletely penetrant (R2). The linkage and LD analysis were performed using Pseudomarker [35]. We inferred the presence or absence of LD blocks by calculating the correlation coefficient between variants using the data from the 1000 Genome project (https://www.internationalgenome.org/data-portal/population/TSI). The study was institutionally approved by the Bios Ethical Committee.
Results
We found a total of 5 variants (rs6277, rs60599314, rs112646785, rs4936274, rs4648317) significantly linked to and/or in LD with PCOS (Table 1). Linkage and association (LD) were statistically significant across different inheritance models (Fig. 1). None of the 5 variants had been previously reported with PCOS.

Parametric analysis results of DRD2-risk single nucleotide polymorphisms (SNPs) in polycystic ovarian syndrome (PCOS)
Legend. For each DRD2-risk SNPs in PCOS, we present the − log10(P) as a function of the significant (p < 0.05) test statistics [(Linkage, linkage disequilibrium (LD)|Linkage, LD|NoLinkage] and per inheritance model. D1: dominant, complete penetrance, D2: dominant, incomplete penetrance, R1: recessive, complete penetrance, R2: recessive, incomplete penetrance
In-silico analysis
We analyzed the DRD2-risk variants by different bioinformatics tools that predict their functional role in transcription factor (TF) binding (SNP Function Prediction) [37], miRNA binding (mirSNP) [38], splicing (SpliceAI) [39], and regulatory potential (RegulomeDB) [40]. We found that the risk allele (T) of the variant rs60599314 disrupts the binding of transcription factor AP-2-alpha (Tfap2a) whose upregulation impairs meiosis in mouse oocytes [41].
Discussion
PCOS is a complexly inherited disorder, and variations in several neuroendocrine-related genes were shown to mediate a risk or susceptibility [42]. In this study, we have shown for the first time that the DRD2 gene is also a potential risk gene in PCOS. We have recently reported the implication of DRD2 gene in the risk of T2D and depression in the same multigenerational Italian families under study [30]. The same risk allele (T) of the variant rs112646785 was previously linked to the risk of T2D and MDD comorbidity [30] and now to PCOS, confirming the closely intertwined genetic and phenotypic relationships between these complex disorders [6, 32] as well as the possible pleiotropic role of DRD2. None of the five DRD2-variants reported in our study have been linked before to the risk of PCOS and therefore are novel. Some risk variants in our study, however, have been previously studied and/or reported with PCOS-related phenotypes. The risk allele (A) of the variant rs6277 was associated with obesity [36]. The same variant has also been studied with endometriosis-associated infertility (along with rs4648317) [43] and insulin resistance and T2D [44], but no association has been found. The non-risk allele (C) of this variant rs6277, however, correlated with hyperglycemia in schizophrenic patients, indicating the presence of possible LD with other undetected contributing variants [45].
The roles played by DRD2-risk variants in the pathogenesis of PCOS have yet to be defined. The effect could be mediated by alteration of TF binding as predicted by our in-silico analysis. The risk allele (T) of the variant rs60599314 was predicted to disrupt the binding of transcription factor AP-2-alpha (Tfap2a), whose upregulation impairs meiosis in mouse oocytes [41]. The pathogenic role could also be mediated by alteration of DRD2 properties. The risk allele (A) of the variant rs6277 was previously associated with increased receptor density and affinity in the striatum [46] which if confirmed at the level of the hypothalamus might mediate the DRD2 effects on GnRH pulsatility in PCOS. However, if the DRD2 density and affinity were to be constitutionally present, the effect would not account for the higher peripheral dopamine levels reported in patients with PCOS by a possible DRD2 resistance [17]. Given that elevated blood levels of PRL were found in more than one third of PCOS women [16], and due to our previous findings of PRL contributing to T2D and depression [31], both phenotypes associated with PCOS [6, 32], it is also possible that some DRD2 variants might impair PRL secretion regulation and indirectly impair gonadotropin secretion and reproductive cycles. Given the complex heterogeneity of PCOS, more than one molecular genetic pattern can contribute to it, and PCOS subjects may be predisposed to one molecular distinct entity vs. another, or present with overlapping molecular underpinnings. Therefore, functional studies are still needed to confirm and explain these results. It is also important to replicate the genetic results in other ethnic groups.
Data Availability
The study data are available on reasonable request, and due to lacking specific patients’ consent and privacy restrictions, they are not publicly available.
References
Azziz R, et al. The prevalence and features of the polycystic ovary syndrome in an unselected population. J Clin Endocrinol Metab. 2004;89(6):2745–9.
Azziz R, et al. Positions statement: criteria for defining polycystic ovary syndrome as a predominantly hyperandrogenic syndrome: an androgen excess society guideline. J Clin Endocrinol Metab. 2006;91(11):4237–45.
Brassard M, AinMelk Y, Baillargeon JP. Basic infertility including polycystic ovary syndrome. Med Clin North Am. 2008;92(5):1163–92.
Norman RJ, et al. Polycystic ovary syndrome. Lancet. 2007;370(9588):685–97.
Dunaif A. Hyperandrogenic anovulation (PCOS): a unique disorder of insulin action associated with an increased risk of non-insulin-dependent diabetes mellitus. Am J Med. 1995;98(1A):33S–9.
Kolhe JV, et al. PCOS and depression: common links and potential targets. Reprod Sci. 2022;29(11):3106–23.
Tsutsumi R, Webster NJ. GnRH pulsatility, the pituitary response and reproductive dysfunction. Endocr J. 2009;56(6):729–37.
Khan MJ, Ullah A, Basit S. Genetic basis of polycystic ovary syndrome (PCOS): current perspectives. Appl Clin Genet. 2019;12:249–60.
Legro RS, Strauss JF. Molecular progress in infertility: polycystic ovary syndrome. Fertil Steril. 2002;78(3):569–76.
Rojas J et al. Polycystic ovary syndrome, insulin resistance, and obesity: navigating the pathophysiologic labyrinth Int J Reprod Med, 2014. 2014: p. 719050.
Acevedo-Rodriguez A, et al. Emerging insights into hypothalamic-pituitary-gonadal axis regulation and interaction with stress signalling. J Neuroendocrinol. 2018;30(10):e12590.
Rosenfield RL, Ehrmann DA. The pathogenesis of polycystic ovary syndrome (PCOS): the hypothesis of PCOS as functional ovarian hyperandrogenism revisited. Endocr Rev. 2016;37(5):467–520.
Hernandez I, et al. Hypothalamic dopaminergic tone and prolactin bioactivity in women with polycystic ovary syndrome. Arch Med Res. 2000;31(2):216–22.
Liu X, Herbison AE. Dopamine regulation of gonadotropin-releasing hormone neuron excitability in male and female mice. Endocrinology. 2013;154(1):340–50.
Quigley ME, Rakoff JS, Yen SS. Increased luteinizing hormone sensitivity to dopamine inhibition in polycystic ovary syndrome. J Clin Endocrinol Metab. 1981;52(2):231–4.
Davoudi Z, et al. Prolactin level in polycystic ovary syndrome (PCOS): an approach to the diagnosis and management. Acta Biomed. 2021;92(5):e2021291.
Hamed EA, et al. Nesfatin-1, dopamine, and NADPH levels in Infertile Women with polycystic ovary syndrome: is there a relationship between their levels and metabolic and hormonal variables. J Reprod Infertil. 2022;23(3):160–8.
Leblanc H, et al. Effects of dopamine infusion on pituitary hormone secretion in humans. J Clin Endocrinol Metab. 1976;43(3):668–74.
Polson DW, Mason HD, Franks S. Bromocriptine treatment of women with clomiphene-resistant polycystic ovary syndrome. Clin Endocrinol (Oxf). 1987;26(2):197–203.
Missale C, et al. Dopamine receptors: from structure to function. Physiol Rev. 1998;78(1):189–225.
Fitzgerald P, Dinan TG. Prolactin and dopamine: what is the connection? A review article. J Psychopharmacol. 2008;22(2 Suppl):12–9.
Ciechanowska M, et al. Neuroendocrine regulation of GnRH release and expression of GnRH and GnRH receptor genes in the hypothalamus-pituitary unit in different physiological states. Reprod Biol. 2010;10(2):85–124.
Chaudhari N, Dawalbhakta M, Nampoothiri L. GnRH dysregulation in polycystic ovarian syndrome (PCOS) is a manifestation of an altered neurotransmitter profile. Reprod Biol Endocrinol. 2018;16(1):37.
Cincotta AH, Tozzo E, Scislowski PW. Bromocriptine/SKF38393 treatment ameliorates obesity and associated metabolic dysfunctions in obese (ob/ob) mice. Life Sci. 1997;61(10):951–6.
de Leeuw JE, et al. Pharmacological modulation of dopamine receptor D2-mediated transmission alters the metabolic phenotype of diet induced obese and diet resistant C57Bl6 mice. Exp Diabetes Res. 2011;2011:928523.
Legro RS, Castracane VD, Kauffman RP. Detecting insulin resistance in polycystic ovary syndrome: purposes and pitfalls. Obstet Gynecol Surv. 2004;59(2):141–54.
Vrbikova J, et al. Insulin sensitivity in women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2004;89(6):2942–5.
Gomez R, et al. Evidences for the existence of a low dopaminergic tone in polycystic ovarian syndrome: implications for OHSS development and treatment. J Clin Endocrinol Metab. 2011;96(8):2484–92.
Amin M, Gragnoli C. The prolactin receptor gene (PRLR) is linked and associated with the risk of polycystic ovarian syndrome. Under Revision; 2023.
Amin M et al. Linkage and association of novel DRD2 variants to the comorbidity of type 2 diabetes and depression. Eur Rev Med Pharm Sci, 2023.
Amin M, et al. Novel implication of the prolactin gene (PRL) in the comorbidity of type 2 diabetes and depression. Eur Rev Med Pharm Sci (In Press; 2023.
Livadas S, et al. Polycystic ovary syndrome and type 2 diabetes mellitus: a state-of-the-art review. World J Diabetes. 2022;13(1):5–26.
Rotterdam EA. .-S.P.c.w.g., revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome (PCOS). Hum Reprod. 2004;19(1):41–7.
Purcell S, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81(3):559–75.
Hiekkalinna T, et al. PSEUDOMARKER: a powerful program for joint linkage and/or linkage disequilibrium analysis on mixtures of singletons and related individuals. Human Hered. 2011;71(4):256–66.
Sardahaee FS, et al. Effects of single genetic variants and polygenic obesity risk scores on disordered eating in adolescents - the HUNT study. Appetite. 2017;118:8–16.
Xu Z, Taylor JA. SNPinfo: Integrating GWAS and candidate gene information into functional SNP selection for genetic association studies Nucleic Acids Research, 2009. 37(SUPPL. 2).
Liu C, et al. MirSNP, a database of polymorphisms altering miRNA target sites, identifies miRNA-related SNPs in GWAS SNPs and eQTLs. BMC Genomics. 2012;2012 13(1):1–10.
Jaganathan K, et al. Predicting Splicing from primary sequence with deep learning. Cell. 2019;176(3):535–548e24.
Boyle AP, et al. Annotation of functional variation in personal genomes using RegulomeDB. Genome Res. 2012;22(9):1790–7.
Lin J et al. Overexpression of Tfap2a in mouse oocytes impaired spindle and chromosome Organization. Int J Mol Sci, 2022. 23(22).
Abbott DH, Dumesic DA, Levine JE. Hyperandrogenic origins of polycystic ovary syndrome - implications for pathophysiology and therapy. Expert Rev Endocrinol Metab. 2019;14(2):131–43.
Szczepańska M, et al. Polymorphic variants in the dopamine receptor D2 in women with endometriosis-related infertility. Mol Med Rep. 2015;12(2):3055–60.
Guigas B, et al. Sex-specific effects of naturally occurring variants in the dopamine receptor D2 locus on insulin secretion and type 2 diabetes susceptibility. Diabet Med. 2014;31(8):1001–8.
Lawford BR, et al. Dopamine 2 receptor genes are Associated with raised blood glucose in Schizophrenia. Can J Psychiatry. 2016;61(5):291–7.
Hirvonen MM, et al. C957T polymorphism of dopamine D2 receptor gene affects striatal DRD2 in vivo availability by changing the receptor affinity. Synapse. 2009;63(10):907–12.
Acknowledgements
We thank the families who participated in the study, and we thank Bios Biotech Multi-Diagnostic Health Center, Rome, Italy, for data access and for financial, medical, and laboratory staff support.
Funding
This publication was supported in part with the funds received under Nebraska Laws 2021, LB 380, Sect. 109 awarded to C.G. (PI), Creighton University School of Medicine, through the Nebraska Department of Health & Human Services (DHHS). Its contents represent the views of the authors and do not necessarily represent the official views of the State of Nebraska or DHHS.
Author information
Authors and Affiliations
Contributions
M.A. (https://orcid.org/0000-0003-2876-0784) helped with manuscript drafting and in silico analysis. N.H. (https://orcid.org/0000-0001-7413-0543) drafted the manuscript and helped with literature search. C.G. (https://orcid.org/0000-0002-3873-6617) conceived and performed the study and critically revised the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Families were recruited following the Helsinki declaration guidelines, and individuals provided written informed consent prior to participation. The Bios Ethical Committee approved this study (Prot.PR/Mg/Cg/311708).
Authors’ information
C.G. is Professor of Medicine and Chief of Endocrinology and Endowed Puller Chair at Creighton University School of Medicine, Omaha, NE, and Adjunct Professor of Public Health Sciences, Penn State University College of Medicine, Hershey, PA; N.H. is a medical student at Creighton University School of Medicine, Omaha, NE; M.A. has an MD and a PhD in Genetics (University of Paris) and is a geneticist at Orphanet | INSERM-US14, Paris, France, and member of the executive committee, European Reference Network for the Intellectual Disabilities (ERN-ITHACA) and the International Consortium of Gene Curation Coalition (GenCC).
Competing interests
The authors have declared that they have no conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Amin, M., Horst, N. & Gragnoli, C. Linkage and association of variants in the dopamine receptor 2 gene (DRD2) with polycystic ovary syndrome. J Ovarian Res 16, 158 (2023). https://doi.org/10.1186/s13048-023-01205-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13048-023-01205-2