
Abstract
Background
After diagnosis, glioblastoma (GBM) patients undertake tremendous psychological problems such as anxiety and depression, which may contribute to GBM progression. However, systematic study about the relationship between depression and GBM progression is still lacking.
Methods
Chronic unpredictable mild stress and chronic restrain stress were used to mimic human depression in mice. Human GBM cells and intracranial GBM model were used to assess the effects of chronic stress on GBM growth. Targeted neurotransmitter sequencing, RNA-seq, immunoblotting and immunohistochemistry were used to detect the related molecular mechanism.
Results
Chronic stress promoted GBM progression and up-regulated the level of dopamine (DA) and its receptor type 2 (DRD2) in tumor tissues. Down-regulation or inhibition of DRD2 abolished the promoting effect of chronic stress on GBM progression. Mechanistically, the elevated DA and DRD2 activated ERK1/2 and consequently inhibited GSK3β activity, leading to β-catenin activation. Meanwhile, the activated ERK1/2 up-regulated tyrosine hydroxylase (TH) level in GBM cells and then promoted DA secretion, forming an autocrine positive feedback loop. Remarkably, patients with high-depression exhibited high DRD2 and β-catenin levels, which showed poor prognosis. Additionally, DRD2 specific inhibitor pimozide combined with temozolomide synergistically inhibited GBM growth.
Conclusions
Our study revealed that chronic stress accelerates GBM progression via DRD2/ERK/β-catenin axis and Dopamine/ERK/TH positive feedback loop. DRD2 together with β-catenin may serve as a potential predictive biomarker for worse prognosis as well as therapeutic target of GBM patients with depression.
Similar content being viewed by others

Background
Among malignant brain tumors, glioblastoma (GBM) is the most fatal one featuring with rapid progression, therapeutic resistance and high rate of recurrence [1]. Clinically, patients with GBM routinely experience physical and psychological pressure caused by tumor diagnosis, repeated treatment and tumor recurrence throughout the whole disease course, leading to depression [2]. Epidemiological studies demonstrate that GBM patients suffer from depression at higher rates than the general population, as well as patients with other forms of cancer [3, 4]. Emerging evidence suggests that depression caused by chronic stress could affect tumor progression by influencing the tumor microenvironment, such as metabolic remodeling, immune function and hormone regulation [5,6,7]. However, the relationship between chronic stress and GBM growth remains poorly defined.
It has been reported that neuroactive substances of tumor microenvironment play important roles in supporting progression and malignancy in multiple human cancers [8]. Under chronic stress conditions, the hypothalamic-pituitary-adrenocortical (HPA) axis and the sympathetic nervous system (SNS) are generally activated to stimulate the secretion of stress-related hormones such as glucocorticoids (GC) and adrenocorticotropic hormone (ACTH), as well as stress-related neurotransmitters, such as epinephrine (E), norepinephrine (NE) and dopamine (DA), to cope with harmful and unpredictable stressors [9]. Numerous studies have demonstrated that chronic stress could promote peripheral tumor progression through NE, E and GC, which could be blocked by the respective receptor blockers. These three stress-related factors have also been considered as key regulators to simulate chronic stress environment in vitro [10, 11]. Although β-adrenergic receptor blockers can inhibit cell proliferation, migration, and induce apoptosis in preclinical studies, they failed to prolong the median survival time of glioma patients [12]. Therefore, the critical molecular mechanisms by which chronic stress induced GBM progression have not been elucidated and novel potential molecular targets for GBM treatment may exist.
In this study, we found that depression induced by chronic stress promoted GBM progression and reduced the median survival time of both tumor-bearing nude mice and immunocompetent mice. After chronic stress, DA and its receptor DRD2(Dopamine Receptor D2) significantly increased in GBM tissues, which activated ERK1/2, leading to GSK3β activity inhibition and β-catenin activation. Meanwhile, the activated ERK1/2 up-regulated tyrosine hydroxylase (TH), the rate-limiting enzyme of DA synthesis, and then promoted DA secretion, forming an autocrine positive feedback loop. These factors together contributed to GBM malignant progression. Remarkably, the levels of DRD2 and β-catenin in tumor tissues from GBM patients with depression were significantly upregulated and negatively correlated with the prognosis of patients. Consistently, our findings identified that the combined administration of pimozide (PIMO), a specific inhibitor of DRD2, and temozolomide (TMZ) exert a stronger antitumor action in GBM.
Methods
Cell cultures and reagents
Luciferase-GL261, HEK 293 T and U251 glioma cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). Human U87 GBM cells were purchased from American Type Culture Collection (ATCC). GBM#1 and GBM#2 primary cells were freshly prepared from surgical samples of GBM patients, which were normally expanded and subcultured according to our previous study [13]. All the above cells were maintained in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum (Gibco) in the standard condition of 37 °C, 5% CO2. Sometimes, cells were treated with different concentrations of NE, PIMO or TMZ (all from Target Mol) for 48 h and used for various cell assays.
Antibodies and plasmids
Antibodies specific for β-catenin (1:1000, #8480), Active-β-catenin (1:800, #19,807), c-Myc (1:1000, #5605), Cyclin D1 (1:1000, #2922), TH (1:800, #58,844), GSK3β (1:1000, #12,456), p-GSK3β (1:1000, #5558), ERK (1:1000, #4695), p-ERK (1:1000, #4370), GAPDH (1:1000, #5174), AKT (1:1000, #2920) and p-AKT (1:1000, #4060) were obtained from Cell Signaling. Antibodies against DRD1(1:500, A2893), DRD2(1:500, A12930 or 1:1000, AB5084P), DRD3(1:500, A4587), DRD4(1:500, A1337), DRD5(1:500, A1719) and β-actin (1:1000, AC026) were obtained from ABclonal or Millipore.
Lentiviral constructs of shDRD2#1 and shDRD2#2 was constructed by Shanghai GeneChem. DRD2 over-expression and doxycycline-inducible DRD2 down-regulation (shDRD2i) lentiviral constructs were constructed by OBiO Technology (Shanghai).
Chronic Unpredictable Mild Stress (CUMS) and Chronic Restrain Stress (CRS)
The Institutional Animal Use Committee of Xuzhou Medical University approved all animal experiments (Process number for animal experiments: 202104A455). Five-week-old male BALB/c nude mice and C57BL/6 J mice were obtained from the GemPharmatech. The CUMS and CRS procedures were performed as previously described [5, 14] with certain modifications. For the CUMS procedure, all stressors were shown in Table 1 and each was carried out twice randomly. For the CRS experiments, mice were individually restrained in a modified well-ventilated 50 mL centrifuge tube and unable to move freely for 2 h every day. Mice were stressed for 4 weeks (CUMS) or 3 weeks (CRS) before tumor transplantation, and continued to be stressed daily until tumor-bearing mice were sacrificed.
Depression behavioral tests
For sucrose preference test (SPT), mice were individually housed in cages with a two-bottles procedure, during which mice can freely access to both water and a 2% sucrose solution. The sucrose preference was calculated as a percentage of the consumed sucrose solution relative to the total amount of liquid intake. Reduced sucrose preference in mice is considered as an anhedonia response, which is suggestive of depression [15].
For tail suspension test (TST), mouse was hung upside down 30 cm above the ground by fixing the tip of the tail. The test lasted for 6 min, and the immobility time, defined as the absence of escape-directed behavior, was measured at last 5 min.
For forced swimming test (FPT), each mouse was placed into a cylinder (height: 25 cm; diameter:18 cm) filled with water (22 °C) to a depth of 15 cm. Following 1 min period of adaptation in water, the immobility duration was recorded during the last 5 min of the total 6-min testing period. The immobility time was defined as absence of any movement except for only slight movements to keep the head above the water surface.
Orthotopic glioma-bearing mouse model
In general, after having been stressed for 4 weeks (CUMS) or 3 weeks (CRS), BALB/c nude mice or C57BL/6 J mice were intracranially injected with luciferase-labelled U87 or GBM#2 (5 × 105) or GL261 glioma cells (2 × 105) respectively. Thereafter, the intracranial tumor growth was detected by bioluminescence imaging on day 7, 14, 21 and 28 after tumor transplantation. Mice were sacrificed when neurological symptoms, such as hemiplegia, listlessness, cachexia, were observed. The median survival times were analyzed using Kaplan–Meier curves. For the doxycycline induced shDRD2 GBM#2 cell mouse model, beginning at day 7 after tumor transplantation, mice were fed with doxycycline (2 mg/ml, 5 consecutive days a week) in drinking water. For the PIMO and TMZ co-administration experiment, beginning at the day 7 after tumor transplantation, mice were administered with PIMO (10 mg/kg) by oral gavage or TMZ (7.5 mg/kg) intraperitoneally alone or together with PIMO (10 mg/kg) 5 days on, 2 days off for 3 weeks.
Enzyme-linked immunosorbent assay (ELISA)
ELISA was performed to measure the level of ACTH, corticosterone, DA and NE from tumor tissues and plasma of mice, according to the manufacturers’ instructions.
Targeted neurotransmitter sequencing
Tumor tissues and plasma were obtained from both depression and control mice and immediately stored in liquid nitrogen for targeted neurotransmitter sequencing. The relative quantification of neurotransmitters was examined by LC–MS/MS analysis at Zhongke New Life Biotech.
RNA sequencing (RNA-seq) and gene set enrichment analysis (GSEA)
Total RNA was extracted from tumor tissues and submitted for RNA-seq at BGI Genomics. For data analysis, genes with |log2FC|> 1.0 and adjusted P values (Q values) < 0.05 were selected as differentially expressed genes (DEGs) using the Dr. Tom online software (BGI Genomics). GSEA was performed online (http://software.broadinstitute.org/gsea/).
Quantitative PCR and primers
Quantitative RT-PCR was performed with SuperReal PreMix Plus (Tiangen) according to the manufacturer’s instructions. The relative mRNA expression levels of genes were normalized to the GAPDH internal control and measured with the 2−ΔΔCt method. All primer sequences were synthesized by Sangon Biotech. as follows:
-
DRD1-F, 5′-TGTGACACGAGGTTGAGC-3′;
-
DRD1-R, 5′-GGTGGTCTGGCAGTTCTT-3′;
-
DRD2-F, 5′-CCCAATGGATCCACTGAACCTG-3′;
-
DRD2-R, 5′- AATCAAGGTGTGCTCCGCTACTG-3′;
-
β-actin-F, 5′-TATAAAACCCGGCGGCGCA-3′;
-
β-actin-R,5′-TCATCCATGGCGAACTGGTG-3′.
Western blotting, CCK8, colony formation, EdU and Immunofluoresence
The above methods were performed as described previously [16].
Patient depression scale
A total of 76 glioma patients were studied at the Affiliated Hospital of Xuzhou Medical University. To assess the symptoms of depression, we combined the Patient Health Questionnaire-9 (PHQ-9, self-rating) depression scale and the Hamilton Depression Rating Scale (HDRS, his-rating) [17] to establish a rating scale 3 days before surgery.
As we know, the PHQ-9 has been identified as the most reliable screening tool according to the international guidelines. It is a self-administered questionnaire aimed at surveying the nine main criteria for major depression [18, 19]. The PHQ-9 is scored from 0 to 27 (5–9 means mild depression, 10–14 means moderate depression, 15–19 means major depression, and 20–27 means severe depression). The Hamilton rating scale for depression is considered as the gold standard for the assessment of major depressive disorder. It is a clinician-rated interview which can better reflect the severity of depressive symptoms and the results are more objective. We used the 17-item HDRS to assess the severity of the depression in this study [20]. Mild depression was defined by scores ranging 8–17, moderate depression with scores ranging 18–24, and serious depression with scores > 25 [21, 22]. Based on the actual situation of glioma patients, our work mainly refers to the HDRS results, and the PHQ-9 scale as an auxiliary use.
This study was approved by the Ethics Committee of the Affiliated Hospital of Xuzhou Medical University (Process number for human experiments: XYFY2022-KL414-01). All the patients were provided informed consents.
Fresh glioma specimens, glioma tissue microarray (TMA) and immunohistochemistry
The 23 fresh glioma samples for western blotting and 45 glioma specimens to construct glioma TMA for immunohistochemical analysis were obtained from the Affiliated Hospital of Xuzhou Medical University. Based on the HAMD and PHQ-9 scale, these total 68 samples included 12 patients without depression, 35 patients with mild depression and 21 patients with severe depression. The glioma TMA construction and immunohistochemical analysis were performed as described in our previous study [23].
Statistical analysis
All experiments were repeated independently at least 3 replicates. SPSS16.0 was applied to analyze the data and the results were presented as mean ± SD. P values were calculated by using the two-tailed t test for comparisons between independent samples and the One-way analysis of variance (ANOVA) for multiple-group analysis followed by Tukey’s test. Chi-square test was used to compare the counting data and overall survival (OS) was determined using Kaplan–Meier curves and log-rank tests. Differences with P < 0.05 were considered statistically significant.
Results
Chronic stress promotes GBM progression and shortens survival time of tumor-bearing mice
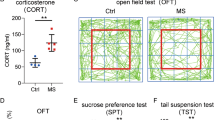
To investigate the potential effects of chronic stress on glioma growth, we took advantage of CUMS and CRS, two well-established stress paradigms, to mimic human depression in nude mice (Fig. 1A). As shown in Fig. 1B and 1C, the stressed mice exhibited more body weight loss, apparent decrease in SPT and longer immobility time in TST under two paradigms. In addition, the levels of corticosterone and ACTH, two classical stress-related hormones, were significantly upregulated after chronic stress both in tumor tissues and plasma of glioma-bearing mice (Fig. 1D). The above results demonstrate the successful construction of the model for mice depression.

Chronic stress promotes glioma progression and shortens the survival of nude mice bearing xenografts. A Diagram of the experimental design for nude mice under CUMS or CRS procedure. B&C Body weight (left) and behavioral tests (right) were used to evaluate the depression of mice after CUMS and CRS. D Levels of corticosterone and ACTH in plasma and glioma tissues were measured by ELISA. E&F Representative bioluminescence images (E) and the photon flux was analyzed quantitatively (F) in both models using U87 glioma cells. G Representative H&E staining of consecutive brain sections (left) and the quantified results (right). H Ki67 immunostaining (left) and positive cell quantification (right). I Kaplan–Meier curve showed the overall survival time of glioma-bearing mice. CUMS: chronic unpredictable mild stress. CRS: chronic restraint stress. SPT: sucrose preference test. TST: tail suspension test. *P < 0.05, **P < 0.01, ***P < 0.001
Next, we constructed the orthotopic GBM xenograft model in above mice by using U87 glioma cells and examined by in vivo bioluminescence imaging. As shown in Fig. 1E&1F, the tumors grew up dramatically as time increase and those in stressed mice grew faster than those in control mice. Consistently, data from H&E and Ki67 staining also showed that chronic stress promoted the xenograft growth (Fig. 1G,1H). Besides, chronic stress shortens the survival time of glioma-bearing mice (Fig. 1I).
To rule out that the above phenomenon was attributed to specific mouse strain, we repeated the above experiments in C57BL/6 J mice using GL261 glioma cells and obtained similar results (sFig.1). In addition, the behavioral tests at one week after tumor inoculation showed that the depression effect was induced by CUMS or CRS, but not by glioma cell inoculation in situ (sFig.2). These results suggest that chronic stress was of great importance for glioma progression, which needs to be studied further in detail.
Dopamine and DRD2 in glioma tissues are significantly enhanced after chronic stress
Studies have demonstrated that neurotransmitters in tumor microenvironment also act as regulators of tumor growth [24]. To elucidate the mechanistic link between chronic stress and glioma growth, we employed targeted metabolomics to detect the changes of 15 common neurotransmitters from the tumor tissues and plasma of mice after chronic stress. As presented in Fig. 2A-2D, majority of these neurotransmitters increased, among which alterations of DA metabolism-associated neurotransmitters such as levodopa (DOPA), 3-methoxytyramine, DA and NE were more prominent. Strikingly, the DA level in tumor tissues of depression mice was about 14 times higher than those of control mice and was the highest increased one among all examined neurotransmitters. Consistently, examined by ELISA, the DA levels, as well as those of NE, increased significantly after chronic stress (Fig. 2E, 2F). Interestingly, the increase of DA was limited in tumor tissues and the plasma DA level showed no change after chronic stress (Fig. 2E,2F). Furthermore, chronic stress upregulated the levels of TH, the rate-limiting enzyme of DA biosynthesis (Fig. 2G,2H), suggesting that the DA generation and secretion by tumor cells increased locally after chronic stress indeed.

Dopamine and DRD2 levels significantly increased after chronic stress. A-D Heatmap of targeted metabolomic alterations in plasma (A) and glioma tissues (B) after chronic stress and the quantification results (C&D). E&F Levels of two stress-related neurotransmitters (DA and NE) in plasma (E) and glioma tissues (F) were measured by ELISA. G&H Immunoblotting (G) and immunofluorescence staining (H) of TH in glioma tissues with or without chronic stress. Scale bar: 50 μm. I Clustergram of RNA-seq data from tumor tissues of mice. J KEGG analysis of DEGs of RNA-seq data. K Five key genes in dopamine metabolic pathway were significantly upregulated after chronic stress. L The mRNA levels of DRD1 and DRD2 in glioma xenografts. M&N The protein levels of DRD1-5 in xenografts (M) and the quantified results (N). Representative WB of three independent experiment is shown. TH: tyrosine hydroxylase. *P < 0.05, **P < 0.01, ***P < 0.001
To perform its function, DA needs to bind to its D1-like (DRD1/DRD5) or D2-like (DRD2-DRD4) receptors. To determine which DA receptor mediated the effects of chronic stress on glioma progression, we conducted high-throughput transcriptome sequencing in tumor tissues and found that 2591 genes were up-regulated and 402 genes were down-regulated with |Log2 FC|≥ 1 and Q-value < 0.05 (Fig. 2I). Take advantage of the Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis, we found that the neuroactive ligand-receptor interaction pathway was the most significant enrichment of DEGs (Fig. 2J). Among DEGs in this pathway, DRD1, DRD2, GRIA3, GRIA4 and GRIN2A were the key genes in dopamine metabolic pathways, out of which the increase in DRD1 and DRD2 levels became pronounced after chronic stress (Fig. 2K). Examined by qPCR, the mRNA levels of DRD2, but not DRD1, were dramatically upregulated after chronic stress (Fig. 2L). Similarly, the protein level of DRD2, but not those of any other four dopamine receptors, increased after chronic stress (Fig. 2M,2N). These data suggest that DRD2 may be the candidate mediator of chronic stress-induced glioma progression.
DRD2 mediates the promoting effects on glioma progression induced by chronic stress
Next, we generated stable DRD2 down-regulation and over-expression glioma cells (sFig.3A) and found that DRD2 down-regulation significantly inhibited glioma cell growth, while DRD2 over-expression significantly promoted the aforementioned effect in both U251, GBM#2 and U87 cells (Fig. 3A-3D, sFig.3B-3F). Notably, shDRD2#1 who exhibited poor down-regulation effect could not inhibit the glioma cell growth (Fig. 3A-3D, sFig.3B-3F), indicating the specific inhibitory effect of DRD2 down-regulation on glioma cell growth.

DRD2 is required for glioma progression induced by chronic stress. A-D Cell proliferation was measured using CCK-8 (A&B) and EdU assay (C&D) in glioma cells over-expressing or knocking-down DRD2.Scale bar: 100 µm. E–H. Representative images and quantitative results of CCK-8 (E&F) and EdU assay (G&H) of glioma cells knocking-down DRD2 with or without NE (20 μM) treatment. I Diagram of in vivo experimental schedule utilizing GBM#2 glioma cells. J&K Time-lapse bioluminescence imaging of mice (J) and the photon flux was analyzed (K). L Kaplan–Meier curve showed the overall survival time of glioma-bearing mice of each group. NE: norepinephrine. *P < 0.05, **P < 0.01, ***P < 0.001
As a crucial stress-related neurotransmitter, NE is involved in the promoting effect of tumor growth caused by chronic stress and also widely used to mimic chronic stress in in vitro experiment [25]. As expected, the proliferation of four glioma cell lines increased significantly after NE treatment (sFig.4A). In addition, down-regulation of DRD2 significantly suppressed the promotion effects of NE in both U251 and GBM#2 cells (Fig. 3E-3H and sFig.4B). Furthermore, we confirmed the above observations in U87 cells (sFig.3G-3I). Similarly, the specific DRD2 antagonist PIMO abolished the promoting effects induced by NE treatment (sFig.4C-4E). Therefore, DRD2 knockdown or inhibition largely abolished the promoting effect of NE on glioma proliferation in vitro.
Next, since DRD2 down-regulated cells generated by shDRD2 in Fig. 3A grew extremely slow, we performed the above experiment in vivo by conducting the xenograft tumor model with doxycycline-inducible DRD2 knockdown (shDRD2i) GBM#2 cells (sFig.4F,4G) together with CRS treatment (Fig. 3I). The results showed that doxycycline-induced DRD2 depletion not only significantly inhibited tumor growth but also blocked the promoting effect of tumor growth induced by chronic stress (Fig. 3J,3K). Consistently, DRD2 depletion extended the survival time of tumor-bearing chronic stressed mice (Fig. 3L). Taken together, both the in vitro and in vivo results indicate that DRD2 indeed mediated the promoting effect of chronic stress on glioma progression.
Chronic stress activates Wnt/β-catenin signaling via DRD2
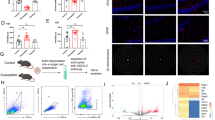
To further explore the molecular mechanism responsible for DRD2-mediated tumor progression, data of 154 primary GBM patients from the TCGA database were utilized to perform GSEA. The results showed that Wnt/β-catenin pathway gene set was enriched significantly in high DRD2 expression group (Fig. 4A). Meanwhile, the GO enrichment analysis from our RNA-seq data also showed that DEGs were significantly enriched in Wnt/β-catenin pathway (sFig.5A). These results sparked our interest in determining the relationship between DRD2 and Wnt/β-catenin pathway in our system. We found that, except for DRD2, the levels of key proteins in Wnt/β-catenin signaling, such as p-GSK3β, Active-β-catenin, β-catenin and its downstream target cyclin D1 were upregulated after chronic stress (Fig. 4B). In addition, β-catenin was translocated into the nucleus after chronic stress (Fig. 4C).

Chronic stress activates DRD2/ERK/β-catenin axis and Dopamine/ERK/TH regulatory loop. A GSEA was performed using TCGA dataset. B Lysates of glioma tissues after chronic stress were examined by immunoblotting. C Immunofluorescence staining of DRD2 and β-catenin in glioma tissues with or without chronic stress. Scale bar: 50 μm. D Lysates of DRD2 down-regulation or over-expression glioma cells were examined by immunoblotting. E Immunoblotting analysis of DRD2 down-regulated U87 and U251 glioma cells treated with NE. F FpClass analysis predicted that ERK was the protein simultaneously interacting with β-catenin, GSK3β and TH. G Lysates of glioma tissues with or without chronic stress were examined by immunoblotting. H Lysates of DRD2 knockdown U87 and U251 glioma cells treated with both NE and U0126 simultaneously were examined by immunoblotting. WB experiments were independently replicated three times. Act-β-catenin: Active-β-catenin
Furthermore, knockdown of DRD2 by shDRD2#2 markedly inhibited the levels of p-GSK3β, Active-β-catenin, β-catenin and its target protein cyclin D1, while over-expression of DRD2 exerted an opposite effect (Fig. 4D). Importantly, the level of p-GSK3β, Active-β-catenin and p-β-catenin was gradually elevated with NE increase in both U251 and U87 glioma cells (sFig.5B). Conversely, they significantly decreased upon PIMO increase (sFig.5C). Strikingly, PIMO treatment abolished NE-induced upregulation of the above proteins (sFig.5D). Consistently, DRD2 knockdown also abolished the NE-induced protein upregulation of Wnt/β-catenin pathway in both glioma cells (Fig. 4E). Together, the above results indicated that chronic stress activates Wnt/β-catenin signaling via DRD2.
ERK is the core mediator between DRD2 and Wnt/β-catenin signaling
We next assessed which is the mediator between DRD2 and Wnt/β-catenin signaling. By utilizing the bioinformatics tool FpClass (http://dcv.uhnres.utoronto.ca/FPCLASS/home/), we identified that ERK was the highest scoring protein simultaneously interacting with GSK3β, β-catenin and TH (Fig. 4F). In addition, the p-ERK1/2 was up-regulated after chronic stress (Fig. 4G), NE treatment (sFig. 5E), and DRD2 over-expression (Fig. 4D). Interestingly, TH level was also up-regulated after NE treatment (sFig. 5E). Furthermore, the ERK inhibitor U0126 treatment decreased the p-ERK1/2, p-GSK3β, β-catenin, as well as TH levels (sFig.5F). However, TH inhibitor metyrosine could not decrease p-ERK1/2 level under either basal (sFig. 5G) or stress condition (sFig. 5E), indicating that TH was the downstream of ERK1/2. Therefore, we conclude that ERK act as the common upstream factor for TH and β-catenin. Importantly, either U0126 treatment or DRD2 down-regulation could abolish NE-induced ERK, TH and Wnt/β-catenin activation (Fig. 4H), suggesting that DRD2-ERK mediates the facilitatory influence on the Wnt/β-catenin pathway and TH after chronic stress. Our findings that chronic stress up-regulated the levels of DA, DRD2 and TH, and U0126 treatment abolished NE-induced TH activation means a Dopamine/ERK/TH positive regulatory loop after chronic stress.
As AKT and ERK are both considered to be classical upstream kinases of GSK3β in mammals and thus able to result in β-catenin accumulation and nuclear translocation indirectly [26], we also examined the level of p-AKT(T308) and p-AKT(S473), two main phosphorylation sites of AKT. However, neither total nor phosphorylation levels of AKT showed any changes, which suggests that chronic stress may activate ERK but not AKT signaling (Fig. 4B,4D).
DRD2 combined with β-catenin act as an indicator for poor prognosis in GBM patients with depression
Since the above results revealed that chronic stress promoted glioma growth, we wondered whether depression caused by chronic stress in glioma patients affected tumor progression and prognosis in the clinic. Firstly, we assessed depressive grade of glioma patients using PHQ-9 and HAMD scales, which were widely recognized and used in depression measurements. As presented in Table 2, the relationships between depression and clinicopathological features in 76 glioma patients were analyzed. No correlations were found between the interactions of age, sex, treatment interventions and depression. However, it is noteworthy that depression itself and depression grade both significantly correlated with glioma grades, as well as patient outcome. Additionally, the grade of glioma increased with depression severity aggravated (Fig. 5A). In parallel, highly depressed glioma patients appeared a worse prognosis than those with mild depression (Fig. 5B).

DRD2 combined with β-catenin act as an indicator for poor prognosis in GBM patients with depression. A Hamilton scores were gradually upregulated as the advanced malignancy of gliomas. B High-depression GBM patients had a worse prognosis than the patients with low-depression. C&D Glioma lysates from patients with or without depression were subjected to immunoblotting (C) and quantified (D). Representative WB of three independent experiment is shown. E–H Representative images of immunohistochemical staining (E) and percentage (F–H) of DRD2 and β-catenin positive cells in glioma patients with or without depression. Scale bar: 100 μm. I-K Kaplan–Meier survival analysis of glioma patients with or without depression expressing indicated proteins. LGG: low grade glioma. HGG: high grade glioma. *P < 0.05, **P < 0.01, ***P < 0.001
Motivated by the above phenomena, we next explored the expression levels of DRD2 and β-catenin in glioma patients with or without depression. As shown in Fig. 5C and 5D, levels of both proteins were elevated in glioma patients with depression and the more severe of depression, the higher level of both proteins. Consistently, examined by immunohistochemistry in tissue microarray, both the percentage of DRD2 and β-catenin positive cells increased with the depression level increase (Fig. 5E-H). In addition, the follow-up results exhibited that glioma patients with higher DRD2 expression showed poorer prognosis than those with lower DRD2 expression, while this was not the case for β-catenin (Fig. 5I,5J). More importantly, glioma patients with concomitantly high DRD2 and β-catenin have shorter overall survival than patients with concomitantly low expression (Fig. 5K), suggesting that the combined detection of DRD2 and β-catenin could be a key outcome measure in glioma patients with depression.
Combination treatment of PIMO and TMZ induces a synergistic anti-glioma effect both in vitro and in vivo
In the foregoing studies, we found that DRD2 antagonist PIMO inhibits glioma cell proliferation in vitro (sFig.4). Considering that PIMO could penetrate the BBB, we wondered whether it would also have tumor-suppressive effects in vivo and synergize with the first-line anti-glioma drug TMZ. As illustrated in Fig. 6A-6D, cell proliferation was significantly inhibited by PIMO, and further inhibited by the combination treatment with different concentration of TMZ. Furthermore, the combination index (CI) of PIMO and TMZ was assessed by Compusyn software. As shown in Table 3, the CI values for both U87 and U251 cells were considerably < 1, indicating a synergism of the inhibition effect.

Combination treatment of PIMO and TMZ induces a synergistic anti-glioma effect both in vitro and in vivo. A-D Relative proliferation of U251 and U87 cells treated with PIMO (5 μM or 10 μM) and indicated doses of TMZ were measured by CCK-8 assay. E Diagram of in vivo experimental schedule. F&G Representative bioluminescence images of intracranial xenografts using GBM#2 glioma cells with different drug administration on the indicated days (F) and quantitative analysis of the fluorescence index (G). H Kaplan–Meier analysis of the median survival time of nude mice. I Schematic model of how chronic stress promotes glioma growth. PIMO: pimozide. TMZ: Temozolomide. *P < 0.05, **P < 0.01, ***P < 0.001
Next, we conducted the above experiments in glioma-bearing mice with or without depression (Fig. 6E). The results showed that either TMZ or PIMO treatment alone significantly delayed tumor growth, while the combined treatment exhibited the most obvious inhibition effects in both control and stress groups (Figs. 6F,G). Consistently, PIMO in combination with TMZ significantly extended the median overall survival of glioma-bearing nude mice compared to either agent alone (Fig. 6H). To rule out the possibility that these results were due to a specific intracranial tumor mouse model, we conducted this in vivo experiment of combination therapy in a murine-derived tumor model using CRS paradigm and obtained consistent results (sFig.6A-6C).
Discussion
Accumulating evidence has shown that chronic stress potentiated peripheral tumor development [10, 14]. However, studies about the relationship between chronic stress and glioma progression is mainly focused on the correlation of depression and clinicopathological features of glioma patients [9, 27]. In this study, we found that chronic stress promotes glioma malignant progression and identified DRD2 as the novel regulator. The schematic model to summarize our findings was presented in Fig. 6I. In 2020, Wang et al. reported that psychological stress promotes proliferation and invasiveness of glioma cells [28], in line with our study. However, Lopes et al. very recently reported that chronic stress does not influence the survival of mouse models of glioblastoma [29]. This difference may be caused by different stress exposure time. Lopes et al. just stressed the mice before the orthotopic injection of GBM cells and stopped the stress procedure after tumor cell injection. During the long unstressed period, the stress effect may be diminished, leading to no change of the animals' overall survival (OS). In our system, the chronic stress lasted for the whole in vivo experiment window, which is closer to the clinic stress feature of patients, and shortened the OS of mice.
By binding to its different receptors, DA showed promotion or inhibition effect on tumor growth [30]. Among five DA receptors, DRD2 is the most frequently studied one. Previous work has revealed that high expression of DRD2 mRNA and protein was observed in glioma tissues [31] and DRD2 contributes to the spheroid formation behavior of U87 glioma cells, promotes glioma cell proliferation and maintains the self-renewal of glioma-initiation cells [32,33,34]. Recently Liu et al., reported that psychological stress drives progression of melanoma and breast cancer via DRD2/HIF-1α signaling [35]. They found that DRD2 interacted with von Hippel-Lindau (VHL) in the nucleus, and competitive binding of DRD2 and HIF-1α to VHL resulted in reduced ubiquitination-mediated degradation of HIF-1α, enhancing the epithelial-mesenchymal transition of tumor cells. In this study, we identified a new signaling regulatory mechanism, namely the DRD2/ERK/β-catenin pathway and Dopamine/ERK/TH regulatory loop, which was activated by chronic stress and in turn promoted tumor progression.
Except for that glioma patients with high depression tended to have a worse prognosis than those with less depression, we further found that DRD2 and β-catenin were upregulated in glioma tissues from patients with depression. Furthermore, the higher expression of them, the poorer prognosis of the patients. Therefore, DRD2 and β-catenin serve as crucial factors mediated glioma progression caused by chronic stress and the combined detection of DRD2 and β-catenin may be a key way for predicting prognosis of glioma patients with depression. In addition, combination of TMZ with PIMO exhibited significant synergism on the suppression of glioma growth both in vitro and in vivo, suggesting that PIMO may potentially serve as combination therapy with TMZ for glioma treatment. However, the mechanism of their joint action is not clear and needs further study.
Conclusion
In this study, we revealed that DRD2/ERK/β-catenin pathway and Dopamine/ERK/TH regulatory loop jointly mediate the promoting effect of chronic stress on glioma malignant progression. Therefore, for glioma patients with high psychological depression, personalized and precise molecular targeted therapy targeting DRD2 and TMZ may be an effective treatment.
Availability of data and material
All data and material during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ACTH:
-
Adrenocorticotropic Hormone
- AKT:
-
Protein kinase B
- ATCC:
-
American Type Culture Collection
- BBB:
-
Blood Brain Barriers
- CI:
-
Combination Index
- CRS:
-
Chronic Restrain Stress
- CUMS:
-
Chronic Unpredictable Mild Stress
- DA:
-
Dopamine
- DEG:
-
Differentially Expressed Genes
- DRD1:
-
Dopamine Receptor D1
- DRD2:
-
Dopamine Receptor D2
- DRD3:
-
Dopamine Receptor D3
- DRD4:
-
Dopamine Receptor D4
- DRD5:
-
Dopamine Receptor D5
- E:
-
Epinephrine
- ELISA:
-
Enzyme-Linked Immunosorbent Assay
- ERK:
-
Extracellular Regulated protein Kinases
- FST:
-
Forced Swimming Test
- GBM:
-
Glioblastoma Multiforme
- GC:
-
Glucocorticoid
- GSEA:
-
Gene Set Enrichment Analysis
- GSK3β:
-
Glycogen Synthase Kinase-3 Beta
- HDRS:
-
Hamilton Depression Rating Scale
- HE:
-
Haematoxylin and Eosin
- HIF-1α:
-
Hypoxia-Inducible Factor 1-Alpha
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- NE:
-
Norepinephrine
- OS:
-
Overall Survival
- PHQ-9:
-
Patient Health Questionnaire-9
- PIMO:
-
Pimozide
- qPCR:
-
Quantitative real-time Polymerase chain reaction
- RNA-seq:
-
RNA sequencing
- SNS:
-
Sympathetic Nervous System
- SPT:
-
Sucrose Preference Test
- TGCA:
-
The Cancer Genome Atlas
- TH:
-
Tyrosine Hydroxylase
- TMA:
-
Tissue Microarray
- TMZ:
-
Temozolomide
- TST:
-
Tail Suspension Test
- VHL:
-
Von Hippel-Lindau
References
Tan AC, Ashley DM, Lopez GY, Malinzak M, Friedman HS, Khasraw M. Management of glioblastoma: State of the art and future directions. CA Cancer J Clin. 2020;70:299–312.
Bortolato B, Hyphantis TN, Valpione S, Perini G, Maes M, Morris G, Kubera M, Kohler CA, Fernandes BS, Stubbs B, Pavlidis N, Carvalho AF. Depression in cancer: The many biobehavioral pathways driving tumor progression. Cancer Treat Rev. 2017;52:58–70.
Wen Y, Zhang Y, Li J, Luo F, Huang Z, Liu K. Low concentration trifluoperazine promotes proliferation and reduces calcium-dependent apoptosis in glioma cells. Sci Rep. 2018;8:1147.
A.M. Bielecka, E. Obuchowicz, Antidepressant drugs can modify cytotoxic action of temozolomide, Eur J Cancer Care (Engl), 26 (2017).
Thaker PH, Han LY, Kamat AA, Arevalo JM, Takahashi R, Lu C, Jennings NB, Armaiz-Pena G, Bankson JA, Ravoori M, Merritt WM, Lin YG, Mangala LS, Kim TJ, Coleman RL, Landen CN, Li Y, Felix E, Sanguino AM, Newman RA, Lloyd M, Gershenson DM, Kundra V, Lopez-Berestein G, Lutgendorf SK, Cole SW, Sood AK. Chronic stress promotes tumor growth and angiogenesis in a mouse model of ovarian carcinoma. Nat Med. 2006;12:939–44.
Cheng Y, Gao XH, Li XJ, Cao QH, Zhao DD, Zhou JR, Wu HX, Wang Y, You LJ, Yang HB, He YL, Li YR, Bian JS, Zhu QY, Birnbaumer L, Yang Y. Depression promotes prostate cancer invasion and metastasis via a sympathetic-cAMP-FAK signaling pathway. Oncogene. 2018;37:2953–66.
Yang H, Xia L, Chen J, Zhang S, Martin V, Li Q, Lin S, Chen J, Calmette J, Lu M, Fu L, Yang J, Pan Z, Yu K, He J, Morand E, Schlecht-Louf G, Krzysiek R, Zitvogel L, Kang B, Zhang Z, Leader A, Zhou P, Lanfumey L, Shi M, Kroemer G, Ma Y. Stress-glucocorticoid-TSC22D3 axis compromises therapy-induced antitumor immunity. Nat Med. 2019;25:1428–41.
Jiang SH, Hu LP, Wang X, Li J, Zhang ZG. Neurotransmitters: emerging targets in cancer. Oncogene. 2020;39:503–15.
Eckerling A, Ricon-Becker I, Sorski L, Sandbank E, Ben-Eliyahu S. Stress and cancer: mechanisms, significance and future directions. Nat Rev Cancer. 2021;21:767–85.
Cui B, Luo Y, Tian P, Peng F, Lu J, Yang Y, Su Q, Liu B, Yu J, Luo X, Yin L, Cheng W, An F, He B, Liang D, Wu S, Chu P, Song L, Liu X, Luo H, Xu J, Pan Y, Wang Y, Li D, Huang P, Yang Q, Zhang L, Zhou BP, Liu S, Xu G, Lam EW, Kelley KW, Liu Q. Stress-induced epinephrine enhances lactate dehydrogenase A and promotes breast cancer stem-like cells. J Clin Invest. 2019;129:1030–46.
Tian W, Liu Y, Cao C, Zeng Y, Pan Y, Liu X, Peng Y, Wu F. Chronic Stress: Impacts on Tumor Microenvironment and Implications for Anti-Cancer Treatments. Front Cell Dev Biol. 2021;9: 777018.
Tewarie IA, Senders JT, Hulsbergen AFC, Kremer S, Broekman MLD. Beta-blockers and glioma: a systematic review of preclinical studies and clinical results. Neurosurg Rev. 2021;44:669–77.
Wang X, Wang Z, Zhang Y, Wang Y, Zhang H, Xie S, Xie P, Yu R, Zhou X. Golgi phosphoprotein 3 sensitizes the tumour suppression effect of gefitinib on gliomas. Cell Prolif. 2019;52: e12636.
Cheng Y, Tang XY, Li YX, Zhao DD, Cao QH, Wu HX, Yang HB, Hao K, Yang Y. Depression-Induced Neuropeptide Y Secretion Promotes Prostate Cancer Growth by Recruiting Myeloid Cells. Clin Cancer Res. 2019;25:2621–32.
You J, Sun L, Wang J, Sun F, Wang W, Wang D, Fan X, Liu D, Xu Z, Qiu C, Chen J, Yan H, Liu B. Role of Adiponectin-Notch pathway in cognitive dysfunction associated with depression and in the therapeutic effect of physical exercise. Aging Cell. 2021;20: e13387.
Wang Y, Pan P, Wang Z, Zhang Y, Xie P, Geng D, Jiang Y, Yu R, Zhou X. beta-catenin-mediated YAP signaling promotes human glioma growth. J Exp Clin Cancer Res. 2017;36:136.
S. Ijaz, P. Davies, C.J. Williams, D. Kessler, G. Lewis, N. Wiles, Psychological therapies for treatment-resistant depression in adults, Cochrane Database Syst Rev, 5 (2018) CD010558.
Accuracy of Patient Health Questionnaire-9 (PHQ-9) for screening to detect major depression: individual participant data meta-analysis, BMJ, 365 (2019) l1781.
L. Costantini, C. Pasquarella, A. Odone, M.E. Colucci, A. Costanza, G. Serafini, A. Aguglia, M. Belvederi Murri, V. Brakoulias, M. Amore, S.N. Ghaemi, A. Amerio, Screening for depression in primary care with Patient Health Questionnaire-9 (PHQ-9): A systematic review, J Affect Disord, 279 (2021) 473–483.
Rohan KJ, Rough JN, Evans M, Ho SY, Meyerhoff J, Roberts LM, Vacek PM. A protocol for the Hamilton Rating Scale for Depression: Item scoring rules, Rater training, and outcome accuracy with data on its application in a clinical trial. J Affect Disord. 2016;200:111–8.
Etkin A, Patenaude B, Song YJ, Usherwood T, Rekshan W, Schatzberg AF, Rush AJ, Williams LM. A cognitive-emotional biomarker for predicting remission with antidepressant medications: a report from the iSPOT-D trial. Neuropsychopharmacology. 2015;40:1332–42.
Kubo H, Setoyama D, Watabe M, Ohgidani M, Hayakawa K, Kuwano N, Sato-Kasai M, Katsuki R, Kanba S, Kang D, Kato TA. Plasma acetylcholine and nicotinic acid are correlated with focused preference for photographed females in depressed males: an economic game study. Sci Rep. 2021;11:2199.
Y. Wang, K. Wang, J. Fu, Y. Zhang, Y. Mao, X. Wang, X. Wang, R. Yu, X. Zhou, FRK inhibits glioblastoma progression via phosphorylating YAP and inducing its ubiquitylation and degradation by Siah1, Neuro Oncol, (2022).
Gao J, Gao Y, Lin S, Zou X, Zhu Y, Chen X, Wan H, Zhu H. Effects of activating GABAB1 receptor on proliferation, migration, invasion and epithelial-mesenchymal transition of ovarian cancer cells. J Ovarian Res. 2020;13:126.
Du P, Zeng H, Xiao Y, Zhao Y, Zheng B, Deng Y, Liu J, Huang B, Zhang X, Yang K, Jiang Y, Ma X. Chronic stress promotes EMT-mediated metastasis through activation of STAT3 signaling pathway by miR-337-3p in breast cancer. Cell Death Dis. 2020;11:761.
Zhang B, Zhang M, Li Q, Yang Y, Shang Z, Luo J. TPX2 mediates prostate cancer epithelial-mesenchymal transition through CDK1 regulated phosphorylation of ERK/GSK3beta/SNAIL pathway. Biochem Biophys Res Commun. 2021;546:1–6.
Rooney AG, McNamara S, Mackinnon M, Fraser M, Rampling R, Carson A, Grant R. Frequency, clinical associations, and longitudinal course of major depressive disorder in adults with cerebral glioma. J Clin Oncol. 2011;29:4307–12.
Wang P, Wang Z, Yan Y, Xiao L, Tian W, Qu M, Meng A, Sun F, Li G, Dong J. Psychological Stress Up-Regulates CD147 Expression Through Beta-Arrestin1/ERK to Promote Proliferation and Invasiveness of Glioma Cells. Front Oncol. 2020;10: 571181.
Lopes M, Vieira de Castro J, Pojo M, Goncalves CS, Martins EP, Coimbra B, Sotiropoulos I, Sousa N, Rodrigues AJ, Costa BM. Chronic Stress Does Not Influence the Survival of Mouse Models of Glioblastoma. Front Oncol. 2022;12: 856210.
Moreno-Smith M, Lu C, Shahzad MM, Pena GN, Allen JK, Stone RL, Mangala LS, Han HD, Kim HS, Farley D, Berestein GL, Cole SW, Lutgendorf SK, Sood AK. Dopamine blocks stress-mediated ovarian carcinoma growth. Clin Cancer Res. 2011;17:3649–59.
He Y, Li J, Koga T, Ma J, Dhawan S, Suzuki Y, Furnari F, Prabhu VV, Allen JE, Chen CC. Epidermal growth factor receptor as a molecular determinant of glioblastoma response to dopamine receptor D2 inhibitors. Neuro Oncol. 2021;23:400–11.
Weissenrieder JS, Reed JL, Green MV, Moldovan GL, Koubek EJ, Neighbors JD, Hohl RJ. The Dopamine D2 Receptor Contributes to the Spheroid Formation Behavior of U87 Glioblastoma Cells. Pharmacology. 2020;105:19–27.
Li Y, Wang W, Wang F, Wu Q, Li W, Zhong X, Tian K, Zeng T, Gao L, Liu Y, Li S, Jiang X, Du G, Zhou Y. Paired related homeobox 1 transactivates dopamine D2 receptor to maintain propagation and tumorigenicity of glioma-initiating cells. J Mol Cell Biol. 2017;9:302–14.
S.P. Caragher, J.M. Shireman, M. Huang, J. Miska, F. Atashi, S. Baisiwala, C. Hong Park, M.R. Saathoff, L. Warnke, T. Xiao, M.S. Lesniak, C.D. James, H. Meltzer, A.K. Tryba, A.U. Ahmed, Activation of Dopamine Receptor 2 Prompts Transcriptomic and Metabolic Plasticity in Glioblastoma, J Neurosci, 39 (2019) 1982–1993.
Liu H, Yang J, Zhang Y, Han J, Yang Y, Zhao Z, Dai X, Wang H, Ding X, Liu Y, Zhong W, Gao W, Sun T. Psychologic Stress Drives Progression of Malignant Tumors via DRD2/HIF1alpha Signaling. Cancer Res. 2021;81:5353–65.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural Science Foundation of China (82273111 and 82072770 to X.Z.; 81902526 to Y.W.); Natural Science Foundation of Jiangsu province (BK20201458 to X.Z.); Key Research & Development Plan (Social Development) of Jiangsu province (BE2022714 to X.Z.).
Author information
Authors and Affiliations
Contributions
XZ and RY were responsible for the conception and design and study supervision. XZ, YW, and XW performed the experiments and wrote the manuscript. KW, JQ, YZ, LZ and LG analyzed the data. XW, XW and YZ collected clinical specimens and followed up. All authors read and approved the final version of this paper.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The use of clinical samples was approved by the Ethics Committee of the Affiliated Hospital of Xuzhou Medical University and informed consent were obtained from all patients (Process number: XYFY2022-KL414-01). All animal experimental protocols were examined and verified by Laboratory Animal Ethics Committee of Xuzhou Medical University in accordance with Guide to Laboratory Animal Ethics Examination of Xuzhou Medical University (Process number: 202104A455).
Consent for publication
All authors consent to the publication of this article.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: sFig. 1
Chronic stress promotes tumor progression and shortens the survival of C57BL/6J glioma-bearing mice (related to Fig. 1). sFig. 2 In situ glioma transplantation does not cause depression in mice (related to Fig. 1). sFig. 3 DRD2 over-expression promotes glioma cell growth, while DRD2 down-regulation inhibits it (related to Fig. 3). sFig. 4 DRD2 is required for glioma progression induced by chronic stress (related to Fig. 3). sFig. 5. Chronic stress activates Wnt/β-catenin and stimulates TH secretion via DRD2/ERK (related to Fig. 4). sFig. 6. Combination treatment of PIMO and TMZ induces a synergistic anti-glioma effect in C57BL/6J mice (related to Fig.6).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, Y., Wang, X., Wang, K. et al. Chronic stress accelerates glioblastoma progression via DRD2/ERK/β-catenin axis and Dopamine/ERK/TH positive feedback loop. J Exp Clin Cancer Res 42, 161 (2023). https://doi.org/10.1186/s13046-023-02728-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-023-02728-8