
Abstract
CD8+ T cells play a central role in anti-tumor immunity. Naïve CD8+ T cells are active upon tumor antigen stimulation, and then differentiate into functional cells and migrate towards the tumor sites. Activated CD8+ T cells can directly destroy tumor cells by releasing perforin and granzymes and inducing apoptosis mediated by the death ligand/death receptor. They also secrete cytokines to regulate the immune system against tumor cells. Mitochondria are the central hub of metabolism and signaling, required for polarization, and migration of CD8+ T cells. Many studies have demonstrated that mitochondrial dysfunction impairs the anti-tumor activity of CD8+ T cells through various pathways. Mitochondrial energy metabolism maladjustment will cause a cellular energy crisis in CD8+ T cells. Abnormally high levels of mitochondrial reactive oxygen species will damage the integrity and architecture of biofilms of CD8+ T cells. Disordered mitochondrial dynamics will affect the mitochondrial number and localization within cells, further affecting the function of CD8+ T cells. Increased mitochondria-mediated intrinsic apoptosis will decrease the lifespan and quantity of CD8+ T cells. Excessively low mitochondrial membrane potential will cause the release of cytochrome c and apoptosis of CD8+ T cells, while excessively high will exacerbate oxidative stress. Dysregulation of mitochondrial Ca2+ signaling will affect various physiological pathways in CD8+ T cells. To some extent, mitochondrial abnormality in CD8+ T cells contributes to cancer development. So far, targeting mitochondrial energy metabolism, mitochondrial dynamics, mitochondria-mediated cell apoptosis, and other mitochondrial physiological processes to rebuild the anti-tumor function of CD8+ T cells has proved effective in some cancer models. Thus, mitochondria in CD8+ T cells may be a potential and powerful target for cancer treatment in the future.
Similar content being viewed by others
Background
Cancer remains the major cause of death even in wealthy countries [1]. T-cell-mediated cellular immune response is known to play a crucial role in anti-tumor immunity. Many previous studies have demonstrated a positive correlation between a high level of T-cell infiltration and a favorable prognosis in breast, lung, ovarian, colorectal, renal, prostate and gastric cancers, and melanoma as well [2]. Mature T cells are categorized into two subgroups: CD4+ and CD8+. Compared with CD4+ T cells, CD8+ T cells are more prominent anti-tumor cells in the body [3, 4].
When stimulated by processed tumor antigen-derived peptide, resting CD8+ T cells are activated via the interaction between T cell antigen receptor (TCR) -CD3 complex and major histocompatibility complex (MHC) I molecule. Simultaneously, costimulatory molecules, CD8 and CD28 provide the second signals for the reaction [5]. Cytokines such as interleukin 2 (IL-2), IL-4, IL-15 and IL-7 are consumed to maintain the later proliferation of activated CD8+ T cells [6, 7]. Upon co-stimulation from CD80, CD70, 4-1BB (CD137) and cytokines secreted from Dendritic cells (DCs) such as IL-12, INF-I and IL-15, naïve CD8+ T cells differentiate into cytotoxic T cells (CTLs) [8]. Subsequently, effector CD8+ T cells, which express CXC-chemokine receptor 3 (CXCR3), migrate to tumors from blood and lymphatic systems in response to TH1-type chemokines CX-chemokine ligand 9 (CXCL9), CXCL10 and CXCL11 [9, 10]. Activated CD8+ T cells directly eliminate tumor cells by releasing perforin and granzymes. They induce tumor cell apoptosis by expressing death ligands, mainly Fas ligand (FasL) and TNF-related apoptosis-inducing ligand (TRAIL) [5, 11]. Besides, activated CD8+ T cells secrete multiple cytokines such as INF-γ, TNF-α and TNF-β to regulate and improve overall anti-tumor response, including innate and specific immune systems [12,13,14].
Mitochondria play crucial and diverse roles during different stages of T cell adaptive immune response [15]. Firstly, as the main organelle for energy supply, mitochondrial energy metabolism provides adenosine triphosphate (ATP) to support all kinds of physiological activities of T cells. Secondly, the subcellular location of mitochondria regulates the directionality of T cell migration. Mitochondria gather near the cell edge where they approach the extracellular chemokines to offer sufficient ATP so that T cells can move towards higher concentrations of chemokines and recruit into tumor sites [15]. As an important signal molecule, the Ca2+ wave, which is prolonged by mitochondria gathering at the immune synapse (IS), is essential for T cell activation [15, 16]. In addition, the low mitochondrial reactive oxygen species (mROS) concentration favors T cell survival and function, and enhances TCR signaling transduction after tumor antigen stimulation in particular [17]. Therefore, the quantity and quality of mitochondria have a significant impact on T cell activity, distribution and function. So, mitochondrial dysfunction would directly affect the anti-tumor effect of CD8+ T cells and promote tumor occurrence and progression. Actually, in chronic infections and cancer, T cells are exposed to continuous antigen stimulation and/or inflammatory signals, and therefore mitochondrial dysfunction is considered a hallmark of resultant deterioration of T cell function, a situation called ‘exhaustion’ [15, 18]. Targeting mitochondria in CD8+ T cells and restoring their original condition to increase CD8+ T cell vitality, or even reverse the dysfunction, may become a feasible treatment strategy for cancer.
In this review, we will introduce the influencing mechanisms of mitochondrial abnormalities on the anti-tumor effect of CD8+ T cells, which may involve mitochondrial energy metabolism, mROS production, mitochondrial dynamic process, CD8+ T cell apoptosis mediated by mitochondria, mitochondrial membrane potential (MMP), and mitochondrial Ca2+ signaling regulation, as well as the possible efficient ways to treat cancer by targeting mitochondria to improve the anti-tumor function of CD8+ T cells.
Mitochondrial dysfunction affects the anti-tumor effect of CD8+ T cells
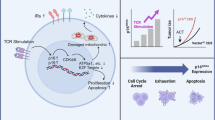
Mitochondrial physiological activities are critical for the various anti-tumor process of CD8+ T cells, and can even directly impact the number of CD8+ T cells. One strategy for cancer cells to burr the anti-tumor effect of CD8+ T cells is to inhibit their mitochondrial function (Fig. 1 and Fig. 2).

Mechanism of mitochondrial dysfunction in the disturbing anti-tumor activity of CD8+ T cells. (The red arrows indicate promotion, and the green arrows indicate inhibition) ①The decrease of mitochondrial membrane potential and the high expression of PD-1 restrict OXPHOS, reducing ATP synthesis. ②mROS accumulation in turn inhibits OXPHOS or limits respiratory metabolism by decreasing the MMP of CD8+ T cells. ③High level of mROS suppresses CD8+ T cell activation and proliferation by inhibiting NF-κB, mTOR, and NFAT5 signaling pathways. ④mROS accumulation plays a role in attritting telomeres and promoting senescence of CD8+ T cells. ⑤An excessively high mROS concentration activates the caspase signaling cascade and induces CD8+ T cell apoptosis. ⑥PD-1 hinders mitochondrial biogenesis through increasing repression of PGC1-α mediated by Akt. ⑦Imbalance in Bax and Bal-2 accelerates the release of cytochrome c from mitochondria and initiates CD8+ T cell apoptosis. ⑧Low MMP causes a high mitochondrial membrane permeability, triggering the subsequent release of cytochrome c and CD8+ T cell apoptosis. ⑨High MMP leads to a high level of mROS production. ⑩Increased cytoplasmic Ca2+ caused by failed mitochondrial Ca2+ buffering initiates the intrinsic apoptosis process. ⑪Accumulated Ca2+ in matrix inhibits mitochondrial biogenesis

Mechanism of TME affecting the mitochondrial function of CD8+ T cells. (The red arrows indicate promotion, and the green arrows indicate inhibition) ①Glucose competition and lactic acid accumulation increase the glycolysis stress of CD8+ T cells. ②Hypoxia inhibits OXPHOS and increases oxidative stress. ③Released K+ and metabolic waste PEG2, and adenosine interfere with the activity of the mTOR pathway. ④Certain fatty acids can directly damage mitochondrial structures. ⑤Soluble Fas released from tumor cells activates mitochondria-mediated intrinsic apoptotic processes
Mitochondrial energy metabolism maladjustment
CD8+ T cells require a massive amount of energy for normal functioning. Once activated, CD8+ T cells proliferate exponentially, migrate from the original site to tumor locations and synthesize inflammatory cytokines. All these activities require plenty of energy, especially mitochondrial ATP [19, 20] However, it was found in a systemic immunosuppressive property (SIP) tumor model [21] that atypical and non-proteinaceous molecules less than 3 kDa released from SIP-positive tumor cells could weaken mitochondrial energy metabolism and proliferation of CD8+ T cells, thus favoring SIP-positive tumor cells to escape anti-tumor immune response CD8+ T cells.
Naïve T cells depend on oxidative phosphorylation (OXPHOS) and fatty acid oxidation (FAO) for energy, while activated CD8+ T cells more rely on glycolysis even in the presence of sufficient oxygen [19]. This metabolic reprogramming phenomenon is similar to the Warburg effect observed in tumor cells [22]. Therefore, abundant glucose, the substrate for OXPHOS and glycolysis, is the key to CD8+ T cells, whether naïve or active. However, potential competition for glucose and other nutrient substances with tumor cells may inhibit CD8+ T cell functioning, and this metabolic stress can greatly affect signal transduction and gene expression of CD8+ T cells [18, 23,24,25]. Some in vitro studies [26,27,28] demonstrated that co-inhibitory molecules programmed cell death protein 1 (PD-1) and cytotoxic T lymphocyte-associated protein 4 (CTLA-4) could attenuate glycolysis in CD8+ T cells and suppress their anti-tumor function. For example, PD-1 can prevent glycolysis by maintaining FAO, and this effect could be achieved by activating the signal transducer and activator of transcription 3 (STAT3) of CD8+ T cells in obese patients [29, 30]. On the other hand, overexpressed PD-1 and CTLA-4 were proved on exhausted CD8+ T cells. So, the deficiency of glucose and energy may exacerbate CD8+ T cell exhaustion.
It cannot be neglectable that OXPHOS is not repressed entirely but continues to increase and plays a role in activated effector CD8+ T cells. Pyruvate, the final product of glycolysis, participates in the tricarboxylic acid cycle (TAC) as a substrate. mROS produced during OXPHOS participates in CD8+ T cell activation by working as an important signal molecule. For instance, it can regulate the nuclear factor of activated T cells (NFAT) to encourage antigen-induced cellular signaling [19, 31]. But studies have found that the level of OXPHOS is very low in tumor-infiltrated CD8+ T cells due to the low mitochondrial mass or MMP [32, 33]. Other studies have also reported that PD-1 engagement could inhibit OXPHOS in human CD8+ T cells through initiating a negative expression of metabolic genes progressively due to seriously impaired mitochondrial cristae [29].
In summary, mitochondrial energy metabolism dysfunction will not only further expand the energy gap of CD8+ T cells but affect the internal signal transduction, eventually resulting in tumor occurrence due to their weakened function in the anti-tumor effect.
Abnormally high level of mROS
Mitochondria are the major site for intracellular ROS formation [34]. ROS, mainly including superoxide anion (O2-), hydrogen peroxide (H2O2), and hydroxyl radical (OH•), are produced from the partially reduced O2, which renders a higher chemical reaction activity. The conventional view is that ROS are destructive agents, in fact, a suitable threshold level of ROS is necessary for intracellular signal transduction, kinase activation, and biologic responses associated with receptor signaling [35]. A low to moderate level of ROS is crucial for CD8+ T cell activation, proliferation, and effective substance secretion [34]. After the TCR complex binds to the tumor antigen, calcium will influx rapidly, resulting in the increased release of mROS [36]. This released mROS can regulate IL-2 and IL-4 secretion, which are essential for maintaining CD8+ T cell activation [34]. mROS also can increase the phosphorylation of c-Jun N-terminal kinase (JNK) and nuclear factor kappa-B (NF-κB), the levels of transcription factor (TF) STAT-1 and T-bet, as well as the secretion of IL-2, IL-4, TNF-α and granulocyte-macrophage colony stimulating factor (GM-CSF), all of which promote CD8+ T cells to synthesize and secret more INF-γ against tumor cells [34].
When mitochondria dysfunction occurs due to their higher activity, the accumulation of mROS will hurt CD8+ T cells in many ways. Dysfunctional mitochondria produce a large amount of ROS, which will directly cause damage to biological macromolecule DNA, protein and lipid [37]. Overproduction of ROS will inhibit NF-κB phosphorylation and activation, reduce CD8+ T cell sensitivity and response, and decrease the mammalian target activity of rapamycin complex 1 (mTORC1), which is vital in CD8+ T cell activation and metabolism [34, 38]. A high concentration of mROS will accelerate CD8+ T cell senescence. It is proved that mROS is a motivator for telomere length shortening in CD8+ T cells [39]. With reduced calcium influx into mitochondria and increased mROS production, respiration in CD8+ T cells will be depressed. All these alterations can reduce the MMP and slow down ATP synthesis [40]. Long-term exposure to ROS can activate the caspase cascade signal and elevate CD8+ T cell apoptosis in the peripheral tissue, and inhibit NFAT5 by combining with IL-6 promoter and preventing CD8+ T cell expansion [34, 41]. As a result, the number of CD8+ T cells is decreased and the anti-tumor function is aberrated.
A study about clear cell renal cell carcinoma (ccRCC) [42] reported that mitochondrial morphology and energy metabolism were abnormal in ccRCC-infiltrated CD8+ T cells. These condensed and fragmented mitochondria showed a hyperpolarized state and generated a large quantity of ROS. Similarly, abnormal and hyperpolarized mitochondria could also be observed in the peripheral blood of ccRCC patients. These experiments further demonstrated that excessive mROS was attributed to reduced mitochondrial superoxide dismutase 2 (SOD2), which could countervail mROS to relieve oxidative stress, but this protein was measured to be expressed less in ccRCC-infiltrated CD8+ T cells.
In conclusion, elevated mROS derived from stressed mitochondria will expedite functional loss of CD8+ T cells and weaken their anti-tumor effect.
Disordered mitochondrial dynamic processes
Mitochondrial dynamics mainly refers specifically to mitochondrial fusion and fission, but other dynamic changing processes, including mitochondrial mobility, biogenesis and autophagy, also have an influence on CD8+ T cells [40]. Mitochondrial fusion can promote OXPHOS and FAO in CD8+ T cells by facilitating cristae and electron transport chain (ETC) complex formation as well as increasing substrate intake. Mitochondrial fission can facilitate aerobic glycolysis and promote mitochondrial autophagy to delete dysfunctional mitochondria in CD8+ T cells [40, 43]. More importantly, mitochondrial fragmentation and accumulation underneath the TCR clusters are required for immune synapse formation. At the established IS, mitochondrial fragmentation continues to be needed for ATP supplement, local calcium buffering, TFs activation and cytokine secretion [16, 44]. Mitochondrial mobility determines mitochondrial subcellular localization and local concentrations in CD8+ T cells. Mitochondria gather at the uropod of CD8+ T cells when more energy is required for cell migration [20, 45]. At parallel, mitochondrial fission promotes their own transportation as the smaller mitochondria are more easily to move. In a solid cancer model, CD8+ T cells with defective mitochondrial fission would cause a reduced infiltration [46, 47]. The balance between mitochondrial biogenesis and autophagy sustains a stable scale of CD8+ T cells. Damaged mitochondria demand to be degraded by mitophagy, otherwise, these damaged mitochondria with lower MMP will promote cell senescence [48]. Autophagy related gene 7 (Atg7)-deficient CD8+ T cells are unable to constrain the mitochondrial content, resulting in ROS overproduction and Bcl-2-associated X protein (Bax) elevation, and finally increasing their apoptosis [49].
Previous studies on non-small cell lung cancer (NSCLC) and chronic viral infections reported that high expression of PD-1 on the surface of activated CD8+ T cells restrained transcriptional coactivator peroxisome proliferator-activated receptor-γ coactivator-1α (PGC1-α), a critical gene transcriptional regulator of mitochondrial metabolism and biogenesis, leading to mitochondrial depolarization accompanied with the reduced number and length of cristae, and a trend towards ROS accumulation. CD8+ T cells with these severe functional and structural changes in mitochondria present deficiency in energy metabolism and immune function [26, 29, 50]. Another study [32] demonstrated that PGC1-α-mediated defects in the mitochondrial structure and dynamics were related to chronic protein kinase B (PKB/Akt) activation. PGC1-α is regulated by several signal pathways, among which Akt mediates an obviously suppressive pathway. Compared with resting ones in lymph nodes, CD8+ T cells infiltrating in B16 tumor show a higher level of Akt activation. Consistent with this alteration, Akt-mediated inhibitory forkhead box O (Foxo) phosphorylation is increased as well.
Therefore, abnormal mitochondrial dynamics will cause abnormal structural and functional changes in mitochondria, which further blunts the anti-tumor response of CD8+ T cells, thus promoting tumor progression.
Increased mitochondria-mediated intrinsic apoptosis
Mitochondrial-mediated intrinsic apoptosis is an important pathway for programmed death in eukaryotic cells. According to the classical cell theory, mitochondrial membrane permeability will increase when cell apoptosis launches, resulting in the release of cytochrome c, apoptosis-inducing factor (AIF), endonuclease G and other nucleases and proteases from mitochondria into the cytoplasm [51]. This process can be regulated by B-cell lymphoma-2 (Bcl-2) family proteins [52]. Released cytochrome c combines with apoptotic protease-activating factor 1 (Apaf-1), caspase-9 and ATP, forming apoptosomes. Apoptosomes can activate caspase-9 and trigger subsequent caspase cascade, inducing the degradation phase of apoptosis [53]. In the meantime, endonuclease G and AIF lead to nuclear DNA degradation in a caspase-independent manner [54]. The increased apoptosis of CD8+ T cells mediated by mitochondria and the decreased amount of CD8+ T cells will inevitably reduce the immune surveillance function, which provides a chance for tumor development.
In patients with squamous cell carcinoma of the head and neck (SCCHN), the inhibited function of tumor-infiltrated CD8+ T cells was possibly caused by an increased proportion of apoptosis, which is visible both at the tumor sites and in the peripheral blood [55,56,57]. The same phenomenon was also observed in patients with other cancer types [58, 59]. Studies have demonstrated that there is an imbalance between pro-apoptosis and anti-apoptosis proteins in the Bcl-2 family in SCCHN CD8+ T cells. Compared with the control group, the expression level of Bax and Bcl-XL in the SCCHN group was upregulated markedly while the expression of Bcl-2 remained unchanged [55], suggesting that the mitochondria-mediated intrinsic apoptosis pathway may participate in SCCHN CD8+ T cell death in the peripheral blood. When mitochondrial Bcl-2 was downregulated, activated CD8+ T cells cultured in vitro quickly transformed into an apoptotic state, probably due to lack of IL-2, a crucial factor for maintaining an appropriate level of intracellular Bcl-2 and CD8+ T cell survival [60].
Therefore, an increased signal for inducing apoptosis in the extracellular environment and consequent enhanced intrinsic apoptosis mediated by mitochondria would decrease the number of CD8+ T cells involved in the anti-tumor response, thus promoting tumor development.
Mitochondrial membrane potential disruption
Stable MMP is essential for mitochondria to maintain their normal structure and physiological function. MMP is generated by proton pumps (ETC complex I, III and IV), by which, protons can be actively transported from inside to outside the inner membrane of mitochondria. The accumulation of a large number of protons in the intermembrane space eventually creates an electrochemical gradient with negative charges inside the mitochondrial matrix [61].
Data from different human CD8+ T cells suggest that tumor-infiltrated CD8+ T cells display an improvement in metabolism as represented by the increase in the MMP and mitochondrial number [62]. MMP is not only involved in ATP production, Ca2+ uptake and storage, ROS generation and decomposition, but is also closely related to cytochrome c release and cell apoptosis [63]. It was found that co-culture of CD8+ T cells with tumor cells decreased or even abolished the MMP of CD8+ T cells, further leading to an accumulation of dysfunctional mitochondria with disrupted membrane structures, cristae structure and declined cristae number and length of crista [64]. One of the reasons may be the exposure to gangliosides GD3 released from the tumor cells [65,66,67]. GD3 has been proven to be a selective inducer of the mitochondrial permeability transition pore (mPTP) in situ [68]. When mPTP opens, protons gathered outside the inner mitochondrial membrane start to flow inward driven by an electrochemical gradient, resulting in the MMP loss. Destructed structure and increased permeability of the inner mitochondrial membrane allow H2O2 and some solute molecules to influx freely, which further leads to mitochondrial swelling and outer mitochondrial membrane rupture. Eventually, cytochrome c is released and the caspase cascade is activated. CD8+ T cells subsequently began apoptosis processing [69].
However, some studies argued that CD8+ T cells with lower MMP had a longer survival time in vivo and a better anti-tumor effect [70]. They believed that lower MMP was directly associated with less ROS, which supports DNA protection and reparation. Compare with CD8+ T cells with high MMP, CD8+ T cells with low MMP own an extensive storage capacity of oxidized glutathione (GSSG) as well as an over expression of ROS-detoxifying enzymes mRNAs such as catalase (Cat), glutathione peroxidase 4 (Gpx4), superoxide dismutase 1 (SOD1) and SOD2. These enzymes are conductive to maintain intracellular redox equilibrium in CD8+ T cells [70]. The same conclusion can be proved in chronic lymphocytic leukemia (CLL) patients. CLL-derived CD8+ T cells showed a high MMP when the genes encoding subunits of ETC complexes I, III, IV and V, and mitochondrial-associated protein phosphatase 2 were expressed at a consistently high level, and the fitness of these CD8+ T cells was damaged [71].
In conclusion, impaired MMP will seriously affect the function and survival of CD8+ T cells in favor of tumor growth.
Fluctuant Ca2+ signaling regulates mitochondria in CD8+ T cells
Ca2+, a second messenger in CD8+ T cells, has been proved to be involved in multiple signaling pathways to regulate a variety of cellular and mitochondrial physiological processes [72]. Upon antigen stimulation, rising cytosolic Ca2+ binds with calmodulin (CaM) to activate protein phosphatase calcineurin, which later phosphorylates NFAT, resulting in IL-2 production and continuous cell proliferation [31, 73]. Ca2+ regulates the energy metabolism of CD8+ T cells in many aspects. At first, Ca2+ can activate three rate-limiting enzymes in TAC [pyruvate dehydrogenase (PDH), NAD+ isocitrate dehydrogenase (NAD-IDH) and α-ketoglutarate dehydrogenase (α-KGDH)], leading to more abundant ATP supply [74]. Secondly, Ca2+ regulates glycolysis in activated T cells by controlling the expression of glucose transporters (GLUT-1 and GLUT-3) and some crucial TFs [c-Myc, interferon regulatory factor 4 (IFR-4) and hypoxia inducible factor 1 (HIF-1)] [75]. Ca2+ also can enhance FAO via activating adenosine 5′-monophosphate (AMP)-activated protein kinase (AMPK) [72]. Extracellular Ca2+ influx after CD8+ T cell activation induces the rearrangement of the cytoskeleton, favoring the interactions between the TCR complex and APCs [76]. And the cytotoxicity of TILs is reported to rely on Ca2+ concentration [77, 78]. In addition, Ca2+ is a key factor in the activation of phosphoinositide 3-kinase (PI3K)-AKT-mTORC1 signaling pathway, further regulating mRNA transcription, protein synthesis and cell differentiation [72, 75].
Upon CD8+ T cells activation, phosphatidylinositol biphosphate (PIP2) is hydrolyzed in inositol triphosphate (IP3) and diacyl glycerol (DAG) by phospholipase C (PLC). Then, IP3 binds to the IP3 receptor (IP3R) on the endoplasmic reticulum (ER) to release stored Ca2+ into the cytoplasm [79]. Free Ca2+ decreased in ER will trigger the opening of Ca2+ release-activated Ca2+ channel (CRAC) on cytomembrane, leading to a further increased level of cytosolic Ca2+ by store-operated Ca2+ entry (SOCE) [80]. However, Ca2+ accumulation is not always beneficial to cells, so mitochondria, also as known as the main intracellular Ca2+ storage organelle, play an essential role in the Ca + buffering process. Mitochondria redistribute to the vicinity of the IS in a microtubule-actin-dependent manner [81], where they uptake Ca2+ through the voltage dependent anion channel (VDAC) in the outer mitochondrial membrane and the mitochondrial Ca2+ uniporter (MCU) in the inner mitochondrial membrane [82]. This reaction effectively prevents CRAC inhibition by Ca2 + −dependent negative feedback [72]. On the other hand, Ca2+ in mitochondria can be slowly extruded into the cytoplasm through mitochondrial Na+/Ca2+ exchanger (NCLX) and Na + independent exchanger mPTP, helping to prolong the Ca2+ signaling as well [76]. Mitochondrial Ca2+ buffering also can avoid the modulation of plasma membrane Ca2+ ATPase (PMCA), which pumps Ca2+ out of T cells, and maintain an appropriate intensity of SOCE [83].
In view of the complexity and importance of Ca2+ signaling, the fluctuant Ca2+ signaling related to mitochondria will inevitably disturb the anti-tumor function of CD8+ T cells. Continual Ca2+ extrusion due to the increased mPTP permeability will cause mitochondrial matrix swelling and outer membrane rupture, resulting in the release of cytochrome c and apoptosis proteins [84]. Because of the failed Ca2+ buffering of depolarized mitochondria, excessive Ca2+ in cytoplasm will inhibit CRAC activation, and even trigger the intrinsic apoptosis pathway. In this situation, Mfn2 has been reported to restrain partial Ca2+ influx to protect cells [85]. Similarly, MCU deletion also will damage mitochondrial Ca2+ uptake, as well as mitochondrial function, such as OXPHOS and mROS production [72]. In addition, mitochondrial Ca2+ overload due to the extra influx or hindered extrusion (e.g. deficiency of Na + or NCLX) can not only influence mROS production, but also mitochondrial biogenesis, which might through altering PGC1-α expression or Drp-1 phosphorylation [72, 83, 86].
Existing evidence has confirmed that a microenvironment with low Ca2+ concentration is more favorable for TILs. A clinical study on NSCLC treated with nivolumab has found that low serum Ca2+ level was parallel to significantly prolonged overall survival (OS) and progression free survival (PFS) [87]. Indeed, the optimum intracellular and extracellular Ca2+ for TILs were respectively defined as 122-334 nmol/L and 23-625 μmol/L, the latter being much lower than the normal physiological concentration [78]. These results suggest properly restraining the Ca2+ signaling by decreasing the extracellular Ca2+ or selectively blocking CRAC to prevent CD8+ T cell exhaustion and enhance the efficacy of TILs [78, 88]. However, regrettably, none of the above studies addressed the potential role of mitochondria.
Although we can safely draw a conclusion that mitochondrial Ca2+ regulation is important to the anti-tumor function of CD8+ T cells, there are not many specific types of research that have focused on this. So, further investigation is required to address this issue.
Mitochondria of CD8+ T cells in the tumor microenvironment
The tumor microenvironment (TME), composed of cellular components (tumor cells, immune cells and other stromal cells) and non-cellular components (extracellular matrix, chemokines, cytokines and growth factors), has a complex effect on anti-tumor immunity and cancer fate [89]. In severe hypoxic TME, CD8+ T cells are under high oxidative stress conditions. Dysfunctional mitochondria generate unbearable mROS, maintaining continuous activation of NFAT, which upregulates transcriptional factors TOX and NR4A to induce the expression of exhaustion-related genes [90, 91]. Hypoxic interferes with mitochondrial fusion through the miR24-Myc-Mfn1 axis. Decreased OXPHOS in fragmented mitochondria cannot provide sufficient ATP for CD8+ T cells [92]. Hypoxia inducible factor-1 (HIF-1) also disturbs OXPHOS by activating pyruvate dehydrogenase kinase 1 (PDK1). In the meantime, HIF-1 activates mTORC1 and lactate dehydrogenase A (LDHA) to enhance glycolysis, but this exacerbates the glucose deficiency in TME [93, 94]. On the one hand, the intense glucose competition in TME limits CD8+ T cells’ energy support [95], and on the other hand, the accumulation of metabolism wastes and negative factors from tumor cells can damage the mitochondria of CD8+ T cells. The vigorous aerobic glycolysis in tumor cells produces massive lactic acid, leading to adverse TME with a low pH, in which glycolysis of CD8+ T cells is declined for inhibited polyphosphate kinase (PPK) [96]. Accumulated adenosine and prostaglandin E2 (PGE2) in TME respectively bind to A2A and EP4 receptors on CD8+ T cells, causing increased activity of protein kinase A (PKA), which negatively regulates mTORC1, the important pathway for mitochondrial function [95, 97, 98]. Dead tumor cells will release abundant K+ into TME. High K+ mediates Akt-mTORC1 inactivation via protein phosphatase 2A (PP2A), with possible involvement of zinc finger proteins 91 (ZFP91) [99, 100]. Excessive lipids in TME, particularly oxidized lipids and some certain long-chain fatty acids, can directly destroy mitochondria structure behind transferred into CD8+ T cells [101, 102]. Another immunosuppressive metabolite is D-2-hydroxyglutarate (D2HG), and it can inhibit ATP synthase located on the inner mitochondrial membrane [103]. Additionally, soluble Fas secreted from tumor cells also can activate mitochondria-mediated intrinsic apoptosis, depressing anti-tumor immunity of CD8+ T cells [104]. At last, sustained antigenic stimulation, severe hypoxia, nutritional deficiencies, acidic environment, cholesterol accumulation, high concentration of K+ and soluble factors [such as vascular endothelial growth factor A (VEGF-A), indoleamine2,3-dioxygenase1 (IDO), INF-γ and TGF-β] secreted by other cells in TME all upregulate expression of co-inhibition receptors (mainly PD-1 and CTLA-4) on CD8+ T cells. They can affect mitochondrial function through multiple pathways [91, 105].
Recent single-cell transcriptome results have revealed some differentially expressed genes that may be associated with dysfunctional mitochondria of exhausted CD8+ T cells in TME. SARDH, NDUFB3 and HSPA1A are involved in the ETC, involving in ATP metabolism and mROS production [106, 107]. FABP5 and TPI1 are implicated in multiple energy metabolic processes, including glucose metabolism (both OXPHOS and glycolysis) and fatty acid metabolism [106, 107]. FABP5 may also directly shape mitochondrial cristae [108]. CD38 not only negatively affects metabolism, but also activates the mitochondria-mediated intrinsic apoptosis [107]. Another two genes PHLDA1 and EPSTI1 facilitate intrinsic apoptosis as well [107, 109]. PRDX3 and HSBP1 can reduce oxidative stress levels and maintain a stable MMP [107]. In addition to cell apoptosis and electron transport, CDK1 alters mitochondrial dynamics by regulating Drp1 [110, 111].
In the meantime, several key TFs were discovered. STAT2 regulates mitochondrial fission, while STAT3 regulates mitochondrial Ca2+ homeostasis through the ETC [112, 113]. FOXP1, FOXP3, FOXO1 and GATA3 all control mitochondrial respiration and oxidative stress as well as mitochondrial dynamics, autophagy and biogenesis [112, 114,115,116]. NF-Κb and JUN are mainly associated with mitochondrial fragmentation and increased cellular apoptosis [112]. Besides, PRDM1, which encodes protein blimp1, is proved as an important TF connected with PGC1-α-mediated mitochondrial biogenesis [117]. Other TFs, like BATF, VHL and ETV1, respectively regulate mitochondrial energy metabolism and intrinsic apoptosis process [115, 118, 119].
Thus, it can be seen that TME mainly affects mitochondrial metabolism, oxidative stress, intrinsic apoptosis and dynamic processes in CD8+ T cells.
Treatment of cancer by targeting mitochondria in CD8+ T cells
Although mitochondrial abnormality and dysfunction can accelerate the loss of the anti-tumor effect of CD8+ T cells, this consequence is not irreversible. We can aim at mitochondria in CD8+ T cells as a target, and recover the anti-tumor activity of CD8+ T cells by regulating the mitochondrial metabolism, dynamics or other physiological processes, and increasing mitochondrial mass and quality (Fig. 3). It may prove to be a new strategy for cancer treatment in the future.

Targeting multiple metabolic pathways of mitochondria can enhance the anti-tumor effect of CD8+ T cells. (The red arrows indicate promotion, and the green arrows indicate inhibition) ①Activating CD28, increasing enolase 1 activity, or adding exogenous pyruvate and PEP can promote the glycolysis of CD8+ T cells. ②Inhibiting LDH activity and supplementing exogenous L-arginine can promote OXPHOS of CD8+ T cells. ③For melanoma, fibrosarcoma, colon cancer and lung cancer, a combination of bezafibrate with PD-1 blockade can promote FAO and function of CD8+ T cells, while inhibiting FAO can improve the anti-tumor effect of CD8+ T cells in breast cancer. ④Up-regulating the expression level of transcriptional activity of PGC1-α in CD8+ T cells can facilitate mitochondrial biogenesis and functional recovery. It can be achieved by stimulating 4-1BB combined with PD-1 blockade, inhibiting the activity of Akt, inhibiting the mTORC2 pathway to activate Foxo1, and using nicotinamide adenine dinucleotide to activate SIRT1. ⑤Regulating mitochondrial fusion and fission by essential GTPase (Drp1, Mfn1, Mfn2 and Opa1) can control CD8+ T cell function. ⑥MAPK/ERK can prevent the release of cytochrome C from mitochondria to cytoplasm and the activation of caspase cascade, decreasing CD8+ T cell apoptosis. ⑦Addition of BH4 can regulate mitochondrial iron transport and respiration in CD8+ T cells, increasing the activation of CD8+ T cells
Targeting mitochondrial energy metabolism of CD8+ T cells
Glycolysis provides the most fuel for activated CD8+ T cells. It was reported [120] that CD28 co-stimulation could enhance glycolysis of ccRCC-infiltrated CD8+ T cells and recover their metabolism mainly by increasing glucose transporter 3 (GLUT3), and under CD28 co-stimulation mitochondria of CD8+ T cell showed more fusion and quantity, boosting mitochondrial activity restoration. Another study [33] reported that enolase 1 activity was downregulated in metabolism-impaired tumor-infiltrated CD8+ T cells. It restricted the production of phosphoenolpyruvate (PEP), which is known to play an important role in glycolysis as an intermediate, as well as production of pyruvate, which is a downstream product of enolase 1. So, increasing enolase 1 activity or adding exogenous PEP and pyruvate is effective for repairing partial glucose metabolism and anti-tumor activity of CD8+ T cells.
Regulating mitochondrial OXPHOS is another means to regulate CD8+ T cell activity. Inhibition of lactate dehydrogenase (LDH) improves pyruvate oxidation, promotes pyruvate into TAC and finally enhances OXPHOS, and the effect is much more efficient with IL-2 synergy [121]. Lots of L-arginine is consumed by activated CD8+ T cells for downstream metabolism, so the intracellular level of L-arginine is significantly reduced inside activated CD8+ T cells. Some groups [122] demonstrated that adding exogenous L-arginine could upregulate the serine biosynthesis pathway, and further strengthen TAC, leading to the transformation of CD8+ T cell metabolism from glycolysis to OXPHOS, which reduces the Warburg effect, and improves CD8+ T cell survival and anti-tumor function as well.
Tumor-infiltrated CD8+ T cells have to experience nutrient limitations due to the intense competition with cancer cells for oxygen, glucose and other key nutrients. This kind of nutrient restriction can cause hyporesponsiveness of CD8+ T cells even under stimulation by highly antigenic tumor cells [123]. In melanoma immunotherapy, enhancing FAO could compensate for the energy deficiency of CD8+ T cells and elevate the anti-tumor effect [124]. More studies in fibrosarcoma, colon cancer and lung cancer mouse models demonstrated that combined treatment by benzabate and PD-1 blockage could activate mitochondria of CD8+ T cells and promote FAO to provide extra energy. Besides, the same treatment also worked through other pathways by ①improving CD8+ T cell glycolysis and OXPHOS; ②upregulating Bcl-2 and carnitine palmitoyl transferase 1 (Cpt1), which can bind together to decrease CD8+ T cell apoptosis, and the latter was also found to participate in FAO; ③promoting CD8+ T cell infiltration by increasing the expression of CXCL9 and CXCL10 from tumor tissues and CXCR3 in CD8+ T cells [125, 126].
So far, it is still difficult to draw a conclusion on whether enhancing FAO is always positive for CD8+ T cells in different types of cancers at present. A study [30] about obesity-associated breast cancer reported that suppressed FAO could recover glycolysis and the anti-tumor capacity of CD8+ T cells. Given the role of obesity is complex in cancer progression and immunity regulation, we won’t discuss it further in this article. In all, the above facts indicate that targeting mitochondrial energy metabolism can regulate the anti-tumor activity of CD8+ T cells.
Targeting mitochondrial dynamic of CD8+ T cells
PGC1-α is an essential transcriptional co-activator that regulates mitochondrial biogenesis by binding to the nuclear receptors and specific sequences in the promoter region of the target genes [127]. The level of PGC1-α depends on energy demand [128]. With CD8+ T cell activation, the PGC1-α expression is upregulated, while post-translational modification can modulate the transcriptional activity of PGC1-α [127]. Phosphorylation of PGC1-αby Akt will largely inhibit its transcriptional activity [128, 129]. As we mentioned previously, abnormal mitochondrial dynamics in tumor-infiltrated CD8+ T cells are related to the downregulation or inhibition of PGC1-α. Thus, increasing PGC1-α expression and activity may be able to rescue mitochondria and the anti-tumor function of CD8+ T cells.
CD8+ T cells with PGC1-α overexpression showed a higher OXPHOS level and a greater spare respiratory capacity (SRC) in vitro, suggesting that their mitochondrial biogenesis is restored. In line with this, these CD8+ T cells also exhibited a longer survival time and an increased anti-tumor effect [32]. The PGC1-α level was low in CD8+ T cells with overexpressed PD-1. Although mitochondrial activity was improved in tumor-bearing mice after the PD-1 block, simply blocking PD-1 could not completely reverse mitochondria dysfunction [21, 32, 130]. Another costimulatory molecule 4-1BB, highly expressed on exhausted T cells, was also found to elevate PGC1-α-mediated mitochondrial biogenesis and fusion by activating the p38- mitogen-activated protein kinase (MAKP) signal pathway. A combination of 4-1BB agonism might help overcome the metabolic dilemma of CD8+ T cells. In addition, 4-1BB co-stimulation combined with PD-1 block could exert a potent anti-tumor effect [40, 130]. Akt-mediated signaling activation repressed the PGC1-α in CD8+ T cells [32]. Akt inhibition helped improve FAO, SRC, as well as survival and expansion of transferred anti-tumor CD8+ T cells in a study about adoptive cell therapy [131]. Although they did not further explore whether the effect had a direct link to PGC1-α, their finding could not be excluded as a possible mechanism. In addition, PGC1-α is also regulated by Foxo1. Inhibition of the mTORC2 pathway to prevent Fxoxo1 phosphorylation and activation could promote PGC1-α and subsequent generation of memory CD8+ T cells [49, 132, 133]. Mouse embryonic fibroblasts treated with nicotinamide adenine dinucleotide (NAD) could deacetylate PGC1-α by activating silent information regulator type 1 (SIRT1), consequently increasing the PGC1-α transcriptional activity and restoring the mitochondrial original function [134,135,136]. In mice fed with nicotinamide riboside (NR), the precursor of NAD generation, melanoma and colon tumors growth was significantly restricted by CD8+ T cell-dependent immune. In vitro, CD8+ T cells treated with NR showed a strong anti-tumor response for reduced mROS generation and more secretion of effector cytokines. But these results were more likely to be related to enhanced autophagy of dysfunctional mitochondria than mitochondrial biogenesis [64]. As the complex regulation of the mitochondrial dynamic process, more studies are required for the specific mechanism.
Besides mitochondrial biogenesis, the anti-tumor ability of CD8+ T cells can be changed through regulating mitochondrial fusion and fission. Several GTPases belonging to the Dynamin family are the core components in mitochondrial dynamics. GTPase Dynamin-related protein 1 (Drp1) oligomerization drives mitochondrial fission. Mitofusins 1 (Mfn1) and Mfn2 actuate outer mitochondrial membrane fusion, followed by inner mitochondrial membrane fusion actuated by optic atrophy 1 (Opa1) [137, 138]. These proteins are essential targets to control mitochondrial dynamics as well.
It is observed that drugs promoting mitochondrial fusion extended the longevity of effector CD8+ T cells and enhanced their secretion of INF-γ and TNF-α in tumor-bearing mice [43]. Mitochondrial fusion recovers cellular function through increasing mitochondrial metabolism and decreasing the sensitivity of cell death signals [139]. Several molecules can directly modulate mitochondrial fusion, such as SAMβA (preventing Mfn 1 phosphorylation) [140]; 15-oxospiramilactone (preventing degradative mitofusins ubiquitination) [141]; Leflunomide (facilitating mitofusins expression) [142]. While other molecules can enhance mitochondrial fusion by inhibiting fission, such as P110 and P259 (blocking Drp1 receptor) [143, 144]; mdivi-1 (inhibiting Drp1 GTPase activity) [145]. But at the same time, we must realize that promoting mitochondrial fusion is not always beneficial to CD8+ T cells. Drp1-mediated mitochondrial fission not only helps chemotaxis of CD8+ T cells and their infiltration into the tumor site but controls mitochondria distribution during cell expansion. It is proved that CD8+ T cell migration and effectors function were hindered when Drp1 expression was inhibited [146]. Therefore, enhancing Drp1-mediated mitochondrial fission may promote CD8+ T cell infiltration towards the tumor, particularly in the absence of effector CD8+ T cells [20, 45, 147]. On the other hand, increased Drp-1-mediated mitochondrial fission has been confirmed to promote aerobic glycolysis, which activated CD8+ T cells relied on [43]. However, there are only a few reports about mitochondria-fission-promoting drugs. This is an area worth exploring in the future.
Since mitochondrial fusion and fission can both help the anti-tumor function of CD8+ T cells, how do we choose or balance between these two seemingly contradictory options in practical application? Based on the current present evidence, mitochondrial fission contributes more to CD8+ T cells activation and infiltration, while mitochondrial fusion is connected to cellular longevity and cytokine secretion after cell activation, so a dynamic therapeutic strategy that promotes mitochondrial fission at the early stage and promotes mitochondrial fusion later and is continuously adjusted is needed.
Targeting mitochondria-mediated intrinsic apoptosis of CD8+ T cell
Interaction between extracellular death ligands and superficial death receptors results in sequential activation of Fas-associated death domain (FADD) protein and caspase-8 in CD8+ T cells. Activated caspase-8 splits Bid protein to cleaved Bid (tBid), which translocates onto mitochondria and causes mitochondrial depolarization. Then, MMP is changed and the mitochondrial membrane becomes more permeable, allowing cytochrome c release [148,149,150]. Mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) can negatively regulate apoptosis by inhibiting caspase-8 and Bit activation, impeding mitochondrial depolarization, and cytochrome c release, protecting CD8+ T cells from death [149, 151, 152].
Targeting other mitochondrial physiological processes in CD8+ T cells
Tetrahydrobiopterin (BH4) is required for CD8+ T cell expansion both in vitro and in vivo, because it is involved in iron metabolism and mitochondrial respiration regulation. After BH4 inhibition by kynurenine, the anti-tumor function of CD8+ T cells is suppressed, and this phenomenon may be mechanistically associated with disturbed iron redox of cytochrome c in mitochondria and resultant mitochondrial dysfunction. Therefore, increasing the BH4 level can to some extent help CD8+ T cells get out of this dilemma and restrain tumor development [153].
Conclusion
In this review, we have discussed the adverse effects of mitochondrial dysfunction on CD8+ T cells, and systematically reviewed the recent research progress of targeting mitochondria to restore the anti-tumor function of CD8+ T cells. Beyond the classical function of energy metabolism and ATP production, we also emphasized the important role of mitochondria in intracellular signal transduction, cell activation and migration, cell senescence and apoptosis and other crucial physiological processes in CD8+ T cells. In particular, single-cell sequencing techniques have unveiled some possible genes with altered expression in the TME, providing additional clues to delve into the mechanism of mitochondrial dysfunction of CD8+ T cells. So, it is important to recognize that mitochondrial dysfunction can disturb the original normal immune function of CD8+ T cells by various means, further leading to tumor occurrence and progression. On the one hand, the complex relationship between tumor growth and anti-tumor immunity has been confirmed again, but on the other hand, several possible treatment targets of mitochondria are revealed at the same time, which may reverse the depressed anti-tumor function of CD8+ T cells.
Prospectively, targeting mitochondria to enhance the anti-tumor function of CD8+ T cells presents a better further application perspective in tumor immunotherapy, especially in chimeric antigen receptor T cell (CAR-T) therapy. CAR-T cells (mainly CD8+ T cells) can specifically recognize tumor antigens in vivo and eliminate tumor cells through cellular immunity. But the efficacy of CAR-T therapy in solid tumors is not satisfactory as that in hematological malignancies. CAR-T cell administration is found to show an impaired anti-tumor effect in vivo (the specific manifestations are limited proliferation, hindered migration and infiltration, transient persistence, premature senescence as well as TME immunosuppressive) [154, 155], an important reason for which is cellular mitochondrial dysfunction in the TME [156, 157]. Therefore, restoring mitochondrial function may be able to help break the dilemma and improve the prognosis of cancer patients. Nonetheless, the treatment strategy of targeting mitochondria to regulate the anti-tumor function of the immune system is still in the early stage. More studies are needed to evaluate the efficacy and safety of this approach in clinical use.
Availability of data and materials
Not applicable.
Abbreviations
- CD:
-
Cluster of differentiation
- APCs:
-
Antigen-presenting cells
- TCR:
-
T cell antigen receptor
- CTLs:
-
Cytotoxic T cells
- ATP:
-
Adenosine triphosphate
- IS:
-
Immune synapse
- mROS:
-
Mitochondrial reactive oxygen species
- MMP:
-
Mitochondrial membrane potential
- MHC:
-
Major histocompatibility complex
- IL:
-
Interleukin
- DCs:
-
Dendritic cells
- CXCXR:
-
CXC-chemokine receptor
- TH:
-
T helper
- CXCL:
-
Chemokines CX-chemokine ligand
- pMHC:
-
Peptide-loaded major histocompatibility complex
- MTOC:
-
Microtubule-organizing center
- FasL:
-
Fas ligand
- TNF:
-
Tumor necrosis factor
- TRAIL:
-
TNF-Related apoptosis-inducing ligand
- INF:
-
Interferon
- ADCC:
-
Antibody- dependent cellular cytotoxicity
- TNFR:
-
TNF receptor
- SIP:
-
Systemic immunosuppressive property
- OXPHOS:
-
Oxidative phosphorylation
- FAO:
-
Fatty acid oxidation
- CTLA:
-
Cytotoxic T lymphocyte-associated protein
- PD:
-
Programmed cell death protein
- STAT:
-
Signal transducer and activator of transcription
- TAC:
-
Tricarboxylic acid cycle
- NFAT:
-
Nuclear factor of activated T cells
- JNK:
-
c-Jun N-terminal kinase
- NF-κB:
-
Nuclear factor kappa-B
- TF:
-
Transcription factor
- Gm-csf:
-
Granulocyte-macrophage colony stimulating factor
- mTORC:
-
Mammalian target activity of rapamycin complex
- ccRCC:
-
Clear cell renal cell carcinoma
- SOD:
-
Mitochondrial superoxide dismutase
- ETC:
-
Electron transport chain
- Atg:
-
Autophagy related gene
- Bcl-2:
-
B-cell lymphoma-2
- Bax:
-
Bcl-2-associated X protein
- NSCLC:
-
Non-small cell lung cancer
- PGC1-α:
-
Peroxisome proliferator-activated receptor-γ coactivator-1α
- PKB/Akt:
-
Chronic protein kinase B
- Foxo:
-
Forkhead box O
- AIF:
-
Apoptosis-inducing factor
- Apaf-1:
-
Apoptotic protease-activating factor 1
- SCCHN:
-
Squamous cell carcinoma of the head and neck
- mPTP:
-
Mitochondrial permeability transition pore
- GSSG:
-
Glutathione
- Cat:
-
Catalase
- Gpx:
-
Glutathione peroxidase
- SOD:
-
Superoxide dismutase
- CLL:
-
Chronic lymphocytic leukemia
- Glut:
-
Glucose transporter
- PEP:
-
Phosphoenolpyruvate
- LDH:
-
Lactate dehydrogenase
- Cpt:
-
Carnitine palmitoyl transferase
- SRC:
-
Pare respiratory capacity
- MAKP:
-
Mitogen-activated protein kinase
- NAD:
-
Nicotinamide adenine dinucleotide
- SIRT:
-
Silent information regulator type
- NR:
-
Nicotinamide riboside
- Drp:
-
Dynamin-related protein
- Mfn:
-
Mitofusins
- Opa:
-
Optic atrophy
- FADD:
-
Fas-associated death domain
- MAPK:
-
Mitogen-activated protein kinase
- ERK:
-
Extracellular signal-regulated kinase
- CAR-T:
-
Chimeric antigen receptor T cell
- CaM:
-
Calmodulin
- PDH:
-
Pyruvate dehydrogenase
- NAD-IDH:
-
NAD+ isocitrate dehydrogenase
- α-KGDH:
-
α-ketoglutarate dehydrogenase
- GLUT:
-
Glucose transporters
- IFR-4:
-
Interferon regulatory factor 4
- HIF-1:
-
Hypoxia inducible factor 1
- AMP:
-
Adenosine 5’-monophosphate
- AMPK:
-
AMP-Activated protein kinase
- PI3K:
-
Phosphoinositide 3-kinase
- PIP2:
-
Phosphatidylinositol biphosphate
- IP3:
-
Inositol triphosphate
- DAG:
-
Diacyl glycerol
- PLC:
-
Phospholipase C
- IP3R:
-
IP3 receptor
- ER:
-
Endoplasmic reticulum
- CRAC:
-
Ca2+ release-activated Ca2+ channel
- SOCE:
-
Store-operated Ca2+ entry
- VDAC:
-
Voltage dependent anion channel
- MCU:
-
Mitochondrial Ca2+ uniporter
- NCLX:
-
Na+/Ca2+ exchanger
- PMCA:
-
Plasma membrane Ca2+ ATPase
- OS:
-
Overall survival
- PFS:
-
Progression free survival
- TME:
-
Tumor microenvironment
- PDK1:
-
Pyruvate dehydrogenase kinase 1
- LDHA:
-
Lactate dehydrogenase A
- PPK:
-
Polyphosphate kinase
- PGE2:
-
Prostaglandin E2
- PKA:
-
Protein kinase A
- PP2A:
-
Protein phosphatase 2A
- ZFP91:
-
Zinc finger proteins 91
- D2HG:
-
D-2-hydroxyglutarate
- VEGF-A:
-
Vascular endothelial growth factor A
- IDO:
-
Indoleamine2, 3-dioxygenase1
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71:209–49.
Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev. 2018;32:1267–84.
Hanson HL, Donermeyer DL, Ikeda H, White JM, Shankaran V, Old LJ, et al. Eradication of established tumors by CD8+ T cell adoptive immunotherapy. Immunity. 2000;13:265–76.
Jiang X, Xu J, Liu M, Xing H, Wang Z, Huang L, et al. Adoptive CD8(+) T cell therapy against cancer: challenges and opportunities. Cancer Lett. 2019;462:23–32.
Martinez-Lostao L, Anel A, Pardo J. How do cytotoxic lymphocytes kill Cancer cells? Clin Cancer Res. 2015;21:5047–56.
Waldman AD, Fritz JM, Lenardo MJ. A guide to cancer immunotherapy: from T cell basic science to clinical practice. Nat Rev Immunol. 2020.
Liu G, Yu Y, Feng F, Zhu P, Zhang H, Zhang D, et al. Human CD8(+)CD28(−) T suppressor cells expanded by common gamma chain (gammac) cytokines retain steady allospecific suppressive capacity in vivo. BMC Immunol. 2020;21:23.
Palucka AK, Coussens LM. The basis of Oncoimmunology. Cell. 2016;164:1233–47.
Nagarsheth N, Wicha MS, Zou W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat Rev Immunol. 2017;17:559–72.
Pontes Ferreira C, Cariste LM, Ferri Moraschi B, Ferrarini Zanetti B, Won Han S, Araki Ribeiro D, et al. CXCR3 chemokine receptor guides Trypanosoma cruzi-specific T-cells triggered by DNA/adenovirus ASP2 vaccine to heart tissue after challenge. PLoS Negl Trop Dis. 2019;13:e0007597.
Golstein P, Griffiths GM. An early history of T cell-mediated cytotoxicity. Nat Rev Immunol. 2018;18:527–35.
Fong TA, Mosmann TR. Alloreactive murine CD8+ T cell clones secrete the Th1 pattern of cytokines. J Immunol. 1990;144:1744–52.
Lang D, Terstesse M, Dohle F, Bangen P, Banas B, Pauels HG, et al. Protein kinase C (PKC) dependent induction of tissue factor (TF) by mesangial cells in response to inflammatory mediators and release during apoptosis. Br J Pharmacol. 2002;137:1116–24.
Calzascia T, Pellegrini M, Hall H, Sabbagh L, Ono N, Elford AR, et al. TNF-alpha is critical for antitumor but not antiviral T cell immunity in mice. J Clin Invest. 2007;117:3833–45.
Desdin-Mico G, Soto-Heredero G, Mittelbrunn M. Mitochondrial activity in T cells. Mitochondrion. 2018;41:51–7.
Quintana A, Hoth M. Mitochondrial dynamics and their impact on T cell function. Cell Calcium. 2012;52:57–63.
Diebold L, Chandel NS. Mitochondrial ROS regulation of proliferating cells. Free Radic Biol Med. 2016;100:86–93.
Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol. 2015;15:486–99.
Patel CH, Powell JD. Targeting T cell metabolism to regulate T cell activation, differentiation and function in disease. Curr Opin Immunol. 2017;46:82–8.
Campello S, Lacalle RA, Bettella M, Manes S, Scorrano L, Viola A. Orchestration of lymphocyte chemotaxis by mitochondrial dynamics. J Exp Med. 2006;203:2879–86.
Kumar A, Chamoto K, Chowdhury PS, Honjo T. Tumors attenuating the mitochondrial activity in T cells escape from PD-1 blockade therapy. Elife. 2020;9.
Xu K, Yin N, Peng M, Stamatiades EG, Shyu A, Li P, et al. Glycolysis fuels phosphoinositide 3-kinase signaling to bolster T cell immunity. Science. 2021;371:405–10.
Schietinger A, Greenberg PD. Tolerance and exhaustion: defining mechanisms of T cell dysfunction. Trends Immunol. 2014;35:51–60.
Siska PJ, Rathmell JC. T cell metabolic fitness in antitumor immunity. Trends Immunol. 2015;36:257–64.
Jiang Y, Li Y, Zhu B. T-cell exhaustion in the tumor microenvironment. Cell Death Dis. 2015;6:e1792.
Bengsch B, Johnson AL, Kurachi M, Odorizzi PM, Pauken KE, Attanasio J, et al. Bioenergetic insufficiencies due to metabolic alterations regulated by the inhibitory receptor PD-1 are an early driver of CD8(+) T cell exhaustion. Immunity. 2016;45:358–73.
Parry RV, Chemnitz JM, Frauwirth KA, Lanfranco AR, Braunstein I, Kobayashi SV, et al. CTLA-4 and PD-1 receptors inhibit T-cell activation by distinct mechanisms. Mol Cell Biol. 2005;25:9543–53.
Patsoukis N, Bardhan K, Chatterjee P, Sari D, Liu B, Bell LN, et al. PD-1 alters T-cell metabolic reprogramming by inhibiting glycolysis and promoting lipolysis and fatty acid oxidation. Nat Commun. 2015;6:6692.
Ogando J, Saez ME, Santos J, Nuevo-Tapioles C, Gut M, Esteve-Codina A, et al. PD-1 signaling affects cristae morphology and leads to mitochondrial dysfunction in human CD8(+) T lymphocytes. J Immunother Cancer. 2019;7:151.
Zhang C, Yue C, Herrmann A, Song J, Egelston C, Wang T, et al. STAT3 activation-induced fatty acid oxidation in CD8(+) T effector cells is critical for obesity-promoted breast tumor growth. Cell Metab. 2020;31(148-61):e5.
Sena LA, Li S, Jairaman A, Prakriya M, Ezponda T, Hildeman DA, et al. Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity. 2013;38:225–36.
Scharping NE, Menk AV, Moreci RS, Whetstone RD, Dadey RE, Watkins SC, et al. The tumor microenvironment represses T cell mitochondrial biogenesis to drive Intratumoral T cell metabolic insufficiency and dysfunction. Immunity. 2016;45:374–88.
Gemta LF, Siska PJ, Nelson ME, Gao X, Liu X, Locasale JW, et al. Impaired enolase 1 glycolytic activity restrains effector functions of tumor-infiltrating CD8(+) T cells. Sci Immunol. 2019;4.
Chen X, Song M, Zhang B, Zhang Y. Reactive oxygen species regulate T cell immune response in the tumor microenvironment. Oxidative Med Cell Longev. 2016;2016:1580967.
Devadas S, Zaritskaya L, Rhee SG, Oberley L, Williams MS. Discrete generation of superoxide and hydrogen peroxide by T cell receptor stimulation: selective regulation of mitogen-activated protein kinase activation and fas ligand expression. J Exp Med. 2002;195:59–70.
Kong H, Chandel NS. Regulation of redox balance in cancer and T cells. J Biol Chem. 2018;293:7499–507.
Yu L, Xie S, Xiao J, Wang Z, Zhang X. Quantitative measurement of cerebral oxygen extraction fraction using MRI in patients with MELAS. PLoS One. 2013;8:e79859.
Li M, Zhao L, Liu J, Liu A, Jia C, Ma D, et al. Multi-mechanisms are involved in reactive oxygen species regulation of mTORC1 signaling. Cell Signal. 2010;22:1469–76.
Sanderson SL, Simon AK. In aged primary T cells, mitochondrial stress contributes to telomere attrition measured by a novel imaging flow cytometry assay. Aging Cell. 2017;16:1234–43.
Chao T, Wang H, Ho PC. Mitochondrial control and guidance of cellular activities of T cells. Front Immunol. 2017;8:473.
Kesarwani P, Murali AK, Al-Khami AA, Mehrotra S. Redox regulation of T-cell function: from molecular mechanisms to significance in human health and disease. Antioxid Redox Signal. 2013;18:1497–534.
Siska PJ, Beckermann KE, Mason FM, Andrejeva G, Greenplate AR, Sendor AB, et al. Mitochondrial dysregulation and glycolytic insufficiency functionally impair CD8 T cells infiltrating human renal cell carcinoma. JCI. Insight. 2017;2.
Buck MD, O'Sullivan D, Klein Geltink RI, Curtis JD, Chang CH, Sanin DE, et al. Mitochondrial dynamics controls T cell fate through metabolic programming. Cell. 2016;166:63–76.
Simula L, Nazio F, Campello S. The mitochondrial dynamics in cancer and immune-surveillance. Semin Cancer Biol. 2017;47:29–42.
Caruana I, Simula L, Locatelli F, Campello S. T lymphocytes against solid malignancies: winning ways to defeat tumours. Cell Stress. 2018;2:200–12.
Xie JH, Li YY, Jin J. The essential functions of mitochondrial dynamics in immune cells. Cell Mol Immunol. 2020;17:712–21.
Baixauli F, Martin-Cofreces NB, Morlino G, Carrasco YR, Calabia-Linares C, Veiga E, et al. The mitochondrial fission factor dynamin-related protein 1 modulates T-cell receptor signalling at the immune synapse. EMBO J. 2011;30:1238–50.
Sun N, Youle RJ, Finkel T. The mitochondrial basis of aging. Mol Cell. 2016;61:654–66.
Pua HH, Guo J, Komatsu M, He YW. Autophagy is essential for mitochondrial clearance in mature T lymphocytes. J Immunol. 2009;182:4046–55.
Thommen DS, Koelzer VH, Herzig P, Roller A, Trefny M, Dimeloe S, et al. A transcriptionally and functionally distinct PD-1(+) CD8(+) T cell pool with predictive potential in non-small-cell lung cancer treated with PD-1 blockade. Nat Med. 2018;24:994–1004.
Kaczanowski S. Apoptosis: its origin, history, maintenance and the medical implications for cancer and aging. Phys Biol. 2016;13:031001.
Pena-Blanco A, Garcia-Saez AJ. Bax, Bak and beyond - mitochondrial performance in apoptosis. FEBS J. 2018;285:416–31.
Sinha K, Das J, Pal PB, Sil PC. Oxidative stress: the mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch Toxicol. 2013;87:1157–80.
Kilbride SM, Prehn JH. Central roles of apoptotic proteins in mitochondrial function. Oncogene. 2013;32:2703–11.
Kim JW, Tsukishiro T, Johnson JT, Whiteside TL. Expression of pro- and antiapoptotic proteins in circulating CD8+ T cells of patients with squamous cell carcinoma of the head and neck. Clin Cancer Res. 2004;10:5101–10.
Hoffmann TK, Dworacki G, Tsukihiro T, Meidenbauer N, Gooding W, Johnson JT, et al. Spontaneous apoptosis of circulating T lymphocytes in patients with head and neck cancer and its clinical importance. Clin Cancer Res. 2002;8:2553–62.
Reichert TE, Strauss L, Wagner EM, Gooding W, Whiteside TL. Signaling abnormalities, apoptosis, and reduced proliferation of circulating and tumor-infiltrating lymphocytes in patients with oral carcinoma. Clin Cancer Res. 2002;8:3137–45.
Bauernhofer T, Kuss I, Henderson B, Baum AS, Whiteside TL. Preferential apoptosis of CD56dim natural killer cell subset in patients with cancer. Eur J Immunol. 2003;33:119–24.
Dworacki G, Meidenbauer N, Kuss I, Hoffmann TK, Gooding W, Lotze M, et al. Decreased zeta chain expression and apoptosis in CD3+ peripheral blood T lymphocytes of patients with melanoma. Clin Cancer Res. 2001;7:947s–57s.
Scheel-Toellner D, Raza K, Assi L, Pilling D, Ross EJ, Lee WY, et al. Differential regulation of nuclear and mitochondrial Bcl-2 in T cell apoptosis. Apoptosis. 2008;13:109–17.
Zorova LD, Popkov VA, Plotnikov EY, Silachev DN, Pevzner IB, Jankauskas SS, et al. Mitochondrial membrane potential. Anal Biochem. 2018;552:50–9.
Girotra M, Thierry AC, Harari A, Coukos G, Naveiras O, Vannini N. Measurement of mitochondrial mass and membrane potential in hematopoietic stem cells and T-cells by flow cytometry. J Vis Exp. 2019.
Nicholls DG. Mitochondrial membrane potential and aging. Aging Cell. 2004;3:35–40.
Yu YR, Imrichova H, Wang H, Chao T, Xiao Z, Gao M, et al. Disturbed mitochondrial dynamics in CD8(+) TILs reinforce T cell exhaustion. Nat Immunol. 2020;21:1540–51.
Derweesh IH, Tannenbaum CS, Rayman PA, Finke JH. Mechanisms of immune dysfunction in renal cell carcinoma. Cancer Treat Res. 2003;116:29–51.
De Maria R, Lenti L, Malisan F, d'Agostino F, Tomassini B, Zeuner A, et al. Requirement for GD3 ganglioside in CD95- and ceramide-induced apoptosis. Science. 1997;277:1652–5.
Gastman BR, Johnson DE, Whiteside TL, Rabinowich H. Tumor-induced apoptosis of T lymphocytes: elucidation of intracellular apoptotic events. Blood. 2000;95:2015–23.
Scorrano L, Petronilli V, Di Lisa F, Bernardi P. Commitment to apoptosis by GD3 ganglioside depends on opening of the mitochondrial permeability transition pore. J Biol Chem. 1999;274:22581–5.
Halestrap AP. What is the mitochondrial permeability transition pore? J Mol Cell Cardiol. 2009;46:821–31.
Sukumar M, Liu J, Mehta GU, Patel SJ, Roychoudhuri R, Crompton JG, et al. Mitochondrial membrane potential identifies cells with enhanced Stemness for cellular therapy. Cell Metab. 2016;23:63–76.
van Bruggen JAC, Martens AWJ, Fraietta JA, Hofland T, Tonino SH, Eldering E, et al. Chronic lymphocytic leukemia cells impair mitochondrial fitness in CD8(+) T cells and impede CAR T-cell efficacy. Blood. 2019;134:44–58.
Wang Y, Tao A, Vaeth M, Feske S. Calcium regulation of T cell metabolism. Curr Opin Physiol. 2020;17:207–23.
Wolf IMA, Guse AH. Ca (2+) microdomains in T-lymphocytes. Front Oncol. 2017;7:73.
Fracchia KM, Pai CY, Walsh CM. Modulation of T cell metabolism and function through calcium signaling. Front Immunol. 2013;4.
Vaeth M, Maus M, Klein-Hessling S, Freinkman E, Yang J, Eckstein M, et al. Store-operated Ca2+ entry controls clonal expansion of T cells through metabolic reprogramming. Immunity. 2017;47:664–+.
Quintana A, Kummerow C, Junker C, Becherer U, Hoth M. Morphological changes of T cells following formation of the immunological synapse modulate intracellular calcium signals. Cell Calcium. 2009;45:109–22.
Meng XH, Wu XY, Zheng YY, Shang K, Jing RR, Jiao P, et al. Exploiting Ca2+ signaling in T cells to advance cancer immunotherapy. Semin Immunol. 2020;49.
Zhou X, Friedmann KS, Lyrmann H, Zhou Y, Schoppmeyer R, Knorck A, et al. A calcium optimum for cytotoxic T lymphocyte and natural killer cell cytotoxicity. J Physiol. 2018;596:2681–98.
Yang PC, Jafri MS. Ca (2+) signaling in T lymphocytes: the interplay of the endoplasmic reticulum, mitochondria, membrane potential, and CRAC channels on transcription factor activation. Heliyon. 2020;6:e03526.
Vaeth M, Kahlfuss S, Feske S. CRAC channels and calcium signaling in T cell-mediated immunity. Trends Immunol. 2020;41:878–901.
Quintana A, Schwindling C, Wenning AS, Becherer U, Rettig J, Schwarz EC, et al. T cell activation requires mitochondrial translocation to the immunological synapse. Proc Natl Acad Sci U S A. 2007;104:14418–23.
Pathak T, Trebak M. Mitochondrial ca (2+) signaling. Pharmacol Ther. 2018;192:112–23.
Trebak M, Kinet JP. Calcium signalling in T cells. Nat Rev Immunol. 2019;19:154–69.
Bernardi P, Rasola A. Calcium and cell death: the mitochondrial connection. Subcell Biochem. 2007;45:481–506.
Singaravelu K, Nelson C, Bakowski D, de Brito OM, Ng SW, Di Capite J, et al. Mitofusin 2 regulates STIM1 migration from the Ca2+ store to the plasma membrane in cells with depolarized mitochondria. J Biol Chem. 2011;286:12189–201.
Uzhachenko R, Ivanov SV, Yarbrough WG, Shanker A, Medzhitov R, Ivanova AV. Fus1/Tusc2 is a novel regulator of mitochondrial calcium handling, Ca2+−coupled mitochondrial processes, and Ca2+−dependent NFAT and NF-kappa B pathways in CD4(+) T cells. Antioxid Redox Signal. 2014;20:1533–47.
Svaton M, Zemanova M, Skrickova J, Jakubikova L, Kolek V, Kultan J, et al. Chronic inflammation as a potential predictive factor of Nivolumab therapy in non-small cell lung Cancer. Anticancer Res. 2018;38:6771–82.
Shao M, Teng X, Guo X, Zhang H, Huang Y, Cui J, et al. Inhibition of calcium signaling prevents exhaustion and enhances anti-leukemia efficacy of CAR-T cells via SOCE-Calcineurin-NFAT and glycolysis pathways. Adv Sci (Weinh). 2022;9:e2103508.
Xiao Y, Yu DH. Tumor microenvironment as a therapeutic target in cancer. Pharmacol Therapeut. 2021;221.
Scharping NE, Rivadeneira DB, Menk AV, Vignali PDA, Ford BR, Rittenhouse NL, et al. Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat Immunol. 2021;22:205–15.
Cheng H, Ma K, Zhang L, Li G. The tumor microenvironment shapes the molecular characteristics of exhausted CD8(+) T cells. Cancer Lett. 2021;506:55–66.
Liu YN, Yang JF, Huang DJ, Ni HH, Zhang CX, Zhang L, et al. Hypoxia induces mitochondrial defect that promotes T cell exhaustion in tumor microenvironment through MYC-regulated pathways. Front Immunol. 2020;11.
Ramalho R, Rao M, Zhang C, Agrati C, Ippolito G, Wang FS, et al. Immunometabolism: new insights and lessons from antigen-directed cellular immune responses. Semin Immunopathol. 2020;42:279–313.
Nachef M, Ali AK, Almutairi SM, Lee SH. Targeting SLC1A5 and SLC3A2/SLC7A5 as a potential strategy to strengthen anti-tumor immunity in the tumor microenvironment. Front Immunol. 2021;12.
Shen L, Xiao Y, Tian J, Lu Z. Remodeling metabolic fitness: strategies for improving the efficacy of chimeric antigen receptor T cell therapy. Cancer Lett. 2022;529:139–52.
Turkcan S, Kiru L, Naczynski DJ, Sasportas LS, Pratx G. Lactic acid accumulation in the tumor microenvironment suppresses F-18-FDG uptake. Cancer Res. 2019;79:410–9.
Wehbi VL, Tasken K. Molecular mechanisms for cAMP-mediated Immunoregulation in T cells - role of anchored protein kinase a signaling units. Front Immunol. 2016;7:222.
Scarfo I, Maus MV. Current approaches to increase CAR T cell potency in solid tumors: targeting the tumor microenvironment. J Immunother Cancer. 2017;5:28.
Ong ST, Ng AS, Ng XR, Zhuang Z, Wong BHS, Prasannan P, et al. Extracellular K(+) dampens T cell functions: implications for immune suppression in the tumor microenvironment. Bioelectricity. 2019;1:169–79.
Wang F, Zhang Y, Yu X, Teng XL, Ding R, Hu Z, et al. ZFP91 disturbs metabolic fitness and antitumor activity of tumor-infiltrating T cells. J Clin Invest. 2021;131.
Xu S, Chaudhary O, Rodriguez-Morales P, Sun X, Chen D, Zappasodi R, et al. Uptake of oxidized lipids by the scavenger receptor CD36 promotes lipid peroxidation and dysfunction in CD8(+) T cells in tumors. Immunity. 2021;54(1561-77):e7.
Manzo T, Prentice BM, Anderson KG, Raman A, Schalck A, Codreanu GS, et al. Accumulation of long-chain fatty acids in the tumor microenvironment drives dysfunction in intrapancreatic CD8+ T cells. J Exp Med. 2020;217.
Bunse L, Pusch S, Bunse T, Sahm F, Sanghvi K, Friedrich M, et al. Suppression of antitumor T cell immunity by the oncometabolite (R)-2-hydroxyglutarate. Nat Med. 2018;24:1192–203.
Martinez M, Moon EK. CAR T cells for solid tumors: new strategies for finding, infiltrating, and surviving in the tumor microenvironment. Front Immunol. 2019;10:128.
Evgin L, Vile RG. Parking CAR T cells in Tumours: oncolytic viruses as valets or vandals? Cancers. 2021;13.
Guo X, Zhang Y, Zheng L, Zheng C, Song J, Zhang Q, et al. Global characterization of T cells in non-small-cell lung cancer by single-cell sequencing. Nat Med. 2018;24:978–85.
Sade-Feldman M, Yizhak K, Bjorgaard SL, Ray JP, de Boer CG, Jenkins RW, et al. Defining T cell states associated with response to checkpoint immunotherapy in melanoma. Cell. 2018;175(998-1013):e20.
Field CS, Baixauli F, Kyle RL, Puleston DJ, Cameron AM, Sanin DE, et al. Mitochondrial integrity regulated by lipid metabolism is a cell-intrinsic checkpoint for Treg suppressive function. Cell Metab. 2020;31(422-37):e5.
Zheng C, Zheng L, Yoo JK, Guo H, Zhang Y, Guo X, et al. Landscape of infiltrating T cells in liver Cancer revealed by single-cell sequencing. Cell. 2017;169(1342-56):e16.
Borcherding N, Vishwakarma A, Voigt AP, Bellizzi A, Kaplan J, Nepple K, et al. Mapping the immune environment in clear cell renal carcinoma by single-cell genomics. Commun Biol. 2021;4:122.
Kashatus DF, Lim KH, Brady DC, Pershing NL, Cox AD, Counter CM. RALA and RALBP1 regulate mitochondrial fission at mitosis. Nat Cell Biol. 2011;13:1108–15.
Peng WS, Zhou X, Yan WB, Li YJ, Du CR, Wang XS, et al. Dissecting the heterogeneity of the microenvironment in primary and recurrent nasopharyngeal carcinomas using single-cell RNA sequencing. Oncoimmunology. 2022;11:2026583.
Yang R, Rincon M. Mitochondrial Stat3, the need for design thinking. Int J Biol Sci. 2016;12:532–44.
Zheng L, Qin S, Si W, Wang A, Xing B, Gao R, et al. Pan-cancer single-cell landscape of tumor-infiltrating T cells. Science. 2021;374:abe6474.
Thommen DS, Schumacher TN. T cell dysfunction in Cancer. Cancer Cell. 2018;33:547–62.
Wang C, Singer M, Anderson AC. Molecular dissection of CD8(+) T-cell dysfunction. Trends Immunol. 2017;38:567–76.
van der Leun AM, Thommen DS, Schumacher TN. CD8(+) T cell states in human cancer: insights from single-cell analysis. Nat Rev Cancer. 2020;20:218–32.
Zhang L, Yu X, Zheng L, Zhang Y, Li Y, Fang Q, et al. Lineage tracking reveals dynamic relationships of T cells in colorectal cancer. Nature. 2018;564:268–72.
Li H, van der Leun AM, Yofe I, Lubling Y, Gelbard-Solodkin D, van Akkooi ACJ, et al. Dysfunctional CD8 T cells form a proliferative, dynamically regulated compartment within human melanoma. Cell. 2019;176(775-89):e18.
Beckermann KE, Hongo R, Ye X, Young K, Carbonell K, Healey DCC, et al. CD28 costimulation drives tumor-infiltrating T cell glycolysis to promote inflammation. JCI. Insight. 2020;5.
Hermans D, Gautam S, Garcia-Canaveras JC, Gromer D, Mitra S, Spolski R, et al. Lactate dehydrogenase inhibition synergizes with IL-21 to promote CD8(+) T cell stemness and antitumor immunity. Proc Natl Acad Sci U S A. 2020;117:6047–55.
Geiger R, Rieckmann JC, Wolf T, Basso C, Feng Y, Fuhrer T, et al. L-arginine modulates T cell metabolism and enhances survival and anti-tumor activity. Cell. 2016;167(829-42):e13.
Chang CH, Qiu J, O'Sullivan D, Buck MD, Noguchi T, Curtis JD, et al. Metabolic competition in the tumor microenvironment is a driver of Cancer progression. Cell. 2015;162:1229–41.
Zhang Y, Kurupati R, Liu L, Zhou XY, Zhang G, Hudaihed A, et al. Enhancing CD8(+) T cell fatty acid catabolism within a metabolically challenging tumor microenvironment increases the efficacy of melanoma immunotherapy. Cancer Cell. 2017;32(377-91):e9.
Chowdhury PS, Chamoto K, Kumar A, Honjo T. PPAR-induced fatty acid oxidation in T cells increases the number of tumor-reactive CD8(+) T cells and facilitates anti-PD-1 therapy. Cancer Immunol Res. 2018;6:1375–87.
Wan H, Xu B, Zhu N, Ren B. PGC-1alpha activator-induced fatty acid oxidation in tumor-infiltrating CTLs enhances effects of PD-1 blockade therapy in lung cancer. Tumori. 2020;106:55–63.
Villena JA. New insights into PGC-1 coactivators: redefining their role in the regulation of mitochondrial function and beyond. FEBS J. 2015;282:647–72.
Fernandez-Marcos PJ, Auwerx J. Regulation of PGC-1alpha, a nodal regulator of mitochondrial biogenesis. Am J Clin Nutr. 2011;93:884S–90.
Li X, Monks B, Ge Q, Birnbaum MJ. Akt/PKB regulates hepatic metabolism by directly inhibiting PGC-1alpha transcription coactivator. Nature. 2007;447:1012–6.
Menk AV, Scharping NE, Rivadeneira DB, Calderon MJ, Watson MJ, Dunstane D, et al. 4-1BB costimulation induces T cell mitochondrial function and biogenesis enabling cancer immunotherapeutic responses. J Exp Med. 2018;215:1091–100.
Crompton JG, Sukumar M, Roychoudhuri R, Clever D, Gros A, Eil RL, et al. Akt inhibition enhances expansion of potent tumor-specific lymphocytes with memory cell characteristics. Cancer Res. 2015;75:296–305.
Noelle RJ, Roy M, Shepherd DM, Stamenkovic I, Ledbetter JA, Aruffo A. A 39-kDa protein on activated helper T cells binds CD40 and transduces the signal for cognate activation of B cells. Proc Natl Acad Sci U S A. 1992;89:6550–4.
Pollizzi KN, Patel CH, Sun IH, Oh MH, Waickman AT, Wen J, et al. mTORC1 and mTORC2 selectively regulate CD8(+) T cell differentiation. J Clin Invest. 2015;125:2090–108.
Lin SJ, Guarente L. Nicotinamide adenine dinucleotide, a metabolic regulator of transcription, longevity and disease. Curr Opin Cell Biol. 2003;15:241–6.
Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, et al. Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science. 2004;305:390–2.
Anderson RM, Barger JL, Edwards MG, Braun KH, O'Connor CE, Prolla TA, et al. Dynamic regulation of PGC-1alpha localization and turnover implicates mitochondrial adaptation in calorie restriction and the stress response. Aging Cell. 2008;7:101–11.
Tilokani L, Nagashima S, Paupe V, Prudent J. Mitochondrial dynamics: overview of molecular mechanisms. Essays Biochem. 2018;62:341–60.
Chan DC. Mitochondrial dynamics and its involvement in disease. Annu Rev Pathol. 2020;15:235–59.
Whitley BN, Engelhart EA, Hoppins S. Mitochondrial dynamics and their potential as a therapeutic target. Mitochondrion. 2019;49:269–83.
Ferreira JCB, Campos JC, Qvit N, Qi X, Bozi LHM, Bechara LRG, et al. A selective inhibitor of mitofusin 1-betaIIPKC association improves heart failure outcome in rats. Nat Commun. 2019;10:329.
Yue W, Chen ZH, Liu HY, Yan C, Chen M, Feng D, et al. A small natural molecule promotes mitochondrial fusion through inhibition of the deubiquitinase USP30. Cell Res. 2014;24:482–96.
L. Miret-Casals, D. Sebastian, J. Brea, E. M. Rico-Leo, M. Palacin, P. M. Fernandez-Salguero, et al. Identification of new activators of mitochondrial fusion reveals a link between mitochondrial morphology and pyrimidine metabolism. Cell Chem Biol 2018;25:268−+.
Qi X, Qvit N, Su YC, Mochly-Rosen D. A novel Drp1 inhibitor diminishes aberrant mitochondrial fission and neurotoxicity. J Cell Sci. 2013;126:789–802.
Kornfeld OS, Qvit N, Haileselassie B, Shamloo M, Bernardi P, Mochly-Rosen D. Interaction of mitochondrial fission factor with dynamin related protein 1 governs physiological mitochondrial function in vivo. Sci Rep. 2018;8:14034.
Rosdah AA, K. Holien J, L. M. Delbridge, G. J. Dusting and S. Y. Lim. Mitochondrial fission - a drug target for cytoprotection or cytodestruction? Pharmacol Res Perspect. 2016;4:e00235.
Simula L, Pacella I, Colamatteo A, Procaccini C, Cancila V, Bordi M, et al. Drp1 controls effective T cell immune-surveillance by regulating T cell migration, proliferation, and cMyc-dependent metabolic reprogramming. Cell Rep. 2018;25(3059-73):e10.
Qian W, Choi S, Gibson GA, Watkins SC, Bakkenist CJ, Van Houten B. Mitochondrial hyperfusion induced by loss of the fission protein Drp1 causes ATM-dependent G2/M arrest and aneuploidy through DNA replication stress. J Cell Sci. 2012;125:5745–57.
Shakibaei M, Sung B, Sethi G, Aggarwal BB. TNF-alpha-induced mitochondrial alterations in human T cells requires FADD and caspase-8 activation but not RIP and caspase-3 activation. Antioxid Redox Signal. 2010;13:821–31.
Soderstrom TS, Poukkula M, Holmstrom TH, Heiskanen KM, Eriksson JE. Mitogen-activated protein kinase/extracellular signal-regulated kinase signaling in activated T cells abrogates TRAIL-induced apoptosis upstream of the mitochondrial amplification loop and caspase-8. J Immunol. 2002;169:2851–60.
Kim WS, Lee KS, Kim JH, Kim CK, Lee G, Choe J, et al. The caspase-8/bid/cytochrome c axis links signals from death receptors to mitochondrial reactive oxygen species production. Free Radic Biol Med. 2017;112:567–77.
Holmstrom TH, Chow SC, Elo I, Coffey ET, Orrenius S, Sistonen L, et al. Suppression of Fas/APO-1-mediated apoptosis by mitogen-activated kinase signaling. J Immunol. 1998;160:2626–36.
Holmstrom TH, Tran SE, Johnson VL, Ahn NG, Chow SC, Eriksson JE. Inhibition of mitogen-activated kinase signaling sensitizes HeLa cells to Fas receptor-mediated apoptosis. Mol Cell Biol. 1999;19:5991–6002.
Cronin SJF, Seehus C, Weidinger A, Talbot S, Reissig S, Seifert M, et al. The metabolite BH4 controls T cell proliferation in autoimmunity and cancer. Nature. 2018;563:564–8.
Sterner RC, Sterner RM. CAR-T cell therapy: current limitations and potential strategies. Blood Cancer J. 2021;11:69.
Ma S, Li X, Wang X, Cheng L, Li Z, Zhang C, et al. Current Progress in CAR-T cell therapy for solid tumors. Int J Biol Sci. 2019;15:2548–60.
Li W, Zhang L. Rewiring mitochondrial metabolism for CD8(+) T cell memory formation and effective Cancer immunotherapy. Front Immunol. 2020;11:1834.
Ah Rad SM, Halpin JC, Mollaei M, Smith Bell SWJ, Hirankarn N, McLellan AD. Metabolic and mitochondrial functioning in chimeric antigen receptor (CAR)-T cells. Cancers (Basel). 2021;13.
Acknowledgments
We thank Ms. Yanling Wan for drawing and authorizing all illustrations in the text.
Funding
This work was financially supported by the National Natural Science Foundation of China [No. 31770131, 81473469], International Cooperation Project of the Belt and Road [SHDC12018119], Shanghai Municipal Commission of Health and Family Plan [No. 20400750600], Shanghai Shen Kang Hospital Development Center Plan [201840056], Shanghai Tenth People's Hospital Project [SYLCYJ06].
Author information
Authors and Affiliations
Contributions
L.Z., W.Z. and Z.L. proposed the study concepts. L.Z., W.Z. and S.L. prepared the manuscript. L.Z., W.Z., T.Z. Y.H., Y.Z. and K.W. edited the manuscript. L.Z., W.Z., C.Q., L.Y. and J.J. were responsible for the manuscript review. All authors have read the final manuscript and approved the version to be published.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, L., Zhang, W., Li, Z. et al. Mitochondria dysfunction in CD8+ T cells as an important contributing factor for cancer development and a potential target for cancer treatment: a review. J Exp Clin Cancer Res 41, 227 (2022). https://doi.org/10.1186/s13046-022-02439-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-022-02439-6