
Abstract
The Hippo signaling pathway was found coordinately modulates cell regeneration and organ size. Its dysregulation contributes to uncontrolled cell proliferation and malignant transformation. YAP/TAZ are two critical effectors of the Hippo pathway and have been demonstrated essential for the initiation or growth of most tumors. Noncoding RNAs (ncRNAs), including miRNAs, lncRNAs, and circRNAs, have been shown to play critical roles in the development of many cancers. In the past few decades, a growing number of studies have revealed that ncRNAs can directly or indirectly regulate YAP/TAZ signaling. YAP/TAZ also regulate ncRNAs expression in return. This review summarizes the interactions between YAP/TAZ signaling and noncoding RNAs together with their biological functions on cancer progression. We also try to describe the complex feedback loop existing between these components.
Similar content being viewed by others


Background
The Hippo signaling pathway was initially discovered in Drosophila and regulates organ growth and cell plasticity during animal development and regeneration [1]. The transcriptional coactivators Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ) are two key effectors that mediate the major gene regulation and biological functions of the Hippo pathway [2]. Importantly, YAP/TAZ exert physiological functions dependent on their transfer from the cytoplasm to the nucleus. Nuclear YAP/TAZ interact with transcription factors, particularly TEA domain (TEAD) family members, to induce the expression of proproliferative and survival-enhancing genes. In recent years, studies have found that YAP/TAZ are hyperactivated in human cancers and that excessive activation of YAP/TAZ correlates with poor outcome in several cancer types [3,4,5,6]. YAP/TAZ, as oncogenes, are known to be involved in cancer cell proliferation, invasion, and metastasis; epithelial-mesenchymal transition (EMT); and resistance to anticancer drugs [7]. Treating tumor cells with YAP/TAZ inhibitors, such as verteporfin, significantly reversed the malignant biological behavior of the tumor cells [8,9,10,11], indicating the therapeutic value of targeting YAP/TAZ signaling in cancer. Notably, an increasing number of studies have reported the effects of YAP/TAZ in the tumor microenvironment, which further emphasizes the essential role of YAP/TAZ in tumorigenesis. Therefore, understanding the regulation and mechanisms of YAP/TAZ signaling in cancer is of great importance.
Noncoding RNAs (ncRNAs) are transcripts that do not encode proteins and are classified as microRNAs (miRNAs), circular RNAs (circRNAs), and long noncoding RNAs (lncRNAs). Although ncRNAs lack protein-coding capacity, subsequent studies have shown that ncRNAs are key regulators mediating many fundamental cellular processes, such as differentiation, proliferation, apoptosis, and cell metabolism. NcRNAs can act as archetypical decoys, signals, guides, and scaffolds to mediate mRNA transcription and translation, protein abundance and localization, and chromatin and protein conformation [12,13,14,15]. Additionally, a limited number of ncRNAs that can encode peptides with biological and pathological functions have been validated [16]. A large body of evidence has demonstrated that ncRNAs are important versatile molecules involved in various tumorigenic processes. In vivo, ncRNAs are remarkably stable and are thus potential biomarkers for the diagnosis and prognosis of cancer patients [17,18,19,20,21].
The major roles of ncRNAs in YAP/TAZ signaling, such as regulating the transcription, localization, and stability of YAP/TAZ, has been highlighted by recent studies. In turn, some studies have reported that YAP/TAZ are upstream mediators of ncRNAs. In this review, we discuss the crosstalk between YAP/TAZ and ncRNAs and describe their biological functions in cancers. We expect that YAP/TAZ-associated ncRNAs could be molecular targets and prognostic biomarkers in various cancers.
The Hippo signaling pathway
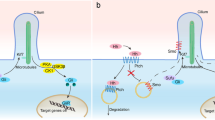
The core of the classical Hippo pathway comprises two Ser/Thr kinases: Ste20-like kinase 1/2 (Mst1/2) and large tumor suppressor kinase 1/2 (LATS1/2). MST1/2, aided by its partner Salvador1 (SAV1), stimulate LATS1/2 and the LATS cofactor MOB1 [22]. Subsequently, the serine/threonine-protein kinase LATS1 directly phosphorylates YAP and TAZ, facilitating their cytoplasmic localization or degradation. Indeed, LATS-mediated phosphorylation at YAP Ser127 generates a unique 14–3-3 protein binding site leading to YAP cytoplasmic localization [23]. YAP is phosphorylated not only at Ser127 but also at Ser381 by LATS1/2, which primes YAP for subsequent phosphorylation by casein kinase 1 (CK1) delta/epsilon in a phosphodegron. The phosphorylated phosphodegron then recruits the β-TrCP E3 ubiquitin ligase, which catalyzes the ubiquitination of YAP, ultimately leading to its proteasomal degradation [24]. Once the Hippo pathway is inactivated, YAP/TAZ accumulate in the nucleus and bind to transcription factors such as TEAD1-4/TEF, p73, RUNX, or SMAD to promote the expression of downstream genes [25].
Continuously accumulating research has identified additional regulators that should be considered novel members of the Hippo core cassette. For instance, Ras association domain family protein 1 isoform A (RASSF1A), a critical upstream regulator of the core Hippo pathway, binds to MST1/2 kinases and SAV1 to activate kinase cascade signaling, leading to YAP/TAZ phosphorylation and inhibition [26]. As another example, nuclear Dbf2-related 1/2 (NDR1/2) kinases, which share a similar NDR domain with LATS1/2 and are phosphorylated by MST1/2 and MOB1, can function as components in an extended Hippo pathway. Active MST1/2 can inhibit YAP/TAZ signaling through NDR1/2-mediated phosphorylation of YAP/TAZ [27]. Notably, Nemo-like kinase (NLK) can phosphorylate YAP at Ser128, blocking its interaction with 14–3-3 and enhancing its nuclear localization [28, 29]. Furthermore, actin-like 6A (ACTL6A) was found to physically interact with YAP/TAZ and disrupt the interaction between YAP and the E3 ubiquitin ligase β-TrCP, thus protecting the YAP protein from degradation [30]. In addition, AJUBA is a multiple LIM domain-containing protein that tends to interact with a large number of proteins. AJUBA has been shown to physically interact with LATS1/2 and inhibit YAP phosphorylation via sequestration of LATS1/2 [31].
YAP/TAZ can also be regulated by other mechanisms in addition to the Hippo signaling pathway (Fig. 1). It has long been appreciated that cell behavior can be influenced by mechanical cues such as the stiffness of the ECM. YAP/TAZ have been acknowledged to be critical mediators of mechanotransduction pathways. Dupont found that YAP/TAZ were nuclear and active in cells grown on a stiff extracellular matrix (ECM), whereas they were predominantly cytoplasmic and inactive in cells grown on a soft ECM. These mechanical cues affect YAP/TAZ nuclear localization and activity through a cytoskeletal pathway that requires Rho, a small GTPase that regulates the formation of actin bundles and stress fibers [32]. A recent study reported that low ECM stiffness induced YAP/TAZ cytoplasmic localization by activating MST1/2 via the small GTPase RAP2, suggesting the involvement of both Hippo-dependent and Hippo-independent mechanisms in mechanoregulated YAP/TAZ signaling [33].

Pathway crosstalk regulates YAP and TAZ YAP/TAZ signaling is mainly regulated by the Hippo signaling pathway. Activation of the Hippo pathway is associated with phosphorylation of the core Hippo kinases MST1/2 and LATS1/2, and their cofactors SAV1 and MOB. Phosphorylated YAP/TAZ are exported from the nucleus and degraded in the cytoplasm. When these kinases are inactive, YAP/TAZ accumulate in the nucleus, bind to transcription factors, and promote the expression of target genes. Additional regulators that compose the Hippo pathway have been identified. Moreover, YAP/TAZ signaling is related to mechanotransduction pathways and is regulated by mechanical forces through the actin cytoskeleton
The function of YAP/TAZ in cancer
Extensive research has demonstrated that YAP/TAZ are involved in cancer cell proliferation, metastasis and invasion; EMT; the acquisition of cancer stem cell (CSC) properties; and drug resistance [34,34,36]. In fact, YAP/TAZ have been identified as signaling hubs in complex networks of various oncogenic signaling pathways such as the EGFR, TGFβ, Wnt, PI3K, GPCR, and KRAS pathways, many of which are considered to be hallmarks of cancer [37, 38]. YAP/TAZ are often also involved in metabolic regulation, such as the promotion of glycolysis, lipogenesis, and glutaminolysis, suggesting that YAP/TAZ are emerging nodes in the provision of energy and necessary synthetic materials for the activities of tumor cells. In addition, YAP/TAZ can regulate the production of secretory proteins, such as amphiregulin (AREG; an epidermal growth factor (EGF) family member), cysteine-rich angiogenic inducer 61 (CYR61), and connective tissue growth factor (CTGF), to influence the tumor microenvironment [39]. Several recent studies have shown that YAP/TAZ mediate the expression of PD-L1, which is involved in cancer immune evasion [40].
Conferring cancer cell stemness is probably one of the best characterized YAP/TAZ-dependent biological responses. CSCs are cancer cells that possess the same self-renewal and differentiation capacities as stem cells, thereby maintaining the ability to regenerate a heterogeneous tumor mass [41]. Recent studies have suggested that CSCs are responsible for tumor initiation, progression, and therapeutic resistance. YAP/TAZ activation can endow cell plasticity by reprogramming genetically normal primary differentiated cells into the corresponding tissue-specific stem or progenitor cells [42]. Many studies have reported that active YAP and TAZ convert benign neoplastic cells into cancer stem cells, contributing to the uncontrolled proliferation, survival, chemoresistance, and metastasis of cancer cells [43,43,45].
Together, these studies reveal the oncogenic role of YAP/TAZ signaling and provide evidence for the therapeutic benefits of YAP/TAZ inhibition in cancer. Therefore, we attempt to provide an overview of different options that can be utilized to target YAP/TAZ for therapeutic interventions in cancer.
The crosstalk between YAP/TAZ and microRNAs
Regulation of YAP/TAZ by microRNAs
MiRNAs are well-known small ncRNAs of 20 to 24 nucleotides in length that regulate the expression of target RNAs through imperfect base pairing with the 3’-untranslated region (3’-UTR) of the target RNAs. A large number of YAP/TAZ-related miRNAs have been discovered in cancer. MiRNAs appear to regulate the YAP/TAZ signaling pathway by targeting YAP/TAZ directly or in a Hippo pathway-dependent manner. Table 1 lists some miRNAs regulating YAP/TAZ expression in cancers.
MicroRNAs regulate YAP/TAZ by directly targeting YAP/TAZ mRNA
Mature miRNAs bind to the 3-’UTRs of YAP/TAZ, resulting in their translational inhibition or mRNA degradation. This is the case for miR-550a-3-5p, which is a tumor suppressor targeting YAP directly in various cancers, including oral squamous cell carcinoma, colon cancer, and melanoma. As elucidated by Cao et al., miR-550a-3-5p directly targets YAP and then reduces YAP/TEAD-mediated C–C motif chemokine ligand 2 (CCL2) expression in HPV-positive oral squamous cell carcinoma cells [60]. Upon downregulation of CCL2, miR-550a-3-5p inhibited M2 macrophage polarization, leading to suppression of migration, invasion, and EMT in HPV-positive oral squamous cell carcinoma. Similar phenomena occur in vemurafenib-resistant colon cancer and melanoma cells. MiR-550a-3-5p was found to directly target YAP to inhibit its expression, and overexpression of miR-550a-3-5p sensitized vemurafenib-resistant colon cancer and melanoma cells to vemurafenib [61]. It is suggested that miRNA-mediated YAP inhibition may be a new strategy for cancer treatment. In the next sections, we summarize miRNAs that target YAP/TAZ directly in different cancers.
In breast cancer, miR-27b-3 was found to induce apoptosis while attenuating cancer cell metastasis and invasion. It was reported that miR-27b-3 bound to the 3′-UTR of YAP to reduce its expression. Furthermore, the researchers showed that the local anesthetic ropivacaine suppressed breast cancer progression by regulating the miR-27b-3p/YAP axis[62]. Intriguingly, the association between miR-205 and YAP1 has been explored in activated cancer-associated fibroblasts (CAFs) of breast tumors. Enhancing miRNA-205 expression in CAFs significantly inhibited the angiogenic process via downregulation of YAP1 [63]. MiRNAs and YAP/TAZ play significant roles in proliferation, metastasis, and therapy response of breast cancer.
Increasing evidence suggests that YAP/TAZ signaling activation favors hepatocellular carcinoma progression. Higashi et al. reported that miR-9-3p functioned as a tumor suppressor that inhibited hepatocellular carcinoma cell proliferation by targeting TAZ [64]. As mentioned above, YAP/TAZ signaling can control chemoresistance in cancers. Chen et al. identified YAP1, which was directly targeted by miR-590-5p, as the major dysregulated molecule in adriamycin-resistant hepatocellular carcinoma cells. Their findings further strengthened the evidence that miR-590-5p significantly inhibited the expression of the stem cell markers OCT4, SOX-2, Notch-1, Nanog, and Nestin to endow chemotherapeutic resistance in hepatocellular carcinoma [65]. Similar to miR-590-5p, miR-375 was found to be downregulated in cisplatin-resistant liver cancer cells and also suppressed cisplatin-induced resistance in liver cancer cells [66]. Downregulation of miR-375 activated YAP signaling, resulting in increases in IL-6 and TGF-β expression.
The miRNA/YAP axis has been shown to be a potential therapeutic target in lung cancer. In lung cancer tissue specimens and cell lines, miR-381 was found to be significantly downregulated, which decreased the expression of its direct target YAP. Enhancing miR-381 expression inhibited non-small-cell lung cancer (NSCLC) cell proliferation, invasion, migration, and EMT. In addition, metformin inhibited lung cancer cell growth and metastasis via the miR-381/YAP axis [67]. MiR-7 is another tumor suppressor and reverses gefitinib resistance by directly targeting YAP in NSCLC cells. Notably, miR-7 was downregulated in serum exosomes from patients with lung carcinoma compared with healthy controls, and miR-7 expression was negatively correlated with the response to gefitinib treatment in lung carcinoma patients as well as to their overall survival [68].
MiR-195-5p is a tumor suppressor in colon cancer, inhibiting colon cancer cell proliferation, migration, invasion, and EMT [69]. MiR-195-5p can bind to the 3’-untranslated region (3’-UTR) of YAP1 mRNA and downregulate its expression. Another miRNA targeting YAP directly, miR-375, has been identified in colorectal cancer. MiR-375 was found to be downregulated in colorectal cancer cell lines and tissues. Overexpression of miR-375 derepressed the expression of YAP and partially reversed resistance to 5-fluorouracil in colorectal cancer cells. The miR-375/YAP axis regulates apoptosis and cell cycle regulators, including cyclin D1 and BIRC5, as well as CTGF, a downstream effector of the Hippo pathway [70].
On the other hand, miR-375 was found to be inversely correlated with EMT signatures in prostate cancer clinical samples. The EMT process is a reversible biological process whereby epithelial cells transition to a mesenchymal phenotype. The reverse process is called mesenchymal-epithelial transition (MET). Indeed, miR-375 inhibited the invasion and migration of prostate cancer cells by facilitating the MET process. YAP1 was identified as a direct target of miR-375 in prostate cancer, and overexpression of YAP1 abrogated the inhibitory effect of miR-375 on prostate cancer cells [71].
Similar to its effect in colorectal cancer, YAP/TAZ overexpression promotes gastric cancer progression. MiR-15a and miR-16–1 could be considered diagnostic factors for gastric cancer, reducing YAP1 expression to inhibit the proliferation, monolayer colony formation, invasion, and migration of gastric adenocarcinoma cells [72].
MicroRNAs regulate the stability or nuclear translocation of the YAP/TAZ proteins
Another mechanism by which miRNAs participate in YAP/TAZ regulatory networks is through effects on the stability or nuclear translocation of the YAP/TAZ proteins. However, the shuttling of YAP/TAZ between the cytoplasm and nucleus is mainly achieved in a phosphorylation-dependent manner. Therefore, these interactions rely primarily on the Hippo pathway or other genes, which have indirect effects.
MiR-103a-3p and miR-429 can directly target LATS2 in colorectal cancer, reducing the phosphorylation of YAP and increasing its nuclear localization. Furthermore, the miR-103a-3p/YAP axis was found to promote the expression of hypoxia-inducible factor 1 alpha (HIF1A), consequently enhancing glycolysis and angiogenesis in colorectal cancer cells [73, 74]. RANBP1 functions as a tumor-promoting factor in colorectal cancer, and it disrupts the Hippo pathway to hyperactivate YAP signaling. It was reported that RANBP1 regulates the Hippo pathway by promoting the processing of precursor miRNAs, including the pri-miRNAs of miR-18a, miR-183, and miR-106 [75].
MiR-25 and miR-107 were found to be expressed at high levels in NSCLC cells compared with normal cells. Through binding to the 3’-UTR of LATS2, miR-25 and miR-107 downregulated its expression. Owing to downregulation of LATS2, YAP signaling was activated to promote the growth and metastasis of NSCLC cells [76, 77]. Moreover, miR-135b, which was upregulated in highly invasive NSCLC cells, was found to improve the migration and invasion abilities of NSCLC in vitro and in vivo. A mechanistic study revealed that miR-135b targeted multiple critical components in the Hippo pathway, including LATS2, β-TrCP, and NDR2, to stabilize the TAZ protein [78].
A similar phenomenon occurs in prostate cancer with upregulation of the miR-302/367 cluster, which consists of miR-302a, miR-302b, miR-302c, miR-302d, and miR-367. The miR-302/367 cluster was found to directly target LATS2 to reduce the phosphorylation of YAP and enhance its nuclear translocation. Functionally, the miR-302/367 cluster enhanced the proliferation, sphere formation, and migration of prostate cancer cells and also drove their resistance to androgen ablation [79].
Myosin phosphatase targeting protein 1 (MYPT1), an upstream regulator of the Hippo pathway, can activate the kinase cascade, resulting in YAP/TAZ inhibition. MiR-30b was found to activate YAP signaling by directly targeting MYPT1, leading to enhanced CSC-like properties as well as resistance to platinum-based therapy in ovarian cancer cells [80].
Wiskott–Aldrich syndrome protein-interacting protein family member 1 (WIPF1) was found to protect YAP/TAZ from proteolysis in a Hippo pathway-independent manner. WIPF1 increased the sequestration of the β-catenin adenomatous polyposis coli (APC)-axin-GSK3 destruction complex to the multivesicular body compartment to reduce the capacity of the complex to degrade YAP/TAZ [81]. Pan et al. demonstrated that miR-141 and miR-200c suppress YAP/TAZ expression by repressing the expression of WIPF1 in pancreatic ductal adenocarcinoma cells [82]. The functional role of miR-141 and miR-200c is to inhibit the EMT process and further suppress cell migration and invasion in pancreatic ductal adenocarcinoma.
Exosomes of endosomal origin are nanoscale membrane vesicles (with a size of 30 ~ 100 nm) that coexist with microvesicles and apoptotic bodies in the extracellular microenvironment [83]. Exosomes can transfer various proteins, DNAs, and RNAs to mediate cell communication. MiR-181a, which was found to be highly expressed in papillary thyroid cancer (PTC) cells, could be delivered to human umbilical vein endothelial cells (HUVECs) by hypoxia-induced exosomes. Exosomal miR-181a promoted the proliferation, migration, and angiogenesis of PTC cells. Specifically, miR-181a inhibited Dishevelled binding antagonist of beta catenin 2 (DACT2) by targeting targeted mixed lineage leukemia 3 (MLL3) [84, 85]. In this way, miR-181a reduced the phosphorylation of YAP and activated the YAP-VEGF axis in HUVECs.
Many studies have reported alternative modulators of YAP/TAZ subcellular localization in addition to the 14–3-3 protein, such as Mastermind-like 1/2 (MAML1/2). MAML1/2 interact with YAP/TAZ through a PPxY motif and WW domain to promote the nuclear localization of YAP/TAZ. Kim et al. showed that miR-30c targeted MAML1 and inhibited MAML1 synthesis in cancer cells, leading to reduced YAP/TAZ nuclear localization [86].
MicroRNAs regulate transcription of YAP/TAZ
In addition to their fundamental interactions with the 3’-UTR of YAP/TAZ or the core components of the Hippo signaling pathway, miRNAs can exert regulatory impacts on transcription factors and affect the transcription of YAP/TAZ.
The cAMP response element-binding (CREB) protein is a CREB/activating transcription factor family member. CREB binds to the promoter of YAP and increases YAP transcriptional output [87]. MiR-1224, functioning as a tumor suppressor, was found to be downregulated in hepatocellular carcinoma and to inhibit the proliferation of hepatocellular carcinoma cells both in vitro and in vivo. By binding to CREB, miR-1224 repressed its transcription and subsequent activation of the YAP signaling pathway in hepatocellular carcinoma cells [88].
RFX5 was identified to bind to the YAP promoter and induce the transcriptional activation of YAP. It was reported that an RNA-binding protein, LIN28, promoted RFX5 mRNA stability. In NSCLC cells, miR-4319 bound to LIN28 and negatively regulated its expression, leading to destabilization of RFX5, which further impeded YAP expression [89]. MiR-4319 thus exerted a tumor-suppressive effect in NSCLC, inhibiting NSCLC cell proliferation and migration while inducing NSCLC cell apoptosis.
Regulation of microRNAs by YAP/TAZ
Previously, we described the role of miRNAs as upstream mediators of YAP/TAZ in cancer progression. Notably, studies have demonstrated that YAP/TAZ can participate in regulating miRNA expression and result in tumorigenesis.
In general, YAP/TAZ act as coactivators and cooperate with other transcription factors to mediate downstream gene transcription. The study conducted by Ma’s team revealed that TAZ/TEAD can bind to the promoter of miR-224 and facilitate the expression of miR-224. Furthermore, the TAZ/miR-224 axis inhibited the tumor suppressor SMAD4 to promote the proliferation and migration of osteosarcoma cells [90].
Interestingly, researchers have identified YAP and TAZ as regulators of global miRNA biogenesis via modulation of miRNA-processing enzymes, microprocessor, or the Dicer complex [91]. Mori et al. reported that YAP regulates miRNA biogenesis in a cell density-dependent manner. At a low cell density, nuclear YAP binds and sequesters p72, a Microprocessor component, causing widespread miRNA suppression in cells. At a high cell density, YAP is inactivated by exclusion from the cell nucleus, thereby allowing p72 to associate with microprocessor and pri-miRNAs, resulting in enhanced miRNA biogenesis [92]. However, this finding is controversial. Chaulk et al. reached the opposite conclusion that nuclear TAZ/YAP, which are abundant at a low cell density, are required for efficient pre-miRNA processing. Knockdown of TAZ/YAP in low-density cells or density-mediated sequestration of TAZ/YAP into the cytoplasm reduced the level of LIN28. The decreased level of LIN28 led to accumulation of let-7a and let-7b miRNA, and let-7a and let-7b miRNA downregulated Dicer, resulting in defective processing of pre-miRNAs [93]. This research suggested that YAP increases miRNA expression by supporting Dicer-mediated pre-miRNA processing, and this idea was further verified by Zhang’s team. Zhang et al. demonstrated that YAP negatively regulated p21 expression by promoting the expression of the miR-17 family. Upon upregulation of p21, loss of YAP1 inhibited chondrosarcoma cell proliferation and induced senescence in chondrosarcoma cells [94].
MiRNAs are frequently derived from protein-coding gene introns and are located in transcripts coding for multiple products [95]. MiR-25, miR-93, and miR-106b can bind to the 3′-UTR of P21 and inhibit its expression. YAP/TAZ was found to induce the transcription of the MCM7 gene to enhance the expression of its host miRNAs (miR-25, miR-93, and miR-106b) in NSCLC cells, thereby promoting cell proliferation through inhibition of p21 [96].
The feedback loop between YAP/TAZ and microRNAs
It is noteworthy that quite a few miRNAs participate in reciprocal feedback loops with the YAP/TAZ signaling pathway (Fig. 2). Shen et al. reported that miR-135b-5p induced by TAZ suppressed glycogen synthase kinase 3β, resulting in EMT induction and promoting the proliferation and invasion of osteosarcoma cells. Moreover, miR-135-5p targeted LATS2 to increase TAZ activity in osteosarcoma cells [97]. The TAZ/miR-135b positive feedback loop played a crucial role in osteosarcoma progression. The positive feedback loop between TAZ and miR-942-3p in bladder cancer is another example. MiR-942-3p was found to be abundantly expressed in bladder cancer, and its upregulation was mediated by TAZ. In turn, overexpression of miR-942-3p amplified TAZ signaling by targeting LATS2, eventually promoting proliferation, angiogenesis, EMT, glycolysis, and reactive oxygen species (ROS) homeostasis in bladder cancer [98].

Feedback loops between YAP/TAZ and microRNAs
MiR-19b-3p was suggested to promote lung cancer progression. In detail, exosomal miR-19b-3p was secreted from lung adenocarcinoma cells and taken up by tumor-associated macrophages, leading to the activation of STAT3 signaling. Activated STAT3 signaling induced M2 macrophage polarization and promoted LINC00273 transcription in M2 macrophages. In contrast, M2 macrophages transferred exosomal LINC00273 to lung adenocarcinoma cells. Investigation of molecular pathways demonstrated that LINC00273 recruits NEDD4 to facilitate LATS2 ubiquitination and degradation, leading to the activation of YAP signaling. In turn, activated YAP induced the transcription of RNA-binding motif protein encoded on the X chromosome (RBMX). By binding to miR-19b-3p, RBMX facilitated the packaging of miR-19b-3p into LUAD cell-derived exosomes [99]. Collectively, the miR-19b-3p/LINC00273/YAP feedback loop contributed to the communication between lung cancer cells and tumor-associated macrophages.
It was confirmed that VGLL4 is a YAP signaling inhibitor that competes with YAP for TEAD binding. MiR-130a, which is directly induced by YAP, activated YAP by suppressing VGLL4 expression. YAP signaling was sustained through this miR-130a-dependent positive feedback loop, leading to tumorigenesis and cancer development [100].
The crosstalk between YAP/TAZ and lncRNAs in cancer
Regulation of YAP/TAZ by lncRNAs
LncRNAs are a class of noncoding RNAs with a transcript length of more than 200 nucleotides. Aberrant expression of lncRNAs has been reported to contribute to the occurrence and development of multiple cancers. Recently, a continuously increasing number of YAP/TAZ-associated lncRNAs have been discovered. LncRNAs can act as signals, decoys, guides, and scaffolds by interacting with proteins or other RNA molecules. LncRNAs can function as miRNA sponges to prevent the association of miRNAs with downstream mRNAs. Some lncRNAs bind to transcription factors to regulate gene expression. Additionally, lncRNAs can regulate gene expression via histone modification and chromatin remodeling. Table 2 lists some lncRNAs regulating YAP/TAZ expression in cancers.
LncRNAs regulate YAP/TAZ signaling by functioning as ceRNAs
The classical mechanism by which lncRNAs participate in YAP/TAZ signaling is through binding to miRNAs as competitive endogenous RNAs (ceRNAs), thereby affecting the expression of YAP/TAZ or various executors in the Hippo pathway.
Yan et al. reported that the lncRNA FLVCR1-AS1 level was elevated in ovarian serous cancer (OSC) tissues and sera. Functionally, FLVCR1-AS1 increased OSC cell proliferation, migration, invasion, and EMT and decreased OSC cell apoptosis in vitro and in vivo. Further study showed that FLVCR1-AS1 activated YAP signaling by acting as a decoy for miR-513 that bound to the 3’ UTR of YAP [122]. Similarly, the lncRNA MLK7-AS1 affects YAP/TAZ signaling in ovarian cancer by regulating the YAP level via the ceRNA mechanism. The lncRNA MLK7-AS1 promoted proliferation, metastasis, and the EMT process via the miR-375/YAP axis in ovarian cancer [123]. Furthermore, the lncRNA MIR205HG was found to deplete endogenous miR-590-3p to increase YAP expression, inducing uncontrolled proliferation of head and neck cancer cells [124]. Additionally, Wu et al. reported that the lncRNA SNHG15 sponged miR-200a-3p to positively modulate YAP1 expression. Knockdown of SNHG15 inhibited proliferation, migration, and EMT and promoted apoptosis in papillary thyroid carcinoma cells [125].
Importantly, lncRNA–miRNA networks can regulate YAP/TAZ signaling by affecting upstream mediators in the Hippo pathway. For example, the lncRNA ASMTL-AS1 sequesters miR-342-3p to enhance the expression of NLK, which can phosphorylate YAP at Ser128, leading to enhanced nuclear translocation of YAP [126]. The lncRNA ASMTL-AS1 was found to be highly expressed in hepatocellular carcinoma tissues, especially in residual hepatocellular carcinoma tissues after insufficient radiofrequency ablation, and its expression level was related to stage and prognosis in hepatocellular carcinoma. Another study revealed the role of the lncRNA MRVI1-AS1 in nasopharyngeal cancer chemosensitivity by inhibiting TAZ activation. Mechanistically, the lncRNA MRVI1-AS1 inhibited the expression of miR-513a-5p and miR-27b-3p by sponging them to upregulate ATF3 (activating transcription factor 3) expression. Furthermore, the MRVI1-AS1/ATF3 axis promoted the expression of RASSF1 (Ras association domain family member 1), a Hippo pathway regulatory factor, thereby inhibiting TAZ expression [127].
Overall, these studies suggest the regulatory roles of lncRNA–miRNA networks in YAP/TAZ signaling in different cancers.
LncRNAs regulate YAP/TAZ signaling by affecting protein–DNA interactions
Another mechanism by which lncRNAs regulate gene expression is through binding to transcription factors to mediate DNA transcription. By binding to C-Myc at the YAP1 promoter, the lncRNA RP11-323N12.5 promotes YAP1 expression in gastric cancer. Furthermore, the lncRNA RP11-323N12.5 was found to mediate immunosuppression by upregulating YAP-induced immunosuppression-related genes, including PSTG2, CSF1, CXCL15, and CSF3 [128]. LINC01048 is another tumor promoter that enhances proliferation while inhibiting apoptosis in cutaneous squamous cell carcinoma cells by increasing YAP1 expression. Further investigation revealed that LINC01048 recruits TAF15 (a transcriptional activator) to the YAP1 promoter to transcriptionally activate YAP1 in cutaneous squamous cell carcinoma cells [129].
Previously, the AJUBA protein was shown to activate YAP signaling by inhibiting MST1/2 and LATS1. The lncRNA MNX1-AS1, which was found to be elevated in intrahepatic cholangiocarcinoma cell lines and tissues, facilitated proliferation, migration, invasion, and angiogenesis in intrahepatic cholangiocarcinoma [130]. Mechanistically, the lncRNA MNX1-AS1 recruited C-Myc and MAZ to the nucleus to facilitate the expression of MNX1. Then, MNX1 promoted the expression of the AJUBA protein by binding to the AJUBA promoter region. Owing to the increased expression of AJUBA, YAP signaling was activated in intrahepatic cholangiocarcinoma.
LncRNAs regulate YAP/TAZ signaling by affecting protein–RNA interactions
LncRNAs also play an essential role in posttranscriptional regulation by interacting with RNA-binding proteins. HuR, a nucleocytoplasmic shuttling protein, can regulate the stability of its target mRNAs [131]. The lncRNA B4GALT1-AS1, whose expression was found to be significantly increased in osteosarcoma tissues and cell lines, promoted proliferation, migration, stemness, and adriamycin-therapeutic sensitivity in osteosarcoma. Mechanistically, the lncRNA B4GALT1-AS1 enhanced YAP mRNA stability by recruiting HuR [132].
LncRNAs regulate YAP/TAZ signaling by affecting protein–chromatin interactions
Chromatin is a macromolecular complex comprising DNA, histone proteins, and RNA [133]. Methylation of histone lysines is a core mechanism that affects gene expression and genome stability. Liu et al. reported that the lncRNA HOTTIP maintained osteosarcoma cell viability, proliferation, migration, and invasion in vitro and contributed to xenograft growth or lung metastasis in vivo. Mechanistically, the lncRNA HOTTIP acted as a scaffold to recruit enhancer of zeste homolog 2 (EZH2) and lysine‐specific demethylase 1 (LSD1) to stimulate histone H3 lysine 27 trimethylation and inhibit histone H3 lysine 4 dimethylation of LATS2. Generally, histone H3 lysine 27 trimethylation contributes to gene silencing, whereas histone H3 lysine 4 dimethylation is associated with gene activation [134, 135]. Via this mechanism, LATS2 was epigenetically silenced, leading to YAP signaling activation [136].
LncRNAs regulate YAP/TAZ signaling by driving aberrant liquid–liquid phase separation
The formation of biomolecular condensates via liquid–liquid phase separation (LLPS) has recently emerged as a widespread mechanism underlying the spatiotemporal coordination of biological activities in cells [137]. LncRNAs contain repetitive sequences and thus contribute to LLPS [138]. Recently, a study revealed the lncRNA SNHG9 as a tumor-promoting factor in breast cancer, and the expression of SNHG9 was positively correlated with breast cancer progression. Mechanistically, the lncRNA SNHG9 and its associated phosphatidic acids (PA) interact with the C-terminal domain of LATS1, driving liquid droplet formation of LATS1. Furthermore, LATS1 kinase activity was impaired by LLPS promoted by SNHG9 and PA, resulting in reduced YAP phosphorylation and cytoplasmic translocation [139].
Regulation of lncRNAs by YAP/TAZ
As has been shown for miRNAs, some studies have demonstrated that YAP/TAZ can regulate lncRNAs and result in cancer progression. For instance, the lncRNA H19, an oncogenic mediator in various cancers, was found to be upregulated by YAP in osteosarcoma [140]. Another study showed that highly expressed YAP facilitated the expression of the lncRNA BCAR4, which coordinated Hedgehog signaling to enhance the transcription of the glycolysis activators HK2 and PFKFB3, thus reprogramming glucose metabolism and conferring invasiveness and a propensity for metastasis on triple-negative breast cancer cells [141, 142]. In colorectal cancer, the lncRNA MALAT1 was found to promote angiogenesis and the EMT process by sponging miR-126-5p to increase VEGFA, Slug, and Twist expression, and LINC00958 was positively regulated by YAP1, which promoted the transcription of LINC00958 by forming a complex with β-catenin/transcription factor 4 that bound to the MALAT1 promoter [143].
Although YAP/TAZ are known to function as transcriptional coactivators, recent transcriptomic studies have revealed that they can also suppress gene expression. Their transcriptional repressive function depends on TEADs and the NuRD complex, and the latter promotes histone deacetylation and nucleosome occupancy at target genes[144]. Tan et al. showed that the lncRNA NORAD was downregulated in lung and breast cancers and that low expression of the lncRNA NORAD in these cancer types was associated with lymph node metastasis and poor prognosis. The YAP/TAZ-TEAD-NuRD complex transcriptionally repressed NORAD expression, which contributed to YAP/TEAD-induced tumor migration and invasion [145].
Feedback loops between lncRNAs and YAP/TAZ
Feedback loops between YAP/TAZ and lncRNAs have also received attention (Fig. 3). In primary renal tumor-initiating cells (T-ICs), lncARSR was found to be transcriptionally upregulated by YAP; afterward, the increased lncARSR bound specifically to the WW1/2 domain of YAP and protected YAP from phosphorylation by LATS1, thus forming a positive feedback loop. According to the study by Qu et al., the YAP/lncARSR interaction enhanced the self-renewal capacity, tumorigenicity, and metastasis of renal T-ICs [146]. Similarly, in pancreatic ductal adenocarcinoma cells, the YAP-induced lncRNA THAP9-AS1 stimulated YAP signaling activation by sponging miR-484 and binding to the YAP protein to inhibit LATS1-mediated YAP phosphorylation, eventually enhancing YAP expression at both the posttranscriptional and posttranslational levels [147]. Additionally, a study showed that the lncRNA SFTA1P bound to TAZ to promote its nuclear translocation, resulting in YAP/TAZ signaling activation, in NSCLC cells. Based on the mutually promotive relationship between the lncRNA SFTA1P and TAZ, they were integrated into a positive feedback loop that induced proliferation and inhibited programmed cell death in NSCLC cells [148] Figs. 2 and 3.

Feedback loops between YAP/TAZ and lncRNAs
Regarding the role of the lncRNA GAS5 as a tumor suppressor in colorectal cancer, Ni et al. uncovered a negative feedback loop of the lncRNA GAS5-YAP-YTHDF3 axis in the progression of colorectal cancer. The lncRNA GAS5 functions as an RNA scaffold binding to the WW domain of YAP to facilitate interactions between YAP and 14–3-3 proteins or LATS1, subsequently increasing the ubiquitin-mediated degradation of YAP. Importantly, the authors showed a novel target of YAP, YTHDF3. The lncRNA GAS5 decreased YAP-mediated transcription of YTHDF3, which reversibly bound to N6-methyladenosine (m6A)-methylated GAS5 to trigger its decay [149].
The crosstalk between circRNAs and YAP/TAZ
Regulation of YAP/TAZ by circRNAs
Circular RNAs (circRNAs) were first discovered in plant viroids and Sendai virus in 1976 [150]. Unlike linear RNAs, circRNAs have a covalently closed continuous loop structure without a 5′ cap or a 3′ poly(A) tail [151]. These structural characteristics increase the stability of circRNAs in plasma and other body fluids; thus, they are potential diagnostic and prognostic biomarkers in cancer. CircRNAs can modify various biological activities, for example, regulating transcription and degradation and occasionally acting as polypeptide-producing templates [152]. In addition, like lncRNAs, circRNAs can sponge miRNAs. Recently, circRNAs have been found to have the potential to either promote or suppress cancer development in a YAP/TAZ-dependent manner.
CircRNAs regulate YAP/TAZ by functioning as ceRNAs
In accordance with the mechanism by which lncRNAs function, circRNAs are becoming a novel research hotspot in the ceRNA family. For instance, circ_0001667 upregulation can promote the proliferation and migration of breast cancer cells by regulating the miR-125a-5p/TAZ signaling pathway [153]. CircRNAs can regulate cancer stemness. CircPIP5K1A is an oncogenic factor in osteosarcoma. Mechanistically, circPIP5K1A was found to enhance YAP expression by targeting miR-515-5p. Knockdown of circPIP5K1A inhibited the CSC properties of osteosarcoma cells, while miR-515-5p inhibitor treatment or YAP overexpression reversed this circPIP5K1A knockdown-induced inhibition [154]. In colorectal cancer, circ_0128846 was found to sponge miR-1184 to release the suppression of AJUBA expression, leading to reduced YAP phosphorylation and activated YAP signaling, thereby promoting proliferation, migration, and invasion and inhibiting apoptosis in colorectal cells [155].
Circ_104075 is a tumor-promoting factor that was found to be highly expressed in hepatocellular carcinoma cell lines and tissues and in the serum of hepatocellular carcinoma patients. The study reported that circ_104075 enhanced YAP expression by acting as a sponge for miR-582-3p, thereby promoting hepatocellular carcinoma progression. Interestingly, an N6-methyladenosine (m6A) motif was identified in the 353–357 base pair region of the YAP 3'UTR, and this m6A modification was essential for the interaction between miR-582-3p and the YAP 3'UTR, indicating that m6A modification of mRNA can impact its stability by regulating its interaction with miRNAs [156].
CircRNAs regulate YAP/TAZ by affecting protein–mRNA interactions
In the translation initiation complex, eukaryotic initiation factor (eIF) 4G binds to Poly(A)-binding protein (PABP), leading to mRNA circularization and enhancing translation. CircYAP, which is derived from the fourth and fifth exons of YAP, was found to negatively regulate YAP expression by suppressing the assembly of the YAP translation initiation machinery in breast cancer cells. CircYAP specifically recognized and bound to YAP mRNA and simultaneously bound to with eIF4G and PABP. These interactions competitively inhibited the interaction of eIF4G and PABP and therefore suppressed the initiation of YAP translation. This study demonstrated the downregulation of circYap expression in breast cancer tissue and indicated that it markedly suppressed the proliferation, migration and colony formation of breast cancer cells [157].
Poly(rC)-binding protein 2 (PCBP2) belongs to the polycytosine-binding protein family, and it plays a crucial role in regulating mRNA stability, protein translation, and protein–protein interactions [158]. Chen et al. reported that PCBP2 could destabilize YAP mRNAs in hepatocellular carcinoma. CircCPSF6, which was upregulated in hepatocellular carcinoma specimens, competitively interacted with PCBP2 to inhibit its binding to YAP1 mRNA, thus attenuating PCBP2-mediated destabilization of YAP1 [159]. Aberrant activation of the circCPSF6-YAP1 axis drove malignancy in hepatocellular carcinoma.
CircRNAs regulate YAP/TAZ by encoding small peptides
Intriguingly, circRNAs have been reported to encode small peptides to regulate tumor pathogenesis. CircPPP1R12A was found to activate YAP signaling by encoding a conserved 73-aa small peptide, circPPP1R12A-73aa, which reduced the phosphorylation and promoted the nuclear translocation of YAP by inhibiting the activation of MST1/2 and LATS1/2. CircPPP1R12A thus played an essential role in promoting the proliferation, migration, and invasion of colon cancer cells [160].
Table 3 circRNAs that regulate the expression of YAP/TAZ and cancer progression.
Regulation of circRNAs by YAP/TAZ
Most studies have focused on the role of circRNAs in regulating YAP/TAZ signaling; some studies have revealed YAP/TAZ as regulators of circRNAs. As elucidated by Verduci et al., circPVT1 was overexpressed in head and neck squamous tissues compared to matched normal tissues, and circPVT1 expression was increased in tumors carrying mutant p53 proteins. YAP and mut-p53 proteins are able to physically interact; mut-p53 and TEAD are recruited to the same site where YAP binds the circPVT1 promoter to promote circPVT1 transcription [169]. Mut-p53/YAP/TEAD complex-induced overexpression of circPVT1 enhanced the malignant phenotype in head and neck squamous cell carcinoma. Recently, another study reported that YAP can promote the expression of QKI; then, QKI, as a trans-acting RNA-binding factor involved in circRNA biogenesis, can facilitate circNOTCH1 generation. Further exploration demonstrated that circNOTCH1 can compete with NOTCH1 mRNA for METTL14 binding to increase NOTCH1 mRNA stability, eventually promoting tumor growth, in non-small-cell lung cancer [170].
Potential clinical applications of YAP/TAZ and ncRNAs
Although cancer is the second leading cause of death, there are still limited numbers of reliable diagnostic, prognostic, and predictive biomarkers. NcRNAs, which exist in different intercellular compartments and can also be identified in biological fluids, may be alternative biomarkers because of their characteristics of abundance and conservation. Moreover, it has been reported that some ncRNAs are cell- and tissue specific. As an example, the circ_104075 level was found to be higher in hepatocellular carcinoma patients than in healthy individuals and patients with hepatitis B, hepatitis C, cirrhosis, lung cancer, gastric cancer, colon cancer, or breast cancer, suggesting that elevated expression of circ_104075 is specific to hepatocellular carcinoma. The area under the receiver operating characteristic curve for circ_104075 was 0.973, with a sensitivity of 96.0% and a specificity of 98.3%, indicating that circ_104075 is a promising serum biomarker for the diagnosis of hepatocellular carcinoma [156]. As another example, circ1662, which directly binds to YAP1 to reduce its phosphorylation and promote its nuclear transfer, exhibited significantly higher expression in colorectal cancer tissues than in paired normal tissues. Expression of circ1662 was positively correlated with poor prognosis and tumor depth in colorectal cancer patients [171]. Similarly, the lncRNA SNHG3, a prognostic factor, was found to contribute to cervical cancer metastasis and initiation via association with YAP1 [172].
Exosomes are usually stable and can exist in body fluids, such as blood, saliva, urine, lacrimal fluid, and cerebrospinal fluid [173]. Chen’s team found that the miR-7 level was significantly higher in serum exosomes from healthy controls than in those from patients with lung carcinoma and that high miR-7 expression was associated with a strong response to gefitinib treatment, as well as longer survival times, in lung carcinoma patients [68]. Therefore, exosomal miR-7 may be a feasible blood-based biomarker for the EGFR T79M resistance mutation.
Previously, YAP/TAZ signaling activation was found to be able to lead to uncontrolled cancer cell proliferation, apoptosis evasion, EMT, and stemness. Notably, EMT and CSCs have been demonstrated to contribute to drug resistance, metastasis, and tumor recurrence. However, due to their unstructured nature, YAP/TAZ are difficult to target using small molecules. YAP/TAZ-associated ncRNAs can be targeted by small molecule inhibitors, antisense oligonucleotides, small interfering RNA/short hairpin RNA-mediated RNA interference techniques and masking of the binding motifs required for interactions between ncRNAs and YAP/TAZ [174]. Clustered regularly interspaced short palindromic repeat (CRISPR)/CRISPR-associated nuclease 9 (Cas9) gene editing can also be applied to manipulate ncRNA sequences [175]. Therefore, biological or pharmacological interventions based on ncRNAs related to YAP/TAZ may become a promising new strategy to reverse drug resistance and improve efficacy of chemotherapy in human cancers. For example, Zhan et al. reported that miR-455-3p inhibited cell proliferation and gemcitabine resistance in pancreatic cancer by targeting TAZ, while inhibition of miR-455-3p had the opposite effects [176]. A similar phenomenon was identified in glioma, in which miR-125b resensitized glioma cells to TNF-related apoptosis-inducing ligand (TRAIL) by suppressing TAZ expression [177]. MiR-509-3p and miR-509-3-5p were markedly downregulated in tissues of recurrent ovarian clear cell carcinoma compared with the paired primary cancer tissues. Furthermore, overexpression of miR-509-3p and miR-509-3-5p reversed cisplatin resistance in ovarian clear cell carcinoma by suppressing the expression of YAP1 [178]. These findings suggest that targeting ncRNAs associated with YAP expression can be considered a strategy to reduce chemoresistance and the relapse rate in cancers.
The lncRNA SNHG29 was found to promote the transcription of PD-L1 by activating YAP signaling, therefore facilitating tumor immune suppression. Moreover, Ni et al. reported that simvastatin suppressed lncRNA SNHG29-mediated YAP activation and promoted antitumor immunity by inhibiting PD-L1 expression [179]. This research uncovered simvastatin as a potential immunotherapeutic drug in colorectal cancer.
MSC-derived exosomes are promising natural nanovectors for drug and molecule delivery [180]. Zhang et al. revealed that exosomal circ_100395 released by AMSCs inhibited the malignant transformation of non-small-cell lung carcinoma cells through the miR-141-3p-LATS2 axis [181]. The transfer of circRNA_100395 via AMSC-derived exosomes is a novel therapeutic strategy for non-small-cell lung carcinoma.
Collectively, these findings indicate that the clinical applications of ncRNAs associated with YAP/TAZ signaling include their use as biomarkers for cancer diagnosis and therapeutic interventions in cancer.
Conclusion
Accumulating evidence indicates that YAP/TAZ and ncRNAs play essential roles in developing human cancers. As there are complex networks contributing to tumorigenesis, it is insufficient to select therapies targeting single genes. Therefore, improving our understanding of the multiple crosstalk mechanisms and potential feedback loops between ncRNAs and YAP/TAZ is particularly necessary. In this review, we described the interplay between YAP/TAZ and ncRNAs, together with their biological functions in cancers. Also, the crosstalks among miRNA, cirRNA and lncRNA can’t be ignored. We also evaluated YAP/TAZ-associated ncRNAs as biomarkers for cancer diagnosis and prognosis and discussed potential clinical therapeutic applications related to the regulatory mechanisms connecting YAP/TAZ and ncRNAs. However, validation experiments based on sufficient clinical samples for predicting prognosis and ensuring safety in the responsive patient population are still challenging for successful implementation of YAP/TAZ-related ncRNAs in the clinic. Overall, the study of the interplay between YAP/TAZ and ncRNAs may open novel research areas for clinical cancer diagnosis and therapy.
Availability of data and materials
Not applicable.
Abbreviations
- YAP:
-
Yes-associated protein
- TAZ:
-
Transcriptional co-activator with PDZ-binding motif
- EMT:
-
Epithelial-mesenchymal transitio: n
- ncRNAs:
-
Non-coding RNAs
- miRNAs:
-
MicroRNAs
- circRNAs:
-
Circular RNAs
- lncRNAs:
-
Long non-coding RNAs
- Mst1/2:
-
Ste20-like kinase1/2
- SAV1:
-
Salvador1
- LATS1/2:
-
Large tumour suppressor kinase 1/2
- CK1:
-
Casein kinase 1
- ACTL6A:
-
Actin-like 6A
- NLK:
-
Nemo-like kinase
- RASSF1A:
-
Ras association domain family protein1 isoform A
- NDR:
-
Nuclear Dbf2-related
- PD-L1:
-
Programmed death 1 ligand 1
- CSCs:
-
Cancer stem cells
- 3’-UTR:
-
3’-Untranslated region
- CAF:
-
Cancer associated-fibroblast
- NSCLC:
-
Non-small-cell lung cancer
- HIF1A:
-
Hypoxia-inducible factor 1 alpha
- VEGFA:
-
Vascular endothelial growth factor-A
- MYPT1:
-
Myosin phosphatase targeting proteins1
- WIPF1:
-
Wiskott–Aldrich syndrome protein interacting protein family member 1
- PTC:
-
Papillary thyroid cancer
- HUVEC:
-
Human umbilical vein endothelial cell
- MLL3:
-
Mixed-lineage leukemia3
- DACT2:
-
Dishevelled binding antagonist of beta catenin 2
- CCL2:
-
C–C motif chemokine ligand 2
- CREB:
-
CAMP response element-binding
- MAML1:
-
Mastermind-like 1
- ROS:
-
Reactive oxygen species
- EZH2:
-
Enhancer of zeste homolog 2
- LSD1:
-
Lysine‐specific demethylase 1
- RBMX:
-
RNA-binding motif protein encoded on the X chromosome
- HMGCR:
-
3-Hydroxy-3-methylglutaryl-coenzyme A (CoA) reductase
- T-Ics:
-
Tumour-initiating cells
- LLPS:
-
Liquid–liquid phase separation
- AMSC:
-
Adipose-derived mesenchymal stem cell
- PABP:
-
Poly(A)-binding protein
- PCBP2:
-
Poly(rC)-binding protein 2
- MSC:
-
Mesenchymal stem cell
- m6A:
-
N6-methyladenosine
- eIF:
-
eukaryotic initiation factor
- CRISPR:
-
Clustered regularly interspaced short palindromic repeat
- Cas9:
-
CRISPR-associated nuclease 9
- TRAIL:
-
TNF-related apoptosis-inducing ligand
References
Zhao B, Li L, Lei Q, Guan KL. The Hippo-YAP pathway in organ size control and tumorigenesis: an updated version. Genes Dev. 2010;24(9):862–74.
Hong W, Guan KL. The YAP and TAZ transcription co-activators: key downstream effectors of the mammalian Hippo pathway. Semin Cell Dev Biol. 2012;23(7):785–93.
Cunningham R, Hansen CG. The Hippo pathway in cancer: YAP/TAZ and TEAD as therapeutic targets in cancer. Clin Sci (Lond). 2022;136(3):197–222.
Qian M, Yan F, Wang W, Du J, Yuan T, Wu R. Deubiquitinase JOSD2 stabilizes YAP/TAZ to promote cholangiocarcinoma progression. Acta Pharm Sin B. 2021;11(12):4008–19.
Hayashi H, Uemura N, Zhao L, Matsumura K, Sato H, Shiraishi Y. Biological Significance of YAP/TAZ in Pancreatic Ductal Adenocarcinoma. Front Oncol. 2021;11:700315.
Strepkos D, Markouli M, Papavassiliou KA, Papavassiliou AG, Piperi C. Emerging roles for the YAP/TAZ transcriptional regulators in brain tumour pathology and targeting options. Neuropathol Appl Neurobiol. 2022;48(2):e12762.
Wu Q, Guo J, Liu Y, Zheng Q, Li X, et al. YAP drives fate conversion and chemoresistance of small cell lung cancer. Sci Adv. 2021;7(40):eabg1850.
Barrette AM, Ronk H, Joshi T, Mussa Z, Mehrotra M, Bouras A, et al. Anti-invasive efficacy and survival benefit of the YAP-TEAD inhibitor verteporfin in preclinical glioblastoma models. Neuro Oncol. 2022;24(5):694–707.
Zanconato F, Cordenonsi M, Piccolo S. YAP/TAZ at the Roots of Cancer. Cancer Cell. 2016;29(6):783–803.
Nguyen CDK, Yi C. YAP/TAZ Signaling and Resistance to Cancer Therapy. Trends Cancer. 2019;5(5):283–96.
Lo Sardo F, Strano S, Blandino G. YAP and TAZ in Lung Cancer: Oncogenic Role and Clinical Targeting. Cancers (Basel). 2018;10(5):137.
Anastasiadou E, Jacob LS, Slack FJ. Non-coding RNA networks in cancer. Nat Rev Cancer. 2018;18(1):5–18.
Oo JA, Brandes RP, Leisegang MS. Long non-coding RNAs: novel regulators of cellular physiology and function. Pflugers Arch. 2022;474(2):191–204.
Wang D, Ye R, Cai Z, Xue Y. Emerging roles of RNA-RNA interactions in transcriptional regulation. Wiley Interdiscip Rev RNA. 2022. https://doi.org/10.1002/wrna.1712.
Wooten S, Smith KN. Long non-coding RNA OIP5-AS1 (Cyrano): A context-specific regulator of normal and disease processes. Clin Transl Med. 2022;12(1):e706.
Wang J, Zhu S, Meng N, He Y, Lu R, Yan GR. ncRNA-Encoded Peptides or Proteins and Cancer. Mol Ther. 2019;27(10):1718–25.
Smolarz B, Durczyński A, Romanowicz H, Szyłło K, Hogendorf P. miRNAs in Cancer (Review of Literature). Int J Mol Sci. 2022;23(5):2805.
Ouyang J, Zhong Y, Zhang Y, Yang L, Wu P, Hou X, et al. Long non-coding RNAs are involved in alternative splicing and promote cancer progression. Br J Cancer. 2022;126(8):1113–24.
Wang H, Meng Q, Qian J, Li M, Gu C, Yang Y. Review: RNA-based diagnostic markers discovery and therapeutic targets development in cancer. Pharmacol Ther. 2022;234:108123.
Toden S, Goel A. Non-coding RNAs as liquid biopsy biomarkers in cancer. Br J Cancer. 2022;126(3):351–60.
Zhang M, Xin Y. Circular RNAs: a new frontier for cancer diagnosis and therapy. J Hematol Oncol. 2018;11(1):21.
Kulaberoglu Y, Lin K, Holder M, Gai Z, Gomez M, Assefa Shifa B, et al. Stable MOB1 interaction with Hippo/MST is not essential for development and tissue growth control. Nat Commun. 2017;8(1):695.
Chen Q, Zhang N, Xie R, Wang W, Cai J, Choi KS, et al. Homeostatic control of Hippo signaling activity revealed by an endogenous activating mutation in YAP. Genes Dev. 2015;29(12):1285–97.
Zhao B, Li L, Tumaneng K, Wang CY, Guan KL. A coordinated phosphorylation by Lats and CK1 regulates YAP stability through SCF(beta-TRCP). Genes Dev. 2010;24(1):72–85.
Heng BC, Zhang X, Aubel D, Bai Y, Li X, Wei Y, et al. An overview of signaling pathways regulating YAP/TAZ activity. Cell Mol Life Sci. 2021;78(2):497–512.
Dubois F, Bergot E, Zalcman G, Levallet G. RASSF1A, puppeteer of cellular homeostasis, fights tumorigenesis, and metastasis-an updated review. Cell Death Dis. 2019;10(12):928.
Hergovich A. The Roles of NDR Protein Kinases in Hippo Signalling. Genes (Basel). 2016;7(5):21.
Hong AW, Meng Z, Yuan HX, Plouffe SW, Moon S, Kim W, et al. Osmotic stress-induced phosphorylation by NLK at Ser128 activates YAP. EMBO Rep. 2017;18(1):72–86.
Moon S, Kim W, Kim S, Kim Y, Song Y, Bilousov O, et al. Phosphorylation by NLK inhibits YAP-14-3-3-interactions and induces its nuclear localization. EMBO Rep. 2017;18(1):61–71.
Ji J, Xu R, Zhang X, Han M, Xu Y, et al. Actin like-6A promotes glioma progression through stabilization of transcriptional regulators YAP/TAZ. Cell Death Dis. 2018;9(5):517.
Das Thakur M, Feng Y, Jagannathan R, Seppa MJ, Skeath JB, Longmore GD. Ajuba LIM proteins are negative regulators of the Hippo signaling pathway. Curr Biol. 2010;20(7):657–62.
Dupont S, Morsut L, Aragona M, Enzo E, Giulitti S, et al. Role of YAP/TAZ in mechanotransduction. Nature. 2011;474(7350):179–83.
Meng Z, Qiu Y, Lin KC, Kumar A, Placone JK, Fang C, et al. RAP2 mediates mechanoresponses of the Hippo pathway. Nature. 2018;560(7720):655–60.
Gao R, Kalathur RKR, Coto-Llerena M, Ercan C, Buechel D, Shuang S, et al. YAP/TAZ and ATF4 drive resistance to Sorafenib in hepatocellular carcinoma by preventing ferroptosis. EMBO Mol Med. 2021;13(12):e14351.
Cho K, Ro SW, Lee HW, Moon H, Han S, Kim HR, et al. YAP/TAZ Suppress Drug Penetration Into Hepatocellular Carcinoma Through Stromal Activation. Hepatology. 2021;74(5):2605–21.
Pobbati AV, Hong W. A combat with the YAP/TAZ-TEAD oncoproteins for cancer therapy. Theranostics. 2020;10(8):3622–35.
Imajo M, Miyatake K, Iimura A, Miyamoto A, Nishida E. A molecular mechanism that links Hippo signaling to the inhibition of Wnt/β-catenin signaling. EMBO J. 2012;31(5):1109–22.
Varelas X, Samavarchi-Tehrani P, Narimatsu M, Weiss A, Cockburn K, Larsen BG, et al. The Crumbs complex couples cell density sensing to Hippo-dependent control of the TGF-β-SMAD pathway. Dev Cell. 2010;19(6):831–44.
Wang Z, Wang F, Ding XY, Li TE, Wang HY, Gao YH, et al. Hippo/YAP signaling choreographs the tumor immune microenvironment to promote triple negative breast cancer progression via TAZ/IL-34 axis. Cancer Lett. 2021;527:174–90.
Kim MH, Kim CG, Kim SK, Shin SJ, Choe EA, Park SH, et al. YAP-Induced PD-L1 Expression Drives Immune Evasion in BRAFi-Resistant Melanoma. Cancer Immunol Res. 2018;6(3):255–66.
Huang T, Song X, Xu D, Tiek D, Goenka A, Wu B, Sastry N, Hu B, Cheng SY. Stem cell programs in cancer initiation, progression, and therapy resistance. Theranostics. 2020;10(19):8721–43.
Panciera T, Azzolin L, Fujimura A, Di Biagio D, Frasson C, Bresolin S, et al. Induction of Expandable Tissue-Specific Stem/Progenitor Cells through Transient Expression of YAP/TAZ. Cell Stem Cell. 2016;19(6):725–37.
Panciera T, Azzolin L, Cordenonsi M, Piccolo S. Mechanobiology of YAP and TAZ in physiology and disease. Nat Rev Mol Cell Biol. 2017;18(12):758–70.
LeBlanc L, Ramirez N, Kim J. Context-dependent roles of YAP/TAZ in stem cell fates and cancer. Cell Mol Life Sci. 2021;78(9):4201–19.
Cao X, Wang C, Liu J, Zhao B. Regulation and functions of the Hippo pathway in stemness and differentiation. Acta Biochim Biophys Sin (Shanghai). 2020;52(7):736–48.
Zhang J, Deng X. Effects of miR-599 targeting YAP1 on proliferation, invasion and apoptosis of bladder urothelial carcinoma cells. Exp Mol Pathol. 2021;118:104599.
Chen X, Cao R, Liu H, Zhang T, Yuan X, Xu S. MicroRNA-15a-5p targeting oncogene YAP1 inhibits cell viability and induces cell apoptosis in cervical cancer cells. Int J Mol Med. 2020;46(4):1301–10.
Huang X, Tang F, Weng Z, Zhou M, Zhang Q. MiR-591 functions as tumor suppressor in breast cancer by targeting TCF4 and inhibits Hippo-YAP/TAZ signaling pathway. Cancer Cell Int. 2019;19:108.
Hu XH, Dai J, Shang HL, Zhao ZX, Hao YD. miR-1285-3p is a potential prognostic marker in human osteosarcoma and functions as a tumor suppressor by targeting YAP1. Cancer Biomark. 2019;25(1):1–10.
Zhang MY, Lin J, Kui YC. MicroRNA-345 suppresses cell invasion and migration in non-small cell lung cancer by directly targeting YAP1. Eur Rev Med Pharmacol Sci. 2019;23(6):2436–43.
Zhang M, Zeng J, Zhao Z, Liu Z. Loss of MiR-424-3p, not miR-424-5p, confers chemoresistance through targeting YAP1 in non-small cell lung cancer. Mol Carcinog. 2017;56(3):821–32.
Zeng G, Xun W, Wei K, Yang Y, Shen H. MicroRNA-27a-3p regulates epithelial to mesenchymal transition via targeting YAP1 in oral squamous cell carcinoma cells. Oncol Rep. 2016;36(3):1475–82.
Zang S, Zhao S, Gao X, Li Y, Zhong C, Gao J. Restoration of miR-26b expression partially reverses the cisplatin resistance of NSCLC by targeting tafazzin. Onco Targets Ther. 2019;12:7551–60.
Liu S, Chu L, Xie M, Ma L, An H, Zhang W, et al. miR-92a-3p Promoted EMT via Targeting LATS1 in Cervical Cancer Stem Cells. Front Cell Dev Biol. 2021;9:757747.
Feng S, Sun H, Zhu W. MiR-92 overexpression suppresses immune cell function in ovarian cancer via LATS2/YAP1/PD-L1 pathway. Clin Transl Oncol. 2021;23(3):450–8.
Song L, Huang Y, Zhang X, Han S, Hou M, Li H. Downregulation of microRNA-224-3p Hampers Retinoblastoma Progression via Activation of the Hippo-YAP Signaling Pathway by Increasing LATS2. Invest Ophthalmol Vis Sci. 2020;61(3):32.
Gao Y, Yi J, Zhang K, Bai F, Feng B, Wang R, et al. Downregulation of MiR-31 stimulates expression of LATS2 via the hippo pathway and promotes epithelial-mesenchymal transition in esophageal squamous cell carcinoma. J Exp Clin Cancer Res. 2017;36(1):161.
Zhang H, Lang TY, Zou DL, Zhou L, Lou M, Liu JS, et al. miR-520b Promotes Breast Cancer Stemness Through Hippo/YAP Signaling Pathway. Onco Targets Ther. 2019;12:11691–700.
Zhao J, Li L, Yang T. MiR-216a-3p suppresses the proliferation and invasion of cervical cancer through downregulation of ACTL6A-mediated YAP signaling. J Cell Physiol. 2020;235(12):9718–28.
Cao MX, Zhang WL, Yu XH, Wu JS, Qiao XW, Huang MC, et al. Interplay between cancer cells and M2 macrophages is necessary for miR-550a-3-5p down-regulation-mediated HPV-positive OSCC progression. J Exp Clin Cancer Res. 2020;39(1):102.
Choe MH, Yoon Y, Kim J, Hwang SG, Han YH, Kim JS. miR-550a-3-5p acts as a tumor suppressor and reverses BRAF inhibitor resistance through the direct targeting of YAP. Cell Death Dis. 2018;9(6):640.
Zhao L, Han S, Hou J, Shi W, Zhao Y, Chen Y. The local anesthetic ropivacaine suppresses progression of breast cancer by regulating miR-27b-3p/YAP axis. Aging (AlbanyNY). 2021;13(12):16341–52.
Du YE, Tu G, Yang G, Li G, Yang D, Lang L, et al. MiR-205/YAP1 in Activated Fibroblasts of Breast Tumor Promotes VEGF-independent Angiogenesis through STAT3 Signaling. Theranostics. 2017;7(16):3972–88.
Higashi T, Hayashi H, Ishimoto T, Takeyama H, Kaida T, Arima K, et al. miR-9-3p plays a tumour-suppressor role by targeting TAZ (WWTR1) in hepatocellular carcinoma cells. Br J Cancer. 2015;113(2):252–8.
Chen M, Wu L, Tu J, Zhao Z, Fan X, Mao J, et al. MiR-590-5p suppresses hepatocellular carcinoma chemoresistance by targeting YAP1 expression. EBioMedicine. 2018;35:142–54.
Yu K, Li H, Jiang Z, Hsu HJ, Hsu HC, Zhang Y, et al. miR 375/Yes associated protein axis regulates IL 6 and TGF β expression, which is involved in the cisplatin induced resistance of liver cancer cells. Oncol Rep. 2021;46(2):162.
Jin D, Guo J, Wu Y, Chen W, Du J, Yang L, et al. Metformin-repressed miR-381-YAP-snail axis activity disrupts NSCLC growth and metastasis. J Exp Clin Cancer Res. 2020;39(1):6.
Chen R, Qian Z, Xu X, Zhang C, Niu Y, Wang Z, et al. Exosomes-transmitted miR-7 reverses gefitinib resistance by targeting YAP in non-small-cell lung cancer. Pharmacol Res. 2021;165:105442.
Sun M, Song H, Wang S, Zhang C, Zheng L, Chen F, et al. Integrated analysis identifies microRNA-195 as a suppressor of Hippo-YAP pathway in colorectal cancer. J Hematol Oncol. 2017;10(1):79.
Xu X, Chen X, Xu M, Liu X, Pan B, Qin J, et al. miR-375-3p suppresses tumorigenesis and partially reverses chemoresistance by targeting YAP1 and SP1 in colorectal cancer cells. Aging (Albany NY). 2019;11(18):7357–85.
Selth LA, Das R, Townley SL, Coutinho I, Hanson AR, Centenera MM, et al. A ZEB1-miR-375-YAP1 pathway regulates epithelial plasticity in prostate cancer. Oncogene. 2017;36(1):24–34.
Kang W, Tong JH, Lung RW, Dong Y, Zhao J, Liang Q, Zhang L, Pan Y, Yang W, Pang JC, Cheng AS, Yu J, To KF. Targeting of YAP1 by microRNA-15a and microRNA-16-1 exerts tumor suppressor function in gastric adenocarcinoma. Mol Cancer. 2015;14:52.
Chen X, Wang AL, Liu YY, Zhao CX, Zhou X, Liu HL, et al. MiR-429 Involves in the Pathogenesis of Colorectal Cancer via Directly Targeting LATS2. Oxid Med Cell Longev. 2020. https://doi.org/10.1155/2020/5316276.
Sun Z, Zhang Q, Yuan W, Li X, Chen C, Guo Y, et al. MiR-103a-3p promotes tumour glycolysis in colorectal cancer via hippo/YAP1/HIF1A axis. J Exp Clin Cancer Res. 2020;39(1):250.
Zheng D, Cao M, Zuo S, Xia X, Zhi C, Lin Y, et al. RANBP1 promotes colorectal cancer progression by regulating pre-miRNA nuclear export via a positive feedback loop with YAP. Oncogene. 2022;41(7):930–42.
Wu T, Hu H, Zhang T, Jiang L, Li X, Liu S, Zheng C, Yan G, Chen W, Ning Y, Li Y, Lu Z. miR-25 Promotes Cell Proliferation, Migration, and Invasion of Non-Small-Cell Lung Cancer by Targeting the LATS2/YAP Signaling Pathway. Oxid Med Cell Longev. 2019. https://doi.org/10.1155/2019/9719723.
Jin D, Guo J, Wu Y, Yang L, Wang X, Du J, et al. m6A demethylase ALKBH5 inhibits tumor growth and metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2-mediated YAP activity in NSCLC. Mol Cancer. 2020;19(1):40.
Lin CW, Chang YL, Chang YC, Lin JC, Chen CC, Pan SH, et al. MicroRNA-135b promotes lung cancer metastasis by regulating multiple targets in the Hippo pathway and LZTS1. Nat Commun. 2013;4:1877.
Guo Y, Cui J, Ji Z, Cheng C, Zhang K, Zhang C, et al. MiR-302/367/LATS2/YAP pathway is essential for prostate tumor-propagating cells and promotes the development of castration resistance. Oncogene. 2017;36(45):6336–47.
Muñoz-Galván S, Felipe-Abrio B, Verdugo-Sivianes EM, Perez M, Jiménez-García MP, et al. Downregulation of MYPT1 increases tumor resistance in ovarian cancer by targeting the Hippo pathway and increasing the stemness. Mol Cancer. 2020;19(1):7.
Pan Y, Lu F, Xiong P, Pan M, Zhang Z, Lin X, et al. WIPF1 antagonizes the tumor suppressive effect of miR-141/200c and is associated with poor survival in patients with PDAC. J Exp Clin Cancer Res. 2018;37(1):167.
Gargini R, Escoll M, García E, García-Escudero R, Wandosell F, Antón IM. WIP Drives Tumor Progression through YAP/TAZ-Dependent Autonomous Cell Growth. Cell Rep. 2016;17(8):1962–77.
Zhou H, He X, He Y, Ou C, Cao P. Exosomal circRNAs: Emerging Players in Tumor Metastasis. Front Cell Dev Biol. 2021;9:786224.
Wang Y, Cen A, Yang Y, Ye H, Li J, Liu S, et al. miR-181a, delivered by hypoxic PTC-secreted exosomes, inhibits DACT2 by downregulating MLL3, leading to YAP-VEGF-mediated angiogenesis. Mol Ther Nucleic Acids. 2021;24:610–21.
Tan Y, Li QM, Huang N, Cheng S, Zhao GJ, Chen H, et al. Upregulation of DACT2 suppresses proliferation and enhances apoptosis of glioma cell via inactivation of YAP signaling pathway. Cell Death Dis. 2017;8(8):e2981.
Kim J, Kwon H, Shin YK, Song G, Lee T, Kim Y, et al. MAML1/2 promote YAP/TAZ nuclear localization and tumorigenesis. Proc Natl Acad Sci U S A. 2020;117(24):13529–40.
Wang J, Ma L, Weng W, Qiao Y, Zhang Y, He J, et al. Mutual interaction between YAP and CREB promotes tumorigenesis in liver cancer. Hepatology. 2013;58(3):1011–20.
Yang S, Jiang W, Yang W, Yang C, Yang X, Chen K, et al. Epigenetically modulated miR-1224 suppresses the proliferation of HCC through CREB-mediated activation of YAP signaling pathway. Mol Ther Nucleic Acids. 2021;23:944–58.
Yang Y, Li H, Liu Y, Chi C, Ni J, Lin X. MiR-4319 hinders YAP expression to restrain non-small cell lung cancer growth through regulation of LIN28-mediated RFX5 stability. Biomed Pharmacother. 2019;115:108956.
Ma J, Huang K, Ma Y, Zhou M, Fan S. The TAZ-miR-224-SMAD4 axis promotes tumorigenesis in osteosarcoma. Cell Death Dis. 2017;8(1):e2539.
Yu FX, Guan KL. Transcription and processing: multilayer controls of RNA biogenesis by the Hippo pathway. EMBO J. 2014;33(9):942–4.
Mori M, Triboulet R, Mohseni M, Schlegelmilch K, Shrestha K, Camargo FD, et al. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer. Cell. 2014;156(5):893–906.
Chaulk SG, Lattanzi VJ, Hiemer SE, Fahlman RP, Varelas X. The Hippo pathway effectors TAZ/YAP regulate dicer expression and microRNA biogenesis through Let-7. J Biol Chem. 2014;289(4):1886–91.
Zhang HT, Gui T, Liu RX, Tong KL, Wu CJ, Li Z, et al. Sequential targeting of YAP1 and p21 enhances the elimination of senescent cells induced by the BET inhibitor JQ1. Cell Death Dis. 2021;12(1):121.
Marco A, Ninova M, Griffiths-Jones S. Multiple products from microRNA transcripts. Biochem Soc Trans. 2013;41(4):850–4.
Lo Sardo F, Forcato M, Sacconi A, Capaci V, Zanconato F, Di Agostino S, et al. MCM7 and its hosted miR-25, 93 and 106b cluster elicit YAP/TAZ oncogenic activity in lung cancer. Carcinogenesis. 2017;38(1):64–75.
Shen S, Huang K, Wu Y, Ma Y, Wang J, Qin F, et al. A miR-135b-TAZ positive feedback loop promotes epithelial-mesenchymal transition (EMT) and tumorigenesis in osteosarcoma. Cancer Lett. 2017;407:32–44.
Wang F, Fan M, Zhou X, Yu Y, Cai Y, Wu H, et al. A positive feedback loop between TAZ and miR-942-3p modulates proliferation, angiogenesis, epithelial-mesenchymal transition process, glycometabolism and ROS homeostasis in human bladder cancer. J Exp Clin Cancer Res. 2021;40(1):44.
Chen J, Zhang K, Zhi Y, Wu Y, Chen B, Bai J, et al. Tumor-derived exosomal miR-19b-3p facilitates M2 macrophage polarization and exosomal LINC00273 secretion to promote lung adenocarcinoma metastasis via Hippo pathway. Clin Transl Med. 2021;11(9):e478.
Shen S, Guo X, Yan H, Lu Y, Ji X, Li L, et al. A miR-130a-YAP positive feedback loop promotes organ size and tumorigenesis. Cell Res. 2015;25(9):997–1012.
Jiang N, Zhao L, Zong D, Yin L, Wu L, Chen C, et al. Long non-coding RNA LUADT1 promotes nasopharyngeal carcinoma cell proliferation and invasion by downregulating miR-1207-5p. Bioengineered. 2021;12(2):10716–28.
Zhu X, Zhu J, Tan T, Bu F, Zhao J, Luo C, et al. RP11–51O6.1 sponges miR-206 to accelerate colorectal cancer carcinogenesis and metastasis through upregulating YAP1. Carcinogenesis. 2021;42(7):984–94.
Zhu X, Bu F, Tan T, Luo Q, Zhu J, Lin K, et al. Long noncoding RNA RP11–757G1.5 sponges miR-139–5p and upregulates YAP1 thereby promoting the proliferation and liver, spleen metastasis of colorectal cancer. J Exp Clin Cancer Res. 2020;39(1):207.
Shen Y, Gao X, Tan W, Xu T. STAT1-mediated upregulation of lncRNA LINC00174 functions a ceRNA for miR-1910-3p to facilitate colorectal carcinoma progression through regulation of TAZ. Gene. 2018;666:64–71.
Wang H, Di X, Bi Y, Sun S, Wang T. Long non-coding RNA LINC00649 regulates YES-associated protein 1 (YAP1)/Hippo pathway to accelerate gastric cancer (GC) progression via sequestering miR-16-5p. Bioengineered. 2021;12(1):1791–802.
Liu Y, Lin W, Dong Y, Li X, Lin Z, Jia J, et al. Long noncoding RNA HCG18 up-regulates the expression of WIPF1 and YAP/TAZ by inhibiting miR-141-3p in gastric cancer. Cancer Med. 2020;9(18):6752–65.
Yu M, Yi B, Zhou W, Gong W, Li G, Yu S. Linc00475 promotes the progression of glioma by regulating the miR-141-3p/YAP1 axis. J Cell Mol Med. 2021;25(1):463–72.
Wang X, Li XD, Fu Z, Zhou Y, Huang X, Jiang X. Long non coding RNA LINC00473/miR-195-5p promotes glioma progression via YAP1 TEAD1 Hippo signaling. Int J Oncol. 2020;56(2):508–21.
Yu W, Xiang D, Jia H, He X, Sheng J, Long Y, Zhu S, Wang K, Liu Q. The lncRNA BCYRN1 Functions as an Oncogene in Human Glioma by Downregulating miR-125a-5p in vitro. Cancer Manag Res. 2020;12:1151–61.
Ding P, Liang B, Shou J, Wang X. lncRNA KCNQ1OT1 promotes proliferation and invasion of glioma cells by targeting the miR 375/YAP pathway. Int J Mol Med. 2020;46(6):1983–92.
Li C, Ye J, Zhang Z, Gong Z, Lin Z, Ding M. Long non-coding RNA RBM5-AS1 promotes the aggressive behaviors of oral squamous cell carcinoma by regulation of miR-1285-3p/YAP1 axis. Biomed Pharmacother. 2020;123:109723.
Zhang C, Bao C, Zhang X, Lin X, Pan D, Chen Y. Knockdown of lncRNA LEF1-AS1 inhibited the progression of oral squamous cell carcinoma (OSCC) via Hippo signaling pathway. Cancer Biol Ther. 2019;20(9):1213–22.
Lu X, Gan Q, Gan C, Zheng Y, Cai B, Li X, et al. Long non-coding RNA PICSAR knockdown inhibits the progression of cutaneous squamous cell carcinoma by regulating miR-125b/YAP1 axis. Life Sci. 2021;274:118303.
Meng Q, Li Z, Pan J, Sun X. Long noncoding RNA DUXAP8 regulates proliferation and apoptosis of ovarian cancer cells via targeting miR-590-5p. Hum Cell. 2020;33(4):1240–51.
Lin X, Feng D, Li P, Lv Y. LncRNA LINC00857 regulates the progression and glycolysis in ovarian cancer by modulating the Hippo signaling pathway. Cancer Med. 2020;9(21):8122–32.
Wang K, Hu YB, Zhao Y, Ye C. Long non-coding RNA ASAP1-IT1 suppresses ovarian cancer progression by regulating Hippo/YAP signaling. Int J Mol Med. 2021;47(4):44.
Wang Q, Ding J, Nan G, Lyu Y, Ni G. LncRNA NOC2L-4.1 functions as a tumor oncogene in cervical cancer progression by regulating the miR-630/YAP1 pathway. J Cell Biochem. 2019;120(10):16913–20.
Guan H, Shang G, Cui Y, Liu J, Sun X, Cao W, et al. Long noncoding RNA APTR contributes to osteosarcoma progression through repression of miR-132-3p and upregulation of yes-associated protein 1. J Cell Physiol. 2019;234(6):8998–9007.
Liu S, Yang Y, Wang W, Pan X. Long noncoding RNA TUG1 promotes cell proliferation and migration of renal cell carcinoma via regulation of YAP. J Cell Biochem. 2018;119(12):9694–706.
Zang Y, Li J, Wan B, Tai Y, Liu H, Li Q, Ji Y. Long non-coding RNA CCAT2 drives the growth of laryngeal squamous cell carcinoma via regulating YAP activity. Hum Cell. 2021;34(6):1878–87.
Huang S, Li Y, Hu J, Li L, Liu Z, Guo H, et al. LncRNA PWAR6 regulates proliferation and migration by epigenetically silencing YAP1 in tumorigenesis of pancreatic ductal adenocarcinoma. J Cell Mol Med. 2021;25(9):4275–86.
Yan H, Li H, Silva MA, Guan Y, Yang L, Zhu L, et al. LncRNA FLVCR1-AS1 mediates miR-513/YAP1 signaling to promote cell progression, migration, invasion and EMT process in ovarian cancer. J Exp Clin Cancer Res. 2019;38(1):356.
Yan H, Li H, Li P, Li X, Lin J, Zhu L, et al. Long noncoding RNA MLK7-AS1 promotes ovarian cancer cells progression by modulating miR-375/YAP1 axis. J Exp Clin Cancer Res. 2018;37(1):237.
Di Agostino S, Valenti F, Sacconi A, Fontemaggi G, Pallocca M, Pulito C, Ganci F, Muti P, Strano S, Blandino G. Long Non-coding MIR205HG Depletes Hsa-miR-590-3p Leading to Unrestrained Proliferation in Head and Neck Squamous Cell Carcinoma. Theranostics. 2018;8(7):1850–68.
Wu DM, Wang S, Wen X, Han XR, Wang YJ, Shen M, et al. LncRNA SNHG15 acts as a ceRNA to regulate YAP1-Hippo signaling pathway by sponging miR-200a-3p in papillary thyroid carcinoma. Cell Death Dis. 2018;9(10):947.
Ma D, Gao X, Liu Z, Lu X, Ju H, Zhang N. Exosome-transferred long non-coding RNA ASMTL-AS1 contributes to malignant phenotypes in residual hepatocellular carcinoma after insufficient radiofrequency ablation. Cell Prolif. 2020;53(9):e12795.
Zhu Y, He D, Bo H, Liu Z, Xiao M, Xiang L, et al. The MRVI1-AS1/ATF3 signaling loop sensitizes nasopharyngeal cancer cells to paclitaxel by regulating the Hippo-TAZ pathway. Oncogene. 2019;38(32):6065–81.
Wang J, Huang F, Shi Y, Zhang Q, Xu S, Yao Y, et al. RP11–323N12.5 promotes the malignancy and immunosuppression of human gastric cancer by increasing YAP1 transcription. Gastric Cancer. 2021;24(1):85–102.
Chen L, Chen Q, Kuang S, Zhao C, Yang L, Zhang Y, et al. USF1-induced upregulation of LINC01048 promotes cell proliferation and apoptosis in cutaneous squamous cell carcinoma by binding to TAF15 to transcriptionally activate YAP1. Cell Death Dis. 2019;10(4):296.
Li F, Chen Q, Xue H, Zhang L, Wang K, Shen F. LncRNA MNX1-AS1 promotes progression of intrahepatic cholangiocarcinoma through the MNX1/Hippo axis. Cell Death Dis. 2020;11(10):894.
Wu M, Tong CWS, Yan W, To KKW, Cho WCS. The RNA Binding Protein HuR: A Promising Drug Target for Anticancer Therapy. Curr Cancer Drug Targets. 2019;19(5):382–99.
Li Z, Wang Y, Hu R, Xu R, Xu W. LncRNA B4GALT1-AS1 recruits HuR to promote osteosarcoma cells stemness and migration via enhancing YAP transcriptional activity. Cell Prolif. 2018;51(6):e12504.
Michalak EM, Burr ML, Bannister AJ, Dawson MA. The roles of DNA, RNA and histone methylation in ageing and cancer. Nat Rev Mol Cell Biol. 2019;20(10):573–89.
Bosselut R. Pleiotropic Functions of H3K27Me3 Demethylases in Immune Cell Differentiation. Trends Immunol. 2016;37(2):102–13.
Zhang L, Wang J, Pan Y, Jin J, Sang J, Huang P, et al. Expression of histone H3 lysine 4 methylation and its demethylases in the developing mouse testis. Cell Tissue Res. 2014;358(3):875–83.
Liu K, Ni JD, Li WZ, Pan BQ, Yang YT, Xia Q, et al. The Sp1/FOXC1/HOTTIP/LATS2/YAP/β-catenin cascade promotes malignant and metastatic progression of osteosarcoma. Mol Oncol. 2020;14(10):2678–95.
Mehta S, Zhang J. Liquid-liquid phase separation drives cellular function and dysfunction in cancer. Nat Rev Cancer. 2022;22(4):239–52.
Adnane S, Marino A, Leucci E. LncRNAs in human cancers: signal from noise. Trends Cell Biol. 2022;S0962–8924(22):00007–11.
Li RH, Tian T, Ge QW, He XY, Shi CY, Li JH, et al. A phosphatidic acid-binding lncRNA SNHG9 facilitates LATS1 liquid-liquid phase separation to promote oncogenic YAP signaling. Cell Res. 2021. https://doi.org/10.1038/s41422-021-00530-9.
Chan LH, Wang W, Yeung W, Deng Y, Yuan P, Mak KK. Hedgehog signaling induces osteosarcoma development through Yap1 and H19 overexpression. Oncogene. 2014;33(40):4857–66.
Xing Z, Lin A, Li C, Liang K, Wang S, Liu Y, et al. lncRNA directs cooperative epigenetic regulation downstream of chemokine signals. Cell. 2014;159(5):1110–25.
Zheng X, Han H, Liu GP, Ma YX, Pan RL, Sang LJ, et al. LncRNA wires up Hippo and Hedgehog signaling to reprogramme glucose metabolism. EMBO J. 2017;36(22):3325–35.
Sun Z, Ou C, Liu J, Chen C, Zhou Q, Yang S, et al. YAP1-induced MALAT1 promotes epithelial-mesenchymal transition and angiogenesis by sponging miR-126-5p in colorectal cancer. Oncogene. 2019;38(14):2627–44.
Kim M, Kim T, Johnson RL, Lim DS. Transcriptional co-repressor function of the hippo pathway transducers YAP and TAZ. Cell Rep. 2015;11(2):270–82.
Tan BS, Yang MC, Singh S, Chou YC, Chen HY, Wang MY, et al. LncRNA NORAD is repressed by the YAP pathway and suppresses lung and breast cancer metastasis by sequestering S100P. Oncogene. 2019;38(28):5612–26.
Qu L, Wu Z, Li Y, Xu Z, Liu B, Liu F, et al. A feed-forward loop between lncARSR and YAP activity promotes expansion of renal tumour-initiating cells. Nat Commun. 2016;7:12692.
Li N, Yang G, Luo L, Ling L, Wang X, Shi L, et al. lncRNA THAP9-AS1 Promotes Pancreatic Ductal Adenocarcinoma Growth and Leads to a Poor Clinical Outcome via Sponging miR-484 and Interacting with YAP. Clin Cancer Res. 2020;26(7):1736–48.
Zhu B, Finch-Edmondson M, Leong KW, Zhang X, V M, Lin QXX, Lee Y, Ng WT, Guo H, Wan Y, Sudol M, DasGupta R. LncRNA SFTA1P mediates positive feedback regulation of the Hippo-YAP/TAZ signaling pathway in non-small cell lung cancer. Cell Death Discov. 2021;7(1):369.
Ni W, Yao S, Zhou Y, Liu Y, Huang P, Zhou A, et al. Long noncoding RNA GAS5 inhibits progression of colorectal cancer by interacting with and triggering YAP phosphorylation and degradation and is negatively regulated by the m6A reader YTHDF3. MolCancer. 2019;18(1):143.
Sanger HL, Klotz G, Riesner D, Gross HJ, Kleinschmidt AK. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc Natl Acad Sci U S A. 1976;73(11):3852–6.
Allegra A, Cicero N, Tonacci A, Musolino C, Gangemi S. Circular RNA as a Novel Biomarker for Diagnosis and Prognosis and Potential Therapeutic Targets in Multiple Myeloma. Cancers (Basel). 2022;14(7):1700.
Sharma AR, Banerjee S, Bhattacharya M, Saha A, Lee SS, Chakraborty C. Recent progress of circular RNAs in different types of human cancer: Technological landscape, clinical opportunities and challenges (Review). Int J Oncol. 2022;60(5):56.
Geng Z, Wang W, Chen H, Mao J, Li Z, Zhou J. Circ_0001667 promotes breast cancer cell proliferation and survival via Hippo signal pathway by regulating TAZ. Cell Biosci. 2019;9:104.
Shi P, Li Y, Guo Q. Circular RNA circPIP5K1A contributes to cancer stemness of osteosarcoma by miR-515-5p/YAP axis. J Transl Med. 2021;19(1):464.
Wang X, Chen Y, Liu W, Liu T, Sun D. Hsa_circ_0128846 promotes tumorigenesis of colorectal cancer by sponging hsa-miR-1184 and releasing AJUBA and inactivating Hippo/YAP signalling. J Cell Mol Med. 2020;24(17):9908–24.
Zhang X, Xu Y, Qian Z, Zheng W, Wu Q, Chen Y, et al. circRNA_104075 stimulates YAP-dependent tumorigenesis through the regulation of HNF4a and may serve as a diagnostic marker in hepatocellular carcinoma. Cell Death Dis. 2018;9(11):1091.
Wu N, Yuan Z, Du KY, Fang L, Lyu J, Zhang C, et al. Translation of yes-associated protein (YAP) was antagonized by its circular RNA via suppressing the assembly of the translation initiation machinery. Cell Death Differ. 2019;26(12):2758–73.
Gu H, Yang J, Zhang J, Song Y, Zhang Y, Xu P, et al. PCBP2 maintains antiviral signaling homeostasis by regulating cGAS enzymatic activity via antagonizing its condensation. Nat Commun. 2022;13(1):1564.
Chen Y, Ling Z, Cai X, Xu Y, Lv Z, Man D, et al. Activation of YAP1 by N6-Methyladenosine-Modified circCPSF6 Drives Malignancy in Hepatocellular Carcinoma. Cancer Res. 2022;82(4):599–614.
Zheng X, Chen L, Zhou Y, Wang Q, Zheng Z, Xu B, et al. A novel protein encoded by a circular RNA circPPP1R12A promotes tumor pathogenesis and metastasis of colon cancer via Hippo-YAP signaling. Mol Cancer. 2019;18(1):47.
Wu D, Jia H, Zhang Z, Li S. Circ_0000511 accelerates the proliferation, migration and invasion, and restrains the apoptosis of breast cancer cells through the miR 326/TAZ axis. Int J Oncol. 2021;58(4):1.
Hu J, Ji C, Hua K, Wang X, Deng X, Li J, et al. Hsa_circ_0091074 regulates TAZ expression via microRNA 1297 in triple-negative breast cancer cells. Int J Oncol. 2020;56(5):1314–26.
Liu G, Huang K, Jie Z, Wu Y, Chen J, Chen Z, et al. CircFAT1 sponges miR-375 to promote the expression of Yes-associated protein 1 in osteosarcoma cells. Mol Cancer. 2018;17(1):170.
Wang X, Ji C, Hu J, Deng X, Zheng W, Yu Y, et al. Hsa_circ_0005273 facilitates breast cancer tumorigenesis by regulating YAP1-hippo signaling pathway. J Exp Clin Cancer Res. 2021;40(1):29.
Niu R, Li D, Chen J, Zhao W. Circ_0014235 confers Gefitinib resistance and malignant behaviors in non-small cell lung cancer resistant to Gefitinib by governing the miR-146b-5p/YAP/PD-L1 pathway. Cell Cycle. 2021;17:1–15.
Liu G, Zhou J, Piao Y, Zhao X, Zuo Y, Ji Z. Hsa_circ_0085576 promotes clear cell renal cell carcinoma tumorigenesis and metastasis through the miR-498/YAP1 axis. Aging (Albany NY). 2020;12(12):11530–49.
Li S, Yan G, Liu W, Li C, Wang X. Circ0106714 inhibits tumorigenesis of colorectal cancer by sponging miR-942-5p and releasing DLG2 via Hippo-YAP signaling. Mol Carcinog. 2020;59(12):1323–42.
Peng QS, Cheng YN, Zhang WB, Fan H, Mao QH, Xu P. circRNA_0000140 suppresses oral squamous cell carcinoma growth and metastasis by targeting miR-31 to inhibit Hippo signaling pathway. Cell Death Dis. 2020;11(2):112.
Verduci L, Ferraiuolo M, Sacconi A, Ganci F, Vitale J, Colombo T, et al. The oncogenic role of circPVT1 in head and neck squamous cell carcinoma is mediated through the mutant p53/YAP/TEAD transcription-competent complex. Genome Biol. 2017;18(1):237.
Shen Y, Li C, Zhou L, Huang JA. G protein-coupled oestrogen receptor promotes cell growth of non-small cell lung cancer cells via YAP1/QKI/circNOTCH1/m6A methylated NOTCH1 signalling. J Cell Mol Med. 2021;25(1):284–96.
Chen C, Yuan W, Zhou Q, Shao B, Guo Y, Wang W, et al. N6-methyladenosine-induced circ1662 promotes metastasis of colorectal cancer by accelerating YAP1 nuclear localization. Theranostics. 2021;11(9):4298–315.
Zhu H, Zhu C, Feng X, Luo Y. Long noncoding RNA SNHG3 promotes malignant phenotypes in cervical cancer cells via association with YAP1. Hum Cell. 2022;35(1):320–32.
Salehi M, Sharifi M. Exosomal miRNAs as novel cancer biomarkers: Challenges and opportunities. J Cell Physiol. 2018;233(9):6370–80.
Winkle M, El-Daly SM, Fabbri M, Calin GA. Noncoding RNA therapeutics - challenges and potential solutions. Nat Rev Drug Discov. 2021;20(8):629–51.
Yang J, Meng X, Pan J, Jiang N, Zhou C, Wu Z, et al. CRISPR/Cas9-mediated noncoding RNA editing in human cancers. RNA Biol. 2018;15(1):35–43.
Zhan T, Huang X, Tian X, Chen X, Ding Y, Luo H, et al. Downregulation of MicroRNA-455-3p Links to Proliferation and Drug Resistance of Pancreatic Cancer Cells via Targeting TAZ. Mol Ther Nucleic Acids. 2018;10:215–26.
Ma W, Cui Y, Liu M, Tan Z, Jiang Y. Downregulation of miR-125b promotes resistance of glioma cells to TRAIL through overexpression of Tafazzin which is a mitochondrial protein. Aging (Albany NY). 2019;11(9):2670–80.
Yoshida K, Yokoi A, Sugiyama M, Oda S, Kitami K, Tamauchi S, et al. Expression of the chrXq27.3 miRNA cluster in recurrent ovarian clear cell carcinoma and its impact on cisplatin resistance. Oncogene. 2021;40(7):1255–68.
Ni W, Mo H, Liu Y, Xu Y, Qin C, Zhou Y, et al. Targeting cholesterol biosynthesis promotes anti-tumor immunity by inhibiting long noncoding RNA SNHG29-mediated YAP activation. Mol Ther. 2021;29(10):2995–3010.
Hassanzadeh A, Rahman HS, Markov A, Endjun JJ, Zekiy AO, Chartrand MS, et al. Mesenchymal stem/stromal cell-derived exosomes in regenerative medicine and cancer; overview of development, challenges, and opportunities. Stem Cell Res Ther. 2021;12(1):297.
Zhang C, Cao J, Lv W, Mou H. CircRNA_100395 Carried by Exosomes From Adipose-Derived Mesenchymal Stem Cells Inhibits the Malignant Transformation of Non-Small Cell Lung Carcinoma Through the miR-141-3p-LATS2 Axis. Front Cell Dev Biol. 2021;9:663147.
Acknowledgements
Not applicable.
Funding
This work was supported by the General Program of National Natural Science Foundation of China(No.81972278), Nanjing Medical University Youth Fund (No. NMUB2019041), and Outstanding Talent Program of SAHNMU (789ZYRC202090147).
Author information
Authors and Affiliations
Contributions
Keming Wang and Jie Ding provided conception and design of this manuscript.Yirao Zhang,Yang Wang,Hao Ji participated in writing the manuscript. Yirao Zhang prepared the tables and figures. Keming Wang and Jie Ding critically reviewed and edited the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, Y., Wang, Y., Ji, H. et al. The interplay between noncoding RNA and YAP/TAZ signaling in cancers: molecular functions and mechanisms. J Exp Clin Cancer Res 41, 202 (2022). https://doi.org/10.1186/s13046-022-02403-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-022-02403-4