
Abstract
Background
Escherichia coli is an important cause of diarrhea in calves and its diarrheagenic properties are related to presence of certain virulence genes. In this study, the prevalence of virulence genes F5, F17, F41, sta, stx1, stx2, eae, and saa in E. coli isolated from pre-weaned calves presenting with (n= 329) or without diarrhea (n= 360) was explored using multiplex polymerase chain reaction. We also evaluated the association between detection of E. coli and the presence of diarrhea.
Results
Escherichia coli was detected in 56.3% (388/689) of the fecal samples and showed the highest prevalence (66.5%) in 21–40-day-old calves and the lowest (46.3%) among those that were 1–20 days old. The prevalence of the enterotoxigenic E. coli (ETEC) and Shiga toxin-producing E. coli (STEC) pathotypes was detected in 73.9% and 15.9%, respectively. The results showed no association between diarrhea and the presence of E. coli in general, ETEC or STEC. The F17 gene was the most frequently detected virulence factor in E. coli of calves of all ages regardless of diarrhea. Interestingly, the results show that the calves aged 41–60 days with F17-positive E. coli are at a higher risk for production of Shiga toxin (Stx1; 95% confidence intervals: 1.86–31.95; P = 0.005) compared to calves aged 1–20 days; no association between this finding and diarrhea was observed among the calves of this age group. Moreover, the virulence genes associated with the ETEC and STEC strains were not significantly associated with pathogenicity in this study cohort.
Conclusions
These results suggest that while the incidence of E. coli is age-related, there was no relationship linking E. coli virulence genes to calf age and diarrhea. Furthermore, the present study demonstrated that detection of E. coli strains either with or without virulence factors was not associated with diarrhea in pre-weaned calves.
Similar content being viewed by others

Background
Escherichia coli is an important cause of diarrhea in calves and has negative economic effects on the livestock industry worldwide due to high mortality and reduced growth rate [1]. Diarrheagenic E. coli are divided into six pathotypes based on the virulence properties of the bacteria and resulting clinical signs of the host; these include enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), Shiga toxin-producing E. coli (STEC; e.g., enterohemorrhagic E. coli [EHEC]), enteroinvasive E. coli (EIEC), enteroaggregative E. coli (EAEC), and diffusely adherent E. coli (DAEC) [2].
Of the six pathotypes, ETEC has been identified as a the major causative agent of calf diarrhea, whereas the others are frequently isolated from diarrheic as well as normal feces [3]. The main virulence factors contributing to diarrhea-associated ETEC are fimbrial adhesins and enterotoxins. Fimbrial adhesins (F5, F17, and F41) mediate the attachment of bacteria to epithelial surfaces in the small intestines, thereby facilitating bacterial colonization with both heat-labile (LT) and heat-stable (STa and STb) enterotoxins; the toxins promote secretion of fluids and electrolytes from the intestinal epithelial cells, which ultimately results in diarrhea [4,5,6]. The role of the STEC as an etiological agent of calf diarrhea is not yet conclusive; however, there are reports of cases of fatal STEC infection in cattle [3]. In humans, infection with STEC can result in hemorrhagic colitis, and hemolytic uremic syndrome (HUS) [7, 8]. STEC produces the Shiga toxins, Shiga toxin 1 (Stx1) and Shiga toxin 2 (Stx2), and expresses several virulence factors, including intimin (encoded by the eae gene) [9] and the STEC auto-agglutinating adhesin (saa) [10]. Domestic ruminants, particularly cattle, are known as the most important reservoirs of STEC.
The fimbrial adhesins, F17, F5, and F41 are mainly associated with ETEC in calves [11]. Likewise, the F5 and F41 virulence factors have been associated with diarrhea in animals [3, 12], although these genes are detected at decreased frequency with increasing age. E. coli-associated F17 fimbrial adhesions have been identified from animals and from humans with diarrhea and/or septicemia [13]. By contrast, STEC is not typically associated with disease in cattle and is detected more frequently in healthy animals [3]. However, STEC carried by healthy cattle may pose a potential risk to humans. The objective of the present study was to investigate the role of E. coli as a causative agent of diarrhea, to explore the prevalence of virulence genes associated with ETEC (F5, F17, F41, and sta) and with STEC (stx1, stx2, eae, and saa), and to determine the relationship between virulence genes and diarrhea in pre-weaned calves in the Republic of Korea (ROK).
Methods
Fecal samples
Fecal samples were collected from a total of 689 pre-weaned calves that were less than 61 days old during March 2017 and October 2018. The samples were taken from nine different regions within the ROK, including Anseong (n = 39), Asan (n = 95), Geochang (n = 37), Gimje (n = 106), Gyeongju (n = 6), Mungyeong (n = 112), Samrye (n = 80), Sangju (n = 8), and Yeongju (n = 206). These herds were maintained as part of a large-scale cattle breeding system; the calves had access to water and concentrates. All calves were fed with colostrum from birth. All fecal samples were collected directly from the calves by rectal palpation and were stored in plastic containers. Fecal samples were categorized based on their consistency using the scoring system (0 to 3) included in the calf health scoring guide created by the University of Wisconsin-Madison School of Veterinary Medicine (https://fyi.extension.wisc.edu/heifermgmt/files/2015/02/calf_health_scoring_chart.pdf). All samples scored at 2 or 3 were categorized as diarrhea. The fecal samples were classified as normal (n = 360) or diarrheic, typically with a pasty and watery (n = 329) consistency. Most calves used in this study were able to stand and suck at feeding time but preferred to remain in sternal recumbency. Of the 689 samples obtained, 140, 197, 129, 89, 79, and 55 were from calves at 1–10 days, 11–20 days, 21–30 days, 31–40 days, 41–50 days, and 51–60 days of age, respectively. The collected fecal samples were placed in 50 mL tubes and shipped on ice to the Animal Immunology Laboratory at Kyungpook National University.
E. coli culture, DNA extraction, and PCR
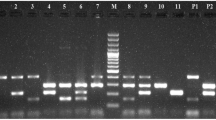
Each fecal sample was inoculated onto MacConkey agar (MBcell, Seoul, ROK) and blood agar (Asan Pharmaceutical Co., Ltd., ROK) and was incubated for 12–18 h at 37 °C. Pure colonies with appearance suggestive of E. coli were selected at random; identities were confirmed via standard biochemical tests (API 20E System, bioMérieux, France). From each sample, 2 to 10 colonies were selected, suspended in 200 μL of water, and boiled for 10 min. After centrifugation at 12,000×g for 10 min, the supernatant was used as a template for polymerase chain reaction (PCR). PCR for ETEC and STEC was performed by two multiplex PCR strategies; the first reaction targeted two factors (F17 and saa) and the second targeted six factors (F5 (= K99), F41, stx1, stx2, eae, and sta) as previously described [14,15,16,17]. PCR cycling conditions for F17 and saa included 95 °C for 10 min followed by 30 cycles of 30 s at 95 °C, annealing at 61 °C for 30 s, and 72 °C for 30 s. PCR cycling conditions for eae, F5, F41, stx1, stx2, and sta included 95 °C for 10 min, followed by 25 cycles of denaturing at 95 °C for 30 s, annealing at 50 °C for 45 s, and at 70 °C for 90 s. The reaction mixtures contained 2 μL of DNA template, 10 pM of each primer, and 10 μL of EmeraldAmp® GT PCR Master Mix (Takara, Shiga, Kusatsu, Japan) in a final volume of 20 μL. The positive controls for each target gene were obtained from Animal and Plant Quarantine Agency (Gimcheon, ROK), and distilled water was used as a negative (no template) control in each PCR experiment. Reactions were conducted in an automated DNA thermal cycler (Bio-Rad, Hercules, CA, USA). PCR products were visualized under UV light after 1.5% agarose gel electrophoresis and ethidium bromide staining.
Statistical analysis
Statistical analysis was performed using the SPSS Statistics 25 software package for Windows (SPSS Inc, Chicago, IL, USA). Binary and multinomial logistic regression analysis was used to evaluate the prevalence of E. coli and to determine any associations between the prevalence and specific independent variables, including age, virulence genes, and fecal consistency. Odd ratios (OR) with 95% confidence intervals (CIs) were calculated to assess the association between age and prevalence of E. coli and virulence genes. A P-value of less than 0.05 was considered to be statistically significant.
Results
Prevalence of E. coli according to age in pre-weaned calves
Escherichia coli was detected in 388 of a total 689 (56.3%) fecal samples. E. coli was found with a prevalence rate of 58.3% in normal feces and 54.1% in diarrheic feces, respectively. These results indicated that E. coli was not directly associated with diarrhea in pre-weaned calves (Table 1, P = 0.420). The age distribution relative to detection of E. coli is shown in Table 1. The prevalence of E. coli was significantly associated with age (P < 0.001). E. coli was detected in 66.5% of the samples from calves aged 21–40 days compared to only 46.3% of those from the 1–20-day-old group. The binomial regression analysis revealed that the prevalence of E. coli increased 2.31-fold in calves aged 21–40 days (95% CI 1.62–3.28; P < 0.001), and 2.15-fold in those aged 41–60 days (95% CI 1.42–3.25; P < 0.001) compared to that observed in calves aged 1–20 days (Table 2).
Distribution of virulence factors
The distribution of virulence factors characteristic of ETEC (F5, F17, F41, and sta) and STEC (stx1, stx2, eae, and saa) was compared to fecal consistency groups. In this study, ETEC-associated virulence factors were identified in fecal samples from 287 calves (73.9%) and STEC-associated virulence factors in samples from 61 calves (15.7%). Of all the virulence factors for ETEC and STEC evaluated, the F17 gene showed the highest prevalence (72.2%), followed by the stx1 gene (9.3%), eae gene (4.4%), and saa gene (1.8%) regardless of diarrhea (Table 3). Combinations of two or three virulence genes were also observed in both calves with and without diarrhea. Stx1 + eae, F17 + stx1, and F17 + stx1 + eae were detected in 3.6%, 3.1%, and 0.5% of the fecal samples, respectively (Table 3). There were no associations between diarrhea and any pattern of virulence genes evaluated in this study.
Distribution of virulence factors according to age
Potential associations between age and detection of virulence factors were evaluated. As shown in Table 4, the F17 gene was detected most frequently. The stx1 and eae were detected in all age groups, but their frequency was low, whereas the saa gene was found only in calves aged 1–20 days. The F5 gene was detected in fecal samples from three calves only. The F41 gene was not observed as a single factor in samples from any of the age groups, although it was detected in combination with F17 in samples from three calves (Table 4). Except for the F17, no statistical significance between any single virulence factors and age was found. Moreover, the combinations of F17 and stx1 were detected more frequently with increasing age, a finding that reached critical statistical significance (P = 0.005). Interestingly, there were no interactive effects of age and diarrhea with respect to the prevalence of virulence genes.
The binomial regression analysis revealed that detection of the F17 gene was significantly associated with age. The F17 gene was more prevalent with increasing age: notably, it was detected 2.62-fold more frequently in fecal samples from calves aged 21–40 days (95% CI 1.79–3.84; P < 0.001) and 2.44-fold more frequently in calves aged 41–60 days (95% CI 1.57–3.81; P < 0.001) than in calves aged 1-20 days. The combinations of the F17 and stx1 genes were statistically significant in calves aged 41–60 days compared to those aged 1–20 days (OR = 7.70, P = 0.005; Table 5). However, no relationship between diarrhea and the combinations of F17 and stx1 genes was observed.
Discussion
Escherichia coli has been reported as among the most common pathogens associated with diarrhea in calves, leading to high morbidity and mortality worldwide. However, remarkably few studies have been performed to evaluate the relationship between E. coli and calf diarrhea in the ROK [18,19,20]. In the present study, we examined the prevalence of E. coli according to age and explored the possibility of associations between E. coli and diarrhea in pre-weaned calves. Moreover, this study provides important new information regarding E. coli as a causative agent and its relationship to the pathogenesis of diarrhea in calves.
With the regard to the relationship between age distribution and prevalence of E. coli in fecal samples, our findings indicate that the prevalence of E. coli in fecal samples was highest among calves aged 21–40 days (66.5%) and lowest in 1–20-day-old calves (46.3%). E. coli is considered to be an important etiological agent of neonatal calf diarrhea, together with rotavirus, coronavirus, and Cryptosporidium parvum. However, our findings revealed that the presence of E. coli was not directly associated with diarrhea in pre-weaned calves. According to previous studies, E. coli can result in diarrhea during the first week of life in calves [1, 3, 11, 21]. However, our findings contradict with these results as the prevalence of E. coli was at its lowest in calves under 20 days of age in our study cohort and no mortality due to E. coli infection was identified. Interestingly, the incidence of E. coli increased significantly in association with the age, most notably after 21 days; we cannot accurately address the reasons underlying this observation, however, this finding also suggests that E. coli is unlikely to be a causative agent for diarrhea in this study. As such, we conclude that diarrhea was not caused by E. coli itself; however, this condition typically results from complex interactions among various pathotypes and virulence factors of E. coli and/or co-infections with other pathogens. While our results cannot fully rule out the possibility that E. coli serves as a risk factor for diarrhea in pre-weaned calves, our findings suggest that it is unlikely to be the main causative agent of diarrhea in this study.
ETEC strains are typically regarded as among the most important agents contributing to the pathogenesis of diarrhea. The first step in the infection process is the colonization of the intestinal epithelium, which is mediated by ETEC-associated fimbrial adhesins, primarily F5, F17, and F41 [3]. F5 and F41 have been associated with the incidence of diarrhea among neonatal calves [11]. F5 was the first virulence factor identified in ETEC, while F41 has been detected in both healthy and diarrheic calves. F41 may also have different virotypes; in other words, F41 present in healthy calves might be expressed specifically in response to immune factors associated with stress and diet, thereby promoting the onset of diarrhea. However, in this study, the F41 gene was detected only in combination with F17, and the F5 gene was found only in three calves. Moreover, the prevalence of F5 may be quite low because farms frequently inoculate calves using a vaccine that targets this antigen.
After their attachment to the intestinal mucosa, ETEC strains produce enterotoxins ST and LT; ST is more prevalent than LT and may play a major role in the pathogenesis of calf diarrhea [11]. The STa enterotoxin is produced by both porcine and bovine ETEC strains, whereas LT and STb enterotoxins are predominantly produced by porcine and human ETEC strains. There is a specific link between ETEC-associated heat-stable toxins and adhesins; STa is significantly linked to the F5/F41 combination [22]. However, the sta enterotoxin gene was detected in two calves only; this finding may explain why F5 and F41 showed a low prevalence. Taken together, these results suggest that the limited expression of fimbrial adhesins may prevent the ETEC-associated virulence factors from initiating their pathogenic actions by secreting toxins, thereby preventing the onset of diarrhea.
F17 fimbriae have been implicated in the pathogenesis of calf diarrhea as they mediate the adherence of bacterial cells to calf intestinal villi. In this study, F17 was the most predominant virulence factor detected in the E. coli strains isolated from pre-weaned calves in the ROK; F17 showed at a slightly higher prevalence in normal feces than in diarrheic feces. This could be because F17 may be related to diseases other than diarrhea in cattle. F17 has been detected in human and ruminant necrotoxigenic E. coli (NTEC) [23,24,25]. NTEC is the etiologic agent of extraintestinal infections in humans, pigs, dogs, and cats [26]; it has also been associated with intestinal infections or septicemia in calves [27]. NTEC is detected frequently in healthy calves, although the risk of NTEC infection increases after the first month of life [28]. Our results demonstrated no associations between F17 detection and diarrhea. According to our findings, the prevalence of F17 significantly increased after 21 days of age (P < 0.001) compared to those aged 1–10 days. Furthermore, F17-positive calves aged 41–60 days were at increased risk of Shiga toxin (Stx1) production (OR = 0.25, 95% CI 0.49–12.57; P = 0.005). However, we were unable to identify any associations between these genes with diarrhea. Another very important point to consider is that these calves may represent potential reservoirs of human pathogenic E. coli. The results presented here suggest that F17 has a stronger association with E. coli colonization compared to F5 and F41. Further studies will be needed to determine whether there are any specific correlations between various F17 subtypes and ETEC and NTEC pathotypes.
Stx1 was identified in 36 (9.3%) of the fecal samples from pre-weaned calves, whereas stx2 was identified in one (0.3%) diarrheic calf only. Several studies have reported that Stx1 is observed among groups of diarrheic calves, whereas Stx2 is identified as predominant among those that are healthy [7, 29]. Our results revealed that neither stx1 nor stx2 were associated with diarrhea among the calves evaluated in this study. One report suggested that Stx2 may be related to the severity of disease [30]. In humans, infection with isolates producing Stx2 toxins can result in hemorrhagic colitis and HUS [31]. In contrast, our findings suggest that STEC may be associated with fewer problems in cattle than in humans. Among the explanations for this observation, Stx toxins may have no activity at the bovine intestinal epithelium and may be unable to translocate across bovine enterocytes, in contrast to what has been observed in humans. Moreover, in the present study, stx1 was detected more frequently than stx2. Although we could not identify a relationship between diarrhea and Shiga toxin, our results also suggest that calves might act as an important reservoir for STEC infection in humans.
The eae gene product promotes interactions between bacteria and epithelial cells in vivo. In this study, the eae gene was detected in 2.4% and 2.5% of E. coli samples taken from diarrheic and normal feces, respectively. Interestingly, the eae gene was not detected in pathogenic E. coli strains isolated from cattle feces and its low prevalence has been reported in many studies [32, 33]. The role of eae in promoting the pathogenesis of diarrhea remains unclear. Of note, the saa was previously associated with an outbreak of HUS [10]. In addition, the saa gene was detected in seven calves (1.8%). These results support our observations regarding the virulence factors of STEC strains and the fact that they are unlikely to be involved in pathogenicity in the gastrointestinal tracts of calves. Consequently, the present study also highlights the important observation that healthy calves may serve as a potential reservoir for STEC.
Conclusions
The present results indicate that F17 is predominant among the virulence genes detected in fecal samples from pre-weaned calves in the ROK. Our findings revealed no associations between diarrhea and detection of ETEC or STEC strains. The incidence of E. coli was found to be age-related, although the detection of E. coli was not associated with diarrhea. Taken together, these results suggest that E. coli may serve as a risk factor and reservoir for STEC infection in humans rather than as a causative agent of calf diarrhea. This study provides useful information on the virulence factors characteristic of E. coli that are prevalent in the ROK. As such, these findings constitute an important contribution to our understanding of gastrointestinal disorders in calves and likewise provide an important database for the implementation of diagnostic, preventive, and treatment measures.
Availability of data and materials
All data generated or analyzed during this study are included in the article.
References
Lorenz I, Fagan J, More SJ. Calf health from birth to weaning. II. Management of diarrhoea in pre-weaned calves. Ir Vet J. 2011;64:9.
Croxen MA, Law RJ, Scholz R, Keeney KM, Wlodarska M, Finlay BB. Recent advances in understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013;26:822–80.
Kolenda R, Burdukiewicz M, Schierack P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. coli. Front Cell Infect Microbiol. 2015;5:23.
Guler L, Gunduz K, Ok U. Virulence factors and antimicrobial susceptibility of Escherichia coli isolated from calves in Turkey. Zoonoses Public Health. 2008;55:249–57.
Vu-Khac H, Cornick NA. Prevalence and genetic profiles of Shiga toxin-producing Escherichia coli strains isolated from buffaloes, cattle, and goats in central Vietnam. Vet Microbiol. 2008;126:356–63.
Vu-Khac H, Holoda E, Pilipcinec E, Blanco M, Blanco JE, Dahbi G, Mora A, Lopez C, Gonzalez EA, Blanco J. Serotypes, virulence genes, intimin types and PFGE profiles of Escherichia coli isolated from piglets with diarrhoea in Slovakia. Vet J. 2007;174:176–87.
Fakih I, Thiry D, Duprez JN, Saulmont M, Iguchi A, Pierard D, Jouant L, Daube G, Ogura Y, Hayashi T, et al. Identification of Shiga toxin-producing (STEC) and enteropathogenic (EPEC) Escherichia coli in diarrhoeic calves and comparative genomics of O5 bovine and human STEC. Vet Microbiol. 2017;202:16–22.
Griffin PM, Tauxe RV. The epidemiology of infections caused by Escherichia coli O157:H7, other enterohemorrhagic E. coli, and the associated hemolytic uremic syndrome. Epidemiol Rev. 1991;13:60–98.
Nataro JP, Kaper JB. Diarrheagenic Escherichia coli. Clin Microbiol Rev. 1998;11:142–201.
Paton AW, Srimanote P, Woodrow MC, Paton JC. Characterization of Saa, a novel autoagglutinating adhesin produced by locus of enterocyte effacement-negative Shiga-toxigenic Escherichia coli strains that are virulent for humans. Infect Immun. 2001;69:6999–7009.
Nagy B, Fekete PZ. Enterotoxigenic Escherichia coli in veterinary medicine. Int J Med Microbiol. 2005;295:443–54.
Toledo A, Gomez D, Cruz C, Carreon R, Lopez J, Giono S, Castro AM. Prevalence of virulence genes in Escherichia coli strains isolated from piglets in the suckling and weaning period in Mexico. J Med Microbiol. 2012;61:148–56.
Le Bouguenec C, Bertin Y. AFA and F17 adhesins produced by pathogenic Escherichia coli strains in domestic animals. Vet Res. 1999;30:317–42.
Franck SM, Bosworth BT, Moon HW. Multiplex PCR for enterotoxigenic, attaching and effacing, and Shiga toxin-producing Escherichia coli strains from calves. J Clin Microbiol. 1998;36:1795–7.
Shabana II, Zaraket H, Suzuki H. Molecular studies on diarrhea-associated Escherichia coli isolated from humans and animals in Egypt. Vet Microbiol. 2013;167:532–9.
Cortes C, De la Fuente R, Blanco J, Blanco M, Blanco JE, Dhabi G, Mora A, Justel P, Contreras A, Sanchez A, et al. Serotypes, virulence genes and intimin types of verotoxin-producing Escherichia coli and enteropathogenic E. coli isolated from healthy dairy goats in Spain. Vet Microbiol. 2005;110:67–76.
Paton AW, Paton JC. Direct detection and characterization of Shiga toxigenic Escherichia coli by multiplex PCR for stx1, stx2, eae, ehxA, and saa. J Clin Microbiol. 2002;40:271–4.
Hur J, Jeon BW, Kim YJ, Oh IG, Lee JH. Escherichia coli isolates from calf diarrhea in Korea and their virulent genetic characteristics. J Vet Med Sci. 2013;75:519–22.
Lee JH, Hur J, Stein BD. Occurrence and characteristics of enterohemorrhagic Escherichia coli O26 and O111 in calves associated with diarrhea. Vet J. 2008;176:205–9.
Kang SJ, Ryu SJ, Chae JS, Eo SK, Woo GJ, Lee JH. Occurrence and characteristics of enterohemorrhagic Escherichia coli O157 in calves associated with diarrhoea. Vet Microbiol. 2004;98:323–8.
Al Mawly J, Grinberg A, Prattley D, Moffat J, Marshall J, French N. Risk factors for neonatal calf diarrhoea and enteropathogen shedding in New Zealand dairy farms. Vet J. 2015;203:155–60.
Valat C, Forest K, Auvray F, Metayer V, Meheut T, Polizzi C, Gay E, Haenni M, Oswald E, Madec JY. Assessment of adhesins as an indicator of pathovar-associated virulence factors in bovine Escherichia coli. Appl Environ Microbiol. 2014;80:7230–4.
Ghanbarpour R, Oswald E. Characteristics and virulence genes of Escherichia coli isolated from septicemic calves in southeast of Iran. Trop Anim Health Prod. 2009;41:1091–9.
Kavitha K, Prabhakar K, Rajendran S, Uma B, Sarayu YL. Isolation of necrotoxigenic Escherichia coli from paediatric patients with acute diarrhoea. J Med Microbiol. 2010;59:503–4.
Mainil JG, Gerardin J, Jacquemin E. Identification of the F17 fimbrial subunit- and adhesin-encoding (f17A and f17G) gene variants in necrotoxigenic Escherichia coli from cattle, pigs and humans. Vet Microbiol. 2000;73:327–35.
De Rycke J, Gonzalez EA, Blanco J, Oswald E, Blanco M, Boivin R. Evidence for two types of cytotoxic necrotizing factor in human and animal clinical isolates of Escherichia coli. J Clin Microbiol. 1990;28:694–9.
Blanco J, Cid D, Blanco JE, Blanco M, Quiteira RSJA, de la Fuente R. Serogroups, toxins and antibiotic resistance of Escherichia coli strains isolated from diarrhoeic lambs in Spain. Vet Microbiol. 1996;49:209–17.
Orden JA, Ruiz-Santa-Quiteria JA, Cid D, Garcia S, de la Fuente R. Prevalence and characteristics of necrotoxigenic Escherichia coli (NTEC) strains isolated from diarrhoeic dairy calves. Vet Microbiol. 1999;66:265–73.
Wieler LH, Vieler E, Erpenstein C, Schlapp T, Steinruck H, Bauerfeind R, Byomi A, Baljer G. Shiga toxin-producing Escherichia coli strains from bovines: association of adhesion with carriage of eae and other genes. J Clin Microbiol. 1996;34:2980–4.
Meng Q, Bai X, Zhao A, Lan R, Du H, Wang T, Shi C, Yuan X, Bai X, Ji S, et al. Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 2014;14:5.
Friedrich AW, Bielaszewska M, Zhang WL, Pulz M, Kuczius T, Ammon A, Karch H. Escherichia coli harboring Shiga toxin 2 gene variants: frequency and association with clinical symptoms. J Infect Dis. 2002;185:74–84.
Hornitzky MA, Mercieca K, Bettelheim KA, Djordjevic SP. Bovine feces from animals with gastrointestinal infections are a source of serologically diverse atypical enteropathogenic Escherichia coli and Shiga toxin-producing E. coli strains that commonly possess intimin. Appl Environ Microbiol. 2005;71:3405–12.
Fremaux B, Raynaud S, Beutin L, Rozand CV. Dissemination and persistence of Shiga toxin-producing Escherichia coli (STEC) strains on French dairy farms. Vet Microbiol. 2006;117:180–91.
Acknowledgements
The authors would like to thank Mr. Jeong-Byoung Chae and Du-Gyeong Han for helping with the collection of feces.
Prior publication
Data have not been published previously.
Funding
This research was supported by the National Research Foundation of Korea (NRF), funded by the Korea government (MSIP) (No. 2018R1D1A1B07048271).
Author information
Authors and Affiliations
Contributions
KSC designed the study and drafted the manuscript. JHR, SK, and JP performed the experiments and data analyses. All authors participated in the subsequent discussions. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study did not receive approval from the Institutional Animal Care and Use Committee (IACUC) at Kyungpook National University, because the IACUC at this University evaluates laboratory animals maintained within indoor facilities and not outdoor animals. A local veterinarian collected all fecal samples and sent them to us.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ryu, JH., Kim, S., Park, J. et al. Characterization of virulence genes in Escherichia coli strains isolated from pre-weaned calves in the Republic of Korea. Acta Vet Scand 62, 45 (2020). https://doi.org/10.1186/s13028-020-00543-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13028-020-00543-1