
Abstract
Background
Epstein–Barr virus (EBV) infection is ubiquitous and in sub-Saharan Africa, occurs early in life. In a population-based rural African cohort, we leveraged historical samples from the General Population Cohort (GPC) in Uganda to examine the epidemiology of infection with EBV over time, in the era of HIV.
Methods
We used 9024 serum samples collected from the GPC in 1992, 2000, 2008, from 7576 participants across the age range (0–99 years of age) and tested for anti-EBV immunoglobulin G (IgG) antibodies to EAd, VCA, and EBNA-1 using a multiplex bead-based assay. The related gammaherpesvirus, Kaposi’s sarcoma-associated herpesvirus (KSHV) seropositivity was also determined by detection of anti-KSHV IgG antibodies to K8.1 or ORF73 measured by recombinant protein enzyme-linked immunosorbent assay. Data on sex, age, and HIV serostatus were also collected. EBV seropositivity was modeled with age (excluding those under one year, who may have had maternal antibodies), sex, HIV serostatus, and KSHV serostatus using generalized linear mixed effects models to produce beta estimates.
Results
More than 93% of children were EBV seropositive by one year of age. EBV seropositivity was significantly associated with KSHV seropositivity. Anti-EBNA-1 antibody levels decreased with increasing age and were lower on average in people living with HIV. In general, anti-EAd antibody levels increased with age, were higher in males and KSHV seropositive persons, but decreased over calendar time. Anti-VCA antibody levels increased with age and with calendar time and were higher in KSHV seropositive persons but lower in males.
Conclusions
This is the first study to identify factors associated with EBV antibodies across the entire life-course in rural sub-Saharan Africa. Consistent with other studies, EBV was near ubiquitous in the population by age one year. Patterns of antibodies show changes by age, sex and calendar time, but no association with HIV was evident, suggesting no relationship between EBV sero-epidemiology and the spread of HIV in the population over time in Uganda.
Similar content being viewed by others


Introduction
Epstein–Barr virus (EBV) is a ubiquitous human herpesvirus and is the etiological agent for several cancers including nasopharyngeal carcinoma and Burkitt lymphoma. Similar to other herpesviruses, infection with EBV is lifelong with symptomatic disease only occurring in a small subset of infected individuals. Although EBV is ubiquitous worldwide, age of infection differs by region and in sub-Saharan Africa occurs early in life, with most children infected by two years of age in some regions [1, 2]. Individuals infected with EBV at earlier ages have been shown to have worse immune control of the virus which potentially increases the risk of developing EBV-associated malignancies [3,4,5]. Elevated antibody levels to EBV proteins including early antigen (EAd), viral capsid antigen (VCA), and EBV-nuclear antigen (EBNA) have previously been found to be associated with several EBV-associated malignancies and so may act as important markers for risk of disease development [6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23].
Immunosuppression impairs the ability of the immune system to control EBV infection. In many populations, the spread of HIV led to an increase in the incidence of EBV-associated cancers [24,25,26]. With the roll-out of antiretroviral therapy (ART) in 2004, Uganda and other sub-Saharan African nations saw a decrease in some HIV-related cancers such as Kaposi’s sarcoma, but EBV-associated malignancies are still a relatively common form of cancer in the population with low survival rates [27, 28]; despite this, aspects of the epidemiology of EBV throughout the life course and, the effects of HIV infection on EBV serological responses at the population level are not well characterized. The General Population Cohort (GPC) is an open population-based cohort of over 22,000 individuals living in rural south-western Uganda [29]. Recruitment into the GPC began in 1989 to study the natural history of HIV and has continued with follow-up over the last 30 + years [30]. We used existing historical samples from this community to identify associations of sex, age, calendar time, and HIV with EBV seroprevalence and anti-EBV antibody levels.
Methods
The GPC in Kalungu District, Uganda was established in 1989, by the UK Medical Research Council and the Uganda Virus Research Institute (UVRI) and includes residents from 25 adjacent villages. Data are collected during annual census rounds, the methods for which are described in more detail elsewhere [29]. Briefly, all individuals within the study area are eligible to participate, regardless of age or sex. At each census round a questionnaire is administered and a blood sample collected. At the time of collection, serum was used to test for HIV-1 serostatus with the remainder stored in Entebbe until further analysis.
We identified serum samples collected during three GPC census rounds (1992, 2000, 2008). Samples from these years were chosen because children were included in enrollment of those rounds. Subjects were selected at random, after stratification to ensure roughly equal sex and age distributions. After stratification, up to 3000 samples from each round were randomly selected with methods described elsewhere [31]. Levels of anti-EBV immunoglobulin G (IgG) antibodies to EAd, VCA, and EBNA-1 [32] were measured in serum using a multiplex bead-based assay (MBBA) on the Luminex platform based on glutathione-S-transferase (GST) fusion capture immunosorbent assays, combined with fluorescent bead technology [1]. Briefly, for each 96-well plate, 22,500 beads were loaded in 50uL assay buffer per well (2500 per region) and 1uL of serum diluted in 49 uL of assay buffer was added. A goat anti-human IgG (y-chain) F(ab)” was used as secondary antibody (100 uL of a 0.5 ng/uL solution). Acquisition target was 50 events per bead region; antibody levels were measured as median fluorescence intensities (MFI). EBV seropositivity was determined by detection of anti-EBV IgG antibodies to either EBNA-1 (MFI > 519), VCA (MFI > 165) or EAd (MFI > 117). Kaposi’s sarcoma-associated herpesvirus (KSHV) seropositivity was determined by detection of anti-KSHV IgG antibodies to recombinant proteins K8.1 or ORF73 measured by enzyme-linked immunosorbent assay (ELISA) as previously described [33].
Serum was also tested for HIV-1 antibodies [29] with remaining samples stored at -80°C. We identified a subset of HIV seropositive individuals (all 15 + years of age) who were also part of an HIV-1 natural history cohort [34] with data on CD4 T cell counts (per mm3) and WHO clinical staging of HIV disease (stages 1–4) [35].Measures of CD4 T cell counts, HIV viral load, or disease staging were matched to participant and exact census year. Several participants had multiple measures of CD4 T cell counts or disease staging in a single year. Analyses included the average of CD4 T cell counts in a single census year. Analyses of HIV disease staging included the highest value measured in a single year, a sensitivity analysis using the lowest value in one year was completed and no difference in magnitude or direction was seen for analyses and so were not included here.
Statistical analysis
EBV seropositivity was modeled with age, sex, KSHV serostatus, HIV serostatus, CD4 T cell count, and WHO clinical stage using generalized linear mixed effects models to account for within person correlations. Anti-EBV antibody levels were log10 transformed for analysis. Associations between log-transformed anti-EBV antibodies and participant age, sex, KSHV serostatus, HIV serostatus, CD4 T cell count, and WHO clinical stage were modeled using linear mixed effects models. Models of EBV seropositivity and anti-EBV antibody levels excluded participants under one year of age to prevent inclusion of maternal antibody levels and models of anti-EBV antibody levels also excluded EBV seronegative participants. Adjusted analyses included all other covariates with age treated as a continuous variable. All analyses were completed using SAS 9.4 [SAS Institute In, Cary, NC].
Results
We tested 9077 samples collected from 7576 participants. The majority of samples came from different individuals in only one of the rounds (n = 6275), though samples from 1101 individuals were available from two rounds, and samples from 200 participants were available from all three census rounds. Overall KSHV seropositivity ranged from 91 to 94%. Participants did not differ in age, sex, or HIV serostatus by census year (Table 1). Slightly higher KSHV and EBV seropositivity rates were seen in samples of participants seen in 2000 compared to 1992 and 2008. HIV seroprevalence was 6% across census years when all participants were included, regardless of age. However, this low HIV seroprevalence was driven by the inclusion of children. In participants 15 + years of age, HIV seroprevalence was 10% in 1992 and 2000, and 11% in 2008. Among people living with HIV (PLWH), average CD4 T cell counts were similar in 1992 and 2000, but were lowest in 2008. In 1992, similar percentages of participants were in WHO HIV disease stage 1, 2, and 3. The largest percentage of participants in 2000 and 2008 were stage 3.
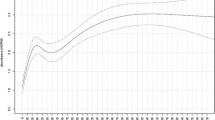
We examined the distribution of EBV seropositivity and anti-EBV antibody levels across the age range. There was a clear decrease in average antibody levels from 0 to 9 months of age for EBNA-1 and VCA antibodies correlating with the expected loss of maternal antibodies acquired during pregnancy (Fig. 1). Anti-EBNA-1 and anti-VCA antibody levels increased until the age of 5 years, after which everyone was infected in this cohort, and then anti-EBNA-1 levels were seen to decrease slightly with age, while anti-VCA levels were relatively maintained in older age groups. This pattern correlates with EBV seropositivity by age as expected. However, the same behavior is not seen with EAd where average antibody levels are low at birth and increase slightly with increasing age.

Average log(MFI) anti-EBV antibody levels by age in years. Mean (•) and standard deviation (\({ \vdash }\)) median fluorescence intensities (MFI) of anti-Epstein–Barr Virus (EBV) log-transformed antibody levels by age in years for A EBV-nuclear antigen (EBNA-1), B early antigen (EAd), C viral capsid antigen (VCA), and D EBV Seropositivity by age in years. EBV seropositivity was measured as detection of anti-EBV antibodies to EBNA-1, EAd, or VCA
Figure 2A shows results for age-specific log transformed MFI of anti-EBV antibodies by enrollment year. Average anti-EBNA-1 levels decreased with age for all enrollment cohorts. Anti-EAd levels appeared to increase with age until around 20 years and then plateau off for each enrollment cohort and anti-VCA levels appeared to have only a slight increase with age across enrollment cohorts. Figure 2B provides a visualization of the enrollment cohort specific log transformed MFI of anti-EBV antibodies by age group. There did not appear to be any effect of enrollment year on anti-EBV antibody levels though the age effect is still present.

Age and Cohort-specific anti-EBV antibody levels. A Age effects average anti-EBV antibody levels by enrollment year and B Enrollment cohort-specific average anti-EBV antibody levels by age group. All antibody levels were log transformed
We modeled risk factors for EBV seropositivity in participants over one year of age. EBV seropositivity was not associated with age or sex (Table 2). After adjusting for age, sex, and KSHV and HIV serostatus the odds of being EBV seropositive was 1.3 times higher [95% Confidence Interval (95%CI) 1.0,1.6] for participants recruited in 2000 compared to 1992. The adjusted odds of being EBV seropositive was reduced in people living with HIV [aOR = 0.7(95% CI 0.5–1.0); p = 0.049] but was twice as high in KSHV seropositive vs seronegative participants [aOR = 2.0 (95% CI 1.6–2.6); p < 0.001]. Among PLWH, there was no association between average CD4 T cell count or the highest WHO HIV disease stage in a census year and EBV seropositivity.
To identify factors associated with anti-EBV antibody levels we used linear mixed effects models restricted to EBV seropositive individuals over age one year (Fig. 3, Additional file 1: Table S1). Increasing age was associated with decreasing anti-EBNA-1 but increasing anti-EAd antibody levels. Anti-VCA antibody levels were significantly lower in 5–9- and 10–14-year-olds compared to 1–4-year-olds but the reduction in antibody levels became less marked in those over 15 years of age. In adjusted analyses, males had higher average anti-EAd levels and lower average anti-VCA levels compared to females. Anti-EBNA-1 and anti-VCA antibody levels increased in each census round while anti-EAd levels decreased with time. KSHV seropositivity was associated with higher average antibody levels for all three antigens. HIV infection was associated with lower anti-EBNA-1 and anti-EAd antibody levels. Among PLWH, there was no association between CD4 T cell count and anti-EBV antibody levels. Anti-EBNA-1 antibody levels were lower in PLWH in stage 3 compared to stage 1 HIV disease, while anti-VCA antibody levels were higher in PLWH at WHO HIV disease stage 4 compared to stage 1.

Risk Factors for anti-EBV antibody levels in EBV seropositive participants aged > 1 year. Antibody levels were measured in serum of 8495 samples from 7150 EBV seropositive participants over 1 year of age and reported as median fluorescence intensities (MFI) which were log transformed. CD4 T cell and WHO HIV Disease Stage analyses included 307 samples from 273 participants and 338 samples from 268 participants, respectively. Linear mixed effects regression modelling was used to estimate the average difference in log transformed antibody levels between groups. For age treated as a continuous variable the estimate represents the average change in log transformed antibody levels for every one-year increase in age. For CD4 T cell count the estimates represent the average change in log transformed antibody levels for every 100-cell increase in CD4 T cell counts/mm3. Beta coefficients (•) and 95% confidence intervals (95% CI) (\({ \vdash }\)) are provided. All models included a random intercept and adjusted for all other co-factors. In adjusted models age was treated as continuous. For estimates refer to Additional file 1: Table S1. Other acronyms include: Human immunodeficiency virus (HIV), Kaposi’s sarcoma-associated herpesvirus (KSHV), World Health Organization (WHO)
Discussion
Using data from the well-established General Population Cohort we were able to identify EBV antibody levels and seroprevalence patterns across the age range and by HIV serostatus in rural Uganda over a 24-year period. Our study is notable because it is population-based, maximizing generalizability, and is one of the few from sub-Saharan Africa, with the ability to look at EBV serology across the age range in otherwise healthy individuals.
We found that in our Ugandan participants EBV infection occurs in early childhood with an EBV seroprevalence of 94% over one year of age. Earlier age of EBV infection in Uganda is consistent with other studies in the region. In a cohort of Kenyan children, 100% were found to be EBV-infected by two years of age as measured by EBV DNA detection [3]. A study of infants from Eastern Uganda reported 43% were EBV-infected by the age of 50 weeks as indicated by detection of EBV DNA [36] and another that 47% of children were EBV-infected by 12 months [37]. In our population, there was a clear reduction in antibody levels against VCA and EBNA-1 from 0 to 6 months, suggesting decay of maternal antibodies, but this was not seen for EAd, where levels were already low at early ages. Our findings are similar to those from a study in Kenya which showed that, in a cohort of infants followed longitudinally, there was a clear decrease in level of maternal antibodies to VCA and EBNA-1 but not in anti-EAd IgG levels [38]. Sharp increases in anti-VCA and anti-EBNA-1 antibodies were seen after six months, likely representing initial EBV infection which peaked in children in the 2–5-year range, consistent with ages of universal infection with EBV in developing countries [5, 39].
After excluding children under one year of age, there was no difference in the probability of being EBV seropositive by age or sex, likely due to the already high infection rates early in childhood. This is consistent with a study of 1–16-year-olds from Ghana that reported that neither age nor sex were associated with EBV seroprevalence [40]. However, when examining individual anti-EBV antibody levels, we found that older age was associated with a decrease in average anti-EBNA-1 antibody levels but with increasing antibody levels to EAd and VCA. In a study of Kenyan 1–15-year-olds, VCA and EBNA-1 antibody levels appeared to be maintained across the age range [1]. In our study we also found that, on average, anti-EAd antibody levels were higher and anti-VCA were lower in males compared to females. EBNA-1 is a latent antigen, EAd is an early antigen and VCA is a late antigen. Decreasing EBNA-1 and increasing EAd and VCA antibody levels with age may be representing reactivation of EBV. Higher anti-EBV antibody levels, resulting from reactivation in an individual, have been shown to be a marker of risk for nasopharyngeal carcinoma (NPC) [7, 13]. Identifying the factors that lead to EBV reactivation are therefore important to mitigate morbidity and mortality due to EBV-associated cancers in sub-Saharan Africa such as Burkitt lymphoma and NPC.
HIV infection was moderately associated with reduced probability of EBV seropositivity after adjustment for age, sex, calendar time and KSHV seropositivity. Similarly, in a study of adults in Tanzania completed prior to ART rollout, a moderate but not statistically significant difference in antibody prevalence and titers for some EBV antigens was seen in adults living with HIV-1 compared to those without [41]. However, in contrast to our findings, in a population-based study based in Ghana, adults living with HIV were significantly more likely to be EBV seropositive [40].
When we examined specific antibody levels, we did not find a difference in anti-VCA or anti-EAd levels in people living with and without HIV in our study, though among PLWH anti-VCA antibody levels were lower in those with more severe disease. We did find that people living with HIV had on average lower EBNA-1 levels and that more severe disease was associated with lower EBNA-1 levels. A case–control study of adult Cameroonians with and without Kaposi’s sarcoma (KS) found no differences in anti-VCA, EAd, or EBNA-1 levels by HIV status, although EBV detection was significantly more frequent in blood and oral fluids of participants with HIV [42]. In another case–control study of adults, HIV infection was associated with increased prevalence of anti-EAd but not anti-EBNA-1 antibodies [43].
Seropositivity to KSHV, another gammaherpesvirus endemic to Uganda, was strongly associated with both EBV seropositivity as well as with higher average anti-EAd and anti-VCA antibody levels. In healthy Kenyan children, anti-EBNA-1 antibody levels were higher in KSHV seropositive children but there was no difference in anti-VCA antibody levels [44]. And among Cameroonian adults living with HIV, higher levels of anti-VCA, anti-gp350, and anti-EAd antibodies were identified in patients with KS, the cancer caused by KSHV, compared to persons without KS, though EBNA-1 levels did not differ [42]. Several studies have previously reported interactions between EBV and KSHV. EBV has been shown to inhibit chemically induced lytic replication of KSHV [45]. In primary effusion lymphoma (PEL), KSHV is necessary but often involves co-infection with EBV, and these two viruses have been shown to interact to alter cell proliferation [46] suggesting KSHV and EBV interactions that are important to disease development. However, the mechanisms behind these interactions are relatively undefined and require further study.
When we looked at changes in EBV serology over calendar time we found high overall EBV seropositivity in the 2000 census round compared to 1992, but no difference between EBV seropositivity in 1992 and 2008. We also identified significant increases in average anti-EBNA-1 and anti-VCA antibody levels but decreases in anti-EAd antibody levels over the three time points. The year 1992 was the peak of the HIV/AIDS epidemic in Uganda, with 18% of the general population infected; HIV prevalence decreased consistently until 2000 [47]. The initial roll out of ART in Uganda did not occur until 2004; ART was provided to patients with HIV who were severely immunocompromised (CD4 count < = 250 cells/ul or WHO clinical stage III or IV) [48]. It could be that continued HIV transmission over time was driving increases in EBV reactivation which led to higher detection of EBV seropositivity in the year 2000, but that the rollout of ART led to better control of HIV infection thus limiting EBV reactivation. However, the relatively long time between census rounds examined in this study is such that we would not be able to make any defining statements regarding the effects of HIV or ART on EBV serology over time.
Our study had some limitations. We measured anti-EBV antibodies to three antigens that are important in the clinical diagnosis of EBV infection, EBNA-1, EAd, and VCA. However, more recent studies focused on the EBV vaccine target, gp350, or the immediate early protein, ZTA, could be more informative to the state of EBV infection and potential for vaccine efficacy in this population [6, 49]. In addition, the large time gap between census rounds does not allow us to identify shorter term changes in serology patterns and their associated risk factors. Among PLWH, we were able to identify a subset of individuals with CD4 T cell counts and WHO HIV disease stage, however, HIV viral loads and date of ART initiation were not available for enough individuals to perform valid analyses and were not included. We did not have data on EBV DNA or viral loads in blood nor on malaria or other co-infections that may affect reactivation of EBV and subsequent EBV antibody levels. However, we were able to leverage a large population-based cohort in rural Africa, with samples collected during the HIV pandemic pre- and post-ART rollout. In addition, since both children and adults were tested, we were able to identify the sero-epidemiology of EBV across the age spectrum in sub-Saharan Africa.
Conclusion
We have identified early age of EBV infection in Uganda as well as several factors associated with changing anti-EBV antibody levels across two decades that included both pre- and post-ART timepoints. Our findings are an important contribution to understanding the epidemiology of EBV in sub-Saharan Africa. Future studies will be required to identify the significance of our findings and to apply them to potential intervention research.
Data availability
The datasets generated and/or analyzed during the current study are not publicly available as consent was not obtained to make data public but data are available from the corresponding author on reasonable request.
References
Piriou E, Kimmel R, Chelimo K, Middeldorp JM, Odada PS, Ploutz-Snyder R, et al. Serological evidence for long-term Epstein–Barr virus reactivation in children living in a holoendemic malaria region of Kenya. J Med Virol. 2009;81(6):1088–93.
Biggar RJ, Henle W, Fleisher G, Bocker J, Lennette ET, Henle G. Primary Epstein–Barr virus infections in African infants. I. Decline of maternal antibodies and time of infection. Int J Cancer. 1978;22(3):239–43.
Piriou E, Asito AS, Sumba PO, Fiore N, Middeldorp JM, Moormann AM, et al. Early age at time of primary Epstein–Barr virus infection results in poorly controlled viral infection in infants from Western Kenya: clues to the etiology of endemic Burkitt lymphoma. J Infect Dis. 2012;205(6):906–13.
Slyker JA, Casper C, Tapia K, Richardson B, Bunts L, Huang ML, et al. Clinical and virologic manifestations of primary Epstein–Barr virus (EBV) infection in Kenyan infants born to HIV-infected women. J Infect Dis. 2013;207(12):1798–806.
de The G, Day NE, Geser A, Lavoue MF, Ho JH, Simons MJ, et al. Sero-epidemiology of the Epstein–Barr virus: preliminary analysis of an international study - a review. IARC Sci Publ. 1975;11:3–16.
Asito AS, Piriou E, Odada PS, Fiore N, Middeldorp JM, Long C, et al. Elevated anti-Zta IgG levels and EBV viral load are associated with site of tumor presentation in endemic Burkitt’s lymphoma patients: a case control study. Infect Agent Cancer. 2010;5:13.
Cao SM, Liu Z, Jia WH, Huang QH, Liu Q, Guo X, et al. Fluctuations of Epstein–Barr virus serological antibodies and risk for nasopharyngeal carcinoma: a prospective screening study with a 20-year follow-up. PLoS ONE. 2011;6(4):e19100.
Carpenter LM, Newton R, Casabonne D, Ziegler J, Mbulaiteye S, Mbidde E, et al. Antibodies against malaria and Epstein–Barr virus in childhood Burkitt lymphoma: a case-control study in Uganda. Int J Cancer. 2008;122(6):1319–23.
Chien YC, Chen JY, Liu MY, Yang HI, Hsu MM, Chen CJ, et al. Serologic markers of Epstein–Barr virus infection and nasopharyngeal carcinoma in Taiwanese men. N Engl J Med. 2001;345(26):1877–82.
Evans AS, Gutensohn NM. A population-based case-control study of EBV and other viral antibodies among persons with Hodgkin’s disease and their siblings. Int J Cancer. 1984;34(2):149–57.
Geser A, de The G, Lenoir G, Day NE, Williams EH. Final case reporting from the Ugandan prospective study of the relationship between EBV and Burkitt’s lymphoma. Int J Cancer. 1982;29(4):397–400.
Hardell K, Carlberg M, Hardell L, Bjornfoth H, Ericson Jogsten I, Eriksson M, et al. Concentrations of organohalogen compounds and titres of antibodies to Epstein–Barr virus antigens and the risk for non-Hodgkin lymphoma. Oncol Rep. 2009;21(6):1567–76.
Ji MF, Wang DK, Yu YL, Guo YQ, Liang JS, Cheng WM, et al. Sustained elevation of Epstein–Barr virus antibody levels preceding clinical onset of nasopharyngeal carcinoma. Br J Cancer. 2007;96(4):623–30.
Levin LI, Chang ET, Ambinder RF, Lennette ET, Rubertone MV, Mann RB, et al. Atypical prediagnosis Epstein–Barr virus serology restricted to EBV-positive Hodgkin lymphoma. Blood. 2012;120(18):3750–5.
Levine PH, Stemmermann G, Lennette ET, Hildesheim A, Shibata D, Nomura A. Elevated antibody titers to Epstein–Barr virus prior to the diagnosis of Epstein-Barr-virus-associated gastric adenocarcinoma. Int J Cancer. 1995;60(5):642–4.
Mueller N, Evans A, Harris NL, Comstock GW, Jellum E, Magnus K, et al. Hodgkin’s disease and Epstein-Barr virus. Altered antibody pattern before diagnosis. N Engl J Med. 1989;320(11):689–95.
Mueller N, Mohar A, Evans A, Harris NL, Comstock GW, Jellum E, et al. Epstein–Barr virus antibody patterns preceding the diagnosis of non-Hodgkin’s lymphoma. Int J Cancer. 1991;49(3):387–93.
Mueller NE, Lennette ET, Dupnik K, Birmann BM. Antibody titers against EBNA1 and EBNA2 in relation to Hodgkin lymphoma and history of infectious mononucleosis. Int J Cancer. 2012;130(12):2886–91.
Mutalima N, Molyneux E, Jaffe H, Kamiza S, Borgstein E, Mkandawire N, et al. Associations between Burkitt lymphoma among children in Malawi and infection with HIV, EBV and malaria: results from a case-control study. PLoS ONE. 2008;3(6):e2505.
Rothman N, Cantor KP, Blair A, Bush D, Brock JW, Helzlsouer K, et al. A nested case-control study of non-Hodgkin lymphoma and serum organochlorine residues. Lancet. 1997;350(9073):240–4.
Shinkura R, Yamamoto N, Koriyama C, Shinmura Y, Eizuru Y, Tokunaga M. Epstein–Barr virus-specific antibodies in Epstein–Barr virus-positive and -negative gastric carcinoma cases in Japan. J Med Virol. 2000;60(4):411–6.
Xu J, Ahmad A, Blagdon M, D’Addario M, Jones JF, Dolcetti R, et al. The Epstein–Barr virus (EBV) major envelope glycoprotein gp350/220-specific antibody reactivities in the sera of patients with different EBV-associated diseases. Int J Cancer. 1998;79(5):481–6.
Yu KJ, Hsu WL, Pfeiffer RM, Chiang CJ, Wang CP, Lou PJ, et al. Prognostic utility of anti-EBV antibody testing for defining NPC risk among individuals from high-risk NPC families. Clin Cancer Res. 2011;17(7):1906–14.
Wabinga HR, Nambooze S, Amulen PM, Okello C, Mbus L, Parkin DM. Trends in the incidence of cancer in Kampala, Uganda 1991–2010. Int J Cancer. 2014;135(2):432–9.
Wabinga HR, Parkin DM, Wabwire-Mangen F, Nambooze S. Trends in cancer incidence in Kyadondo County, Uganda, 1960–1997. Br J Cancer. 2000;82(9):1585–92.
Biological agents. IARC Monogr Eval Carcinog Risks Hum. 2012;100(Pt B):1–441.
Mutyaba I, Wabinga HR, Orem J, Casper C, Phipps W. Presentation and outcomes of childhood cancer patients at Uganda Cancer Institute. Glob Pediatr Health. 2019;6:2333794X19849749.
Stefan C, Bray F, Ferlay J, Liu B, Maxwell PD. Cancer of childhood in sub-Saharan Africa. Ecancermedicalscience. 2017;11:755.
Asiki G, Murphy G, Nakiyingi-Miiro J, Seeley J, Nsubuga RN, Karabarinde A, et al. The general population cohort in rural south-western Uganda: a platform for communicable and non-communicable disease studies. Int J Epidemiol. 2013;42(1):129–41.
Nunn AJ, Kengeya-Kayondo JF, Malamba SS, Seeley JA, Mulder DW. Risk factors for HIV-1 infection in adults in a rural Ugandan community: a population study. AIDS. 1994;8(1):81–6.
Newton R, Labo N, Wakeham K, Miley W, Asiki G, Johnston WT, et al. Kaposi sarcoma-associated herpesvirus in a rural Ugandan cohort, 1992–2008. J Infect Dis. 2018;217(2):263–9.
Fachiroh J, Paramita DK, Hariwiyanto B, Harijadi A, Dahlia HL, Indrasari SR, et al. Single-assay combination of Epstein–Barr Virus (EBV) EBNA1- and viral capsid antigen-p18-derived synthetic peptides for measuring anti-EBV immunoglobulin G (IgG) and IgA antibody levels in sera from nasopharyngeal carcinoma patients: options for field screening. J Clin Microbiol. 2006;44(4):1459–67.
Mbisa GL, Miley W, Gamache CJ, Gillette WK, Esposito D, Hopkins R, et al. Detection of antibodies to Kaposi’s sarcoma-associated herpesvirus: a new approach using K8.1 ELISA and a newly developed recombinant LANA ELISA. J Immunol Methods. 2010;356(1–2):39–46.
Morgan D, Malamba SS, Maude GH, Okongo MJ, Wagner HU, Mulder DW, et al. An HIV-1 natural history cohort and survival times in rural Uganda. AIDS. 1997;11(5):633–40.
World Health Organization. Interim WHO clinical staging of HVI/AIDS and HIV/AIDS case definitions for surveillance: African region. Geneva: World Health Organization; 2005.
Montoya-Ferrer A, Sanosyan A, Fayd’herbe de Maudave A, Pisoni A, Bollore K, Moles JP, et al. Clinical and biological factors associated with early Epstein–Barr VIRUS infection in human immunodeficiency virus-exposed uninfected infants in eastern Uganda. Clin Infect Dis. 2021;72(6):1026–32.
Minab R, Bu W, Nguyen H, Wall A, Sholukh AM, Huang ML, et al. Maternal Epstein–Barr virus-specific antibodies and risk of infection in Ugandan infants. J Infect Dis. 2021;223(11):1897–904.
Reynaldi A, Schlub TE, Piriou E, Ogolla S, Sumba OP, Moormann AM, et al. Modeling of EBV infection and antibody responses in Kenyan infants with different levels of malaria exposure shows maternal antibody decay is a major determinant of early EBV infection. J Infect Dis. 2016;214(9):1390–8.
Gantt S, Orem J, Krantz EM, Morrow RA, Selke S, Huang ML, et al. Prospective characterization of the risk factors for transmission and symptoms of primary human herpesvirus infections among Ugandan infants. J Infect Dis. 2016;214(1):36–44.
Adjei AA, Armah HB, Gbagbo F, Boamah I, Adu-Gyamfi C, Asare I. Seroprevalence of HHV-8, CMV, and EBV among the general population in Ghana. West Africa BMC Infect Dis. 2008;8:111.
Essers S, Schwinn A, ter Meulen J, von Lips H, Dietz K, Mhalu FS, et al. Seroepidemiological correlations of antibodies to human herpesviruses and human immunodeficiency virus type 1 in African patients. Eur J Epidemiol. 1991;7(6):658–64.
Labo N, Marshall V, Miley W, Davis E, McCann B, Stolka KB, et al. Mutual detection of Kaposi’s sarcoma-associated herpesvirus and Epstein–Barr virus in blood and saliva of Cameroonians with and without Kaposi’s sarcoma. Int J Cancer. 2019;145(9):2468–77.
Kayamba V, Monze M, Asombang AW, Zyambo K, Kelly P. Serological response to Epstein–Barr virus early antigen is associated with gastric cancer and human immunodeficiency virus infection in Zambian adults: a case-control study. Pan Afr Med J. 2016;23:45.
Oluoch PO, Oduor CI, Forconi CS, Ong’echa JM, Munz C, Dittmer DP, et al. Kaposi Sarcoma-associated herpesvirus infection and endemic Burkitt’s lymphoma. J Infect Dis. 2020;222:111.
Xu D, Coleman T, Zhang J, Fagot A, Kotalik C, Zhao L, et al. Epstein–Barr virus inhibits Kaposi’s sarcoma-associated herpesvirus lytic replication in primary effusion lymphomas. J Virol. 2007;81(11):6068–78.
Boni M, Rieble L, Munz C. Co-infection of the Epstein–Barr virus and the Kaposi sarcoma-associated herpesvirus. Viruses. 2022;14(12):2709.
Stoneburner RL, Low-Beer D. Population-level HIV declines and behavioral risk avoidance in Uganda. Science. 2004;304(5671):714–8.
Ssempijja V, Namulema E, Ankunda R, Quinn TC, Cobelens F, Hoog AV, et al. Temporal trends of early mortality and its risk factors in HIV-infected adults initiating antiretroviral therapy in Uganda. EClinicalMedicine. 2020;28:100600.
Smith NA, Baresel PC, Jackson CL, Ogolla S, Toko EN, Heit S, et al. Differences in the Epstein–Barr virus gp350 IgA antibody response are associated with increased risk for coinfection with a second strain of Epstein–Barr virus. J Infect Dis. 2019;219(6):955–63.
Acknowledgements
We would like to acknowledge our field staff, as well as, all the study participants, without whom this work would not have been possible.
Funding
This work was supported by the Welcome Trust (Grant number: 090132) and with federal funds from the National Cancer Institute, National Institutes of Health, under Contract No. HHSN261201500003I and NCI contract 75N91019D00024 to DW. The GPC is funded by the UK Medical Research Council (MRC) and the UK Department for International Development (DFID) under the MRC/DFID Concordat agreement. The funders had no role in study design, data collection, analysis or interpretation of the data. The content of this manuscript is solely the responsibility of the authors and does not necessarily represent the official views of NCI or NIH.
Author information
Authors and Affiliations
Contributions
KRS completed final analyses, assisted in interpretation of data, and drafted the manuscript. JM, GA, and AN contributed to study deign, data collection and manuscript development NL and WM analyzed EBV serology data and contributed to data interpretation, and manuscript development. RB contributed to data interpretation and manuscript development. RR assisted substantially with study concept, data interpretation and manuscript development. TWJ assisted with study design, interpretation of data, and manuscript development. RN and DW made substantial contributions to the study conception and design, data collection, interpretation, and manuscript development. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval was obtained by the Uganda Virus Research Institute Research Ethics Committee (UVRI-REC) and Uganda National Council for Science and Technology (UNCST). All participants provided informed consent.
Consent for publication
Not applicable.
Previous presentation
These data have not previously been presented at a scientific meeting.
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Risk Factors for log transformed anti-EBV antibody levels (MFI) in participants 1+ years of age (n=8495)1 using linear mixed effects regression2.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sabourin, K.R., Mugisha, J., Asiki, G. et al. Epstein–Barr virus (EBV) antibody changes over time in a general population cohort in rural Uganda, 1992–2008. Infect Agents Cancer 18, 55 (2023). https://doi.org/10.1186/s13027-023-00534-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13027-023-00534-7