
Abstract
Background
Both SARS-CoV-2 mRNA-based vaccines [BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna)] have shown high efficacy, with very modest side effects in limiting transmission of SARS-CoV-2 and in preventing the severe COVID-19 disease, characterized by a worrying high occupation of intensive care units (ICU), high frequency of intubation and ultimately high mortality rate. At the INT, in Naples, only the BNT162b2/Pfizer vaccine has been administered to cancer patients and healthcare professionals aged 16 and over. In the present study, the antibody response levels and their decline were monitored in an interval of 6–9 months after vaccine administration in the two different cohorts of workers of the INT – IRCCS "Fondazione Pascale" Cancer Center (Naples, Italy): the group of individuals previously infected with SARS-CoV-2 and vaccinated with a single dose; and that of individuals negative for previous exposure to SARS-CoV-2 vaccinated with two doses 21 days apart.
Methods
Specific anti-RBD (receptor-binding domain) titers against trimeric spike glycoprotein (S) of SARS-CoV-2 by Roche Elecsys Anti-SARS-CoV-2 S ECLIA immunoassay were determined in serum samples of 27 healthcare workers with a previously documented history of SARS-CoV-2 infection and 123 healthcare workers without, during antibody titers’ monitoring. Moreover, geometric mean titers (GMT) and relative fold changes (FC) were calculated.
Results
Bimodal titer decline was observed in both previously infected and uninfected SARS-CoV-2 subjects. A first rapid decline was followed by a progressive slow decline in the 6/9 month-period before the further vaccine boost. The trend was explained by 2 different mathematical models, exponential and power function, the latter revealing as predictive of antibody titer decline either in infected or in not previously infected ones. The value of the prolonged lower vaccine titer was about 1 log below in the 6/9-month interval after the single dose for previously infected individuals with SARS-CoV-2 and the two doses for those not previously infected. The titer change, after the boost dose administration, on the other hand, was ≥ 1.5 FC higher than the titers at the 6/9-month time-points in both cohorts. A similar quantitative immune titer was observed in both cohorts 8 days after the last boost dose. The subsequent immunoresponse trend remains to be verified.
Discussion
The results show that a very rapid first decline, from the highest antibody peak, was followed by a very slow decline which ensured immune protection lasting more than 6 months. The apparent absence of adverse effects of the rapid decline on the vaccine's immune protective role has been related to a large majority of low avidity antibodies induced by current vaccines. High avidity antibodies with prolonged anti-transmission efficacy show a longer half-life and are lost over a longer interval period. The cellular immunity, capable of preventing severe clinical diseases, lasts much longer. The unbalanced dual activity (cellular vs humoral) while effective in limiting ICU pressure and overall mortality, does not protect against transmission of SARS-CoV-2, resulting in high circulation of the virus among unvaccinated subjects, including the younger population, and the continuous production of variants characterized by changes in transmissibility and pathogenicity. The high mutation rate, peculiar to the RNA virus, can however lead to a dual opposite results: selection of defective and less efficient viruses up to extinction; risk of more efficiently transmitted variants as the current omicron pandemic.
Conclusions
In conclusion the current bimodal antibody-titer decline, following BNT162b2 mRNA anti-SARS-CoV-2 vaccination, needs a further extended analysis to verify the protective borderline levels of immunity and the optimal administration schedule of vaccine boosters. Our current results can contribute to such goal, besides a direct comparison of other FDA-approved and candidate vaccines.
Similar content being viewed by others
Background
Severe Acute Respiratory Syndrome Coronavirus-2 (SARS-CoV-2), a positive-sense single-stranded RNA virus belonging to the Coronaviridae family, causes coronavirus disease 2019 (COVID-19), a respiratory syndrome evolving frequently to severe pneumonia, respiratory and multi-visceral failure and causing death in some patients with comorbidity [1].
Preventive protection against many infectious diseases is mediated by a functional, persistent antibody response, which is therefore a critical immune correlate for many licensed human vaccines. The durability of vaccine-acquired antibody responses varies greatly among antigens [2]. Antibodies induced by viral infections, or by vaccination with live-attenuated viruses, can persist for decades. However, most vaccines based on protein antigens require repeated immunizations to generate immunological memory, and to maintain antibody responses above protective levels [3]. The level of antigen–antibody binding avidity, a qualitative response index, can also correlate with protection. Nevertheless, low-avidity antibodies have been associated with antibody-mediated disease enhancement following pandemic influenza vaccinations [4, 5].
Moreover, inadequate levels of avidity maturation (the latter defined as the increase of avidity over time) can heighten susceptibility to viral infection [6]. Thus, both quantitative and qualitative yardsticks can determine vaccine efficacy. Vaccine adjuvants are linked to both of these aspects of the antibody response. By enhancing innate immunity, they promote activation of naïve B cells and CD4 + T cells [7].
The immune system represents an important component against the viral infection by the titer and avidity of neutralizing antibodies production. The trimeric spike glycoprotein (S) of SARS-CoV-2 is a key target for virus neutralizing antibodies and the prime candidate for vaccine development [8]. The protein S binds its cellular receptor on the host cells, human angiotensin converting enzyme 2 (ACE2), through a receptor-binding domain (RBD) [9].
Both SARS-CoV-2 mRNA-based vaccines containing the messenger RNA that encodes the SARS-CoV-2 S in small lipid particles [BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna)] have shown high efficacy, with very modest side effects in limiting transmission of SARS-CoV-2 and in preventing the severe COVID-19 disease, characterized by a worrying high occupation of intensive care units (ICU), high frequency of intubation and ultimately high mortality rate.
On 22nd December 2020, the Italian regulatory agency for drugs AIFA (Agenzia Italiana del Farmaco) authorized in Italy the use of BNT162b2/Pfizer vaccine, in 2 doses with an interval of 21 days between the doses [10]. Recent studies have found that subjects infected with COVID-19 develop higher antibody titers after vaccination [11] and present protective immunity for at least 6 months [12], but the impact of previous exposure to SARS-CoV-2 on immune response elicited by the vaccines needs to be further verified in larger trial studies. Several studies have shown that the immune response to the vaccine after the first dose is substantially more pronounced in individuals with pre-existing immunity and it is similar to the immune response developed after the second dose in individuals not previously infected [11, 13,14,15]. However, few data are available on the accurate monitoring of the titers’ decline, response to further boosters, optimal vaccine dosage and role of different adjuvants on vaccine efficacy.
In the present study, the antibody response levels and their decline were monitored in an interval of 6/9 months after vaccine administration in the two different cohorts of workers of the INT – IRCCS "Fondazione Pascale" Cancer Center (Naples, Italy), established since 2020 [16]: the group of individuals previously infected with SARS-CoV-2 and vaccinated with a single dose; and that of individuals negative for previous exposure to SARS-CoV-2 vaccinated with two doses 21 days apart.
Materials and methods
Sample size
According to our national recommendations, the internal health surveillance program for healthcare workers implemented the following BNT162b2/Pfizer vaccination schedule: 1 dose was administered to subjects previously infected to SARS-CoV-2, and 2 doses (with an interval of 21 days) to subjects not infected. Most of healthcare providers underwent also boost dose. The program contemplated the evaluation of antibody responses by determining anti-RBD titers at three times: the day before vaccination (baseline anti-S antibody titer), 20 days after the first dose and 8 days after the second dose for not previously infected workers and 8 days after the unique dose for previously infected healthcare providers. Further data were collected by determining antibody titers 1 month after the preliminary completion of vaccination cycle and 8 days after the boost dose administration for both cohorts. Time points were defined as follows:
-
For not infected subjects:
-
T0 = baseline pre-vaccination.
-
T1 = 20 days after the first dose.
-
T2 = 8 days after the second dose, distinguishing:
-
T2 I = 8 days after the second dose.
-
T2 II = 1 month after the second dose.
-
-
T3 = 3 months after the second dose.
-
T6 = 6 months after the second dose.
-
T9 = 9 months after the second dose.
-
T boost = 8 days after the administration of a further boost dose.
-
-
For previously infected subjects:
-
T0 = baseline pre-vaccination after SARS-CoV-2 infection.
-
T1 = 8 days after the unique dose.
-
T2 = 1 month after the unique dose.
-
T3 = 3 months after the unique dose.
-
T6 = 6 months after the unique dose.
-
T9 = 9 months after the unique dose.
-
T boost = 8 days after the administration of a further boost dose.
-
Data regarding 150 healthcare workers of INT – IRCCS “Fondazione Pascale” Cancer Centre (27 and 123 with history/no history of COVID-19 infection, defined as infected and not infected subjects, respectively) were analyzed. Moreover, in order to exactly quantify and extend the analysis to a 9-month interval two subgroups were extrapolated for the previously COVID-infected (n = 3) and not infected subjects (n = 20). The study was performed under the statements of Declaration of Helsinki and approved by the local Institutional Ethics Committee. All participants gave their informed consent.
Assay
Roche Elecsys Anti-SARS-CoV-2 S electrochemiluminescence immunoassay (ECLIA) for the in vitro quantitative determination of antibodies (including IgG) against spike RBD of SARS-CoV-2 in human serum was performed on Roche Cobas e 801 module. According to the manufacturer, the correlation test between Roche Elecsys Anti-SARS-CoV-2 S units per mL and WHO International Standards for anti-SARS-CoV-2 immunoglobulins showed an excellent correlation (r2 = 0.9992, slope = 0.972, intercept = 0.0072), thus allowing to consider specific Roche Elecsys Anti-SARS-CoV-2 S U/mL units equivalent to WHO International Standard BAU/mL (Binding Arbitrary Units per mL). Measuring range spanned from 0.4 BAU/mL to 250.0 BAU/mL, requiring a 1:10 dilution for samples with concentrations > 250.0 BAU/mL, extending the measuring range until to 2500.0 BAU/mL; values higher than 0.8 BAU/mL were considered positive. Serum samples with antibody titers > 2500.0 BAU/mL, from the selected subgroups above specified, were further diluted in order to obtain the exact quantitative concentration.
Statistical analysis
Statistical analysis was performed by using the Statistical Package for Social Science (SPSS Inc., Chicago, IL, USA), version 27.0. Distribution of variables was evaluated by Shapiro–Wilk test; parametric data were represented as mean ± standard deviation (SD), whilst non-parametric variables were expressed as median (IR—Interquartile Range). Two-tailed Mann–Whitney for independent variables test was used to compare groups. Values lower than 0.4 BAU/mL were assumed as 0.4 and values higher than 2500.0 BAU/mL were reported as 2500.0; p values < 0.05 were considered statistically significant. Moreover, geometric mean titers (GMT) and relative fold changes (FC) were calculated. To overcome the novelty of general bead-based linear models, used to evaluate post-to-pre vaccination antibody titer increase or subsequent titer decline, the used statistic methods were extrapolated from Zaccaro et al. [17]. Different mathematical models were tested and evaluated by fitting median values extrapolated by immunoassays at different time-points for previously and not previously infected healthcare providers, in order to study and explain the antibody decline kinetics.
Results
The enrolled cohort of healthcare workers (27 previously infected and 123 not infected) was represented as follows: 27 seropositive cases, 17 female and 10 male subjects with an overall mean age of 49.1 years (SD ± 8.9, range 34–65); and 123 seronegative cases, 80 female and 43 male subjects with an overall mean age of 46.2 years (SD ± 11.4, range 23–67) (Table 1).
Data regarding monitoring during the interval from baseline to boost dose administration are showed in Table 2 and Fig. 1. At each time-point, two-tailed Mann–Whitney U test showed significantly higher antibody titer concentrations in infected subjects in comparison with not previously infected ones, except for T boost at which previously infected workers presented higher (but not statistically significant) levels of antibodies.

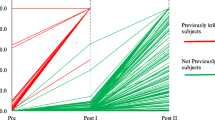
Vaccine immune response monitoring in previously infected and not previously infected cohorts of healthcare providers. For not infected subjects (n = 123): T0 = baseline pre-vaccination, T1 = 20 days after the first dose, T2 = 8 days after the second dose, T3 = 3 months after the second dose, T6 = 6 months after the second dose, T9 = 9 months after the second dose, T boost = 8 days after the administration of boost dose. For previously infected subjects (n = 27): T0 = baseline pre-vaccination after SARS-CoV-2 infection, T1 = 8 days after the unique dose, T2 = 1 month after the unique dose, T3 = 3 months after the unique dose, T6 = 6 months after the unique dose, T9 = 9 months after the unique dose, T boost = 8 days after the administration of a further boost dose. Orange lines: subjects previously infected with SARS-CoV-2. Yellow lines: subjects not previously infected and with T2 titers < 500.0 BAU/mL. Green lines: subjects not previously infected and with T2 titers < 1000.0 BAU/mL and ≥ 500.0 BAU/mL. Grey lines: subjects not previously infected and with T2 titers < 2000.0 BAU/mL and ≥ 1000.0 BAU/mL. Blue lines: subjects not previously infected and with T2 titers ≥ 2000.0 BAU/mL
Despite the upper limit of 2500.0 BAU/mL imposed by the Roche method, a bimodal titer decline, before the further boost dose administration, was observed in most subjects. This finding was confirmed on data obtained from further diluted samples. In order to better define antibody titers’ trend in the interval immediately after the administration of the unique dose or the second dose, the T2 for previously infected subjects, and T2 I–T2 II time-points for not infected ones, respectively, were included (Table 3, Fig. 2a–b). Such analysis allowed to show for not previously infected workers a first rapid decline in the interval T2 I–T2 II with a negative slope of − 2495.5 followed by a progressive slow decline in the 6/9 month-period before the further vaccine boost (slope of − 847.3 in the interval T2 II–T3, − 342.2 in the interval T3–T6, − 123.1 in the interval T6–T9), with a slope of − 3342.9 in the interval T2 I–T3 and an overall negative slope of − 3808.2 from T2 I to T9 (Table 3, Fig. 2a). Peculiarly, in the cohort of previously infected workers, after an unexpected and unreported initial median increase of antibody titer in the T1–T2 interval (positive slope of 2550), a rapid decline followed by a slower decrease was observed (Table 3), with a negative slope of − 7562 in the interval T2–T3, − 1009 (T3–T6), − 582 (T6–T9) and an overall negative slope of − 9153 reported from T2 to T9 (Table 3, Fig. 2b). Both in previously infected cohort (n = 20) and in not previously infected one (n = 3), 2 different mathematical models were analyzed to study and calculate the antibody decline kinetics: an exponential vs. a power function model.

a Vaccine immune response decline in not previously infected healthcare providers’ diluted samples. T0 = baseline pre-vaccination, T1 = 20 days after the first dose, T2 I = 8 days after the second dose, T2 II = 1 month after the second dose, T3 = 3 months after the second dose, T6 = 6 months after the second dose, T9 = 9 months after the second dose. Each colored line represents a subject (n = 20). b Vaccine immune response decline in previously infected healthcare providers’ diluted samples. T0 = baseline pre-vaccination after SARS-CoV-2 infection, T1 = 8 days after the unique dose, T2 = 1 month after the unique dose, T3 = 3 months after the unique dose, T6 = 6 months after the unique dose, T9 = 9 months after the unique dose. Each colored line represents a subject (n = 3)
In not previously infected workers the following equations were detected:
for exponential and power function models, with R2 = 0.9502 and 0.9971, respectively (Fig. 3a–b).

a Exponential model to study and calculate the antibody decline kinetics in not previously infected subjects. Blue dot line indicates the graphic representation of the exponential model, the single blue dots the median values at different time-points and black vertical lines the interquartile range of distributions. b Power function model to study and calculate the antibody decline kinetics in not previously infected subjects. Blue dot line indicates the graphic representation of the power function model, the single blue dots the median values at different time-points and black vertical lines the interquartile range of distributions
In previously infected subjects the following equations were detected:
for exponential and power function models, with R2 = 0.9136 and 0.988, respectively (Fig. 4a–b).

a Exponential model to study and calculate the antibody decline kinetics in previously infected subjects. Blue dot line indicates the graphic representation of the exponential model, the single blue dots the median values at different time-points and black vertical lines the interquartile range of distributions. b Power function model to study and calculate the antibody decline kinetics in previously infected subjects. Blue dot line indicates the graphic representation of the power function model, the single blue dots the median values at different time-points and black vertical lines the interquartile range of distributions
By calculating GMT at each time-point and relative FC, it was found that the prolonged lower vaccine titer was about 1 log below in the 6/9-month interval after the single dose for previously infected individuals with SARS-CoV-2 and the two doses for those not previously infected (Table 4). The mean change in titer after the boost dose administration, on the other hand, was equal to or more than 1.5 FC higher than the T6–T9 time-points in both cohorts (1.5 and 1.7 higher than T6 and T9 for previously infected workers, and 1.7 and 1.9 higher than T6 and T9 for not previously infected ones, respectively) (Table 4). A mean increase of 0.7 and 0.9 FC was found comparing T boost–T2 time-points in infected subjects and in not previously infected ones, respectively (Table 4). At T boost time-point, previously infected workers presented higher (but not statistically significant) levels of antibodies in respect to not previously infected cohort (Table 4, Fig. 5a–b). At T boost time-point, however, a similar anti-S antibody titer was observed in previously infected and not-infected subjects suggesting that the vaccination boost was able to induce an equivalent immune response among the two groups eliminating any difference related to the previous infection.

a Vaccine immune response increase after boost dose administration in not previously infected healthcare providers’ diluted samples. T0 = baseline pre-vaccination, T1 = 20 days after the first dose, T2 I = 8 days after the second dose, T2 II = 1 month after the second dose, T3 = 3 months after the second dose, T6 = 6 months after the second dose, T9 = 9 months after the second dose, T boost = 8 days after the administration of a further boost dose. Each colored line represents a subject. b Vaccine immune response increase after boost dose administration in previously infected healthcare providers’ diluted samples. T0 = baseline pre-vaccination after SARS-CoV-2 infection, T1 = 8 days after the unique dose, T2 = 1 month after the unique dose, T3 = 3 months after the unique dose, T6 = 6 months after the unique dose, T9 = 9 months after the unique dose, T boost = 8 days after the administration of a further boost dose. Each colored line represents a subject
Discussion
In accordance with first data reported in literature [11, 13, 18], our findings confirm that administration of a single dose to previously infected subjects is sufficient to elicit an adequate immunoresponse, the latter resulting more sustained in terms of concentration and duration if compared to antibody response of not previously infected cohort. Nevertheless, for the first time in literature, we demonstrated a bimodal decline kinetic characterized by a very rapid first decline from the highest antibody peak (1 month after the unique vaccine dose in previously infected workers and 8 days after the second dose in not previously infected one), followed by a slow decline which ensures a long-term immune protection lasting more than 6 months. Our data fit a power function decline curve, which allows the calculation and identification of a specific trend that can predict the decline rate of anti-RBD antibody titer, surrogate of neutralizing protective antibodies [19, 20], with excellent accuracy in both previously infected and uninfected individuals. Moreover, the initial more pronounced decline observed in previously infected subjects (demonstrated by higher negative slopes) was counterbalanced by the less robust response to the additional boost dose, resulting in overlapping of antibody titers in the 2 different cohorts 8 days after boost vaccine administration (T boost).
The apparent absence of negative effects related to the rapid decline on the vaccine's immune protective role has been referred to a large majority of low avidity antibodies induced in the early post-vaccination stages [21,22,23]. High avidity antibodies with prolonged anti-transmission efficacy show a longer half-life and are lost over a longer interval period. Protective humoral immune response is generally characterized by a parabolic trend with a protective plateau, several months long, following natural infection as well as vaccine induced active immunity, also for the latest developed subunits vaccines. As paradigm the long-lasting immune response following a single dose of L1-based Virus-like particles (VLPs), the anti- DNA papillomavirus (HPV) vaccine [24], and following multiple doses of a recombinant E2-based vaccine against the RNA-hepatitis E virus (HEV) [25] can be considered. The trend observed in the current anti-SARS-CoV-2 vaccination shows, instead, after a rapid increment an asymptotic hyperbolic trend towards a modest not-protective immune response, with higher risk of further infections, gradually declining below a titer which should represent the protective immune threshold. The high titer stage is lost in less than 30 days and is referred mainly to low-avidity antibodies, leaving the protective activity to the low-titer stage ranging from 1893.4 to 469.5 BAU/mL for the subject without previous COVID infection and from 2806.4 to 1152.2 BAU/mL for those with a previous COVID infection (GMT from Fig. 2a and b, respectively). The alternative dramatic inference would be that the protective period is just a month.
The identification and quantification of the protective lower antibody level, whose avidity along with neutralizing activity increases over time [26], becomes relevant for monitoring the individual protection level and selecting the appropriate further boosting time. Moreover, it becomes a reference value to compare the efficacy of other vaccines as well as their delivery and formulation (including adjuvants) besides the possibility to develop pre-vaccination strategies to improve the immune response to vaccines [27]. The limitation of the current study is the low number (n = 3) of previously infected vaccinees. Such number limit, however, is not crucial for this specific manuscript given that the immunoresponse decline of the previously infected follows the same curve (although starting from a higher antibody titer) of the non-previously infected vaccinees. It suggests that also the humoral immune response following the vaccination is very similar for the lower antibody level with a FC of 0.4 between the previously infected and non-previously infected at 9 months after the vaccination.
Fortunately, the cellular immunity, capable of preventing severe clinical diseases, seems to last much longer. The complementary immune response (short-lived humoral immunity vs long-lasting cellular immunity), although effective in limiting overall mortality, with the consequent reduction of pressure in the ICU, does not prevent and does not protect against transmission of SARS-CoV-2, resulting in high circulation of the virus among unvaccinated subjects, including the younger population, and the continuous production of variants characterized by changes in transmissibility and pathogenicity. The high mutation rate, peculiar to the RNA virus, can however lead to a dual opposite result: selection of defective and less efficient viruses up to extinction [28, 29]; risk of more efficiently transmitted variants as the current omicron pandemic [30, 31].
Conclusions
In conclusion, further extended analyses are mandatory to confirm our preliminary findings in larger cohorts of subjects (including patients and immunocompromised individuals), in order to monitor antibody titers also after the administration of further boost doses, verify the protective borderline levels of immunity and define the optimal administration schedule of vaccine boosters. Our results, highlighting the current bimodal antibody-titer decline after BNT162b2 mRNA anti-SARS-CoV-2 vaccination, can contribute to such goal, besides a direct comparison of other FDA-approved and candidate vaccines.
Availability of data and materials
All relevant data and their evaluation are reported in the manuscript.
Abbreviations
- ACE2:
-
Human Angiotensin Converting Enzyme 2
- AIFA:
-
Agenzia Italiana del Farmaco, the Italian regulatory agency for drugs
- BAU/mL:
-
Binding Arbitrary Units per mL
- COVID-19:
-
Coronavirus disease 2019
- ECLIA:
-
Electrochemiluminescence Immunoassay
- FC:
-
Fold Change
- GMT:
-
Geometric Mean Titer
- HPV:
-
Human papilloma virus
- HEV:
-
Hepatitis E Virus
- ICU:
-
Intensive Care Units
- IR:
-
Interquartile Range
- RBD:
-
Receptor-Binding Domain
- S:
-
Trimeric spike glycoprotein
- SARS-CoV-2:
-
Severe Acute Respiratory Syndrome caused by Coronavirus 2
- SD:
-
Standard Deviation
- Tx:
-
Time Points
- T0:
-
Baseline pre-vaccination
- T1:
-
20 Days after the first dose
- T2:
-
8 Days after the second dose, distinguishing
- T2 I:
-
8 Days after the second dose
- T2 II:
-
1 Month after the second dose
- T3:
-
3 Months after the second dose
- T6:
-
6 Months after the second dose
- T9:
-
9 Months after the second dose
- T boost:
-
8 Days after the administration of a further boost dose
- T0:
-
Baseline pre-vaccination after SARS-CoV-2 infection
- T1:
-
8 Days after the unique dose
- T2:
-
1 Month after the unique dose
- T3:
-
3 Months after the unique dose
- T6:
-
6 Months after the unique dose
- T9:
-
9 Months after the unique dose
- T boost:
-
8 Days after the administration of a further boost dose
- VLP:
-
Virus-like particles
References
Zhou F, Yu T, Du R, Fan G, Liu Y, Liu Z, et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: a retrospective cohort study. Lancet. 2020;395(10229):1054–62. https://doi.org/10.1016/S0140-6736(20)30566-3.
Hangartner L, Zinkernagel RM, Hengartner H. Antiviral antibody responses: the two extremes of a wide spectrum. Nat Rev Immunol. 2006;6(3):231–43. https://doi.org/10.1038/nri1783.
Antia A, Ahmed H, Handel A, Carlson NE, Amanna IJ, Antia R, et al. Heterogeneity and longevity of antibody memory to viruses and vaccines. PLoS Biol. 2018;16(8): e2006601. https://doi.org/10.1371/journal.pbio.2006601.
Smatti MK, Al Thani AA, Yassine HM. Viral-induced enhanced disease illness. Front Microbiol. 2018;9:2991. https://doi.org/10.3389/fmicb.2018.02991.
Eidem S, Tete SM, Jul-Larsen Å, Hoschler K, Montomoli E, Brokstad KA, et al. Persistence and avidity maturation of antibodies to A(H1N1) pdm09 in healthcare workers following repeated annual vaccinations. Vaccine. 2015;33(33):4146–54. https://doi.org/10.1016/j.vaccine.2015.05.081.
Lee YC, Kelly DF, Yu LM, Slack MP, Booy R, Heath PT, et al. Haemophilus influenzae type b vaccine failure in children is associated with inadequate production of high-quality antibody. Clin Infect Dis. 2008;46(2):186–92. https://doi.org/10.1086/524668.
Del Giudice G, Rappuoli R, Didierlaurent AM. Correlates of adjuvanticity: a review on adjuvants in licensed vaccines. Vaccine. 2015;33(33):4146–54. https://doi.org/10.1016/j.vaccine.2015.05.081.
Vogel AB, Kanevsky I, Che Y, Swanson KA, Muik A, Vormehr M, et al. BNT162b vaccines protect rhesus macaques from SARS-CoV-2. Nature. 2021;592(7853):283–9. https://doi.org/10.1038/s41586-021-03275-y.
Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020;579(7798):270–3. https://doi.org/10.1038/s41586-020-2012-7.
Filia A, Rota MR, D’Ancona FP. Comirnaty (BNT162b2), Il primo vaccino contro il COVID-19 approvato in Europa e in Italia: Istituto Superiore di Sanità -Dipartimentomalattieinfettive; 2021. https://www.epicentro.iss.it/vaccini/covid-19-vaccino-pfizer-biontech
Cavalcanti E, Isgrò MA, Rea D, Di Capua L, Trillò G, Russo L, et al. Vaccination strategy and anti - SARS-CoV-2 S titers in healthcare workers of the INT – IRCCS “Fondazione Pascale” Cancer Center (Naples, Italy). Infect Agent Cancer. 2021;16(1):32. https://doi.org/10.1186/s13027-021-00375-2.
Dan JM, Mateus J, Kato Y, Hastie KM, Yu ED, Faliti CE, et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science. 2021;371(6529):eabf4063. https://doi.org/10.1126/science.abf4063.
Krammer F, Srivastava K, Alshammary H, Amoako AA, Awawda MH, Beach KF, et al. Antibody responses in seropositive persons after a single dose of SARS-CoV-2 mRNA vaccine. N Engl J Med. 2021;384(14):1372–4. https://doi.org/10.1056/NEJMc2101667.
Padoan A, Dall’Olmo L, Rocca FD, Barbaro F, Cosma C, Basso D, et al. Antibody response to first and second dose of BNT162b2 in a cohort of characterized healthcare workers. Clin Chim Acta. 2021;519:60–3. https://doi.org/10.1016/j.cca.2021.04.006.
Blain H, Tuaillon E, Gamon L, Pisoni A, Miot S, Picot MC, et al. Spike antibody levels of nursing home residents with or without prior COVID-19 3 weeks after a single BNT162b2 vaccine dose. JAMA. 2021;325(18):1898–9. https://doi.org/10.1001/jama.2021.6042.
Buonaguro FM, Botti G, Ascierto PA, Pignata S, Ionna F, Delrio P, et al. The clinical and translational research activities at the INT – IRCCS “Fondazione Pascale” cancer center (Naples, Italy) during the COVID-19 pandemic. Infect Agents Cancer. 2020;15:69. https://doi.org/10.1186/s13027-020-00330-7.
Zaccaro DJ, Wagener DK, Whisnant CC, Staats HF. Evaluation of vaccine-induced antibody responses: impact of new technologies. Vaccine. 2013;31(25):2756–61. https://doi.org/10.1016/j.vaccine.2013.03.065.
Levi R, Azzolini E, Pozzi C, Ubaldi L, Lagioia M, Mantovani A, et al. One dose of SARS-CoV-2 vaccine exponentially increases antibodies in individuals who have recovered from symptomatic COVID-19. J Clin Investig. 2021;131(12): e149154. https://doi.org/10.1172/JCI149154.
Pang NY, Pang AS, Chow VT, Wang DY. Understanding neutralising antibodies against SARS-CoV-2 and their implications in clinical practice. Military Med Res. 2021;8:47. https://doi.org/10.1186/s40779-021-00342-3.
Wang L, Zhou T, Zhang Y, Yang ES, Schramm CA, Shi W, et al. Ultrapotent antibodies against diverse and highly transmissible SARS-CoV-2 variants. Science. 2021;373(6556):eabh1766. https://doi.org/10.1126/science.abh1766.
Budroni S, Buricchi F, Cavallone A, Bourguignon P, Caubet M, Dewar V, et al. Antibody avidity, persistence, and response to antigen recall: comparison of vaccine adjuvants. NPJ Vaccines. 2021;6(1):78. https://doi.org/10.1038/s41541-021-00337-0.
Tretyn A, Szczepanek J, Skorupa M, Jarkiewicz-Tretyn J, Sandomierz D, Dejewska J, et al. Differences in the concentration of anti-SARS-CoV-2 IgG antibodies post-COVID-19 recovery or post-vaccination. Cells. 2021;10(8):1952. https://doi.org/10.3390/cells10081952.
Levin EG, Lustig Y, Cohen C, Fluss R, Indenbaum V, Amit S, et al. Waning immune humoral response to BNT162b2 Covid-19 vaccine over 6 months. N Engl J Med. 2021;385(24):e84. https://doi.org/10.1056/NEJMoa2114583.
Harper DM, DeMars LR. HPV vaccines—a review of the first decade. Gynecol Oncol. 2017;146(1):196–204. https://doi.org/10.1016/j.ygyno.2017.04.004.
Wen GP, He L, Tang ZM, Wang SL, Zhang X, Chen YZ, et al. Quantitative evaluation of protective antibody response induced by hepatitis E vaccine in humans. Nat Commun. 2020;11:3971. https://doi.org/10.1038/s41467-020-17737-w.
Scheiblauer H, Nübling CM, Wolf T, Khodamoradi Y, Bellinghausen C, Sonntagbauer M, et al. Antibody response to SARS-CoV-2 for more than one year—kinetics and persistence of detection are predominantly determined by avidity progression and test design. J Clin Virol. 2022;146: 105052. https://doi.org/10.1016/j.jcv.2021.105052.
Tsang JS, Dobaño C, VanDamme P, Moncunill G, Marchant A, Othman RB, et al. Improving vaccine-induced immunity: can baseline predict outcome? Trends Immunol. 2020;41(6):457–65. https://doi.org/10.1016/j.it.2020.04.001.
Jogalekar MP, Veerabathini A, Gangadaran P. SARS-CoV-2 variants: A double-edged sword? Exp Biol Med (Maywood). 2021;246(15):1721–6. https://doi.org/10.1177/15353702211014146.
Schiøler H, Knudsen T, Brøndum RF, Stoustrup J, Bøgsted M. Mathematical modelling of SARS-CoV-2 variant outbreaks reveals their probability of extinction. Sci Rep. 2021;11(1):24498. https://doi.org/10.1038/s41598-021-04108-8.
Chaudhry MZ, Eschke K, Hoffmann M, Grashoff M, Abassi L, Kim Y, et al. Rapid SARS-CoV-2 adaptation to available cellular proteases. J Virol. 2022. https://doi.org/10.1128/jvi.02186-21.
Gerdol M, Dishnica K, Giorgetti A. Emergence of a recurrent insertion in the N-terminal domain of the SARS-CoV-2 spike glycoprotein. Virus Res. 2022;310: 198674. https://doi.org/10.1016/j.virusres.2022.198674.
Acknowledgements
The authors thank all the health workers of the INT – IRCCS “Fondazione Pascale” involved in the COVID monitoring and those of the Laboratory Medicine for their valuable contribution.
[Lab Medicine Unit: Domenica Rea, Lucia Di Capua, Francesco Labonia, Serena Meola, Annamaria Piscopo, Sergio Arpino, Carmine Di Napoli, Gerardo Esposito, Vincenzo Pane, Valentina Delle Donne; Molecular Biology and Viral Oncology Unit: Noemi Starita, Andrea Cerasuolo; Innovative Immunological Models Lab: Mariella Tagliamonte; Epidemiology Unit: Egidio Celentano, Anna Crispo, Concetta Montagnese, Giuseppe Porciello, Emanuela Rotondo, Roberto Simioli, Maria Grimaldi, Flavia Nocerino; Clinical Director Office: Paola Murino, Stefania D’Auria; Data Processing Center: Rocco Saviano]
Funding
Ricerca Corrente Ministero Salute 2021 – Progetto “Caratterizzazione Bio-Molecolare Del Virus SARS-CoV-2 E Dei Cofattori Dell’infiammazione Implicati Nella Patogenesi Della Covid-19” programma anti COVID-19 della Regione Campania Anno 2020 dal titolo “Misure Urgenti in materia di contenimento e gestione della emergenza epidemiologica da COVID-19 DRG N.140/2020”.
Author information
Authors and Affiliations
Consortia
Contributions
EC and FMB conceived, designed and supervised the project. NN, LM and AAMB supported the internal health surveillance program. GT, ALT, LB and MLT assisted with data collection. LR performed the tests. MAI performed the statistical analysis. EC, MAI, LR and FMB analysed, interpreted the data and wrote the manuscript draft. All authors reviewed and edited the manuscript. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The Institutional Ethics Committee approved the study, performed under the statements of Declaration of Helsinki, document #34/21 “RUI-SARS” (16/06/21).
Consent for publication
All authors gave their consent for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Isgrò, M.A., Trillò, G., Russo, L. et al. Bimodal antibody-titer decline following BNT162b2 mRNA anti-SARS-CoV-2 vaccination in healthcare workers of the INT – IRCCS “Fondazione Pascale” Cancer Center (Naples, Italy). Infect Agents Cancer 17, 40 (2022). https://doi.org/10.1186/s13027-022-00451-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13027-022-00451-1