
Abstract
Background
Falciparum and endemic Burkitt lymphoma (eBL) are co-endemic in Africa, but the malaria experience in eBL patients is unknown. A lower prevalence of falciparum has been reported in eBL patients, but those results are anecdotally attributed to pre-enrollment anti-malaria treatment.
Methods
We studied 677 eBL patients and 2920 community controls aged 0–15 years enrolled in six regions in Uganda, Tanzania, and Kenya during 2010–2016. Falciparum was diagnosed using thick blood film microscopy (TFM) and antigen-capture rapid diagnostic tests (RDTs). Guardians of the children answered a 40-item structured questionnaire about their child’s pre-enrollment lifetime malaria history and treatment, demographics, socioeconomics, animal exposures, fevers, and hospitalizations. We utilized exploratory factor analysis to reduce the 40 questionnaire variables into six factors, including Inpatient malaria and Outpatient malaria factors that were surrogates of pre-enrollment anti-malaria treatment. The six factors accounted for 83–90% of the variance in the questionnaire data. We calculated odds ratios and 95% confidence intervals (OR 95% CI) of association of eBL with falciparum positivity, defined as positive both on TFM or RDTs, or only RDTs (indicative of recent infection) or TFM (indicative of current falciparum infection) versus no infection, using multivariable logistic regression, controlling for group of age (0–2, 3–5, 6–8, 9–11 and 12–15 years), sex, and study site and the afore-mentioned pre-enrollment factors.
Results
The prevalence of falciparum infection was 25.6% in the eBL cases and 45.7% in community controls (aOR = 0.43, 95% CI: 0.40, 0.47; P < 0.0001). The results were similar for recent falciparum infection (6.9% versus 13.5%, aOR = 0.44, 95% CI: 0.38, 0.50; P < 0.0001) and current falciparum infection (18.7% versus 32.1%, aOR = 0.47, 95% CI: 0.43, 0.51; P < 0.0001). These aORs for any, recent and current falciparum infection did not change when we adjusted for pre-enrollment factors (aORs = 0.46, =0.44, and = 0.51, respectively) were significantly lower in stratified analysis for any infection in children < 5 years (aOR = 0.46; 95% CI: 0.29, 0.75) or ≥ 10 years (aOR = 0.47; 95% CI: 0.32, 0.71).
Conclusion
Our study results reduce support for pre-enrollment antimalaria treatment as a sole explanation for the observed lower falciparum prevalence in eBL cases and open a space to consider alternative immunology-based hypotheses.
Similar content being viewed by others


Introduction
Endemic Burkitt lymphoma (eBL) is an aggressive germinal-center (GC) B-cell non-Hodgkin lymphoma [1] first described in African children by Denis Burkitt in 1958 [2]. Epidemiological studies noted a geographical correlation of Plasmodium falciparum with eBL, which opened the hypothesis about whether this correlation was evidence of causation [3]. This hypothesis has been supported by findings that eBL patients are more likely than controls to have evidence of prior malaria, as measured by higher anti-malaria antibody titers in plasma [4,5,6], and less likely to be protected from malaria, as measured by carriage of genetic variants that protect against malaria [7, 8]. A causal role is also supported by immunological data. Recurrent falciparum infection stimulates proliferation of polyclonal B cells in the GC, where BL originates, and activates the MYC oncogene in B cells by promoting chromosomal translocations of MYC on chromosome 8q24 into the vicinity of immunoglobulin genes [9, 10]. Furthermore, falciparum preferentially stimulates B cells infected with Epstein-Barr virus via the cysteine-rich inter-domain region 1 alpha (CIDR1alpha) of the Plasmodium falciparum membrane protein 1 (PfEMP1) [11, 12].
However, as falciparum infection is associated with high malaria morbidity and mortality in children below age 3 years who lack immunity [13,14,15], it is unclear where eBL falls in spectrum of immunity to malaria. Malaria morbidity is due to blockage of vessels due to parasite sequestration and inflammatory response and loss of red blood cells (rbcs) [16]. Acquired immunity blocks sequestration, inflammation, and parasite infection of rbcs and promotes splenic clearance of parasites [17], although people remain susceptible and suffer frequent asymptomatic microscopic or submicroscopic infection [18]. The observation that the incidence of eBL peaks in children aged 5–9 years [19] suggests that eBL occurs in children who may have adequate protective immunity [20], but this possibility has not been fully tested [21].
Two studies reported a lower prevalence of falciparum in eBL patients than controls in Uganda [22] and Kenya [23]. This unexpected result was attributed to pre-enrollment anti-malaria treatment, although this hypothesis was not tested because pre-enrollment data were not collected. However, we recently observed this unexpected result (odds ratio [OR] of RDT positivity = 0.26, p < 0.0001) in our study of 697 eBL cases and 2934 community controls enrolled in the Epidemiology of Burkitt lymphoma in East African children and minors (EMBLEM) study in Uganda, Tanzania, and Kenya [24]. These results were not explained by malaria-related fevers reported up to 6 months pre-enrollment, which were lower in eBL cases than controls (OR = 0.47, p < 0.0001) or by other variables that captured pre-enrollment anti-malaria treatment. These results challenged the hypothesis that the lower prevalence of falciparum infection is due to pre-enrollment treatment of malaria, while also raising the possibility that falciparum prevalence is low in children with eBL because they have robust acquired resistance to falciparum.
In the current study, we conducted a more comprehensive adjustment of pre-enrollment exposures by using exploratory factor analysis (EFA) to reduce 40 questionnaire variables about the child’s pre-enrollment lifetime malaria history and treatment, demographics, socioeconomics, animal exposures, fevers, and hospitalizations into a few informative factors [25] and used those obtained factors to control for pre-enrollment anti-malaria treatment. EFA was suitable because it allowed us to adjust for all information without the risk of overfitting the data [26].
Methods
Population and setting
The study population has been described previously [24]. Briefly, eBL cases aged 0–15 years and community controls of a similar age, sex, and geographical area were enrolled in six regions in Uganda, Tanzania, and Kenya [24]. Both eBL cases and controls were tested for falciparum infection using thick blood film microscopy (TFM) and commercial antigen rapid diagnostic tests (RDTs [24]. Pre-enrollment malaria history and treatment, demographics, socioeconomics, animal exposures, fevers, and hospitalizations was elicited by questionnaire (https://emblem.cancer.gov/resources/EMBLEM_Interview_Questionnaire.pdf) (Supplementary Table 1).
Statistical methods -- imputation, exploratory factor analysis, association analyses
The 40 data elements analyzed in this study are shown in Supplementary Table 1. Because complete data are required for EFA, missing data were imputed using sequential regression imputation with 5 imputed datasets created in IVEware (http://www.isr.umich.edu/src/smp/ive) in SAS 9.4 [27]. Imputation affected < 1% of subjects and < 10% of individual data elements where data were missed/not collected. The imputations were performed for controls and cases separately using combined and country specific datasets. The imputation coefficients were combined using Rubin’s rules in Proc MIANALYZE in SAS [28].
EFA was performed on the complete combined and country-specific datasets (Supplementary Table 2) and yielded six factors that we extracted using the principal component method and according to Kaiser’s rule (eigenvalue > 1) [29]. The six factors were transformed using the varimax rotation method to obtain orthogonal (uncorrelated) factors. To facilitate presentation and interpretation of the results, the extracted factors were named according to the questionnaire variables based on the highest absolute factor loadings contributing to the factor and colors used to improve visualization (as shown in Fig. 1). The factor with high (red, positive value) loading from number of rooms in the house and bed net (ownership and use) was named Socioeconomic status; the factor with high (red) loading from keeping animals inside or near the house (all except pigs and birds) was named the Animals factor; the factor with high (red) loading from the inpatient malaria treatment and hospital admission was named the Inpatient malaria factor; the factor with high (red) loading from outpatient malaria treatment and fever (within 6 or 12 months of study enrollment) was named the Outpatient malaria factor; the factor with high (red) loadings from fevers not due to malaria was named the Non-specific fevers factor. Non-specific fevers may be due to undiagnosed viral or parasitic infections [30] or due to fevers of undetermined origin in patients with cancer, also known as B-symptoms [31]. The factor with high (red) loading for birth order (number of children) and inverse (dark green) loading for maternal and paternal education and occupation was named the Home factor.

Heatmap showing factors obtained from exploratory factor analysis (EFA). The rows are questionnaire variables and the columns are the factors. The questionnaire variables are grouped into 7 domains, and the columns are the six factors that were extracted. The factors are presented country-specifically A-C in the following order: Uganda, Tanzania, and Kenya, and D for all countries combined (primary analysis). For each factor, the results are based on five imputed datasets (see Supplementary Table 2). For simplicity, the factor loadings are color coded, with significant factor loadings that have a positive value coded red and pink, while those with a significant negative value are coded light green, and dark green, and those with significant but comparatively small absolute factor loadings are coded yellow. Non-significant factor loadings are shaded gray. Domains contributing to each of the six factors are outlined with a thick black box. The scale bar shows the correlation between variables and the factor loadings
We calculated crude and adjusted ORs and 95% confidence intervals (CIs) to estimate the association of eBL with falciparum using multivariable logistic regression. We adjusted for group of age (0–2, 3–5, 6–8, 9–11 and 12–15 years), sex, and study site, which were considered apriori as confounders. Falciparum infection was defined two ways: First, as a two-level variable with no infection, defined as negative on both TFM and RDT (coded 0), or infected, defined as positive either on TFM or RDT (coded 1). Second, as a three-level variable with no infection defined as before; recent infection defined as positive on RDT but negative on TFM (coded 1), or current infection defined as positive on TFM regardless of RDT result (coded 2; Supplementary Table 1). These definitions allowed us to classify individuals as having any infection (current or recent; definition 1) and distinguish them from those with recent infection, eg, asmay be observed in children who have been treated for malaria but still have degraded falciparum antigens in their blood for 3–4 weeks after treatment of malaria [32] and from those with current infection in whom blood-stage parasite forms are visualized. If the previously reported inverse association of falciparum with eBL is due to pre-enrollment anti-malaria treatment, we would expect to observe an inverse association between eBL and current falciparum infection but a positive association betwen eBL and recent falciparum infection. Additionally, we would expect that adjustment for pre-enrollment anti-malaria treatment would attenuate or eliminate the association of eBL with measures of falciparum infection. We report the combined results primarily and country-specific results secondarily to explore the consistency of effects across the three countries.
Results
Characteristics of included children
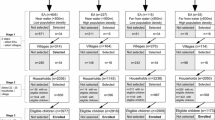
We analyzed 3631 children (692 eBL cases) enrolled in Uganda, Tanzania, and Kenya during 2010–2016 (Fig. 2). Of these, 34 children (15 eBL patients) were excluded because they were missing all questionnaire data. The remaining 3597 children (677 eBL cases; 62% with pathology confirmation) were included in the EFA. Table 1 shows the characteristics of the eBL cases and community controls analyzed. Overall, 55.3% were male, 12.2% had recent falciparum infection and 29.6% had current falciparum infection. Uganda contributed most (41%), followed by Kenya (33%), and the least number was from Tanzania (26%).

Flowchart showing the study process of selection of subjects, data checking, imputation of missing data, and case-control analysis
Association of the factors with falciparum among community controls
Table 2 shows, for all study sites combined, the univariate and adjusted associations with falciparum infection with the factors among the controls. Higher scores on the Socioeconomic status and Inpatient malaria factors were inversely associated with falciparum positivity. The association of falciparum infection with the Socioeconomic factor suggested a monotonic trend (aOR was 0.79 for Q2 vs. Q1, declining to aOR 0.45 for Q4 vs. Q1; P < 0.0001), whereas the association of falciparum infection with the Inpatient malaria factor was more consistent with a threshold (vs Q1 aORs were 0.75 for Q2, 0.79 for Q3, and 0.71 for Q4). Higher scores on the Animals, Non-specific fevers, and Home factors were associated with falciparum positivity, with a pattern suggesting monotonic trend for the Animals factor (aORs was 1.28 for Q2 rising to 1.69 for Q4 vs. Q1; P < 0.0001) and a threshold for the Non-specific fevers (aORs were 1.29 to 1.40 for Q2 to Q4 vs. Q1) and the Home factors (aORs were 1.14 to 1.38 for Q2 to Q4 vs. Q1). The Outpatient factor was unrelated to falciparum positivity (P = 0.20). Although there were slight variations, the associations observed in the combined analyses were present in country-specific analyses (Supplementary Table 3).
Association of the factors with eBL
All the factors were associated with increased odds of eBL in crude and adjusted analyses that included the factors (Table 2). Although falciparum positivity was inversely related to the Socioeconomic status and Inpatient malaria factors, these two facors were associated with higher odds of eBL (Table 2, Fig. 3). However, the associations of eBL with the Animals, Non-specific fevers, and Home factors were consistent, ie, in the same direction as the associations of those factors with falciparum positivity, while the Outpatient malaria factor, which was unrelated to falciparum positivity, was associated with higher odds of eBL (Fig. 3).

Heat map showing direction of associations with P < 0.05 between extracted factors and malaria infection in the controls and with eBL in the case-control analysis for both the combined and country-specific analysis, which are separated by dark column bars. The green color indicates an inverse association (lower risk), red color indicates a positive association (increased risk), while no coloring indicates a null association. For example, the SES factor was inversely associated with falciparum infection, but positively associated with eBL risk
Association of falciparum positivity with eBL diagnosis
We observed lower falciparum positivity in eBL cases than community controls (25.6% versus 45.7%, P < 0.0001) resulting in lower odds of eBL (aOR = 0.43, 95% CI: 0.40, 0.47, Table 2) in those with infection, in analyses adjusting for group of age, sex, and study site. The associations did not change when we included the aforementioned factors in the logistic regression model to control for pre-enrollment anti-malaria treatment (aOR = 0.46, 95% CI: 0.37, 0.56). The inverse association remained robust (aOR = 0.45, 95% CI 0.35, 0.59, P < 0.0001) when we further adjusted for mean platelet count, hemoglobin, and WBC count as confounders [33] and in stratified analysis for children < 5 years (aOR = 0.46; 95% CI: 0.29, 0.75) and those ≥10 years (aOR = 0.47; 95% CI: 0.32, 0.71), and was significant in country-specific analyses (aOR = 0.54 in Uganda, aOR = 0.48 in Tanzania and aOR = 0.30 in Kenya; Supplementary Table 4).
The adjusted results were similar for recent falciparum infection (aOR = 0.44, 95% CI 0.29, 0.65, P < 0.0001) or current falciparum infection (aOR = 0.51, 95% CI 0.39, 0.66, P < 0.0001).
Discussion
Our study found significantly lower prevalence of falciparum positivity, defined as any, recent, or current falciparum infection, in eBL patients than controls in crude and adjusted analyses. The ORs did not change when we adjusted for Inpatient malaria and Outpatient malaria factors as surrogates of pre-enrollment anti-malaria treatment, for the other four factors obtained from EFA, for platelets, low hemoglobin, and elevated white cell counts, which are risk factors for eBL [33]. The results were significant in stratified analyses by age (< 5 years versus ≥10 years or older), which is a surrogate for age-related acquired immunity against malaria. These results are similar to those reported in two studies that did not control for pre-enrollment anti-malaria treatment [23, 34] and those in our previous analyses that included a limited adjustment for a few variables about pre-enrollment anti-malaria treatment [24, 33]. Our findings, based on the most comprehensive adjustment of pre-enrollment exposures and for other risk factors of eBL and stratified analyses, reduce support for the null hypothesis that pre-enrollment anti-malaria treatment is the major explanation for the observed low falciparum positivity among eBL patients, on one hand, and increase support for exploring alternative hypotheses, on the other hand.
One alternative hypothesis, that is usually dismissed because it is counter-intuitive, is that lower prevalence of falciparum infection in eBL children is a surrogate of robust immunity to malaria. Immune response to falciparum spans a spectrum from lack of immunity, which is associated with severe malaria to strong age-related acquired anti-malaria and anti-parasite immunity [13], which is associated with mild or asymptomatic falciparum infection. We suggest from our results that eBL cases possess strong anti-malaria and anti-parasite immunity. Lack of immunity represents a serious risk of death from severe malaria in areas with high eBL incidence, thus, immunity against severe malaria usually develops quickly, in some estimates occurs after exposure to 1–2 falciparum infections and is usually well-developed in the first 1–3 years of life in malaria endemic countries [13]. However, immunity against severe malaria does not prevent falciparum infection or the development of mild/moderate malaria, which are also associated with serious life-threatening morbidity and mortality from anemia, stunting, malnutrition, bacterial co-infections [35], and seizures [36]. The risk for these complications reduces progressively with acquisition of age-related anti-malaria and anti-parasite immunity at a rate of 6% per year of age and 2% per episode of clinical malaria in some areas [14]. Despite having well-developed anti-malaria and anti-parasite immunity by age 5 years in high malaria burden areas, low-grade mild, asymptomatic microscopic or submicroscopic infections continue to afflict children [20]. Because eBL, which by definition occurs in children afflicted by heavier malaria at a median age of 5–9 years, we infer from this pattern that children predisposed to eBL have strong anti-parasite immunity before and after disease onset. This inference is intuitively consistent with the prediction that children with eBL would be expected to have capacity to control falciparum infection to survive in those areas, and it is supported by our finding that eBL cases have significantly lower parasite density than controls of a similar age [20].
The inferences above also suggest that there are some important differences between malaria versus viral carcinogenesis [37, 38]. In the latter, inability to control viral infection promotes inflammation and malignancy [39] and viral infections can be grouped into high-risk versus low-risk types depending on their association with cancer risk, e.g. cervical cancer and high-risk human papillomavirus types (eg, 16 or 18) [40]. This model does not apply to falciparum because lack of immunity to malaria would predispose to severe malaria and high risk of death before eBL [41]. Thus, children who develop immunity to severe malaria relatively quickly and acquire immunity to resist new infections [13] as a central strategy to thrive in hostile environments with heavy exposure to falciparum parasites [42]. Our studies in Ghana and Uganda have confirmed that eBL children as well as similarly aged controls have high antibody titers against falciparum variants linked to severe malaria, indicating prior exposure to and immune response to such variants [6]. Although the development of immunity is hierarchical, is correlated with age-dependent antibodies, and progressively limits parasitemia [43], it does not prevent infection and development of low-grade asymptomatic or sub-microscopic falciparum parasitemia [44]. Children with immunity to malaria have a lower risk of death from severe malaria [16, 18], but they are exposed to chronic falciparum parasitemia and increased activity by splenic macrophages to clear parasites [17]. We suggest that this increased falciparum antigenic load to the spleen is a risk factor for eBL [45] in children in malaria endemic regions. We further infer that it is not high-risk falciparum variants (as is the case for severe malaria), but the multiplicity of diverse variants the children are exposed to that are the risk factor for eBL [46]. Falciparum parasites can establish new infections in immune populations only by allelic switching of falciparum multigene families, such as PfEMP1 [47], to express new epitopes that are new to a particular host and, therefore, not recognized by the immunity acquired by that host up to that time. Our recent studies are consistent with this notion that infection with multiple (swarms of) falciparum variants, presumably expressing novel epitopes that are not recognized by the hosts' acquired immunity, may be related to eBL [19, 48, 49].
Our results highlight potential value of establishing collaboration between malaria and eBL research to jointly address overlapping interests. A better appreciation of the link between eBL and exposure to high falciparum burden could encourage better coordination between eBL and malariologists. eBL may be a suitable sentinel condition to detect pockets of high localized community falciparum transmission, which can be identified and targeted for investigation to suppress reservoirs that may threaten malaria prevention and intervention programs [20]. In view of our findings that eBL is associated with multiplicity of falciparum infection [19, 48, 49] and recent studies suggesting that markers of falciparum genotypic complexity are better measures of force of falciparum infection, co-transmission and superinfection events [50], close collaboration could facilitate the discovery of reliable markers of falciparum complexity that could also be used as an intermediate phenotype for eBL.
The strengths of our study include having a large sample size, data from three countries, detailed data about anti-malaria treatment before study enrollment, and measurement of falciparum using two methods. Our use of EFA is novel in eBL-malaria research, which made it possible for us to rigorously control for confounding without overfitting the data. The use of two tests for falciparum allowed us to test associations of eBL with recent anti-malaria treatment, which would have detected a positive association of recent falciparum with eBL, presumably because such children may have unreported pre-enrollment anti-malaria treatment. We acknowledge that the case-control design is a weakness of our study because it is susceptible to reverse causality that cannot be fully addressed. Nonetheless, our results substantially increase our understanding about the relationship between eBL and falciparum infection, the role of pre-enrollment exposures, and offer support for considering novel hypotheses.
In conclusion, we show that falciparum infection was significantly lower in eBL cases than controls in crude and adjusted analyses. The results were robust in analyses controlling for pre-enrollment factors and in analyses stratified by age. These results reduce support for pre-enrollment antimalaria treatment as a sole explanation for the observed lower falciparum prevalence in eBL cases, and open up space to consider alternative hypotheses such as the state of immunity to falciparum infection in children before and after eBL onset.
Availability of data and materials
Data and code used in the current analysis will be made available upon request from the corresponding author.
Abbreviations
- CIDR1alpha:
-
Cysteine-rich inter-domain region 1 alpha
- EMBLEM:
-
Epidemiology of Burkitt lymphoma in East African children and minors
- EFA:
-
Exploratory factor analysis
- eBL:
-
endemic Burkitt lymphoma
- OR:
-
Odds ratio
- 95% CI:
-
95% confidence interval
- RDTs:
-
Rapid diagnostic tests
- PfEMP1:
-
Plasmodium falciparum membrane protein 1
- Q:
-
Quartile
- TFM:
-
Thick film microscopy
References
Leoncini L, Raphael M, Stein H, Harris NL, Jaffe ES, Kluin PM. Burkitt lymphoma. 4th ed. Lyon: International Agency for Research on Cancer (IARC); 2008.
Burkitt D. A sarcoma involving the jaws in African children. Brit J Surg. 1958;46(197):218–23.
Burkitt DP. Etiology of Burkitts Lymphoma - an Alternative Hypothesis to a Vectored Virus. J Nat Cancer Inst. 1969;42(1):19.
Carpenter LM, Newton R, Casabonne D, Ziegler J, Mbulaiteye S, Mbidde E, et al. Antibodies against malaria and Epstein-Barr virus in childhood Burkitt lymphoma: a case-control study in Uganda. Int J Cancer. 2008;122(6):1319–23.
Aka P, Vila MC, Jariwala A, Nkrumah F, Emmanuel B, Yagi M, et al. Endemic Burkitt lymphoma is associated with strength and diversity of Plasmodium falciparum malaria stage-specific antigen antibody response. Blood. 2013;122(5):629–35.
Derkach A, Otim I, Pfeiffer RM, Onabajo OO, Legason ID, Nabalende H, et al. Associations between IgG reactivity to Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) antigens and Burkitt lymphoma in Ghana and Uganda case-control studies. EBioMedicine. 2019;39:358–68.
Legason ID, Pfeiffer RM, Udquim KI, Bergen AW, Gouveia MH, Kirimunda S, et al. Evaluating the causal link between malaria infection and endemic Burkitt lymphoma in northern Uganda: a Mendelian randomization study. Ebiomedicine. 2017;25:58–65.
Williams AO. Haemoglobin genotypes, ABO blood groups, and Burkitt's tumour. J Med Genet. 1966;3(3):177–9.
Robbiani DF, Deroubaix S, Feldhahn N, Oliveira TY, Callen E, Wang Q, et al. Plasmodium infection promotes genomic instability and AID-dependent B cell lymphoma. Cell. 2015;162(4):727–37.
Torgbor C, Awuah P, Deitsch K, Kalantari P, Duca KA, Thorley-Lawson DA. A multifactorial role for P. falciparum malaria in endemic Burkitt’s lymphoma pathogenesis. PLoS Pathogens. 2014;10(5):e1004170.
Chene A, Donati D, Guerreiro-Cacais AO, Levitsky V, Chen Q, Falk KI, et al. A molecular link between malaria and Epstein-Barr virus reactivation. PLoS Pathog. 2007;3(6):e80.
Lam KM, Syed N, Whittle H, Crawford DH. Circulating Epstein-Barr virus-carrying B cells in acute malaria. Lancet. 1991;337(8746):876–8.
Gupta S, Snow RW, Donnelly CA, Marsh K, Newbold C. Immunity to non-cerebral severe malaria is acquired after one or two infections. Nat Med. 1999;5(3):340–3.
Rodriguez-Barraquer I, Arinaitwe E, Jagannathan P, Boyle MJ, Tappero J, Muhindo M, et al. Quantifying heterogeneous malaria exposure and clinical protection in a cohort of Ugandan children. J Infect Dis. 2016;214(7):1072–80.
Chizzolini C, Delaporte E, Kaufmann MH, Akue JP, Verdini AS, Pessi A, et al. Age-related prevalence of antibody response against three different, defined Plasmodium falciparum antigens in children from the Haut-Ogooue province in Gabon. Trans R Soc Trop Med Hyg. 1989;83(2):147–51.
Chizzolini C, Grau GE, Geinoz A, Schrijvers D. T lymphocyte interferon-gamma production induced by Plasmodium falciparum antigen is high in recently infected non-immune and low in immune subjects. Clin Exp Immunol. 1990;79(1):95–9.
Engwerda CR, Beattie L, Amante FH. The importance of the spleen in malaria. Trends Parasitol. 2005;21(2):75–80.
Engwerda CR, Good MF. Interactions between malaria parasites and the host immune system. Curr Opin Immunol. 2005;17(4):381–7.
Emmanuel B, Kawira E, Ogwang MD, Wabinga H, Magatti J, Nkrumah F, et al. African Burkitt lymphoma: age-specific risk and correlations with malaria biomarkers. Am J Trop Med Hyg. 2011;84(3):397–401.
Redmond LS, Ogwang MD, Kerchan P, Reynolds SJ, Tenge CN, Were PA, et al. Endemic Burkitt lymphoma: a complication of asymptomatic malaria in sub-Saharan Africa based on published literature and primary data from Uganda, Tanzania, and Kenya. Malar J. 2020;19(1):239.
Bouvard V, Baan RA, Grosse Y, Lauby-Secretan B, El Ghissassi F, Benbrahim-Tallaa L, et al. Carcinogenicity of malaria and of some polyomaviruses. Lancet Oncol. 2012;13(4):339–40.
De-The G, Geser A, Day NE, Tukei PM, Williams EH, Beri DP, et al. Epidemiological evidence for causal relationship between Epstein-Barr virus and Burkitt’s lymphoma from Ugandan prospective study. Nature. 1978;274(5673):756–61.
Asito AS, Piriou E, Odada PS, Fiore N, Middeldorp JM, Long C, et al. Elevated anti-Zta IgG levels and EBV viral load are associated with site of tumor presentation in endemic Burkitt’s lymphoma patients: a case control study. Infect Agents Cancer. 2010;5:13.
Peprah S, Ogwang MD, Kerchan P, Reynolds SJ, Tenge CN, Were PA, et al. Risk factors for Burkitt lymphoma in east African children and minors: a case-control study in malaria-endemic regions in Uganda, Tanzania and Kenya. Int J Cancer. 2020;146(4):953–69.
Fonseca MJ, Costa P, Lencastre L, Tavares F. A statistical approach to quantitative data validation focused on the assessment of students’ perceptions about biotechnology. Springerplus. 2013;2:496.
Vittinghoff E, McCulloch CE. Relaxing the rule of ten events per variable in logistic and cox regression. Am J Epidemiol. 2007;165(6):710–8.
Raghunathan TE, Lepkowski JM, Van-Hoewyk M, Solenberger PW. A multivariate technique for multiply imputing missing values using a sequence of regression models. Survey Methodol. 2001;27:10.
Yuan Y. Multiple imputation using SAS software. J Stat Softw. 2011;45(6):1–25.
Kaiser HF. The application of electronic computers to factor analysis. Educ Psychol Meas. 1960;20(1):141–51.
Rooth I, Bjorkman A. Fever episodes in a holoendemic malaria area of Tanzania: parasitological and clinical findings and diagnostic aspects related to malaria. Trans R Soc Trop Med Hyg. 1992;86(5):479–82.
Horowitz HW. Fever of unknown origin or fever of too many origins? N Engl J Med. 2013;368(3):197–9.
Grandesso F, Nabasumba C, Nyehangane D, Page AL, Bastard M, De Smet M, et al. Performance and time to become negative after treatment of three malaria rapid diagnostic tests in low and high malaria transmission settings. Malar J. 2016;15(1):496.
Peprah S, Ogwang MD, Kerchan P, Reynolds SJ, Tenge CN, Were PA, et al. Mean platelet counts are relatively decreased with malaria but relatively increased with endemic Burkitt lymphoma in Uganda, Tanzania, and Kenya. Br J Haematol. 2020;190(5):772–82.
De-The G. Is Burkitt’s lymphoma related to perinatal infection by Epstein-Barr virus? Lancet. 1977;1(8007):335–8.
Scott JA, Berkley JA, Mwangi I, Ochola L, Uyoga S, Macharia A, et al. Relation between falciparum malaria and bacteraemia in Kenyan children: a population-based, case-control study and a longitudinal study. Lancet. 2011;378(9799):1316–23.
Idro R, Ndiritu M, Ogutu B, Mithwani S, Maitland K, Berkley J, et al. Burden, features, and outcome of neurological involvement in acute falciparum malaria in Kenyan children. JAMA. 2007;297(20):2232–40.
Evans AS, Mueller NE. Viruses and cancer. Causal associations. Ann Epidemiol. 1990;1(1):71–92.
Mueller N, Evans A, Harris NL, Comstock GW, Jellum E, Magnus K, et al. Hodgkin's disease and Epstein-Barr virus. Altered antibody pattern before diagnosis. N Engl J Med. 1989;320(11):689–95.
Schiffman M, Herrero R, Desalle R, Hildesheim A, Wacholder S, Rodriguez AC, et al. The carcinogenicity of human papillomavirus types reflects viral evolution. Virology. 2005;337(1):76–84.
Schiffman M, Herrero R, Hildesheim A, Sherman ME, Bratti M, Wacholder S, et al. HPV DNA testing in cervical cancer screening: results from women in a high-risk province of Costa Rica. JAMA. 2000;283(1):87–93.
WHO. World Malaria Report 2018. Geneva: WHO; 2018.
Kafuko GW, Burkitt DP. Burkitts Lymphoma and Malaria. Int J Cancer. 1970;6(1):1.
Cham GK, Turner L, Kurtis JD, Mutabingwa T, Fried M, Jensen AT, et al. Hierarchical, domain type-specific acquisition of antibodies to Plasmodium falciparum erythrocyte membrane protein 1 in Tanzanian children. Infect Immun. 2010;78(11):4653–9.
Goncalves BP, Kapulu MC, Sawa P, Guelbeogo WM, Tiono AB, Grignard L, et al. Examining the human infectious reservoir for Plasmodium falciparum malaria in areas of differing transmission intensity. Nat Commun. 2017;8(1):1133.
Agerbaek MO, Pereira MA, Clausen TM, Pehrson C, Oo HZ, Spliid C, et al. Burkitt lymphoma expresses oncofetal chondroitin sulfate without being a reservoir for placental malaria sequestration. Int J Cancer. 2017;140(7):1597–608.
Eldh M, Hammar U, Arnot D, Beck HP, Garcia A, Liljander A, et al. Multiplicity of asymptomatic Plasmodium falciparum infections and risk of clinical malaria: a systematic review and pooled analysis of individual participant data. J Infect Dis. 2020;221(5):775–85.
Saito F, Hirayasu K, Satoh T, Wang CW, Lusingu J, Arimori T, et al. Immune evasion of Plasmodium falciparum by RIFIN via inhibitory receptors. Nature. 2017;552(7683):101–5.
Johnston WT, Mutalima N, Sun D, Emmanuel B, Bhatia K, Aka P, et al. Relationship between Plasmodium falciparum malaria prevalence, genetic diversity and endemic Burkitt lymphoma in Malawi. Sci Rep. 2014;4:3741.
Arisue N, Chagaluka G, Palacpac NMQ, Johnston WT, Mutalima N, Peprah S, et al. Assessment of mixed Plasmodium falciparumsera5 infection in endemic Burkitt lymphoma: a case-control study in Malawi. Cancers. 2021;13(7):1692.
Mueller I, Schoepflin S, Smith TA, Benton KL, Bretscher MT, Lin E, et al. Force of infection is key to understanding the epidemiology of Plasmodium falciparum malaria in Papua new Guinean children. Proc Natl Acad Sci U S A. 2012;109(25):10030–5.
Acknowledgements
We thank the study population and communities for their participation. We thank Ms. Janet Lawler-Heavener at Westat Inc., (Rockville, MD, USA) and Mr. Erisa Sunday at the African Field Epidemiology Network (Kampala, Uganda) for managing the study. We are grateful to the leadership of the collaborating countries and institutions for hosting local field offices and laboratories and supporting the fieldwork. We thank Ms. Laurie Buck, Dr. Carol Giffen, Mr. Greg Rydzak and Mr. Jeremy Lyman at Information Management Services Inc. (Calverton, MD, USA) for coordinating data, and preparing data analysis files.
Funding
This work was supported by funding from the Intramural Research Program of the Division of Cancer Epidemiology and Genetics, National Cancer Institute at the National Institutes of Health’ [Contracts HHSN261201100063C, HHSN261201100007I] plus by the Intramural Research Program, National Institute of Allergy and Infectious Diseases at the National Institutes of Health [support to SJR], Department of Health and Human Services. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the US Government. Open Access funding provided by the National Institutes of Health (NIH).
Author information
Authors and Affiliations
Contributions
SMM conceived the idea, designed the study and supervised the work. MDO, PK, SJR, CNT, RTK, WNW, PAW, NM, KB, LAW, RMP, and EK contributed to study design and supervised field work. IO, IDL, PAW, MDO, PK, SJR, CNT, RTK, WNW, PAW, NM, and EK conducted fieldwork. SP conducted statistical analysis and interpreted data. RMP supervised statistical analysis. SP drafted the manuscript; JJG, RMP and SMM critically edited the paper. All authors contributed to the manuscript, read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was performed in accordance with ethical guidelines and regulations consistent with the Declaration of Helsinki. Ethical approval was given by ethics committees at Uganda Virus Research Institute, Uganda National Council for Science and Technology (H816), Tanzania National Institute for Medical Research (NIMR/HQ/R.8c/Vol. IX/1023), Moi University/Moi Teaching and Referral Hospital (000536), and National Cancer Institute (10-C-N133). The guardians of the children (mostly parents) gave written informed consent. Children aged 7 years or older gave written marked assent.
Consent for publication
Not applicable because the manuscript does not include identifiable individual data.
Competing interests
SMM is co-Editor-in-Chief of Infectons Agents and Cancer, but was not resposible for handling this manuscript during review; all the other authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Table 1.
Questionnaire variables/data elements and how they were coded for exploratory factor analysis. Supplementary Table 2. Eigenvalue and variance explained by extracted factors shown for each imputed dataset and by study country. Supplementary Table 3. Odds ratios and 95% confidence intervals (CIs) of the association of factors with falciparum infection risk in controls, by country. Supplementary Table 4. Odds ratios and 95% confidence intervals (CIs) of the association of factors with eBL risk, by country.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Peprah, S., Ogwang, M.D., Kerchan, P. et al. Inverse association of falciparum positivity with endemic Burkitt lymphoma is robust in analyses adjusting for pre-enrollment malaria in the EMBLEM case-control study. Infect Agents Cancer 16, 40 (2021). https://doi.org/10.1186/s13027-021-00377-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13027-021-00377-0