
Abstract
Background
Gynostemma pentaphyllum (Thunb.) Makino, commonly known as “southern ginseng”, contains high amounts of ginsenoside derivatives and exhibits similar biological activities with Panax ginseng (C. A. MEY) (ginseng), which is usually used as a low-cost alternative to ginseng. G. pentaphyllum has therapeutic effects on liver diseases. However, the mechanisms underlying its hepatoprotective action have not been fully elucidated.
Methods
The protective effects of the ethanolic extract of G. pentaphyllum (GPE) were evaluated using an experimental carbon tetrachloride (CCl4)-induced liver disease model. Potential targets of GPE were predicted using the “Drug-Disease” bioinformatic analysis. Furthermore, comprehensive network pharmacology and transcriptomic approaches were employed to investigate the underlying mechanisms of GPE in the treatment of liver disease.
Results
The pathological examinations showed that GPE significantly alleviated hepatocyte necrosis and liver injury. GPE significantly downregulated Bax and cleaved-PARP expression and upregulated Bcl-2 expression during CCl4-induced hepatocyte apoptosis. We compared the effects of four typical compounds in GPE -a ginsenoside (Rb3) shared by both GPE and ginseng and three unique gypenosides in GPE. Notably, Gypenoside A (GPA), a unique saponin in GPE, markedly reduced hepatocyte apoptosis. In contrast, ginsenoside Rb3 had a weaker effect. Network pharmacology and transcriptomic analyses suggested that this anti-apoptotic effect was achieved by upregulating the PI3K/Akt signaling pathway mediated by PDK1.
Conclusions
These results suggested that G. pentaphyllum had a promising hepatoprotective effect, with its mechanism primarily involving the upregulation of the PDK1/Bcl-2 signaling pathway by GPA, thereby preventing cell apoptosis.
Graphic Abstract

Similar content being viewed by others


Introduction
Gynostemma pentaphyllum (Thunb.) Makino has a long history of use as a traditional medicine and dietary supplements in China. In 2002, the Chinese Ministry of Public Health of China recognized G. pentaphyllum as a functional food. Saponins are the primary bioactive components of G. pentaphyllum. They play a significant role in its diverse biological activities and clinical effects. G. pentaphyllum is a unique plant outside the family Araliaceae that contains ginseng saponins and at least eight ginsenosides identical to the saponins found in Panax ginseng (C. A. MEY) (ginseng). These shared compounds provide G. pentaphyllum with a tonic function similar to ginseng and for this reason, G. pentaphyllum is praised as “Southern ginseng” and widely used as a low-cost alternative to ginseng [1]. G. pentaphyllum has excellent research and development value.
Liver injury is a critical stage in the progression from chronic liver disease to cirrhosis and poses a serious threat to patient health [2]. The occurrence and development of liver injury are complex processes involving both liver parenchymal cells and non-parenchymal cells, including sinusoidal endothelial cells, hepatic stellate cells (HSCs), Kupffer cells, and different types of lymphocytes. Among them, HSCs are the primary source of collagen synthesis and secretion in the liver, and are commonly considered as effector cells of liver fibrosis. Kupffer cells are special macrophages participating in HSC activation and regulation alongside other lymphocyte types. Excessive and persistent hepatocyte apoptosis is a crucial hallmark of liver injury progression and serves as a trigger for liver injury and fibrosis [3, 4]. Therefore, targeting hepatocyte apoptosis is a critical approach for treating liver injury. Treatments and drugs that focus on targeting liver parenchymal cell damage while considering other hepatic cells are required to treat clinical liver diseases.
The phosphatidylinositol-3 kinase/protein kinase B (PI3K/Akt) pathway, is a key survival pathway that targets multiple molecules involved in anti-apoptosis, antioxidant defense, and protein synthesis [5]. Phosphoinositide dependent protein kinase-1 (PDK1), a serine/threonine kinase of the Protein Kinase A, G, and C family, is a fundamental component of the PI3K/PDK1/Akt pathway [6]. However, PDK1 expression and function in liver injury remains unexplored. Further exploration of PDK1 expression and function and its relationship with liver injury may provide new targets for treating liver diseases.
In vivo and in vitro studies have indicated that the ethanol extract of G. pentaphyllum (GPE) exhibits a range of favorable bioactivities, including antioxidant, anti-inflammatory, and antilipidemic effects [7], suggesting that GPE can regulate hepatic cellular homeostasis. This study investigated the protective effects of GPE in mice with carbon tetrachloride (CCl4)-induced liver injury. Network pharmacology and transcriptomics analyses revealed that GPE and Gypenoside A (GPA) upregulated the expression of proteins involved in the PI3K/PDK1/Akt signaling pathway, alleviating CCl4-induced liver injury and hepatocyte apoptosis. Therefore, G. pentaphyllum is expected to be a safe dietary supplement and medicinal material for treating liver injury.
Materials and methods
Chemicals and reagents
G. pentaphyllum leaves (Cat: 20230615) were acquired from Beijing Bencao Fangyuan Co., Ltd. (Beijing, China). CCl4 was obtained from Boer’s (Shanghai, China). Olive oil was purchased from Yuanye Biotechnology Co., Ltd. (Shanghai, China). Ginsenoside Rb3 (Cat: S9208) and Gypenoside XLIX (Cat: S9177) were purchased from Selleck (Shanghai, China). Gypenoside A (Cat: HY-N2440), Gypenoside XLVI (Cat: HY-N6252) and GSK2334470 (Cat: HY-14981) were purchased from MedChem Express (Shanghai, China). The aspartate transaminase (AST, Cat: BC1565), alanine transami-nase (ALT, Cat: BC1555) and hydroxyproline (HYP, Cat: BC0255) were purchased from Solarbio Life Sciences (Beijing, China).
Preparation of GPE
100 g leaves and 1 liter of 80% ethanol aqueous solution were mixed thoroughly and sonicated for 1 h, gfiltered, and the filtrate collected and stored in the refrigerator at 4 ℃. Next, the residue was added to 1L of 80% aqueous ethanol aqueous solution, and the procedure was repeated to collect the filtrate. The filtrates from the two filtration and vacuum drying rounds were combined to obtain crude GPE powder.
Animals
We obtained 30 male C57BL/6 J mice (16–18 g) from Beijing Vital River Laboratory Animal Technology Co., Ltd. (SCXK (JING) 2021–0011). The mice were kept under controlled environmental conditions, with a temperature of 25 ± 2 °C and a relative humidity of 50 ± 10%. A 12 h light/dark cycle was established, and all mice were provided water and standard chow throughout the study period. The mice were intraperitoneally with 5% CCl4 or an equivalent amount of olive oil daily for 6 weeks. The mice were injected once daily for the first 2 weeks and once every 2 d for the next 4 weeks. GPE was administered by gavage at a dose of 500 mg/kg/day from the beginning of the study for 6 weeks. An equal volume of 0.5% sodium carboxymethyl cellulose was administered to the control group. The Capital Medical University Animal Ethics Committee approved this study (Permit number: AEEI-2023-132).
Histopathological examination
Hepatic pathological changes were examined using hematoxylin and eosin (H&E), Masson’s trichrome, and Sirius red staining. Collagen deposition and fibrosis were evaluated using ImagePro Plus 6 software (Media Cybernetics, Inc.). Hepatocyte apoptosis was assessed by terminal deoxynucleotidyl transferase-mediated dUTP Nick-End Labeling (TUNEL) staining (Servicebio, Wuhan, China).
Network pharmacology and bioinformatics-based analysis
G. pentaphyllum compounds were sourced from the TCMSP database (http://lsp.nwsuaf.edu.cn/tcmsp.php) and screened based on criteria of oral bioavailability critrria ≥ 30% and drug-likeness ≥ 0.18. G. pentaphyllum potential target proteins were predicted using SwissTargetPrediction (http://www.swisstargetprediction.ch/) Using the Keyword “Liver Injury” we identified liver injury-related targets from the Online Mendelian Inheritance in Man database (http://www.omim.org/) and Genecards (http://www.genecards.org), a comprehensive database that provides information on genes, their products, and biomedical applications.
We constructed an interaction network for G. pentaphyllum active components and putative targets in treating liver injury using their interaction data. The interaction network was visualized using Cytoscape software (Version 3.9.0). We analyzed the degree, betweenness, and closeness centrality, using the network Analyzer plugin in Cytoscape. Thus, providing insights into the topological significance of nodes within the network. In addition, a protein-protein interaction network was established to explore the interplay between these targets using Cytoscape (version 3.9.0). Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis was conducted using the Database for Annotation, Visualization, and Integrated Discovery (https://david.ncifcrf.gov/). Molecular docking was conducted to screen for proteins exhibiting superior binding affinities for GPA using the SYBYL-X 2.0 software.
Transcriptomic analysis
Total RNA was isolated from primary mouse hepatocytes using Trizol reagent after treatment with DMSO, GPE, or GPA in the presence of 8 mM CCl4 for 24 h. Subsequently, cDNA libraries were generated following the manufacturer’s protocols, including the use of the NEBNext® Ultra™ Directional RNA Library Prep Kit, NEBNext® Poly (A) mRNA Magnetic Isolation Module, and NEBNext® Multiplex Oligos (New England Biolabs, Ipswich, MA, USA). Differentially expressed genes (DEGs) were identified based on the criteria of |log2FC|> 0.585 and a p-value < 0.05, which indicated statistical significance.
Cell culture-primary hepatocyte isolation
Primary hepatocytes were isolated using the standard 2-step perfusion technique [8]. The liver was perfused with 50 mL of HBSS buffer through the inferior vena cava at a controlled flow rate of 5 mL/min. Subsequently, a 25 mL solution containing 0.5 mg/mL collagenase was perfused at a controlled flow rate of 3 mL/min. Following perfusion, the liver was extracted from the abdominal cavity, and the hepatocytes were liberated into DMEM using sterile surgical scissors. The resulting cell suspension underwent filtration through a 70 μm cell strainer. After centrifugation, hepatocytes were subjected to low-speed gradient centrifugation (50 g, 3 min). The viability of the isolated hepatocytes reached approximately 90%, as determined by Trypan blue staining. Finally, the hepatocytes were seeded into 6-well plates and cultured in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin.
Measurement of AST, ALT and HYP
AST, ALT, and HYP serum levels of the mice were quantified using commercial kits according to the manufacturer’s protocol (Solarbio Life Sciences). For primary mouse hepatocytes, supernatants were collected post-treatment, and AST and ALT enzymatic activities were determined according to the manufacturer’s protocols (Solarbio Life Sciences).
Western blotting assay
Liver and hepatocyte proteins were extracted using RIPA buffer supplemented with protease inhibitor and centrifuged at 12,000 g for 15 min. The protein concentration was determined using the BCA method. Subsequently, the protein samples were mixed with a loading buffer, denatured in boiling water for 5 min, and subjected to electrophoretic separation on sodium dodecyl sulfate-polyacrylamide gels. The separated proteins were then transferred to PVDF membranes. Following the transfer, the membranes were incubated overnight at 4 °C with primary antibodies, washed, and then incubated with secondary antibodies (Table S1). Finally, the target protein bands were visualized using a Tanon imaging device and chemiluminescence kit.
Statistical analysis
All values presented in the figures and text are depicted as mean ± standard error of the mean (SEM). Statistical analyses were conducted using a one-way ANOVA, followed by the Bonferroni post hoc test for multiple comparisons. P-value < 0.05 was considered significant. *p < 0.05, **p < 0.0001: versus indicated group.
Results
GPE improved CCl 4 -induced liver injury in mice
The CCl4-induced chronic liver fibrosis model is a widely accepted experimental model for studying liver injury and fibrosis [9]. In the organism, CCl4 initiates lipid peroxidation through hepatic activation, resulting in hepatocyte degeneration, necrosis and liver fibrosis formation [10]. To confirm the GPE curative effect on liver injury, we first observed its effects in a long-term low-dose CCl4-induced liver fibrosis model mice (Fig. 1A). Despite having no significant influence on ratio of liver weight to body weight, GPE reduced the liver weight gained in CCl4-induced mice (Fig. 1B). Long-term CCl4 use resulted in severe liver injury. In contrast, oral administration of GPE reduced the elevated levels of serum AST and ALT (Fig. 1C). Compared with the control group, GPE reversed the CCl4-induced impairment of liver surface smoothness (Fig. 1D). H&E staining showed that GPE reduced inflammatory cell infiltration and relieved hepatic cell edema. Masson’s and Sirius red staining further confirmed that GPE reduced hepatic collagen deposition. Consistently, GPE administration reduced HYP levels in circulating blood (Fig. 1C). Besides, we examined the levels of collagen 1A1(COL1A1) and alpha smooth muscle (α-SMA) in the liver and the protein levels of COL1A1 and α-SMA were significantly decreased in the GPE treatment group (Fig. 1E). These results demonstrated that GPE plays a protective role against liver injury and fibrosis.

GPE ameliorated CCl4-induced liver injury in mice. Mice were injected intraperitoneally with 5% carbon tetrachloride (CCl4) or an equivalent amount of olive oil daily for 6 weeks. The mice were injected once daily for the first 2 weeks and then once every 2 d for the next 4 weeks. The ethanol extract of G. pentaphyllum (GPE) was administered by gavage at a dose of 500 mg/kg/day from the beginning of modeling for a total of 6 weeks. An equal volume of 0.5% sodium carboxymethylcellulose was administered to the control group. A Workflow of mouse experiments. B Ratio of liver weight to body weight and liver weight in various groups (n = 6). C Serum alanine aminotransferase (ALT), aspartate aminotransferase (AST), and hydroxyproline (HYP) (n = 6). D Hematoxylin and eosin (H&E) and Masson and Sirius Red staining of liver tissues (n = 3). Scale bar: 50 μm. (E) Protein expression of collagen 1A1(COL1A1) and alpha smooth muscle (α-SMA) in the liver (n = 6). * p < 0.05, ** p < 0.001: versus indicated group
Network pharmacology predicted potential signaling pathways in the inhibitory effect of GPE on liver injury
Next, a network pharmacology analysis was conducted to understand the mechanism by which GPE inhibits CCl4-induced liver injury (Fig. S1A). Initially, 296 potential targets of G. pentaphyllum and 3416 potential targets of liver injury were identified using online databases. From this, 177 overlapping targets were selected as G. pentaphyllum-regulated liver injury targets (Fig. S1B), and 16 compounds associated with these targets were deemed effective (Table S2). Subsequently, a compound-target network was established, and related functional enrichment analysis was performed (Fig. 2A). KEGG enrichment analysis indicated a close association of these targets with various metabolic signaling pathways, especially the apoptosis signaling pathway (Fig. 2D). Furthermore, a PPI network of the 177 targets was constructed using the Cytoscape software, revealing 33 hub targets (Fig. 2B, C), with AKT1 exhibiting the highest degree value. These findings suggested that GPE therapeutic effect on liver fibrosis might be mediated by regulating apoptotic signaling pathways and AKT1 plays an essential role in this process.

Network pharmacology predicted potential signaling pathways in G. pentaphyllum inhibitory effect on liver injury. G. pentaphyllum targets in liver injury was predicted using network pharmacological analysis. A The compound-target network was established using Cytoscape 3.9.0 software. The yellow hexagon represents G. pentaphyllum, the olive-green circles represent the bioactive compounds of G. pentaphyllum, and the bottle-green diamonds represent G. pentaphyllum-regulated liver injury targets. B, C Hub targets were identified from the protein-protein interaction (PPI) network using Cytoscape 3.9.0 software. D Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis of 177 G. pentaphyllum-regulated liver injury targets using Metascape
GPE inhibited CCl4-induced hepatocyte apoptosis in vivo
Hepatocyte apoptosis is involved in the occurrence and development of various of acute and chronic liver diseases. It is an essential mechanism of hepatocyte injury [11]. This study investigated whether GPE is beneficial against liver injury by inhibiting hepatocyte apoptosis. We investigated GPE anti-apoptosis activity in mouse hepatocytes using TUNEL staining. The result showed that a large number of TUNEL-positive cells appeared in the CCl4-induced group, indicating that apoptosis occurred. GPE reversed the increase in TUNEL-positive cells induced by CCl4 and reduced hepatocyte apoptosis (Fig. 3A). Cleaved-PARP, cleaved caspase-3, and cleaved caspase-7 are typical features of apoptosis. Western blotting showed that the protein expression of Cleaved-PARP, Cleaved-caspase 3, and Cleaved-caspase 7 increased in the CCl4-induced group and was decreased when GPE treatment was administered simultaneously (Fig. 3B), further verifying GPE beneficial actions on hepatocyte apoptosis. Bcl-2 and Bax protein are two essential proteins that regulate apoptosis, and the reduced the Bcl-2/ Bax ratio strongly confirmed the role of GPE in hepatocyte apoptosis (Fig. 3C). These results demonstrated that GPE inhibits CCl4-induced hepatocyte apoptosis in vivo.

GPE inhibited CCl4-induced hepatocyte apoptosis. Mice were intraperitoneally injected with 5% carbon tetrachloride (CCl4) or an equivalent amount of olive oil daily for 6 weeks. The mice were injected once daily for the first 2 weeks and then once every 2 d for the next 4 weeks. The ethanol extract of G. pentaphyllum (GPE) was administered by gavage at a dose of 500 mg/kg/day from the beginning of modeling for a total of 6 weeks. An equal volume of 0.5% sodium carboxymethyl cellulose was administered to the control group. A Terminal deoxynucleotidyl transferase dUTP nick-end labeling (TUNEL) staining of the liver tissue (n = 3). Scale bar: 50 μm. B Protein expression levels of Cleaved-PARP, Cleaved-caspase 3, and cleaved-caspase 7 in the livers and hepatocytes isolated from the livers (n = 6). C Protein expression levels of Bcl-2/Bax in the liver and hepatocytes isolated from the liver (n = 6). * p < 0.05, ** p < 0.001: versus indicated group
GPE and GPA prevented CCl4-induced hepatocyte apoptosis in vitro
We studied the GPE anti-apoptotic effect and the main components of GPE anti-apoptotic mouse hepatocytes. GPA, GP-XLIX and GP-XLVI were the main triterpenoids isolated from GPE, whereas Rb3 is one of the dominant active components in G. pentaphyllum and ginseng [12]. Fig S2A shows the structures of these compounds. We investigated the anti-apoptotic effects of GPE, GPA, GP-XLIX, GP-XLVI, and Rb3 in a CCl4-induced hepatocyte apoptosis model. ALT and AST levels in the cell culture medium showed that GPE and GPA alleviated hepatocyte damage dose-dependently when exposed to CCl4 (Fig. 4A, B). The reduced expression of cleaved-PARP, cleaved caspase-3, and cleaved caspase-7 further confirmed the anti-apoptotic effects of GPE and GPA (Fig. 4C, D). GPE and GPA consistently decreased the Bcl-2/Bax ratio in CCl4-induced hepatocytes (Fig. 4E, F). However, GP-XLIX and GP-XLVI had no significant effect on the anti-apoptotic phenotype (Fig. S2A). Rb3 showed a tendency to alleviate apoptosis, but its effects were weaker than those of GPA (Fig. S2B). These results indicated that GPE and its unique active component, GPA, exhibits remarkable hepatoprotective and anti-apoptotic effects in vitro. And GPA might as well be the main active component contributing to the anti-apoptotic effect of GPE.

GPE and Gypenoside A (GPA) prevented CCl4-induced hepatocyte apoptosis in vitro. Primary mouse hepatocytes were treated with DMSO or the ethanol extract of G. pentaphyllum (GPE) or Gypenoside A (GPA) in the presence of 8 mM carbon tetrachloride (CCl4) for 24 h. A, B The levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in the hepatocytes culture-medium (n = 6). C, D The protein expression levels of Bcl-2/Bax in the hepatocytes (n = 6). E, F The protein expression levels of PARP/Cleaved-PARP, Cleaved-caspase 3 and Cleaved-caspase 7 in the hepatocytes (n = 6). * p < 0.05, ** p < 0.001: versus indicated group
Transcriptomics analysis revealed the signaling pathways involved in the inhibitory effect of GPE and GPA on liver injury
We explored how GPE and GPA reduce apoptosis by performing hepatocyte transcriptomics approach which clarified the potential mechanisms. Unsupervised hierarchical clustering and principal component analysis (PCA) showed that the GPE and GPA groups were close to the control group and far from the CCl4 group, suggesting that the gene tended to be normal after the treatment (Fig. 5A). The volcano map showed that compared to the CCl4 group, GPE treatment up-regulated 1326 DEGs and down-regulated 1683 DEGs, whereas GPA treatment up-regulated 570 DEGs and down-regulated 116 DEGs (Fig. 5B, C). The Venn diagram showed that GPE treatment reversed the expression of 582 DEGs, whereas GPA treatment reversed the expression of 482 DEGs induced by CCl4 (Fig. S3B). These 582 DEGs and 482 DEGs displayed in the heatmaps were considered as the therapeutic targets of GPE and GPA in CCl4-induced liver injury (Fig. 5D, E).

Transcriptomics analysis revealed the signaling pathways involved in the inhibitory effect of GPE and GPA on liver injury. Primary mouse hepatocytes were treated with DMSO, an ethanolic extract of G. pentaphyllum (GPE), or Gypenoside A (GPA) in the presence of 8 mM carbon tetrachloride (CCl4) for 24 h. mRNA was extracted for transcriptomic analysis. A Unsupervised hierarchical clustering and principal component analysis (PCA) diagram (n = 3). B Volcano map showing the number of DEGs in the CCl4 group versus those in the GPE group (n = 3). C Volcano map showing the number of DEGs in the GPA and CCl4 group (n = 3). D, E Heatmap of DEGs in the model group reversed by GPE and GPA administration. Blue indicates downregulated genes, and red indicates upregulated genes (n = 3). F, G KEGG enrichment analysis of 582 and 482 therapeutic targets of GPE and GPA, respectively
KEGG pathway analysis showed that GPE and GPA therapeutic targets of were mainly enriched in PI3K/Akt and MAPK signaling pathways (Fig. 5F). This was consistent with the results of the network pharmacology analysis, which suggested that PI3K/Akt signaling pathway plays a vital role in GPE and GPA in liver injury (Fig. 5F, G). These results implied that GPE and GPA relieved CCl4-induced liver injury mainly by inhibiting apoptosis through PI3K/Akt signaling pathway regulation.
GPE and GPA inhibited liver injury by controlling the PI3K/PDK1/Akt signaling pathway in vivo and in vitro
We measured the protein expression of key molecules in the PI3K/PDK1/Akt in CCl4-significantly inhibited induced mice and hepatocytes to confirm whether GPE and GPA treatment affected the pathway. When CCl4 significantly inhibited PI3K/PDK1/Akt signaling, GPE treatment restored PI3K/PDK1/Akt signaling by enhancing the phosphorylation of PI3K, PDK1, and Akt (Fig. 6A). Consistent with in vivo, GPE and GPA promoted the expression of p-PI3K, p-PDK1, and p-Akt in the CCl4-induced hepatocyte apoptosis model (Fig. 6B). Furthermore, the effect of promoting phosphorylation was dose-dependent. These results demonstrated the inhibitory effects of GPE and GPA on liver injury associated with the PI3K/PDK1/Akt signaling pathway.

GPE and GPA inhibited liver injury by controlling the PI3K/PDK1/Akt signaling pathway in vivo and in vitro. Mice were intraperitoneally injected with 5% carbon tetrachloride (CCl4) or an equivalent amount of olive oil daily for 6 weeks. The mice were injected once daily for the first 2 weeks and then once every 2 d for the next 4 weeks. The ethanol extract of G. pentaphyllum (GPE) was administered by gavage at a dose of 500 mg/kg/day from the beginning of modeling for a total of 6 weeks. An equal volume of 0.5% CMC-Na was administered to the control group. A Representative western blots for p-PI3K, PI3K, p-PDK1, PDK1, p-Akt, and Akt protein expression in the liver or hepatocytes isolated from the livers of different groups (n = 6). Primary mouse hepatocytes were treated with DMSO, GPE, or GPA in the presence of 8 mM (CCl4) for 24 h. B Representative western blots showing p-PI3K, PI3K, p-PDK1, PDK1, p-Akt, and Akt protein expression (n = 6). C Interaction between the Gypenoside (GPA) and PDK1, as determined by molecular docking. The green dotted line represents the hydrogen bond and interaction between GPA and AKT1 determined by molecular docking. The green dotted line represents the hydrogen bonds. * p < 0.05, ** p < 0.001: versus indicated group
We performed a docking analysis to further analyze the direct relationship between GPA and the PI3K/PDK1/Akt signaling pathway. The results of silicon molecular docking showed that GPA had a strong binding ability with Akt and PDK1 (total score: 10.1398 and 4.0415, respectively) but could not bind to PI3K (Fig. 6C). Studies have shown that PDK1 plays an important role in regulating apoptosis via the PI3K/Akt pathway [13]. Therefore, we hypothesized that PDK1 is a potential target protein for GPE and GPA to regulate the PI3K/Akt signaling pathway and improve liver injury.
PDK1 was required for the effects of GPE and GPA on PI3K/Akt signaling pathway activation and hepatocyte apoptosis
Silicon docking analysis provided essential clues that GPA mediated PI3K/Akt signaling pathway might interact directly with PDK1. Studies have shown that PDK1 can independently activate the PI3K/Akt signaling pathway. The molecular ablation of PDK1 function is associated with the deactivation of the PI3K/Akt signaling pathway [14]. We used GSK2334470 (1 μM), a novel and highly specific inhibitor of PDK1 [15], to verify the importance of PDK1 to GPE and GPA in CCl4-induced hepatocyte apoptosis model. Elevated ALT and AST levels in the cell culture medium showed that GSK2334470 treatment reversed the inhibitory effect of GPE and GPA on cell damage in hepatocytes when exposed to CCl4 (Fig. 7A, B). In the present study, GPE and GPA promoted the phosphorylation of PDK1, PI3K, and Akt; however, those effect were abrogated by GSK2334470 treatment (Fig. 7C, D). The results indicated that GSK2334470 inhibited the phosphorylation expression of PDK1, thereby suppressing PI3K and Akt phosphorylation. Concordantly, GSK2334470 treatment attenuated the enhanced effects of GPE and GPA on the anti-apoptotic protein Bcl-2 expression (Fig. 7C, D). These results showed that GPE and GPA enhance the phosphorylation of PDK1, thereby stimulating the PI3K/Akt signaling pathway and subsequently increasing the expression of Bcl-2. This inhibits hepatocyte apoptosis and ameliorates liver injury. GPE and GPA regulated the PI3K/Akt signaling pathway and enhanced the expression of Bcl-2 in a PDK1-dependent manner.

The role of PDK1 inhibition on the modulation of GPE and GPA on hepatocyte apoptosis. Primary mouse hepatocytes were treated with DMSO, the ethanol extract of G. pentaphyllum (GPE), or Gypenoside A (GPA), and with or without the PDK1 inhibitor GSK2334470 in the presence of 8 mM carbon tetrachloride (CCl4) for 24 h. A, B Levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in the hepatocyte culture medium (n = 6). C, D Expression levels of p-PDK1, PDK1, PI3K, p-PI3K, p-Akt, Akt and Bcl-2 in hepatocytes (n = 6). * p < 0.05, ** p < 0.001: versus indicated group
Discussion
In chronic liver disease, hepatocyte injury can stimulate collagen accumulation and increase matrix stiffness by affecting the metabolic levels in the hepatic microenvironment [16]. Inhibiting hepatocyte apoptosis offers a promising therapeutic approach for liver injury. G. pentaphyllum, a perennial plant belonging to the family-Cucurbitaceae, is mainly cultivated in southern China, Japan, and Korea [17]. In this study, we demonstrated that GPE ameliorated liver injury and fibrosis by improving hepatocyte morphology and reducing fibrous accumulation, ultimately inhibiting the liver injury progression.
Liver injury is a pathological consequence of hepatocyte apoptosis. Oxidative stress, toxic injury, inflammation, hypoxia, and viral infections can induce hepatocyte damage, leading to apoptosis [18]. CCl4, a hepatorenal toxic agent, effectively simulates the physiological process from hepatocyte damage to apoptosis both in vivo and in vitro. Therefore, the use of CCl4-induced liver injury model and hepatocyte apoptosis models is a promising strategy for drug screening. Our results revealed that GPE reduced the number of TUNEL-positive cells, increased the Bcl-2/Bax ratio, and decreased hepatocyte apoptosis. The discovery of GPE inhibitory effect on hepatocyte apoptosis supplements the insufficient understanding of the GPE mechanism of action in regulating liver disease.
The major bioactive constituents of G. pentaphyllum are dammarane-type triterpene saponins known as gypenosides [19]. Approximately 180 gypenosides have been identified in G. pentaphyllum, including GPA, GP-XLIX, GP-XLVI, and Rb3. Interestingly, although far from ginseng botanically, G. pentaphyllum also contains ginsenosides including Rb3. The ginsenosides in G. pentaphyllum constitute approximately 25% of the total saponins in the plant. G. pentaphyllum is an essential and unique plant outside the family-Araliaceae (i.e., ginseng, notoginseng, and American ginseng) that contains ginsenosides. Therefore, G. pentaphyllum has been praised as “Southern ginseng” and “poor man’s ginseng” [17], which can be explained by the chemical resemblance of G. pentaphyllum saponins to those of Araliaceae family Panax species. Studies have shown that G. pentaphyllum has some of the benefits similar to those of ginseng. In this study, we found that GPA could prevent CCl4-induced hepatocyte apoptosis in vitro, GP-XLIX and GP-XLVI had no significant influence on the anti-apoptotic phenotype, and although Rb3 tended to alleviate apoptosis, its effects were weaker than those of GPA. Therefore, we believe that GPA is the primary active substance in G. pentaphyllum that exerts anti-apoptotic activity, which explains the distinction between the hepatoprotective effects of G. pentaphyllum and ginseng.
We conducted a transcriptomic analysis using bioinformatics to elucidate further the specific mechanism by which GPE and GPA exert anti-apoptotic effects on hepatocytes. KEGG enrichment analysis revealed that the anti-apoptotic mechanisms of GPE and GPA may be associated with the PI3K/Akt, MAPK, and Rap1 signaling pathways. Akt activation promotes phosphorylation of the Bcl-2/Bcl-XL-related death promoter (Bad), thereby inhibiting caspase-3-mediated cell death [20]. This phosphorylation event leads to the dissociation of Bcl-2 from phosphorylated Bad and promotes cell survival [21]. Research has demonstrated that PI3K/Akt pathway activation can elevate Bcl-2 protein levels, decrease Bax and caspase 3 proteins expression, confer protection against apoptosis, and ameliorate liver diseases [22, 23]. Given that our previous results confirmed that GPE can elevate the decrease in the Bcl-2/Bax ratio induced by CCl4 and considering its relationship with the PI3K/Akt pathway, we hypothesized that GPE and GPA might exert their anti-apoptotic effects through the PI3K/Akt signaling pathway. As expected, we found that GPE and GPA increased the protein levels of p-PI3K and p-Akt in the PI3K/Akt pathway, thereby enhancing the Bcl-2/Bax ratio.
PDK1 is a phosphorylation-regulated kinase expressed in various eukaryotic organs that plays a central role in activating diverse cell signaling pathways [24, 25]. PI3K activation generates phosphatidylinositol-3,4,5-triphosphate [26], and subsequently recruits PDK1 and Akt to the plasma membrane. Membrane-bound PDK1 then phosphorylates Akt at Thr308, thereby activating Akt and initiating the PI3K/Akt pathway, ultimately suppressing apoptosis and autophagy [27]. According to previous research, PDK1 serves as an independent activator of the PI3K/Akt signaling pathway, and silencing the expression of PDK1 affects the expression of the PI3K/Akt signaling pathway [14]. Consequently, we further investigated the critical role of PDK1 in GPE inhibition of cell apoptosis and improvement of liver injury. GPE treatment restored PI3K/PDK1/Akt signaling, which was inhibited by CCl4 by enhancing the phosphorylation of PI3K, PDK1, and Akt in vivo. GPE and GPA promoted the expression of p-PI3K, p-PDK1, and p-Akt in a CCl4-induced hepatocyte apoptosis model. Docking analysis showed that GPA had a strong binding ability to Akt and PDK1, but could not bind to PI3K. In the CCl4-induced hepatocyte apoptosis model, the addition of GSK2334470, a novel and highly specific PDK1 inhibitor, weakened the effects of GPE and GPA on hepatocyte apoptosis. This was achieved by deactivating PDK1 phosphorylation and inhibiting the phosphorylation of PI3K and Akt. These findings suggest that GPE and GPA enhance the activity of PDK1 by regulating its phosphorylation, which in turn activates the PI3K/Akt signaling pathway. These results indicated that GPE and GPA regulated the PI3K/Akt signaling pathway and enhanced the expression of Bcl-2 in a PDK1-dependent manner.
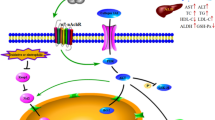
In conclusion, G. pentaphyllum exerts significant therapeutic effects against CCl4-induced liver injury and fibrosis by inhibiting hepatocyte apoptosis and controlling the PDK1-mediated PI3K/Akt signaling pathway to regulate Bcl-2 and caspase family expression (Fig. 8). Our findings contribute to a comprehensive understanding of the protective effects of GPE and GPA against liver injury and may provide a potential remedy for treating liver diseases in the future.

The ethanol extract of G. pentaphyllum (GPE) and its unique saponin Gypenoside A (GPA) significantly ameliorates CCl4-induced liver injury and fibrosis via inhibiting hepatocyte apoptosis activation by controlling PDK1-mediated PI3K/Akt signaling pathway to regulate the Bcl-2 and caspase families expression
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Abbreviations
- AST:
-
Aspartate aminotransferase
- ALT:
-
Alanine aminotransferase
- Akt:
-
Protein kinase B
- CCl4 :
-
Carbon tetrachloride
- COL1A1:
-
Collagen 1A1
- DEGs:
-
Differentially expressed genes
- Ginseng:
-
Panax ginseng (C. A. MEY)
- GPA:
-
Gypenoside A
- GPE:
-
Ethanol extract from Gynostemma pentaphyllum (thunb.) makino
- GP-XLIX:
-
Gypenoside XLIX
- GP-XLVI:
-
Gypenoside XLVI
- H&E:
-
Haematoxylin and eosin
- HYP:
-
Hydroxyproline
- HSCs:
-
Hepatic stellate cells
- KEGG:
-
Kyoto Encyclopedia of Genes And Genomes
- PCA:
-
Principal component analysis
- PDK1:
-
Phosphoinositide dependent protein kinase-1
- PI3K:
-
Phosphatidylinositol-3-kinase
- PPI:
-
Protein–protein interaction
- Rb3:
-
Ginsenoside Rb3
- α-SMA:
-
Alpha smooth muscle
- TCM:
-
Traditional Chinese medicine
- TUNEL:
-
Terminal Deoxynucleotidyl Transferase mediated dUTP Nick-End Labeling
References
Su C, Li N, Ren R, Wang Y, Su X, Lu F, et al. Progress in the medicinal value, bioactive compounds, and pharmacological activities of gynostemma pentaphyllum. Molecules. 2021;26(20).
Marrero JA, Kulik LM, Sirlin CB, Zhu AX, Finn RS, Abecassis MM, et al. Diagnosis, staging, and management of hepatocellular carcinoma: 2018 practice guidance by the American association for the study of liver diseases. Hepatology. 2018;68(2):723–50.
Malhi H, Guicciardi ME, Gores GJ. Hepatocyte death: a clear and present danger. Physiol Rev. 2010;90(3):1165–94.
Schuppan D, Kim YO. Evolving therapies for liver fibrosis. J Clin Investig. 2013;123(5):1887–901.
Miao Z, Miao Z, Wang S, Shi X, Xu S. Quercetin antagonizes imidacloprid-induced mitochondrial apoptosis through PTEN/PI3K/AKT in grass carp hepatocytes. Environ Pollut. 2021;290:118036.
Choi JH, Yang YR, Lee SK, Kim SH, Kim YH, Cha JY, et al. Potential inhibition of PDK1/Akt signaling by phenothiazines suppresses cancer cell proliferation and survival. Ann N Y Acad Sci. 2008;1138:393–403.
Wong WY, Lee MM, Chan BD, Ma VW, Zhang W, Yip TT, et al. Gynostemma pentaphyllum saponins attenuate inflammation in vitro and in vivo by inhibition of NF-κB and STAT3 signaling. Oncotarget. 2017;8(50):87401–14.
Hernández-Alvarez MI, Sebastián D, Vives S, Ivanova S, Bartoccioni P, Kakimoto P, et al. Deficient endoplasmic reticulum-mitochondrial phosphatidylserine transfer causes liver disease. Cell. 2019;177(4):881-895.e17.
Eguchi A, De Mollerat Du, Jeu X, Johnson CD, Nektaria A, Feldstein AE. Liver bid suppression for treatment of fibrosis associated with non-alcoholic steatohepatitis. J Hepatol. 2016;64(3):699–707.
Aram G, Potter JJ, Liu X, Wang L, Torbenson MS, Mezey E. Deficiency of nicotinamide adenine dinucleotide phosphate, reduced form oxidase enhances hepatocellular injury but attenuates fibrosis after chronic carbon tetrachloride administration. Hepatology. 2009;49(3):911–9.
Shao S, Zhang Y, Zhou F, Meng X, Yu Z, Li G, et al. LncRNA-Airn alleviates acute liver injury by inhibiting hepatocyte apoptosis via the NF-κB signaling pathway. Acta Biochim Biophys Sin. 2022;54(11):1619–29.
Ru W, Wang D, Xu Y, He X, Sun YE, Qian L, et al. Chemical constituents and bioactivities of panax ginseng (C. A. Mey.). Drug Discov Ther. 2015;9(1):23–32.
Meng F, Zhang Y, Li X, Wang J, Wang Z. Clinical significance of miR-138 in patients with malignant melanoma through targeting of PDK1 in the PI3K/AKT autophagy signaling pathway. Oncol Rep. 2017;38(3):1655–62.
Bamodu OA, Chang HL, Ong JR, Lee WH, Yeh CT, Tsai JT. Elevated PDK1 expression drives PI3K/AKT/MTOR signaling promotes radiation-resistant and dedifferentiated phenotype of hepatocellular carcinoma. Cells. 2020;9(3).
Najafov A, Sommer Eeva M, Axten Jeffrey M, Deyoung MP, Alessi DR. Characterization of GSK2334470, a novel and highly specific inhibitor of PDK1. Biochem J. 2010;433(2):357–69.
Zhang D, Zhang Y, Sun B. The molecular mechanisms of liver fibrosis and its potential therapy in application. Int J Mol Sci. 2022;433(2):357–69.
Nguyen NH, Ha TKQ, Yang JL, Pham HTT, Oh WK. Triterpenoids from the genus gynostemma: chemistry and pharmacological activities. J Ethnopharmacol. 2021;268: 113574.
Wang K. Molecular mechanisms of hepatic apoptosis. Cell Death Dis. 2014;5(1): e996.
Kim JH, Han YN. Dammarane-type saponins from gynostemma pentaphyllum. Phytochemistry. 2011;72(11–12):1453–9.
Fruman DA, Meyers RE, Cantley LC. Phosphoinositide kinases. Annu Rev Biochem. 1998;67:481–507.
Nakazato PCG, Victorino JP, Fina CF, Mendes KDS, Gomes MCJ, Evora PRB, et al. Liver ischemia and reperfusion injury pathophysiology and new horizons in preconditioning and therapy. Acta cirurgica brasileira. 2018;33(8):723–35.
Rong YP, Huang HT, Liu JS, Wei L. Protective effects of geniposide on hepatic ischemia/reperfusion injury. Transpl Proc. 2017;49(6):1455–60.
Sheng M, Zhou Y, Yu W, Weng Y, Xu R, Du H. Protective effect of berberine pretreatment in hepatic ischemia/reperfusion injury of rat. Transpl Proc. 2015;47(2):275–82.
Ericson K, Gan C, Cheong I, Rago C, Samuels Y, Velculescu VE, et al. Genetic inactivation of AKT1, AKT2, and PDPK1 in human colorectal cancer cells clarifies their roles in tumor growth regulation. Proc Natl Acad Sci USA. 2010;107(6):2598–603.
Mucha BE, Banka S, Ajeawung NF, Molidperee S, Chen GG, Koenig MK, et al. Correction: a new microdeletion syndrome involving TBC1D24, ATP6V0C, and PDPK1 causes epilepsy, microcephaly, and developmental delay. Genet Med. 2019;21(9):2159–60.
Obata A, Kimura T, Obata Y, Shimoda M, Kinoshita T, Kohara K, et al. Correction to: vascular endothelial PDPK1 plays a pivotal role in the maintenance of pancreatic beta cell mass and function in adult male mice. Diabetologia. 2019;62(12):2375.
Hu B, Zhang Y, Deng T, Gu J, Liu J, Yang H, et al. PDPK1 regulates autophagosome biogenesis by binding to PIK3C3. Autophagy. 2021;17(9):2166–83.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (82074112) and the Key Project at the central government level: the ability to establish the sustainable use of valuable Chinese medicine resources (2060302). The instruments and equipment used in this study were provided by the School of Traditional Chinese Medicine at Capital Medical University.
Author information
Authors and Affiliations
Contributions
Yawen Lu made substantial contributions to the conceptualization, supervision, and interpretation of the data, as well as to drafting the article. Linlan Hu performed the experiments, analyzed the data, and drafted the manuscript. Xin Zhao, Hanxiao Cheng, and Yafei Guo performed the experiments and collected data, while Xian He and Shaoting Chen collected the data. Jiabo Wang and Guangde Zhou contributed to the discussion and manuscript review. All authors approved the final version of the manuscript. Guangde Zhou, Jiabo Wang, and Yawen Lu were responsible for the integrity of the study.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The care and treatment of these mice were performed in accordance with the Provisions and General Recommendations of the Chinese Experimental Animals Administration Legislation and approved by the Animal Ethics Committee of the Capital Medical University (Permit number: AEEI-2023-132).
Competing interests
The authors do not have any disclosures to report.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hu, L., Zhao, X., He, X. et al. Gynostemma Pentaphyllum ameliorates CCl4-induced liver injury via PDK1/Bcl-2 pathway with comprehensive analysis of network pharmacology and transcriptomics. Chin Med 19, 70 (2024). https://doi.org/10.1186/s13020-024-00942-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13020-024-00942-w