
Abstract
The morbidity of lung cancer ranks first among all cancers. Lung adenocarcinoma (LUAD) is a classification of lung cancer, and cell invasion and migration of LUAD are the main causes for its high mortality. Therefore, further exploring the potential mechanism of LUAD metastasis may provide bases for following targeted drug development and treatment of LUAD. In this study, clinical data as well as gene expression profiles were obtained from TCGA-LUAD and GEO to analyze CTHRC1 expression. The result found that CTHRC1 was significantly high in LUAD. Similar results were also discovered in 4 cancer cell lines. Moreover, overexpressed/knock-down CTHRC1 cell lines were constructed. It was uncovered that overexpressing CTHRC1 promoted LUAD cell migration and invasion, and inhibited cell adhesion, while knocked down CTHRC1 had the opposite effect. Afterward, the upstream miRNAs that regulated CTHRC1 were predicted by several bioinformatics websites. It was testified by dual-luciferase method that CTHRC1 was negatively mediated by miR-30a-5p. Overexpressed miR-30a-5p suppressed cell invasion/migration, and increased cell adhesion, while overexpressing CTHRC1 as well reversed such impacts. In conclusion, it was disclosed in this study that CTHRC1 worked as a cancer promoter in LUAD, and miR-30a-5p could target and downregulate CTHRC1 to regulate cell adhesion, and inhibited LUAD cell invasion and migration. These results elucidated at cellular level that upregulated CTHRC1 may be a marker protein for LUAD metastasis.
Similar content being viewed by others


Introduction
Lung cancer is a recognized human health killer, with its mortality ranks first among all cancers for several years (about 11.6% in 2018) and its morbidity is relatively high [1]. After years of research, lung cancer is divided into many subtypes, one of which is lung adenocarcinoma (LUAD). Over 500,000 people died from lung cancer every year, and the amount has been significantly increasing over the past few decades [2,3,4]. Although great efforts have been devoted to developing new treatment of lung cancer in recent years, the prognosis of malignant patients remains poor with a 5-year survival rate less than 10% [5, 6]. Shortage of understanding of LUAD-related biological mechanism limits the improvement of therapeutic effect. Hence, it is important to dig related genes of LUAD occurrence and development, and explore its effective mechanism for increasing clinical efficacy.
CTHRC1 is a chondrocyte-secreted glycoprotein first found in the rat balloon-injured artery model and can inhibit collagen matrix synthesis [7, 8]. As revealed in recent years, CTHRC1 is upregulated in various tumors, and promote cancer cell invasion and migration [9,10,11], which also works as a potential biomarker of various cancers. For example, MEI ZHENG et al. [12] disclosed that CTHRC1 overexpression promotes cervical cancer development by simulating Wnt/PCP signaling pathway. Moreover, upregulated CTHRC1 promotes the invasion of epithelial ovarian cancer via stimulating EGFR signaling pathway. However, CTHRC1 high expression in LUAD might pertain to the angiogenesis of LUAD and indicate poor prognosis of LUAD [13]. But reasons for high expression of CTHRC1 and its regulatory mechanism in LUAD are not clear.
Effects of miRNAs in vivo have been neglected for a long time. However, recent study represented that miRNA mediates gene expression via targeting 3’-untranslated region (UTR) of mRNA [14], so as to modulate the progression of various diseases. Studies revealed that CTHRC1 is regulated by miRNAs in cancers. For example, miR-155 targets CTHRC1 to inhibit colorectal cancer [15]. MiR-30c-mediated CTHRC1 accelerates the metastasis of LUAD cells [16]. MiR-98 targets CTHRC1 to suppress liver cancer cell progression [17]. Nonetheless, there are no reports alike in LUAD. This work scrutinized CTHRC1 and LUAD occurrence and development, and uncovered the molecular mechanism of miRNA targeting CTHRC1, which offers researching directions for targeted treatment of LUAD.
Materials and methods
Bioinformatics method
LUAD related data sets were acquired from Gene Expression Omnibus (GEO) and The Cancer Genome Atlas (TCGA), as shown in Table 1. Expression differences of CTHRC1 between normal tissue and LUAD tissue were tested by t-test, and the effect of CTHRC1 expression on patient’s prognosis was detected with R “survival” package. Upstream miRNAs that regulated CTHRC1 were predicted with starBase, TargetScan, miRDB and mirDIP databases, and verified via Pearson correlation analysis. Pathway enrichment analysis was undertaken on CTHRC1 by using Gene Set Enrichment Analysis (GSEA) software.
Cell culture and transfection
LUAD cell lines H1650 (BNCC100260), Calu-3 (BNCC338514), A549 (BNCC337696), H1975 (BNCC100301), and human bronchial epithelial cell line BEAS-2B (BNCC338205) were bought from BeNa Culture Collection. All cells were kept in Roswell Park Memorial Institute (RPMI)-1640 (Thermo Fisher Scientific Company, Waltham, Massachusetts, USA) medium containing 5% fetal bovine serum (FBS), and cultured in an incubator under general conditions.
NC mimic, miR-30a-5p mimic (mimic) were offered by GenePharma (Shanghai, China). oe-CTHRC1 vector, 3 si-CTHRC1 vectors and their negative control lentivirus packing vectors were acquired from Invitrogen (Carlsbad, CA, USA). Vectors were transiently transfected into Calu-3 cells with Lipofectamine 2000 (Thermo Fisher Scientific, Inc.). All cells were cultivated for at least 24 h in the complete medium before transfection, and collected after 36–48 h of transfection.
qRT-PCR
Total RNA was separated with TRIzol Reagent (Invitrogen). MRNA was reversely transcribed into cDNA with M-MLV Reverse Transcriptase Kit (TaKaRa). MiRNA was reversely transcribed with Superscript II Kit (Invitrogen). PCR system was constructed with miScript SYBR Green PCR Kit (Qiagen, Hilden, Germany). Applied Biosystems 7300 Real-Time PCR System (Applied Biosystems, USA) was applied for qRT-PCR to detect gene expression level, with U6 and GAPDH as the internal references. Relative transcription level of the target gene was calculated with 2−△△CT method. Primer sequences were shown in Table 2.
Western blot
After Calu-3 cells were lysed, the protein concentration was measured with bicinchoninic acid (BCA) kit (Thermo, USA). Polyacrylamide gel electrophoresis (PAGE) was applied on 30 μg total proteins, and the proteins were then transferred onto a polyvinylidene fluoride membrane (Amersham, USA). Later, the membrane was blocked with 5% skim milk under room temperature and incubated with primary antibodies at 4 °C overnight after removing the seal solution. The membrane was washed with phosphate-buffered saline with 0.1% Tween-20 (PBST) 3 times, with 10 min of each time. Afterward, the membrane was incubated with horseradish peroxidase labelled secondary antibody for 1 h, and washed with PBST 3 times for 10 min of each time. At last, the membrane was scanned by an optical luminometer (GE, USA) for development. Antibody information was shown in Table 3.
Transwell invasion assay
A 24-well Transwell chamber (8 μm aperture, BD Biosciences) was applied here. The upper chamber was coated with Matrigel (Corning, Corning, NY) and the lower chamber was supplemented with DEME medium containing 10% FBS. About 5 × 104 Calu-3 cells were added into the upper chamber. After being cultured at 37 °C for 24 h, cells that did not pass the membrane were removed with a swab applicator. Cells under the membrane were stained with crystal violet (0.3%) and observed under a microscope in 4 random fields to calculate invaded cells.
Wound healing assay
When Calu-3 cells were grown into about 80% fusion in the well, a 200 μL pipette tip was used to scratch the cell monolayer. The well was washed with medium briefly twice to remove separated cells. Fresh medium was added for another 24 h of cell culture. Cells at 0 h and 24 h were photographed with a microscope for measuring the wound width to calculate cell migratory rate. Migratory rate = (0 h wound width—24 h wound width) /0 h wound width.
Cell adhesion assay
The 96-well plates were precoated with 100 mg/ml fibronectin at 4 °C overnight and blocked with 1% BSA at 37 °C for 1 h. Next, 2 × 104 Calu-3 cells were inoculated into the 96-well plates and cultured in serum-free DMEM. After 2 h of culture, cells were rinsed with PBS 3 times to gently remove nonadherent cells. Thereafter, attached cells were fixed with 4% paraformaldehyde, and stained with 0.5% crystal violet (Sangon Biotech). The stained crystal violet was dissolved with lauryl sodium sulfate (Amresco, Solon, OH, USA). Absorbance at 570 nm was read with a microplate reader.
Dual-luciferase reporter assay
Wild-type or mutant 3’UTR sequences of CTHRC1 (CTHRC1 WT, CTHRC1 MUT) were cloned into pmirGLO (Promega, WI, USA) vector to construct 2 luciferase reporter vectors. Taken renilla luciferase expression vector pRL-TK (TaKaRa, Dalian, China) as an internal reference, miR-30a-5p mimic and NC mimic were co-transfected into HEK-293T cells with luciferase reporter vectors, respectively. Dual-luciferase activity detection was conducted based on Dual-Luciferase Reporter Assay System of Promega (Promega, Madison, WI, USA).
Statistical analysis
All data were treated on SPSS21.0 statistical software (SPSS, Inc, Chicago, IL, USA). Measurement data were displayed as MEAN ± SD and comparison between 2 groups were analyzed by t-test. Patient’s overall survival curve was calculated with Kaplan–Meier and patient’s survival differences were analyzed by log-rank. P < 0.05 represents that the difference is statistically remarkable, and p < 0.01 indicates that the difference is extremely remarkable.
Results
CTHRC1 is significantly high and associated with poor prognosis
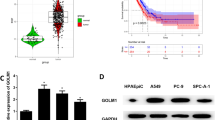
Previous studies indicated that high CTHRC1 expression is closely linked with tumor metastasis [18,19,20,21]. However, the action mechanism of CTHRC1 in LUAD is rarely studied, hence it was chosen for research in this study. It was represented by analyzing TCGA-LUAD data set and 3 GEO data sets that CTHRC1 was significantly high in LUAD tissue (Fig. 1A), and CTHRC1 was upregulated in invasive CL1-5 cell line than that in non-migrated CL1-0 cell line (Fig. 1B). Moreover, it was revealed by GEO data sets (GSE31210, GSE72094) and TCGA-LUAD data set with follow-up records that high CTHRC1 expression was remarkably detrimental to patient’s prognosis (Fig. 1C), which indicated that aberrant expression of CTHRC1 may affected LUAD progression. In addition, similar result was found in LUAD cell lines that CTHRC1 expression was higher in 4 LUAD cells than that inBEAS-2B (Fig. 1D). The above results suggested that CTHRC1 was significantly high and related to poor prognosis in LUAD.

CTHRC1 is significantly high in LUAD cell lines, and related to prognosis of LUAD. A Boxplot of CTHRC1 expression in normal and tumor tissues and several GEO data sets (GSE31210, GSE32863, GSE43458, GSE75037, GSE116959); B Histogram of CTHRC1 expression in CL1-0 and CL1-5 analyzed by GEO data set GSE42407; C Survival curve of CTHRC1 expression on patient’s prognosis in GSE31210, GSE72094 and TCGA-LUAD data sets, with the red line referring to high expression and blue line referring to low expression; D CTHRC1 expression in normal human bronchial epithelial cell line and LUAD cell lines; * represents p < 0.05, ** represents p < 0.01, **** represents p < 0.0001
CTHRC1 accelerates LUAD cell invasion and migration, and inhibits cell adhesion
Studies indicated that high CTHRC1 expression promotes the invasive and migratory abilities of tumor cells [22,23,24]. To testify that CTHRC1 could regulate LUAD cell invasion/migration, it was overexpressed or knocked down in Calu-3 cell line. Firstly, transfection efficiency was measured. It was found that CTHRC1 expression elevated after transfecting overexpressed vector. The interfering efficiency of si-CTHRC1#2 was the highest, therefore it was chosen for the following experiments (Fig. 2A). Overexpressing CTHRC1 noticeably increased the invasive and migratory capabilities of LUAD cells, while the interference group showed the opposite results (Fig. 2B, C). Hence, it was speculated that CTHRC1 impacted LUAD cell migration and invasion. The expression of tumor metastasis-related proteins supported our speculation. Relative to the control group, the expression of 2 proteins noticeably upregulated in oe-CTHRC1 group while those in si-CTHRC1 group was downregulated (Fig. 2D). Based on above results, it was speculated that CTHRC1 could affect cell invasion and migration in LUAD.

CTHRC1 promotes LUAD cell invasion and migration, and inhibits cell adhesion. A CTHRC1 expression in Calu-3 cells in each group; B, C Changes of invasive and migratory abilities of Calu-3 cells after transfecting overexpressing vector, si-CTHRC1 and its negative control; D Expression levels of metastasis-related proteins MMP2 and MMP9 after transfection; E Results of GSEA pathway enrichment analysis; F The effect of CTHRC1 on cell adhesion detected after transfection; * represents p < 0.05, ** represents p < 0.01
To further study the pathways of CTHRC1 regulating tumor cell invasion and migration, univariate GSEA pathway enrichment analysis was performed. It was suggested that CTHRC1 was remarkably pertinent to extracellular matrix (ECM) receptor interaction and focal adhesion signaling pathways (Fig. 2E). These signaling pathways were closely related to cell adhesion ability, adhesion ability measurement was therefore conducted. Results showed that overexpressed CTHRC1 markedly declined the adhesion ability of LUAD, while interfering CTHRC1 elevated cell adhesion activity (Fig. 2F), which illustrated that CTHRC1 inhibited LUAD cell adhesion.
CTHRC1 is targeted and modulated by miR-30a-5p
To further research regulatory mechanism of CTHRC1, upstream miRNAs that regulated CTHRC1 were predicted with 4 databases and then intersected to obtain 5 miRNAs (miR-30e-5p, miR-30d-5p, miR-30b-5p, miR-30a-5p, miR-30c-5p,) (Fig. 3A). By detecting mature miRNA data set in TCGA-LUAD, it was manifested that miR-30a-5p was notably low in tumor tissue (Fig. 3B). Fold change |logFC|> 1, while fold change of other miRNAs was less than 1 (Additional file 1: Table S1). The correlation between miR-30a-5p and CTHRC1 was significantly negative (Fig. 3C). Hence, miR-30a-5p was chosen for research. miR-30a-5p low expression was confirmed in cell lines, which was similar to bioinformatics analysis (Fig. 3D). Afterward, binding sequences between miR-30a-5p and CTHRC1 were predicted via TargetScan database (Fig. 3E). Dual-luciferase method validated the reliability of predicted sites. It was represented that the luciferase activity of cells after co-transfecting WT CTHRC1 and miR-30a-5p mimic was markedly decreased, which indicated that miR-30a-5p could targeted CTHRC1 3’UTR (Fig. 3F). Moreover, it was suggested by qRT-PCR and western blot that the expression of CTHRC1 mRNA and protein significantly declined after cells transfecting miR-30a-5p mimic (Fig. 3G). The above results implied that miR-30a-5p was markedly low in LUAD, and could target and downregulate CTHRC1 expression.

MiR-30a-5p downregulates CTHRC1 expression. A Venn plot of target miRNAs predicted via bioinformatics databases; B Boxplot of the expression of 5 predicted upstream miRNA in normal tissues and tumor tissues; C Pearson correlation analysis between miR-30a-5p and CTHRC1 expression in GSE119269 and TCGA-LUAD data set; D The expression of miR-30a-5p in normal human bronchial epithelial cell line and LUAD cell lines; E Targeted binding sequences between miR-30a-5p and CTHRC1 predicted with starBase database; F Binding relationship between miR-30a-5p and CTHRC1; G The expression of CTHRC1 mRNA and protein after overexpressing miR-30a-5p. * represents p < 0.05 and ** represents p < 0.01
MiR-30a-5p targets CTHRC1 to regulate LUAD cell invasion, migration and cell adhesion
Calu-3 cell lines were divided into 3 groups: mimic + oe-NC group, NC mimic + oe-NC group, and mimic + oe-CTHRC1 group. mRNA and protein levels of CTHRC1 of cells were detected. The result elaborated that overexpressing miR-30a-5p markedly suppressed CTHRC1 expression while simultaneously overexpressing CTHRC1 reversed the results in some degree (Fig. 4A, B). Afterward, invasive and migratory abilities in each group were detected. The result showed that invading cells and migratory rate of cells in mimic + oe-NC group markedly declined (Fig. 4C, D), and MMP2 and MMP9 expression declined (Fig. 4E). However, after simultaneously overexpressing miR-30a-5p and CTHRC1, invasive and migratory abilities of Calu-3 cells were recovered (Fig. 4C, D) and metastasis-related protein expression was also elevated (Fig. 4E). It could be concluded that overexpressing miR-30a-5p strengthened adhesion of Calu-3 cells while overexpressing CTHRC1 reversed that effect (Fig. 4F). Above results proved that miR-30a-5p affected LUAD cell invasion, migration and cell adhesion through targeting and downregulating CTHRC1.

MiR-30a-5p affected the invasion, migration and adhesion of LUAD cells via targeting and downregulating CTHRC1. A CTHRC1 expression of Calu-3 cells in each group; B CTHRC1 protein expression of Calu-3 cells in each group; C, D Changes of invasive and migratory abilities of Calu-3 cells in each group; E Expression of metastasis-related proteins MMP2 and MMP9 of Calu-3 cells in each group; F Cell adhesion of Calu-3 cells in each group. * represents p < 0.05
Discussion
Nowadays, with the development of precise medicine, molecular targeted therapy has been a research hotspot besides conventional therapy. Anaplastic lymphoma kinase (ALK), epidermal growth factor receptor (EGFR), c-ros Repressor of Silencing 1 (ROS1) as crucial lung cancer gene targets have been unanimously recognized by today academic circles, and the era of targeted therapy has begun [24]. More and more biomarkers related to LUAD diagnosis, treatment and prognosis need to be studied. CTHRC1 is an ECM protein associated with atherosclerosis [8], and is upregulated in various cancers and involved in several biological functions of tumors [19, 21, 25, 26], which has value as a potential therapeutic target for cancer. It was manifested by several bioinformatics database profiles and clinical information that CTHRC1 expression was significantly upregulated and related to poor prognosis in LUAD. To further verify the function of CTHRC1, a series of cell experiments in vitro were undertaken. The results indicated that overexpressed CTHRC1 promotes LUAD cell invasion and migration, which coincided with a study about NSCLC [22].
Pathway enrichment analysis was performed for mechanism investigation. It was exhibited that CTHRC1 was mainly enriched in signaling pathways like ECM receptor interaction, focal adhesion, and actin skeleton regulation. These pathways were related to cell adhesion. Proteolytic degradation of the stromal ECM accelerates malignant invasion and metastasis of tumor cells [27, 28]. Additionally, MMPs is a type of zincdependent endopeptidases, and involved in degrading ECM and promoting tumor invasion [29, 30]. After detecting the expression of MMP2 and MMP9 proteins after overexpressing CTHRC1, it was disclosed that the expression of the 2 matrix metalloproteinases was significantly elevated. Moreover, high CTHRC1 expression declined the adhesive ability of LUAD cells, which promoted cancer cell invasion and metastasis.
Our study discovered that CTHRC1 played a vital role in LUAD progression, with the reason for its aberrant expression to be further discussed. MiRNA can target 3’UTR of mRNA so as to regulate mRNA expression. Many studies reported that CTHRC1 is regulated by miRNA in various cancers [15, 16, 31], therefore upstream miRNAs that may target CTHRC1 were predicted via many databases. Only miR-30a-5p was dramatically low in tumor tissues among 5 predicted miRNAs. The binding between miR-30a-5p and CTHRC1 was verified. MiR-30a-5p is found to exert as a cancer-inhibitor in various cancers. For example, CTHRC1 can inhibit tumor growth by suppressing glycolysis via adenosine triphosphate ATP generation, and extracellular acidification rate (ECAR), while increasing oxygen consumption rate OCR in breast cancer cells [32]. MiR-30a-5p inhibits migration of osteosarcoma cells through modulating FOXD1 [33]. In NSCLC, miR-30a-5p suppresses epithelial-mesenchymal transition of cell lines in highly invasive NSCLC via targeting profilin-2 [34], and can strengthen the sensitivity of paclitaxel to NSCLC by targeting BCL-2 expression [35]. In our study, it was uncovered that miR-30a-5p could downregulate CTHRC1 to inhibit LUAD cell invasion and migration, and increase cell adhesive ability. Nevertheless, miR-30a-5p low expression may cause abnormal expression of CTHRC1.
In conclusion, it was revealed in this study that CTHRC1 was remarkably high in LUAD and related to prognosis. High CTHRC1 stimulated LUAD cell invasion and migration, and inhibited cell adhesion. Additionally, CTHRC1 was targeted and negatively regulated by miR-30a-5p to influence LUAD progression. However, the specific regulatory mechanism of CTHRC1 needs to be further researched and it will be further explored in our future studies.
Availability of data and materials
The data used to support the findings of this study are included within the article. The data and materials in the current study are available from the corresponding author on reasonable request.
References
Bray F, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424. https://doi.org/10.3322/caac.21492.
Wang R, et al. EIF4A3-induced circular RNA MMP9 (circMMP9) acts as a sponge of miR-124 and promotes glioblastoma multiforme cell tumorigenesis. Mol Cancer. 2018;17:166. https://doi.org/10.1186/s12943-018-0911-0.
An Y, Furber KL, Ji S. Pseudogenes regulate parental gene expression via ceRNA network. J Cell Mol Med. 2017;21:185–92. https://doi.org/10.1111/jcmm.12952.
Hirsch FR, et al. Lung cancer: current therapies and new targeted treatments. Lancet. 2017;389:299–311. https://doi.org/10.1016/S0140-6736(16)30958-8.
Ni M, et al. Epithelial mesenchymal transition of non-small-cell lung cancer cells A549 induced by SPHK1. Asian Pac J Trop Med. 2015;8:142–6. https://doi.org/10.1016/S1995-7645(14)60305-9.
Riaz SP, et al. Trends in incidence of small cell lung cancer and all lung cancer. Lung Cancer. 2012;75:280–4. https://doi.org/10.1016/j.lungcan.2011.08.004.
Takeshita S, et al. Osteoclast-secreted CTHRC1 in the coupling of bone resorption to formation. J Clin Invest. 2013;123:3914–24. https://doi.org/10.1172/JCI69493.
Pyagay P, et al. Collagen triple helix repeat containing 1, a novel secreted protein in injured and diseased arteries, inhibits collagen expression and promotes cell migration. Circ Res. 2005;96:261–8. https://doi.org/10.1161/01.RES.0000154262.07264.12.
Kim HC, et al. Collagen triple helix repeat containing 1 (CTHRC1) acts via ERK-dependent induction of MMP9 to promote invasion of colorectal cancer cells. Oncotarget. 2014;5:519–29. https://doi.org/10.18632/oncotarget.1714.
Ma MZ, et al. CTHRC1 acts as a prognostic factor and promotes invasiveness of gastrointestinal stromal tumors by activating Wnt/PCP-Rho signaling. Neoplasia. 2014;16:265–78. https://doi.org/10.1016/j.neo.2014.03.001.
Eriksson J, et al. Gene expression analyses of primary melanomas reveal CTHRC1 as an important player in melanoma progression. Oncotarget. 2016;7:15065–92. https://doi.org/10.18632/oncotarget.7604.
Zheng M, Zhou Q, Liu X, Wang C, Liu G. CTHRC1 overexpression promotes cervical carcinoma progression by activating the Wnt/PCP signaling pathway. Oncol Rep. 2019;41:1531–8. https://doi.org/10.3892/or.2019.6963.
Chen Y, et al. High CTHRC1 expression may be closely associated with angiogenesis and indicates poor prognosis in lung adenocarcinoma patients. Cancer Cell Int. 2019;19:318. https://doi.org/10.1186/s12935-019-1041-5.
Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120:15–20. https://doi.org/10.1016/j.cell.2004.12.035.
Liu J, Chen Z, Xiang J, Gu X. MicroRNA-155 acts as a tumor suppressor in colorectal cancer by targeting CTHRC1 in vitro. Oncol Lett. 2018;15:5561–8. https://doi.org/10.3892/ol.2018.8069.
Lai YH, et al. Collagen triple helix repeat containing-1 negatively regulated by microRNA-30c promotes cell proliferation and metastasis and indicates poor prognosis in breast cancer. J Exp Clin Cancer Res. 2017;36:92. https://doi.org/10.1186/s13046-017-0564-7.
Wang CY, et al. MicroRNA-98 suppresses cell proliferation, migration and invasion by targeting collagen triple helix repeat containing 1 in hepatocellular carcinoma. Mol Med Rep. 2016;13:2639–44. https://doi.org/10.3892/mmr.2016.4833.
Thijssen JM, Oosterveld BJ, Wagner RF. Gray level transforms and lesion detectability in echographic images. Ultrason Imaging. 1988;10:171–95. https://doi.org/10.1177/016173468801000302.
Ni S, et al. CTHRC1 overexpression predicts poor survival and enhances epithelial-mesenchymal transition in colorectal cancer. Cancer Med. 2018;7:5643–54. https://doi.org/10.1002/cam4.1807.
Chen G, et al. miR-155-5p modulates malignant behaviors of hepatocellular carcinoma by directly targeting CTHRC1 and indirectly regulating GSK-3beta-involved Wnt/beta-catenin signaling. Cancer Cell Int. 2017;17:118. https://doi.org/10.1186/s12935-017-0469-8.
Wang C, et al. High expression of Collagen Triple Helix Repeat Containing 1 (CTHRC1) facilitates progression of oesophageal squamous cell carcinoma through MAPK/MEK/ERK/FRA-1 activation. J Exp Clin Cancer Res. 2017;36:84. https://doi.org/10.1186/s13046-017-0555-8.
He W, et al. CTHRC1 induces non-small cell lung cancer (NSCLC) invasion through upregulating MMP-7/MMP-9. BMC Cancer. 2018;18:400. https://doi.org/10.1186/s12885-018-4317-6.
Ye J, et al. Upregulated CTHRC1 promotes human epithelial ovarian cancer invasion through activating EGFR signaling. Oncol Rep. 2016;36:3588–96. https://doi.org/10.3892/or.2016.5198.
Jin XF, Li H, Zong S, Li HY. Knockdown of collagen triple helix repeat containing-1 inhibits the proliferation and epithelial-to-mesenchymal transition in renal cell carcinoma cells. Oncol Res. 2016;24:477–85. https://doi.org/10.3727/096504016X14685034103716.
Guo B, et al. Collagen triple helix repeat containing 1 (CTHRC1) activates Integrin beta3/FAK signaling and promotes metastasis in ovarian cancer. J Ovarian Res. 2017;10:69. https://doi.org/10.1186/s13048-017-0358-8.
Lee J, et al. CTHRC1 promotes angiogenesis by recruiting Tie2-expressing monocytes to pancreatic tumors. Exp Mol Med. 2016;48:e261. https://doi.org/10.1038/emm.2016.87.
Johnsen M, Lund LR, Romer J, Almholt K, Dano K. Cancer invasion and tissue remodeling: common themes in proteolytic matrix degradation. Curr Opin Cell Biol. 1998;10:667–71. https://doi.org/10.1016/s0955-0674(98)80044-6.
Butcher DT, Alliston T, Weaver VM. A tense situation: forcing tumour progression. Nat Rev Cancer. 2009;9:108–22. https://doi.org/10.1038/nrc2544.
Basset P, et al. Matrix metalloproteinases as stromal effectors of human carcinoma progression: therapeutic implications. Matrix Biol. 1997;15:535–41. https://doi.org/10.1016/s0945-053x(97)90028-7.
Kenny HA, Kaur S, Coussens LM, Lengyel E. The initial steps of ovarian cancer cell metastasis are mediated by MMP-2 cleavage of vitronectin and fibronectin. J Clin Invest. 2008;118:1367–79. https://doi.org/10.1172/JCI33775.
Zhou S, et al. MiR-9 inhibits Schwann cell migration by targeting Cthrc1 following sciatic nerve injury. J Cell Sci. 2014;127:967–76. https://doi.org/10.1242/jcs.131672.
Li L, et al. miR-30a-5p suppresses breast tumor growth and metastasis through inhibition of LDHA-mediated Warburg effect. Cancer Lett. 2017;400:89–98. https://doi.org/10.1016/j.canlet.2017.04.034.
Tao J, et al. MiR-30a-5p inhibits osteosarcoma cell proliferation and migration by targeting FOXD1. Biochem Biophys Res Commun. 2018;503:1092–7. https://doi.org/10.1016/j.bbrc.2018.06.121.
Yan J, Ma C, Gao Y. MicroRNA-30a-5p suppresses epithelial-mesenchymal transition by targeting profilin-2 in high invasive non-small cell lung cancer cell lines. Oncol Rep. 2017;37:3146–54. https://doi.org/10.3892/or.2017.5566.
Xu X, et al. miR-30a-5p enhances paclitaxel sensitivity in non-small cell lung cancer through targeting BCL-2 expression. J Mol Med (Berl). 2017;95:861–71. https://doi.org/10.1007/s00109-017-1539-z.
Acknowledgements
Not applicable.
Funding
Funding was provided by Respiratory medicine is a key discipline of Guangxi medical and health care (Grant No. 2021QZD01).
Author information
Authors and Affiliations
Contributions
All authors contributed to data analysis, drafting and revising the article, gave final approval of the version to be published, and agreed to be accountable for all aspects of the work. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors consent to submit the manuscript for publication.
Competing interests
The authors declare that they have no potential conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
LogFC Value of 5 Predicted Upstream miRNAs That May Regulate CTHRC1.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yang, C., Huang, T., Liang, Y. et al. CTHRC1 targeted by miR-30a-5p regulates cell adhesion, invasion and migration in lung adenocarcinoma. J Cardiothorac Surg 17, 46 (2022). https://doi.org/10.1186/s13019-022-01788-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13019-022-01788-9