
Abstract
Objective
This study aims to investigate the correlation of long non-coding RNA HOX transcript antisense RNA (lncRNA HOTAIR) with the PTEN/PI3K/AKT pathway and clinical-related indicators in osteoarthritis (OA) and determine the effect of baicalin intervention.
Methods
The levels of clinical lipid metabolism indexes and immune-inflammatory indexes in OA patients and normal controls was detected. OA chondrocytes (OA-CHs) were induced with peripheral blood mononuclear cells (PBMCs), followed by baicalin treatment (50 ug/mL). RT-qPCR was performed to measure lncRNA HOTAIR expression. The levels of inflammatory cytokines and adiponectin were detected using ELISA kits. CCK-8 assay was used to assess the viability of CHs. The related protein expression was measured using Western blot analysis.
Results
LncRNA HOTAIR might act as a biomarker of OA in vivo. LncRNA HOTAIR was positively correlated with TC, hs-CRP, IgA, TNF-α, and VAS score. Overexpression of lncRNA HOTAIR in vitro inhibited cell proliferation, reduced IL-10 and PTEN expression, but augmented TNF-α, p-PI3K, and p-AKT proteins in OA-CHs stimulated by OA-PBMCs. The changes of above indexes were also observed in OA-CHs stimulated by OA-PBMCs treated with si-lncRNA HOTAIR or baicalin, implying the synergistic effects of baicalin and lncRNA HOTAIR silencing on OA.
Conclusions
Conclusively, lncRNA HOTAIR was highly expressed in OA-CHs, which facilitated OA inflammatory responses by orchestrating inflammatory cytokines and the PTEN/PI3K/AKT pathway. Baicalin exerted therapeutic effects by inhibiting the expression of lncRNA HOTAIR, decreasing the protein levels of p-PI3K and p-AKT, and increasing the protein levels of PTEN, APN, and ADIPOR1.
Similar content being viewed by others


Introduction
Osteoarthritis (OA) is a chronic degenerative joint disease featured by low-level inflammation and cartilage degeneration [1, 2], leading to irreversible structural and functional changes in the joint. Clinical signs associated with OA include joint pain, swelling, and significant functional impairment. Multiple risk factors for OA have been reported, including age, joint trauma, obesity, and genetic susceptibility [3]. To a certain extent, obesity is accompanied with a state of chronic inflammation. Hyperlipidemia is a common metabolic change characterized by an abnormal increase in serum total cholesterol (TC), triglycerides, and low-density lipoprotein cholesterol, as well as an abnormal decrease in high-density lipoprotein cholesterol and apolipoprotein A1, all of which contribute to the inflammatory response and OA deterioration. In addition, body fat, insulin resistance index, and body mass index are negatively related to adiponectin (APN) [4, 5]. APN, a recently discovered cytokine released by adipose tissue, is a regulator for inflammatory resolution in OA [6]. Emerging evidence [7] has demonstrated the involvement of long non-coding RNAs (lncRNAs) in the inflammation, lipid metabolism, and other processes of OA, serving a fundamental role in the development and progression of OA [8].
A prior research [9] has observed that lncRNA HOX transcript antisense RNA (HOTAIR) expression in OA-chondrocytes (CHs) was apparently higher than that in normal CHs, and OA-CHs transfected with small interfering RNA (si)-lncRNA HOTAIR showed a stronger proliferation capacity than those transfected with si-negative control (NC). The tumor suppressor gene phosphatase and tensin homolog deleted on chromosome ten (PTEN) can dephosphorylate and inactivate phosphatidylinositol-3,4,5-trisphosphate (PIP3), an important molecule in the phosphoinositide 3-kinase (PI3K)/AKT pathway, to downregulate the PI3K/AKT pathway. The PTEN/PI3K/AKT pathway is considered a classic inflammatory pathway. PTEN can downregulate the expression of fatty acid synthase by inhibiting the PTEN/PI3K/AKT pathway [10]. Interleukin (IL)-10 and tumor necrosis factor-α (TNF-α) are the critical pro-inflammatory and anti-inflammatory factors of OA [11], respectively. Of note, the PTEN/PI3K/AKT pathway can increase TNF-α expression and decrease IL-10 expression [12]. Furthermore, lncRNA HOTAIR silencing can obviously reduce the content of TNF-α and enhance the content of IL-10 [13]. However, whether lncRNA HOTAIR can assume a role in immune regulation by affecting the PTEN/PI3K/AKT pathway in the inflammatory response of OA has not been clearly clarified yet.
Baicalin, an important flavonoid isolated from the dried roots of Scutellaria Baicalensis Georgi, is a powder of light-yellow color with bitter taste at room temperature. Baicalin possesses diverse pharmacological properties such as anti-inflammation, cholesterol lowering, and anti-allergic. Importantly, the effectiveness of baicalin in controlling OA progress has also attracted considerable concern. For example, it has been reported that treatment with baicalin not only prevents cartilage destruction but also alleviates IL-1β-induced inflammatory injury and represses the production of IL-6 and TNF-α in OA [14, 15]. These findings suggest that baicalin may be a potential agent in the treatment of OA. However, the exact mechanism of baicalin in OA is still unclear.
Therefore, this study focused on the expression pattern lncRNA HOTAIR in OA patients and analyzed its correlation with clinical indicators and visual analog scale (VAS) scores. Finally, lncRNA HOTAIR expression was intervened in cell experiments to assess its regulatory effect on the PTEN/PI3K/AKT pathway, inflammatory factors, and lipid metabolism indexes. Moreover, the involvement of baicalin in the development of immune inflammatory response in OA-CHs was determined, and the synergistic effect between baicalin and HOTAIR in the treatment of osteoarthritis was explored.
Materials and methods
Subjects and samples
OA patients and normal control (NC) subjects matched on gender and age were enrolled from the First Affiliated Hospital of Anhui University of Traditional Chinese Medicine between November 2020 and May 2021. Patients who did not meet the following criteria were excluded: not conforming to the 2019 Guidelines for the Diagnosis of OA [16], severe mental illness, significant liver or renal function impairment, the administration of immunosuppressive drugs, or pregnancy. This study was conducted following the relevant provisions of the Declaration of Helsinki [17] and was approved by the Ethical Committee of Scientific Research of Anhui University of Traditional Chinese Medicine's First Affiliated Hospital [2014AH-06(J)].
Co-culture of peripheral blood mononuclear cells (PBMCs) with OA-CHs and cell transfection
Totally, 5 mL venous blood was collected from normal healthy subjects and OA patients, and diluted with an equal volume of normal saline. The same amount of lymphocyte separation solution was slowly added into the prepared samples and centrifuged at 2000 r/min for 20 min. The white floc in the centrifuge tube was transferred into a clean centrifuge tube. The same amount of normal saline was added into the tube and mixed, followed by centrifugation at 800 r/min for 8 min and rinsing twice. Afterward, the samples were transferred into a culture flask and incubated for later use.
OA-CHs and baicalin were purchased from Saibaikang Biotechnology Co., Ltd. (Shanghai, China). PBMCs were seeded into the Transwell chamber and cultured in 75 µL Dulbecco’s Modified Eagle Medium (DMEM). CHs were digested and centrifuged. Then, the medium was discarded, and CHs were washed twice with phosphate buffer saline, resuspended with medium, and seeded into the basolateral chamber, followed by the supplementation of 100 µL DMEM. Following cell adherence, different proportions of OA-PBMCs were added into the apical chamber. Finally, the optimal stimulation concentration of PBMCs was screened by cell counting kit-8 (CCK-8) assay. On the basis of PBMC stimulation concentration (3:1, 48 h), the above steps were repeated, and baicalin at different concentrations was added into the basolateral chamber to screen the optimal concentration of baicalin. Co-culture was conducted in DMEM containing 100 U/mL penicillin and 0.1 mg/mL streptomycin with 5% CO2 at 37 °C until the confluence of CHs reached 70–90%. OA-CHs were transfected with PCDNA3.1-lncRNA HOTAIR plasmid, si-lncRNA HOTAIR plasmid, and corresponding NC plasmids using Lipofectamine 2000, respectively. After 24 h, the cells were harvested for subsequent analysis. All plasmids were purchased from GenePharma (Shanghai, China).
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
The total RNA was extracted from OA-CHs using TRIzol reagent, followed by reverse transcription reaction and amplification reaction. Agarose-gel electrophoresis was implemented for semi-quantitative analysis of PCR products using the Gelpro32 gel image analysis software. Relative quantitative analysis was performed using 2−ΔΔCt with β-actin as the internal reference. All used primers were as follows: lncRNA HOTAIR: the forward primer: 5'-CCATAGCCGATTAGCTGTCA' and the reverse primer: 5'-AATGCCGAACTGGAGGTG-3'; β-actin: the forward primer: 5'-GGCAAATGTCAGAGGGTTCT-3' and the reverse primer: 5'-TTCTTAAATTGGGCTGGGTC-3'.
Enzyme-linked immunosorbent assay (ELISA)
The supernatant of OA-CHs was collected and centrifuged at 2500 r/min for 20 min, and the precipitation was discarded. The supernatant was added to the ELISA plate (50 µL per well) and incubated at 37 °C for 30 min. After the plate was sealed with a sealing plate membrane, the levels of TNF-α, IL-10, APN, and adiponectin receptor 2 (ADIPOR2) were detected using the ELISA method strictly in the light of the kit instructions.
CCK-8 assay
The viability of OA-CHs was measured using CCK-8 assay kit (BIOSS, Beijing, China). At first, 3 × 104 OA-CHs were seeded into each well of 96-well plates and cultured until reaching 70–90% confluence. Logarithmically growing cells were transfected by the above method. OA-CHs in 3 wells in each group were cultured for 0, 12, 24, 48, and 72 h, respectively. Next, 10 mL CCK-8 solution was supplemented into each well at each point of time, followed by incubation at 37 °C for 1–4 h. The viability of OA-CHs was determined by measuring the absorbance value of each well at 450 nm.
Western blot analysis
The cells were lysed with radio-immunoprecipitation assay lysis solution and centrifuged at 12000 r/min for 10 min to harvest the total proteins, followed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) (Beyotime, Shanghai, China). The 5 × SDS-PAGE protein loading buffer was added into cells at the ratio of 1:4 to obtain protein samples. The samples were heated for 10 min in a boiling water bath and supplemented into SDS-PAGE gel wells (5–10 µL per well). Filter papers and polyvinylidene fluoride membrane of the same size as the rubber strip were cut in advance (soaked in methanol for 2–3 min in advance) and immersed in the rotating film buffer for 5 min. Bubbles were removed at every step and the flow membrane was constant. After the protein was transferred into the membranes, the membranes were cooled to room temperature before the insertion of the membranes into the prepared Western washing solution and the removal of the membrane transfer solution by 5-min washing. The membranes were blocked with 5% skim milk powder at room temperature for 2 h and then washed with tris-buffered saline-tween (TBST) buffer 3 times (10 min/time). The membranes underwent overnight incubation with primary pig antibodies to phosphorylation (p)-AKT, PI3K, PTEN, AKT, P-PI3K, APN, ADIPOR1, and ADIPOR2 at 4 °C. The horseradish peroxidase-tagged secondary antibody was diluted with secondary antibody diluent at the ratio of 1:20,000 and used for incubation with the membranes at room temperature for 1 h. The ratio of the absorbance of target proteins to that of glyceraldehyde-3-phosphate dehydrogenase was calculated after electrogenerated chemiluminescence development, darkroom exposure, fixation, and photography.
Molecular modeling
The two-dimensional structure diagram of active components of baicalin was downloaded from Traditional Chinese Medicine Systems Pharmacology (TCMSP) database. The compound structure and AKT1 were converted to PDBQT format by AutoDockTools-1.5.7 software, and the active pockets were formed. Finally, the PDBQT ligand obtained by entering CMD command and the PDBQT core protein obtained by AutoDockTools-1.5.7 were imported into Pymol for docking.
Statistical analysis
SPSS statistical software 23.0 (IBM Corp. Armonk, NY, USA) was applied for statistical analysis, and GraphPad Prism software 8.2 (GraphPad Software, La Jolla, CA, USA) was utilized to capture images. The difference between groups was compared using Student's paired two-tail t test or Kruskal–Wallis nonparametric test. Classification variables were compared using chi-square test. Spearman correlation analysis was employed to evaluate the correlation of lncRNA HOTAIR with erythrocyte sedimentation rate (ESR), high sensitivity C-reactive protein (hs-CRP), immunoglobulin A (IgA), and other indicators. Data were expressed as mean ± standard deviation or median (quartile range). p < 0.05 was indicative of statistical significance.
Results
Clinical characteristics of OA patients and NC subjects
A total of 80 subjects were enrolled, including 30 NC subjects (10 males and 20 females, the age quartile was 55.50 (42.75, 70.0)) in the control group and 50 OA patients (17 males and 33 females, the age quartile was 55.0 (48.5, 65.25)) in the OA group. There were no statistically significant differences in age and gender distribution between the two groups, but there were significant differences in ESR, TC, TG, IgA, and C3 (Table 1).
LncRNA HOTAIR expression was upregulated in the PBMCs of OA patients
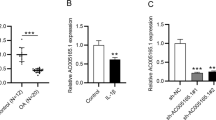
This study intended to assess the lncRNA HOTAIR expression pattern in OA. LncRNA HOTAIR expression in the PBMCs of 50 OA patients and 30 NC subjects was detected using RT-qPCR. The findings revealed that lncRNA HOTAIR expression was increased in the PBMCs of OA patients (Fig. 1A). The diagnostic efficacy of lncRNA HOTAIR was evaluated using receiver-operating characteristic (ROC) curve analysis, and the area under the ROC curve (AUC) was 0.8310 [95% confidence interval (CI): 0.74–0.92]. According to the Youden index, the ideal cut-off value for identification of OA patients from NC subjects was 1.185 with a sensitivity of 60.00% and a specificity of 92.67% (Fig. 1B).

LncRNA HOTAIR was highly expressed in the PBMCs of OA patients. A LncRNA HOTAIR expression in OA patients was higher than that in NC subjects. B Comparison of ROC curves of lncRNA HOTAIR expression in PBMCs of OA patients and NC patients. **p < 0.01
Differential expression of inflammatory cytokines and APN in OA had a positive correlation with lncRNA HOTAIR expression
IL-10 was dramatically reduced (p < 0.01), while TNF-α was elevated (p < 0.01) in the OA group,, as compared to the control group, indicating the significant alterations in the levels of pro-inflammatory and anti-inflammatory factors in OA (Fig. 2A–B). Furthermore, the levels of APN and ADIPOR2 were considerably reduced in the OA group compared with those in the control group (p < 0.01; Fig. 2C–D). Then, we conducted correlation analyses of lncRNA HOTAIR with TC, hs-CRP, IgA, TNF-α, and VAS, respectively. The results displayed that lncRNA HOTAIR was a risk factor for TC, hs-CRP, IgA, TNF-α, and VAS [18] (Fig. 2E–I). These results further suggested the close relationship between lncRNA HOTAIR and the occurrence and development of OA.

Differential expression of inflammatory cytokines in OA. A–D The levels of TNF-α (A), IL-10 (B), APN (C), and ADIPOR2 (D) in the serum of OA patients and normal subjects. *** p < 0.001. (F-I) Correlation analyses of lncRNA HOTAIR with TC (E), hs-CRP (F), IgA (G), TNF-α (H), and VAS (I) in the PBMCs of OA patients
Association rule analysis of lncRNA HOTAIR with laboratory indexes in OA patients
Association rule analysis showed that the elevation of lncRNA HOTAIR was strongly correlated with the elevation of GSH, ESR, and VAS in OA patients. There was a strong correlation between the increase of lncRNA HOTAIR and the decrease of LDL-C, with a support degree greater than 20% and a confidence degree greater than 30%. The results are shown in Table 2.
Optimal concentration of PBMCs, baicalin, and si-lncRNA HOTAIR, and effect of overexpression or knockdown of lncRNA HOTAIR on cell viability
OA-CHs were treated with different concentrations of OA-PBMCs and baicalin. The optimal stimulation concentration of PBMCs (3:1, 48 h), baicalin (50 ug/ml, 48 h), and si-lncRNA HOTAIR 2# was determined by CCK-8 assay (Fig. 3A–C) for subsequent experiments. The effect of lncRNA HOTAIR on the viability of OA-CHs was assessed by CCK-8 assay. The results exhibited that the PCDNA3.1-lncRNA HOTAIR group had dramatically boosted cell viability when compared to the PCDNA3.1-NC group. In contrast to the si-NC group, the cell viability of the si-lncRNA HOTAIR group was evidently diminished. Among the four groups, the viability of cells in the si-lncRNA HOTAIR group was the highest and that of cells in the lncRNA HOTAIR group was the lowest (Fig. 3D). LncRNA HOTAIR expression in the PCDNA3.1-lncRNA HOTAIR group was substantially higher (p < 0.01) than that in the PCDNA3.1-NC group, whereas lncRNA HOTAIR expression in the si-lncRNA HOTAIR group was significantly lower than that in the si-NC group (p < 0.01) (Fig. 3E).

The optimal stimulation concentration of PBMCs (3:1, 48 h), baicalin (50 ug/ml, 48 h), and si-lncRNA HOTAIR 2# was screened and lncRNA HOTAIR silencing facilitated the viability of OA-CHs. A PBMCs: OA-CHs screening of the optimal concentration. B Baicalin screening of the optimal concentration. C Screening of si-lncRNA HOTAIR. D Effect of lncRNA HOTAIR on the viability of OA-CHs was tested by CCK-8 assay. E LncRNA HOTAIR expression in OA-CHs was measured by RT-qPCR. PC, PBMCs: OA-CHs. pc, pcDNA3.1. All tests were repeated three times and results were expressed as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001
LncRNA HOTAIR overexpression facilitated TNF-α expression but repressed IL-10 expression in OA-CHs
The levels of cytokines were evaluated by ELISA. Compared with that in the PCDNA3.1-NC group, TNF-α expression in the PCDNA3.1-lncRNA HOTAIR group was prominently augmented and IL-10 expression was remarkably decreased (p < 0.01). Compared with that in the si-NC group, TNF-α expression was markedly reduced and IL-10 expression was noticeably elevated in the si-lncRNA HOTAIR group (p < 0.01; Fig. 4A–B).

LncRNA HOTAIR overexpression caused TNF-α upregulation and IL-10 downregulation in OA-CHs. A–B The levels of IL-10 (A) and TNF-α (B) in OA-CHs were determined by ELISA. pc, pcDNA3.1. **p < 0.01, ***p < 0.001
LncRNA HOTAIR manipulated the PTEN/PI3K/AKT pathway in OA-CHs
To ascertain the impact of lncRNA HOTAIR on the PTEN/PI3K/AKT pathway, we adopted Western blot analysis to identify the expression of PI3K, p-AKT, p-PI3K, AKT, APN, ADIPOR1, ADIPOR2, and PTEN. As discovered in Fig. 5, overexpression of lncRNA HOTAIR enhanced the expression of p-PI3K and p-AKT and reduced the expression of APN, ADIPOR1, and PTEN. On the contrary, lncRNA HOTAIR silencing contributed to the opposite trends. However, the overexpression or silence of lncRNA HOTAIR did not have a significant effect on the expression of PI3K, AKT, and ADIPOR2.

Effect of lncRNA HOTAIR overexpression or silencing on the PTEN/PI3K/AKT pathway in OA-CHs. A Representative Western blots of PI3K, p-AKT, p-PI3K, AKT, APN, ADIPOR1, ADIPOR2, and PTEN in OA-CHs. B Quantitative analysis of the expression of p-AKT/AKT, p-PI3K/PI3K, and PTEN in OA-CHs. C Quantitative analysis of the expression of APN, ADIPOR1, and ADIPOR2 in OA-CHs. pc, pcDNA3.1. All experiments were repeated three times and data were expressed as mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001
Molecular docking between baicalin and AKT1
To explore the mechanism of baicalin, we next performed computational molecular docking analysis to evaluate whether there was any affinity between baicalin and RAC-alpha serine/threonine-protein kinase (AKT1) protein. The chemical structure of baicalin was employed, and it was discovered that baicalin interacted with and docked at the AKT1 binding site (Fig. 6A). The space-filling model was also used to illustrate the interaction between baicalin and AKT1 (Fig. 6B). High-affinity (− 6.06 kcal/mol) hydrogen binding events were observed between the residues of ARG-15, THR-87, GLY-16, and GLU-85 in baicalin and AKT1. These results indicated that baicalin probably inhibited the development of OA by interacting with the AKT1.

Molecular Docking between Baicalin and AKT1. A Macro- and local-level views between the residues of ARG-15, THR-87, GLY-16, and GLU-85 in baicalin and AKT1. B Space-filling model
Baicalin and si-lncRNA HOTAIR exerted synergistic effects on the PTEN/PI3K/AKT signaling pathway, APN, and inflammatory factors
To evaluate the contribution of baicalin relieving OA, we explored the effect of baicalin on the PTEN/PI3K/AKT signaling pathway, APN, and inflammatory factors after si-lncRNA HOTAIR treatment in OA-PBMCs + OA-CHs. The results showed that baicalin or si-lncRNA HOTAIR inhibited the expression of lncRNA HOTAIR and reduced the levels of IL-6 and TNF-α (Fig. 7A–C). In addition, baicalin reduced the levels of p-AKT and p-PI3K proteins and increased the levels of PTEN, APN, and ADIPOR1 proteins, showing a similar effect with si-lncRNA HOTAIR. Besides, there was no synergistic effect in reducing IL-6 and increasing APN. For the rest of indicator, baicalin and si-lncRNA HOTAIR had a certain degree of synergy.

Baicalin and si-lncRNA HOTAIR had synergistic effects on the PTEN/PI3K/AKT signaling pathway, APN, and inflammatory factors. After lncRNA HOTAIR knockdown and/or baicalin intervention, the protein expression of lncRNA HOTAIR, IL-6, and TNF-α was evaluated by RT-qPCR and ELISA (A–C). The protein expression of p-AKT/AKT, p-PI3K/PI3K, PTEN, APN, and ADIPOR1 was evaluated by Western blot assay (D–F). PC, PBMCs: OA-CHs. All results were represented as the mean ± standard deviation. *p < 0.05, **p < 0.01, ***p < 0.001; n = 3
Discussion
As the most prevalent chronic joint disease, OA causes cartilage loss, joint pain, and stiffness [19, 20]. Aging and obesity are well-known risk factors for OA, leading to a high susceptibility to joint injuries [21]. The critical role of inflammation in the pathophysiology of OA has been well-documented. Synovial inflammation resulting in the changes in the levels of inflammatory factors such as IL-10 and TNF-α has been associated with the radiographic and pain progression of OA [22,23,24]. TNF-α as an essential pro-inflammatory factor can induce the activation of fibroblast-like synoviocytes in OA and cause the gradual destruction of cartilage. IL-10 is known to be a potent anti-inflammatory cytokine, and IL-10 deregulation plays a role in the development of OA. Moreover, existing studies [25, 26] have shown that lipid metabolism imbalance contributes to the occurrence and development of OA, and hyperlipidemia is closely related to the immune-inflammatory response of OA patients. APN, an indicator related to lipid metabolism, induces inflammatory responses [27]. Kabalyk et al. [28] have confirmed that hyperlipidemia directly acts on articular tissues and causes cellular stress, manifested as changes in morphological and functional characteristics of CHs. Processes including cell death, pathological mineralization of articular cartilages, and enhanced pathological angiogenesis can be observed in hyperlipidemia. The quantity of inflammatory cytokines (including IL-1β, IL-6, IL-8, and TNF-α) is significantly augmented in a lipopolysaccharide-induced C28/I2 human chondrocyte cell model compared with the corresponding control group [29, 30].
LncRNA research constitutes a novel and promising field in understanding the complexity of OA pathogenesis. The involvement of lncRNA HOTAIR in OA progression has been reported by several previous studies [31,32,33]. For example, it is reported that lncRNA HOTAIR is highly expressed in OA, and participates in IL-1β-induced matrix metalloproteinase overexpression and promotes inflammatory responses in CHs [34]. A prior research has also elucidated that the PI3K/AKT pathway assumes a key role in the development of OA inflammation [35]. Moreover, the PI3K/AKT pathway exerts a certain effect on lipid metabolism [36].
This study provided evidence of substantially high expression of lncRNA HOTAIR in OA-PBMCs and OA-CHs with the AUC of 0.8310, the optimal truncation value of 1.185, the sensitivity of 60.00%, and the specificity of 92.67%, illustrating that lncRNA HOTAIR had the high diagnostic value. Our results also exhibited that IL-10, APN, and ADIPOR2 levels were diminished and TNF-α level was augmented in OA patients. APN can control inflammation, functioning as a potential therapeutic target for OA [37]. All changes in the levels of cytokines, APN, and its receptors can be indicated as the deregulation of pro-inflammatory and anti-inflammatory factors. In addition, clinical trials have depicted that lncRNA HOTAIR shares a positive correlation with TNF-α, clinical immune-inflammatory indicators, lipid metabolism indexes, and patient perception score scales, like TC, hs-CRP, IgA, and VAS. LncRNA HOTAIR dysregulation is partially responsible for the imbalance of inflammatory factors. To further dissect out the effects of lncRNA HOTAIR on the proliferation, inflammation, and lipid metabolism of OA-CHs, we conducted in vitro cell experiments by stimulating OA-CHs with OA-PBMCs.
Subsequently, the impact of lncRNA HOTAIR on the function of OA-CHs was assessed using CCK-8 assay. The results showed that the viability of OA-CHs was conspicuously decreased after overexpressing lncRNA HOTAIR. On the contrary, the cell viability was enhanced following the transfection of lncRNA HOTAIR silencing plasmids. Under the condition of lncRNA HOTAIR overexpression and silencing, inflammatory cytokines were further measured using ELISA method. The results displayed that overexpression of lncRNA HOTAIR significantly enhanced TNF-α level and reduced IL-10 level. In contrast, the deletion of lncRNA HOTAIR triggered an opposite tendency to inflammation. SiRNAs, as small non-coding RNA fragments, are one of RNA interference inducers for gene modulations, and the activity of inflammatory responses can be affected by using specific siRNAs to regulate the expression of particular inflammatory cytokines [38, 39]. These results suggested that si-lncRNA HOTAIR might be implicated in the occurrence and development of OA by orchestrating the levels of inflammatory factors. Consistently, lncRNA HOTAIR silencing has also been demonstrated to restrain cell viability via the PI3K/AKT pathway, reduce the cellular inflammatory response, inhibit drug resistance, and improve the quality of life of patients [40,41,42,43].
The effect of lncRNA HOTAIR on inflammatory factors may be related to its regulation of APN and its receptors, as well as the inflammatory pathway PTEN/PI3K/AKT. The current study noted the potential correlation between lncRNA HOTAIR and PTEN/PI3K/AKT pathway in OA-CHs through cell experiments. Western blot analysis was implemented to determine the influence of lncRNA HOTAIR on the pathway-related proteins, APN, and its receptors. The results manifested that overexpression of lncRNA HOTAIR elevated the expression of p-PI3K and p-AKT but reduced the expression of PTEN, APN, and ADIPOR1 proteins. The reason why overexpression of lncRNA HOTAIR had no significant effect on the expression of ADIPOR2 may be that ADIPOR1 is expressed abundantly in cartilages, bones, and synovial tissues, whereas ADIPOR2 is rarely detected [44]. Notably, baicalin as a natural flavonoid glycoside with significant anti-inflammation activity can inhibit the PI3K/AKT signaling pathway to induce the apoptosis and autophagy of human osteosarcoma cells [45]. Similarly, our subsequent cell experiments also confirmed that baicalin and lncRNA HOTAIR silencing exerted synergistic effects on the PTEN/PI3K/AKT signaling pathway inflammatory factors.
In summary, our obtained findings uncovered that lncRNA HOTAIR was highly expressed in the PBMCs of OA patients. LncRNA HOTAIR modulated the viability of OA-CHs and participated in inflammatory responses of OA by influencing the levels of inflammatory cytokines, APN, and the PTEN/PI3K/AKT pathway in OA-CHs, thus afflicting the occurrence and development of OA. These findings suggest the potential of lncRNA HOTAIR as a biomarker for OA treatment and provide novel insights into the pathogenesis of OA. Molecular docking analysis revealed that baicalin and si-lncRNA HOTAIR had synergistic effects on the PTEN/PI3K/AKT signaling pathway, APN, and inflammatory factors. However, whether baicalin alleviates immune inflammatory responses in OA patients by interfering the expression of lncRNA HOTAIR or the activation of the PTEN/PI3K/AKT signaling pathway remains to be further verified.
References
Tu J, Huang W, Zhang W, et al. The emerging role of lncRNAs in chondrocytes from osteoarthritis patients. Biomed Pharmacother. 2020;131:110642. https://doi.org/10.1016/j.biopha2020.110642.
Wang XD, Zhang JN, Gan YH, et al. Current understanding of pathogenesis and treatment of TMJ osteoarthritis. J Dent Res. 2015;94:666–73. https://doi.org/10.1177/0022034515574770.
Schram B, Orr R, Pope R, et al. Risk factors for development of lowerlimb osteoarthritis in physically demanding occupations: a narrativeumbrella review. J Occup Health. 2019;62(1):e12103. https://doi.org/10.1002/1348-9585.12103.
Arita Y, Kihara S, Ouchi N, et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 1999;257:79–83.
Cnop M, Havel PJ, Utzschneider KM, et al. Relationship of adiponectin to body fat distribution, insulin sensitivity and plasma lipoproteins: evidence for independent roles of age and sex. Diabetologia. 2003;46:459–69.
Chen TH, Chen L, Hsieh MS, et al. Evidence for a protective role for adiponectin in osteoarthritis. Biochim Biophys Acta. 2006;1762(8):711–8. https://doi.org/10.1016/j.bbadis.2006.06.008.
Larrañaga-Vera A, Lamuedra A, Pérez-Baos S, et al. Increased synovial lipodystrophy induced by high fat diet aggravates synovitis in experimental osteoarthritis. Arthritis Res Ther. 2017;19(1):264. https://doi.org/10.1186/s13075-017-1473-z.
Kim D, Song J, Han J, et al. Two non-coding RNAs, MicroRNA-101 and HOTTIP contribute cartilage integrity by epigenetic and homeotic regulation of integrin-α1 [published correction appears in Cell Signal. 2020 Jan; 65:109455]. Cell Signal. 2013;25(12):2878–887. doi:https://doi.org/10.1016/j.cellsig.2013.08.034
Van De Sande T, De Schrijver E, Heyns W, et al. Role of the phosphatidylinositol3’-kinase/PTEN/Akt kinase pathway in the overexpression of fatty acid synthase in LNCaP prostate cancer ce1ls. Cancer Res. 2002;62(3):642–6.
Wang H, Luo QF, Peng AF, et al. Positive feedback regula- tion between Akt phosphorylation and fatty acid synthase ex- pression in osteosarcoma. Int J Mol Med. 2014;33(3):633–9.
Wojdasiewicz P, Poniatowski ŁA, Szukiewicz D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014;2014:561459. https://doi.org/10.1155/2014/561459.
Bao B, Liu J, Wan L, et al. Nan Fang Yi Ke Da Xue Xue Bao. 2021;41(4):483–94. doi:https://doi.org/10.12122/j.issn.1673-4254.2021.04.02.
Wang J, Luo X, Cai S, et al. Blocking HOTAIR protects human chondrocytes against IL-1β-induced cell apoptosis, ECM degradation, inflammatory response and oxidative stress via regulating miR-222–3p/ADAM10 axis. Int Immunopharmacol. 2021;98:107903. https://doi.org/10.1016/j.intimp.2021.107903.
Chen C, Zhang C, Cai L, et al. Baicalin suppresses IL-1β-induced expression of inflammatory cytokines via blocking NF-κB in human osteoarthritis chondrocytes and shows protective effect in mice osteoarthritis models [published correction appears in Int Immunopharmacol. 2021 Jun;95:107546]. Int Immunopharmacol. 2017; 52:218–26. doi:https://doi.org/10.1016/j.intimp.2017.09.017
Yang X, Zhang Q, Gao Z, et al. Baicalin alleviates IL-1β-induced inflammatory injury via down-regulating miR-126 in chondrocytes. Biomed Pharmacother. 2018;99:184–90. https://doi.org/10.1016/j.biopha.2018.01.041.
Kolasinski SL, Neogi T, Hochberg MC, et al. 2019 American college of rheumatology/arthritis foundation guideline for the management of osteoarthritis of the hand, hip, and knee [published correction appears in Arthritis Care Res (Hoboken). 2021 May;73(5):764]. Arthritis Care Res (Hoboken). 2020;72(2):149–62. doi:https://doi.org/10.1002/acr.24131.
Shrestha B, Dunn L. The declaration of Helsinki on medical research involving human subjects: a review of seventh revision. J Nepal Health Res Counc. 2020;17:548–52.
Schneider A, Hommel G, Blettner M. Linear regression analysis: part 14 of a series on evaluation of scientific publications. Dtsch Arztebl Int. 2010;107(44):776–82. https://doi.org/10.3238/arztebl.2010.0776.
Martel-Pelletier J, Barr AJ, Cicuttini FM, et al. Osteoarthritis. Nat Rev Dis Primers. 2016;2:16072.
Di Paola R, Fusco R, Impellizzeri D, et al. Adelmidrol, in combination with hyaluronic acid, displays increased anti-inflammatory and analgesic effects against monosodium iodoacetateinduced osteoarthritis in rats. Arthritis Res Ther. 2016;18:291.
Sacitharan PK. Ageing and osteoarthritis. Subcell Biochem. 2019;91:123–59.
Yusuf E, Kortekaas MC, Watt I, et al. Do knee abnormalities visualised on MRI explain knee pain in knee osteoarthritis? Asystematic review. Ann Rheum Dis. 2011;70:60e7.
Berenbaum F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr Cartil. 2013;21:1621.
Brouwers H, von Hegedus J, Toes R, et al. Lipid mediators of inflammation in rheumatoid arthritis and osteoarthritis. Best Pract Res Clin Rheumatol. 2015;29(6):741–55. https://doi.org/10.1016/j.berh.2016.02.003.
Kabalyk MA, Kovalenko TS, Nevzorova VA, et al. Effect of arterial hypertension and hyperlipid emiaonre modeling of articul arcartilage and the development of osteoarthritis (experimental study). Adv Gerontol. 2019;32(4):550–7.
Ebrahim Hasnaa A, Alzamil Norah M, Bahjat A-A, et al. Suppression of knee joint osteoarthritis induced secondary to type 2 diabetes mellitus in rats by resveratrol: role of glycated haemoglobin and hyperlipidaemia and biomarkers of inflammation and oxidative stress. ArchPhysiol Biochem. 2020. https://doi.org/10.1080/13813455.2020.1771378.
Koskinen A, Juslin S, Nieminen R, et al. Adiponectin associates with markers of cartilage degradation in osteoarthritis and induces production of proinflammatory and catabolic factors through mitogen-activated protein kinase pathways. Arthritis Res Ther. 2011;13(6)
Kabalyk MA, Kovalenko TS, Nevzorova VA, et al. Effect of arterial hypertension and hyperlipidemia on remodeling of articular cartilage and the development of osteoarthritis (experimental study). Adv Gerontol. 2019;32(4):550–7.
Chen H, Qi J, Bi Q, et al. Expression profile of long noncoding RNA (HOTAIR) and its predicted target miR-17–3p in LPS-induced inflammatory injury in human articular chondrocyte C28/I2 cells. Int J Clin Exp Pathol. 2017;10(9):9146–57.
Luo X, Wang J, Wei X, et al. Knockdown of lncRNA MFI2-AS1 inhibits lipopolysaccharide-induced osteoarthritis progression by miR-130a-3p/TCF4. Life Sci. 2020;240:117019. https://doi.org/10.1016/j.lfs.2019.117019.
Yingchun C, Lin Z, Ernan L, et al. Long-chain non-coding RNA HOTAIR promotes the progression of osteoarthritis via sponging miR-20b/PTEN axis. Life Sci. 2020;253:117685. https://doi.org/10.1016/j.lfs.2020.117685.
Yang Y, Dan X, Yawei W, et al. A long non-coding RNA, HOTAIR, promotes cartilage degradation in osteoarthritis by inhibiting WIF-1 expression and activating Wnt pathway. BMC Mol Cell Biol. 2020;21:53. https://doi.org/10.1186/s12860-020-00299-6.
Marques-Rocha JL, Samblas M, et al. Noncoding RNAs, cytokines, and inflammation-relateddiseases. FASEBJ. 2015;29:3595–611. https://doi.org/10.1096/fj.14-260323.
Jiang M, Liu J, Luo T, et al. LncRNA PACER is down-regulated in osteoarthritis and regulates chondrocyte apoptosis and lncRNA HOTAIR expression. Biosci Rep. 2019;39(6):BSR20190404.
Liu N, Fu D, Yang J, et al. Asiatic acid attenuates hypertrophic and fibrotic differentiation of articular chondrocytes via AMPK/PI3K/AKT signaling pathway. Arthritis Res Ther. 2020;22(1):112. https://doi.org/10.1186/s13075-020-02193-0.
Li L, Liu F, Huang W, et al. Ricolinostat (ACY-1215) inhibits VEGF expression via PI3K/AKT pathway and promotes apoptosis in osteoarthritic osteoblasts. Biomed Pharmacother. 2019;118:109357. https://doi.org/10.1016/j.biopha.2019.109357.
Lee YA, Choi HM, Lee SH et al. Synergy between adiponectin and interleukin-1β on the expression of interleukin-6, interleukin-8, and cyclooxygenase-2 in fibroblast-like synoviocytes. Exp Mol Med. 2012;44(7).
Gargano G, Oliva F, Oliviero A, et al. Small interfering RNAs in the management of human rheumatoid arthritis. Br Med Bull. 2022;142(1):34–43. https://doi.org/10.1093/bmb/ldac012.
Gargano G, Oliviero A, Oliva F, et al. Small interfering RNAs in tendon homeostasis. Br Med Bull. 2021;138(1):58–67. https://doi.org/10.1093/bmb/ldaa040.
Li Z, Qian J, Li J, et al. Knockdown of lncRNA-HOTAIR downregulates the drug-resistance of breast cancer cells to doxorubicin via the PI3K/AKT/mTOR signaling pathway. Exp Ther Med. 2019;18(1):435–42. https://doi.org/10.3892/etm.2019.7629.
Qi K, Zhong J. LncRNA HOTAIR improves diabetic cardiomyopathy by increasing viability of cardiomyocytes through activation of the PI3K/Akt pathway. Exp Ther Med. 2018;16(6):4817–23. https://doi.org/10.3892/etm.2018.6755.
Zhang XH, Hu P, Xie YQ, et al. Long Noncoding RNA HOTAIR promotes endometrial carcinoma cell proliferation by binding to PTEN via the activating phosphatidylinositol 3-Kinase/Akt signaling pathway. Mol Cell Biol. 2019;39(23):e00251-e319. https://doi.org/10.1128/MCB.00251-19.
Liu G, Zhang W. Long non-coding RNA HOTAIR promotes UVB-induced apoptosis and inflammatory injury by up-regulation of PKR in keratinocytes [retracted in: Braz J Med Biol Res. 2021 Jun 14;54(6):e6896retraction]. Braz J Med Biol Res. 2018;51(8):e6896. Published 2018 Jun 11. doi:https://doi.org/10.1590/1414-431X20186896.
Chen T-H, Chen L, Hsieh M-S, et al. Evidence for a protective role for adiponectin in osteoarthritis. Biochim Biophys Acta. 2006;1762:711–8.
Pang H, Wu T, Peng Z, et al. Baicalin induces apoptosis and autophagy in human osteosarcoma cells by increasing ROS to inhibit PI3K/Akt/mTOR, ERK1/2 and β-catenin signaling pathways. J Bone Oncol. 2022;33:100415.
Acknowledgements
Not applicable.
Funding
This work was supported by Grants from Ministry of Science and Technology National Key Research and Development Program Chinese Medicine Modernization Research Key Project (2018YFC1705204); National Nature Fund Program (81973655); National Natural Science Foundation of China (82074373); Open Foundation of Key Laboratory of Xin'an Medical Ministry of Anhui University of Traditional Chinese Medicine (No.2020xayx10); The Key Research and Development Program Foreign Science and Technology Cooperation Project of Anhui (201904b11020011); Anhui Provincial Quality Engineering Teaching and Research Project (2018jyxm1068); Anhui Famous Traditional Chinese Medicine Liu Jian Studio Construction Project (Traditional Chinese Medicine Development Secret [2018] No. 11); National Key Innovative Talents Training Project of Traditional Chinese Medicine (National Education Letter of Traditional Chinese Medicine [2019] No.128); Key Research and Development Plan Project of Anhui Province (201904a07020004); Anhui Provincial Laboratory of Applied Basis and Development of Internal Medicine of Modern Traditional Chinese Medicine (2016080503B041); and 12th batch of "115" Innovation Team of Anhui Province (Anhui Talent Office [2019] No. 1).
Author information
Authors and Affiliations
Contributions
CXL and LJ contributed to the study design. CXL contributed to data analysis, wrote the first draft, and revised the manuscript. SYQ and WJT contributed to the specimen and data collection. ZXH, ZQ, and DX supervised the project and contributed to the manuscript revision. All authors reviewed and accepted the content of the final manuscript.
Corresponding author
Ethics declarations
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chen, X., Liu, J., Sun, Y. et al. Correlation analysis of differentially expressed long non-coding RNA HOTAIR with PTEN/PI3K/AKT pathway and inflammation in patients with osteoarthritis and the effect of baicalin intervention. J Orthop Surg Res 18, 34 (2023). https://doi.org/10.1186/s13018-023-03505-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13018-023-03505-1