
Abstract
Foot-and-mouth disease (FMD) is a contagious viral disease that affects the livelihoods and productivity of livestock farmers in endemic regions. It can infect various domestic and wild animals with cloven hooves and is caused by a virus belonging to the genus Aphthovirus and family Picornaviridae, which has seven different serotypes: A, O, C, SAT1, SAT2, SAT3, and Asia-1. This paper aims to provide a comprehensive overview of the molecular epidemiology, economic impact, diagnosis, and control measures of FMD in Ethiopia in comparison with the global situation. The genetic and antigenic diversity of FMD viruses requires a thorough understanding for developing and applying effective control strategies in endemic areas. FMD has direct and indirect economic consequences on animal production. In Ethiopia, FMD outbreaks have led to millions of USD losses due to the restriction or rejection of livestock products in the international market. Therefore, in endemic areas, disease control depends on vaccinations to prevent animals from developing clinical disease. However, in Ethiopia, due to the presence of diverse antigenic serotypes of FMD viruses, regular and extensive molecular investigation of new field isolates is necessary to perform vaccine-matching studies to evaluate the protective potential of the vaccine strain in the country.
Similar content being viewed by others


Introduction
Ethiopia has a large and diverse livestock population that contributes significantly to the national economy and the livelihoods of rural communities [1]. However, the livestock sector faces major challenges from transboundary animal diseases such as foot-and-mouth disease (FMD), which is a highly contagious viral disease that affects domestic and wild cloven-hoofed animals [2]. FMD causes severe economic losses due to reduced animal productivity, trade restrictions, and control costs. FMD is caused by a virus with seven serotypes (O, A, C, SAT 1, SAT 2, SAT 3, and Asia 1) [3] that vary in their genetic and antigenic characteristics [4]. The virus can infect cattle, sheep, goats, pigs, camels, and several wildlife species [5], causing fever, vesicles, erosions, salivation, lameness, and sometimes death [6].
The disease can spread rapidly through direct or indirect contact with infected animals or materials or through the air [4]. It is common in Africa, the Middle East and Asia, and some parts of South America. FMD-free countries without vaccination are always at risk of outbreaks. FMD prevention and control are costly and mostly borne by low- and lower middle-income countries [7]. The development of novel FMD control measures requires a better understanding of the molecular mechanisms of FMD virus replication and evolution [8]. In Ethiopia, FMD is a notifiable disease that has been reported since 1957 [9]. The disease affects cattle of all ages and breeds, as well as small ruminants and wildlife. The seroprevalence of FMD in Ethiopia ranges from 5.6 to 72.1% in cattle [10, 11] and from 4 to 11% in small ruminants [12]. Four serotypes (O, A, SAT 1, and SAT 2) are endemic in Ethiopia, with serotype O being the most dominant and widespread [13].
The epidemiology and evolution of FMD virus strains circulating in Ethiopia are poorly understood, limiting the effectiveness of control and prevention measures [11]. FMD outbreaks occur frequently across the country, with higher frequency and intensity during the dry season [14]. The occurrence and distribution of FMD outbreaks are influenced by various factors, such as production system, geographic location, species, age of animals, contact with wildlife, season of the year, mixed animal species, breed, and agroecology [15, 16].
New FMDV topotypes in Ethiopia challenge the livestock industry and need better understanding of the disease and its serotypes. This will help design effective control and prevention measures such as vaccination, rapid diagnostic tests (RDTs), and early warning systems [17]. FMD has significant impacts on livestock production and trade in Ethiopia due to reduced animal performance and welfare, market access barriers and consumer confidence, and vaccination costs. Therefore, the aim of this review is to provide a comprehensive overview of FMD with global up-to-date information on its epidemiology and current status in Ethiopia.
General features of FMD
FMD is a viral disease that affects various cloven-hoofed animals and causes huge economic losses in the livestock sector worldwide. It is caused by a virus from the genus Aphthovirus in the family Picornaviridae, which has a single-stranded RNA genome of approximately 8.5 kb [18,19,20]. The genome contains one open reading frame (ORF) surrounded by 5′ and 3′ untranslated regions (UTRs) and a poly-A tail at the 3′ end [21]. These regions play important roles in FMDV replication and translation by affecting viral RNA stability, translation efficiency, genome circularization, and interaction with cellular factors [22]. The RNA has two parts that are important for this explanation: the 5′UTR and the 3′UTR. The 5′UTR helps the virus to start making proteins, while the 3′UTR helps the virus to control how much protein is made. The 5′UTR and the 3′UTR can also interact with each other to regulate protein expression [23]. The poly-A tail of viral RNA interacts with cellular factors that affect its stability, translation initiation, and genome synthesis. These factors include Poly (A) binding proteins (PABPs), Exoribonucleases (ExoNs), and Poly (A) polymerases (PAPs). PABPs regulate the poly-A tail function and fate by modulating its recognition by host pattern recognition receptors (PRRs) and signaling molecules. ExoNs degrade RNA from the 3’ end and can alter the length and function of the poly-A tail. PAPs synthesize the poly-A tail by adding adenine nucleotides to the 3’ end of RNA and can influence the replication, transcription, and translation of viral RNA. The untranslated regions (UTRs) and the poly-A tail are essential for foot-and-mouth disease virus (FMDV) infection and pathogenesis [23,24,25, 31,32,33].
All the seven FMDV serotypes are differ in their antigenicity, virulence, and host range [34]. Antigenicity is the capacity of a molecule or an antigen to induce an immune response, that is to be recognized by and interact with an immunologically specific antibody or T-cell receptor [35]. Antigenicity depends on several factors, such as the antigen processing, presentation, and recognition by antigen-presenting cells (APCs), the B-cell receptor (BCR) epitope interactions, and the affinity and avidity of the antibody-antigen binding. Antigenicity also determines the memory and protection of the immune system on re-encounter with the same antigen [36]. Serotype C has not been detected since 2004. The VP1 protein of FMDV contains the major B-cell epitope, which is the part of the antigen that is recognized by antibodies and induces protective immunity. VP1 also shows high variability and phylogenetic clustering among FMDV strains. Therefore, sequencing and classification of FMDV strains are based on the VP1 gene, which helps to track their evolution and spread [4, 26,27,28,29,30].
The capsid of FMDV is made up of 60 copies of four structural proteins, called VP1, VP2, VP3, and VP4. These proteins are also known as 1A-1D. The capsid has a symmetrical shape with 20 triangular faces (Fig. 1) [37]. The capsid has an icosahedral shape with 20 faces that are each triangular. The proteins VP1, VP2, and VP3 are exposed on the capsid surface, while VP4 is internal and contributes to the structural integrity. The capsid proteins originate from a precursor protein called P1-2A, which is cleaved by a viral protease named 3Cpro into smaller fragments [37, 38]. The capsid proteins can self-assemble into empty shells that resemble the virus but lack any genetic material. These empty shells can serve as vaccines to induce immunity against FMDV in animals [37].

FMDV structure (icosahedral capsid structure), adapted from https://viralzone.expasy.org/98. (Accessed on 17 July 2022)
VP-1 is a multifunctional structural protein that mediates host-cell attachment and elicits immune responses and cell apoptosis [39]. It is the most variable region of the capsid gene and contains highly immunogenic and neutralizing sites [38]. Viral protein 2 (VP2) is a structural protein that contributes to the stability and maturation of the FMDV capsid. It is relatively conserved among FMDV serotypes, with 47% of its amino acids being identical across and within serotypes [20]. However, the extent of conservation varies depending on the region of VP2. The N-terminal end is highly conserved, with 67.4% invariant amino acids, while the C-terminal end is more variable. VP2 also has some antigenic and surface-exposed sites that are conserved among serotypes and may be important for vaccine development [40].
The capsid is composed of four structural proteins: VP1, VP2, VP3, and VP4. Among them, viral protein-1 (VP1) is the most important (major antigenic site of FMDV) for determining the serotype and the immune response of the host. VP4 is the most conserved protein (nucleotide sequence that is 73% to 84% conserved across all FMDV isolates) and has a T-cell epitope for swine [20]. The variability of the capsid proteins is influenced by selective pressures from the host immune system, cell culture adaptation, and vaccine escape [41]. The antigenic sites of FMDV are located on the surface-exposed loops of VP1-3, especially the βG–βH loop in VP1 [42].
The open reading frame (ORF) encodes four structural proteins (VP1, VP2, VP3, and VP4) that form the viral capsid and eight nonstructural proteins (NSPs) (Lpro, 2A, 2B, 2C, 3A, 3B, 3Cpro, and 3Dpol) that help the virus replicate and evade host defenses [25] (Fig. 2). These viral proteins play various roles in FMDV pathogenicity and can inhibit the functions of various host proteins to counteract host antiviral responses [39]. The mutation of the H138 residue in Lpro, an NSP that inhibits host translation, reduces FMDV virulence in vitro and in vivo [43].

The RNA strand of FMDV and its protein-coding parts. The RNA strand has a 5′UTR, an ORF with L, VP4, VP2, VP3, VP1, 2A, 2B, 2C, 3A, 3B (3B1, 3B2, and 3B3), 3C, and 3D parts, and a 3′UTR.; source: [25]
Leader proteinase (Lpro) is the first mature viral protein in FMDV and a key virulence factor that inhibits host cell translation. FMDV Lpro is a viral enzyme that belongs to the papain-like protease family. It cuts the viral polyprotein into smaller proteins and targets some host proteins. This affects the ability of the host cell to make its own proteins and activate its immune system against the virus [43]. FMDV has a genome composed of a single strand of RNA without an envelope surrounded by a protein shell that is not symmetrical [44]. The virus makes mistakes when copying its RNA, leading to high variation among the different serotypes of the virus, which have approximately 86% similarity and cause repeated outbreaks of the disease [45]. Therefore, FMDV serotypes comprise antigenically distinct strains and topotypes, leading to partial cross-protection within serotypes [46].
Morbidity and mortality rate
FMDV is a highly contagious virus that causes FMD, a viral disease that infects various cloven-hoofed animals. FMD can impair the health and productivity of these animals and result in huge economic losses worldwide. The disease is endemic in more than 100 countries, mostly in Africa, Asia, and South America [2]. FMD is classified as the most dangerous animal disease on list A by the World Organization for Animal Health (WOAH), but it does not affect humans. The disease has an incubation period of 2–14 days. The disease is rarely fatal in adult animals (1–5%), but high in young calves, lambs, and piglets (20% or more) [47]. However, the morbidity and mortality rates of FMDV depend on various factors, such as the animal species, breed, production type, age, immunity, virus dose, and animal movement [48]. In a fully susceptible livestock population, the morbidity rate of FMDV can be as high as 100% with a high mortality rate in young animals due to myocarditis [47]. In Ethiopia, FMDV is endemic, with different serotypes circulating at different prevalence levels [2].
The disease impairs the performance and productivity of livestock, especially cattle, which are vital for many farmers' income and food security. A recent study found that FMDV morbidity rates in cattle herds were 68.1% in mixed crop-livestock systems and 54.5% in commercial dairy farms. Another study reported a lower rate of 38.9% during major outbreaks [49]. FMDV infection causes low mortality (0.4%) in cattle and high morbidity (35.7%) in sheep and goats, which can also be asymptomatic carriers of the virus. The herd-level morbidity of FMDV is higher in cattle (57.2%) than in small ruminants (8%) in mixed systems [49]. Most animals recover from FMDV infection within 2 weeks if there are no complications [47]. Sheep and goats are less susceptible to FMDV infection than cattle, but they can still transmit the virus in mixed farming systems [50].
Pathogenesis of FMD in animals
FMDV infection causes different clinical signs, pathogenesis, and immune responses depending on the host and serotype [51]. The virus can enter the host cell by binding to cellular receptors through its capsid proteins. The virus makes copies of itself in the pharyngeal mucosa and travels through the lymphatic and blood systems to other epithelial tissues in the mouth, feet, mammary glands, and skin [52]. The virus can be found in various body fluids before and after the symptoms of FMD appear. The virus can also stay in the oral cavity of infected animals for a long time. The host antiviral system quickly responds to stop virus replication and clear the virus. However, viruses evade host antiviral responses and affect host resistance to other pathogens [34].
FMDV has evolved different immune escape mechanisms to overcome the host immune response in long-term coevolution. These mechanisms can be classified into three types: (1) changing or masking the antigenic structure of the viral surface to avoid recognition by the humoral immune response; (2) inhibiting the expression or activity of cytokines, chemokines, or major histocompatibility complex (MHC) molecules to interfere with the function of the cellular immune response; and (3) inducing immunosuppression, apoptosis, or tolerance to interfere with the host’s immune response to the virus [53].
The epidemiological features of FMDV
Host range
Foot-and-mouth disease virus (FMDV) is a virus that causes FMD, a viral disease that affects animals with split hooves, such as cattle, sheep, goats, pigs, and many wild ruminants. Cattle are the main hosts for FMDV and the most vulnerable to infection. However, some FMDV strains (serotype O) can adapt to pigs after being isolated in cell culture. The role of small ruminants in FMDV transmission is unclear, as it is unknown whether the virus can stay in these animals for long periods without infecting cattle. However, FMDV strains that infect cattle have also been found in wild pigs, antelopes, and deer [54]. Therefore, these hosts have clinical and epidemiological importance in natural infection. Other susceptible species (deer, antelope, wild pigs, elephants, and giraffes) are infected incidentally or accidentally and have little or no epidemiological significance [55]. Horses are resistant to FMD, but the mechanism of resistance is unknown [56]. FMD does not harm human health or food safety, but only healthy animals should be used for consumption [57]. However, FMD can devastate the livelihoods and food security of poor communities, as it reduces animal productivity and income [58].
FMDV transmission and risk factors across species and regions
FMD is mainly transmitted by inhaling virus particles from the breath of infected animals. However, the disease can also spread through the air under certain conditions. The virus can travel through the air under favorable conditions and infect animals outside quarantine zones, making control measures difficult [59]. Cattle are mostly infected by the respiratory or airborne route, while pigs can be infected by the digestive route through contaminated food, water, or fomites [60].
The virus can also spread indirectly through the environment if it is contaminated by infected animals. The virus can survive for a long time under favorable conditions, such as temperatures below 50 °C, relative humidity above 55%, and neutral pH [61, 62]. This makes FMD epidemiology and control more complex, as the virus can have multiple sources of infection. The excretions and secretions of infected animals can transmit the disease and serve as noninvasive samples for diagnosis and surveillance [62].
The occurrence of FMD outbreaks in Ethiopia is influenced by various factors, such as production system, geographic location, species, age of animals, contact with wildlife, season of the year, mixed animal species, and breed [2, 17]. FMD outbreaks are more prevalent in market-oriented systems than in subsistence systems due to frequent movement and mixing of animals [63]. The central, southern, and southeastern regions have a higher incidence of outbreaks than the northern and western regions, which may be related to wildlife reservoirs, climatic factors, or livestock density [63]. Younger animals are more prone to FMD infection than older animals, as they have lower immunity and higher virus shedding. They also tend to show more severe clinical signs and higher mortality rates [12, 14].
Contact with wildlife is another risk factor for FMD occurrence [15], although the only confirmed wildlife reservoir is the African buffalo, Syncerus caffer [7]. Wildlife can also be affected by FMD outbreaks and act as bridge or maintenance hosts for disease transmission. Livestock and wildlife can come into contact by sharing the same grazing or watering areas or breaking through fences [12, 64]. This increases the risk of FMD outbreaks, especially in the dry season when water and pasture are scarce. The virus can also survive better in the environment during the dry season [14].
Keeping different species of livestock together can increase the risk of FMD transmission, as different species may have different susceptibilities and immunity to the virus. For example, sheep and goats can act as silent carriers of FMD and infect cattle without showing clinical signs. Some breeds of cattle are more resistant to FMD than others due to genetic factors that modulate their immune response to the virus. For instance, Borana cattle in Ethiopia are less likely to become infected and show less severe symptoms than other breeds [14]. FMD virus poses challenges for diagnosis, surveillance, and vaccination because it has many genetic variants. The disease also affects the livelihoods and incomes of livestock farmers and traders in developing countries, where livestock are important for food security and social status. Therefore, the socioeconomic factors that influence the attitudes and knowledge of livestock stakeholders are crucial for preventing and controlling FMD.
Historical background
Frosch and Loeffler discovered FMDV as the cause of FMD in 1897, paving the way for virology. Fracastorius described the symptoms of FMD in cattle in 1514 [65]. His description matched the disease seen in Germany (1751) and the UK (1839). The disease spread beyond New Zealand and is still common in parts of Asia, Africa, and South America [66]. The virus also diversified into seven serotypes and originated from the Mediterranean region before reaching Europe, Asia, and South America [27, 67].
Global distribution
FMDV may have emerged in the Middle Ages and spread from the Mediterranean area to other parts of the world through trade and migration. FMDV can survive for long periods in favorable conditions. FMDV has seven serotypes that vary in their molecular characteristics and evolutionary relationships. FMDV also undergoes frequent recombination events that increase its genetic variation and pose challenges for vaccine development and selection [68]. Most of the World’s livestock (77%) is affected by FMD, especially in Africa, the Middle East, Asia, and some parts of South America. The disease can spread to countries that are FMD-free despite vaccination, as some vaccinated animals may still be susceptible to infection or become carriers of the virus. For example, in 2019, an outbreak of FMD occurred in Mongolia, which had been FMD-free since 2017 and had implemented a vaccination program. The outbreak was caused by a new strain of FMDV that was not covered by the vaccine [7]. FMD prevention and control costs are mostly (75%) borne by low- and lower-middle-income countries. These costs are highest in Africa and Eurasia, which account for half and a third of the global total, respectively [7].
The official data show that China has experienced 140 FMD outbreaks since 2010. China has reported a total of 109 FMD outbreaks from 2010 to 2019, affecting cattle, pigs, sheep and goats in 24 provinces [69, 70]. The most recent event occurred in October 2020, affecting 76 cattle in Heshuo County, Xinjiang Uyghur Autonomous Region [71]. Moreover, the common practice of small-scale farming and grazing in these regions could also increase the risk of FMD transmission [71].
Brazil and Kenya are the last countries where serotype C of FMD virus was detected in 2004. Serotype C has been considered extinct since then [65]. The United Kingdom experienced a major FMD outbreak in 2001 that resulted in the slaughter of over six million animals and an estimated cost of £8 billion [72]. India is one of the countries with the highest economic impact of FMD due to its large livestock population and low vaccination coverage. The annual losses due to FMD in India are estimated at USD$4.5 billion [73].
In 2012, the FAO and OIE launched a 15-year global strategy to control FMD. The strategy aims to mitigate the adverse effects of FMD on animal health and welfare, food security and economic development. The strategy involves enhancing the capabilities of veterinary services and adopting progressive control measures informed by risk assessment and surveillance [74]. Early detection, warning systems and surveillance are crucial for preventing the disease. The WOAH maintains an official list of FMD-free countries that can be recognized as free of the disease in their whole territory or in specific zones and compartments (Fig. 3). FMD is a viral disease of livestock that can spread across national borders and infect many animals. Most countries in Latin America have achieved FMD-free status by implementing zoning and vaccination strategies. FMD is absent from some other regions of the world, but it can still emerge sporadically [7].

Map of FMD official status; adapted from https://www.woah.org/app/uploads/2023/03/fmd-world-eng-2023.jpg (accessed on 17 July 2022)
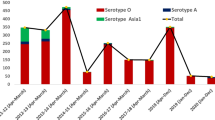
FMDV has seven serotypes: O, A, C, Asia 1, SAT 1, SAT 2, and SAT 3. These serotypes have different sub-lineages or topotypes based on the genetic and antigenic diversity of the capsid genes, especially the VP1 region. The VP1 region can vary by 30–50% between serotypes, while the overall similarity between serotypes is approximately 86%. FMDV is constantly evolving in the field, producing new strains that can cause outbreaks and spread to new areas. FMDV serotypes and their sub-lineages or topotypes are classified into seven virus pools based on their geographic and characteristic features (Fig. 4) [68]. These pools are distributed across Europe, the Middle East, Asia, Africa, and the Americas [68, 80].

Endemic serotypes of FMDV in different pools; adapted from https://www.woah.org/app/uploads/2021/05/fmd-world-eng.png (accessed on 17 August 2023). Source [68]
Viral protein1 (VP1) is the most important structural protein of FMDV, as it determines the serotype and the immune response of the host. VP1 forms part of the capsid, the outer layer of the virus that protects its genetic material and interacts with the host cell receptors. VP1 is the most exposed and variable protein on the surface of the capsid, and it contains the major antigenic site of FMDV, which is recognized by the host’s antibodies. VP1 is also involved in the attachment of FMDV to the host cell receptors, such as integrins and heparan sulfate proteoglycans. VP1 can change its shape and antigenicity to escape from the host’s immune response and infect new cells [77, 78].
Viral protein1 (VP1) is the most important target for vaccine development and selection against FMDV. Vaccines are substances that stimulate the immune system to produce antibodies against a specific pathogen, without causing the disease. Vaccines can protect the host from future infections by the same or similar strains of the pathogen. However, due to the high variability of FMDV, especially in the VP1 region, the vaccines need to match the circulating strains of the virus to be effective. Therefore, it is essential to monitor the evolution and circulation of FMDV serotypes and their sub-lineages or topotypes and to update the vaccines accordingly [79].
FMDV serotypes vary in their geographic distribution and impact. Serotypes O, A, and C can cause outbreaks in Europe, America, Africa, and Asia. Serotypes SAT 1–3 and Asia-1 are restricted to Africa and Asia, respectively [75]. Serotype O is the most common in Africa, except for Southern Africa. Serotype O is also the dominant serotype in Ethiopia [76]. Serotype C has not been detected since 2004 in Brazil and Kenya and is considered extinct [27]. Therefore, the evolution and circulation of viruses within different geographic pools help to assess the suitability of vaccines [17].
Current status of FMD in Ethiopia
FMD was first detected in Ethiopia in 1957, when serotypes O and C were isolated from cattle [9]. Since then, other serotypes, such as A, SAT 1, and SAT 2, have been reported in different regions of the country, with serotype O being the most dominant and serotype C being absent since 1983 [3, 81]. FMD is endemic and widely prevalent in Ethiopia, affecting various livestock species and causing economic losses [82]. A recent study conducted by Woldemariyam, et al. [83] revealed the spatiotemporal trends and hotspots of FMD outbreaks in Ethiopia over 10 years. They observed that the dry season (October–March) was associated with a higher frequency and intensity of outbreaks, with a peak in March 2012. They also noted a decreasing trend in outbreaks over time, but with a potential duration of up to 5 years. They identified four dominant serotypes (O, A, SAT-2, and SAT-1), but only 12% of outbreaks had a known serotype. According to Ayelet, et al. [13], FMD was more prevalent in the central and southern parts of Ethiopia, indicating that the disease was endemic and had multiple serotypes.
[84] reported that serotypes A and SAT 2 were first detected in 1969 and 1989, respectively, and that serotypes O and SAT 2 caused an outbreak between 1988 and 1991. Only one outbreak of serotype SAT 1 was studied from 2007 to 2008 [85]. Asfaw and Sintaro [86] noted that FMD outbreaks had become more frequent since 1990 and were not effectively controlled due to the lack of regular vaccination, except for some dairy animals. This led to substantial economic losses for farmers and the country due to trade restrictions on livestock and livestock products [87]. FMD control in Ethiopia faces major challenges due to the presence of multiple serotypes and subtypes of the virus, the diversity of host animals, the unregulated movement of livestock, and the shortage of effective and affordable FMD vaccines in the country. Therefore, a long-term progressive risk reduction approach for FMD control is recommended for endemic countries such as Ethiopia [79, 88].
Sero-prevalence of FMD
Table 1 shows the FMD seroprevalence in Ethiopia from 2008 to 2021 based on different studies. The FMD seroprevalence in Ethiopia varies by location, from 5.6% to 42.7% in cattle, 4% to 11% in small ruminants, and 30% in ungulate wildlife [12]. In the Borena zone, 42.7% of 363 cattle sera samples were positive for FMD antibody using the 3ABC-ELISA test [82]. The highest prevalence was in Dire district (52.8%). However, in North and South Gondar, the FMD seroprevalence in cattle was lower (14.9%) [89]. This could be due to more FMD outbreaks during a period of major socioeconomic and political crisis (Guerrini et al., 2019). In the Afar region, Dubie and Negash [15] found that 19.8% of animals and 56.94% of herds were FMD seropositive, based on 384 sera samples. Shurbe, et al. [16] also reported that 26.8% of cattle in the Gamo zone had FMD antibodies. In northern Amhara, the FMD seroprevalence in cattle was low (3.4%) [90], which was similar to the 3.1% reported by Gezahegn, et al. [91].
According to Bahiru and Assefa [90], a study conducted in northern Amhara, Ethiopia, found that the overall seroprevalence of FMD was 3.4%, with no significant association with risk factors such as age, sex, district, and body condition score of the cattle. The prevalence was lower than other parts of the country. This result was much lower than the 10.88% prevalence reported in Jijiga [92] and the Amhara region [93], which could be due to different animal management practices, study years, and environmental factors [90].
A cross-sectional study around Addis Ababa found that the FMD seroprevalence among dairy cattle was high (72.1%). The highest seroprevalence was in the Ada Berga, Holeta, and Sululta districts (97.2%, 71.4%, and 57.6%, respectively). This study also found some risk factors for FMD that were statistically significant and detected FMDV serotype O from outbreak cases [11]. However, lower seroprevalence was reported in the West Shewa Zone (40.4%) [94] and in feedlot cattle in Meki (Oromia) (37.4%) [95].
The prevalence of FMD varies by region and country due to different factors, such as sampling method, study design, virus characteristics, and local conditions [16]. Some of these factors are agroecology, wildlife contact, animal movement, grazing and watering practices, age, body condition, management system, vaccination status, vaccine quality, diagnostic capacity, and livestock species interaction [15, 16]. Awel, et al. [11] also reported that age, body condition, and management system of the animals affected FMD seroprevalence in Ethiopia. They suggested that the high seroprevalence in some areas could be related to the diversity of FMDV subtypes and/or topotypes and the low FMD vaccination coverage in those areas.
Several studies have also reported differences in FMD seroprevalence among cattle based on sex, age, breed, and agroecological zone. Jenbere, et al. [100] reported higher seroconversion rates in male (15.7%) than female (8.3%) animals, while Mesfine, et al. [101] reported the opposite trend (8.9% in females and 3.0% in males). Desissa, et al. [10] reported higher FMD seroprevalence in female (27.17%) than male (15.34%) animals in the Kellem Wollega Zone of Ethiopia. They also noted that older cattle (> 4 years) had the highest seroprevalence (24.22%), followed by middle-aged (2–4 years) and young (< 2 years) cattle. Shurbe, et al. [16] reported that in the Gomo zone of southern Ethiopia, FMD seropositivity was higher among adult, local, and lowland cattle than among young, crossbred, and highland cattle. However, this finding was contradicted by G, et al. [102], who asserted that crossbred and productive cattle were more susceptible to FMD than local cattle. They recommended restricting animal movement and contact to reduce FMDV antibody levels in stationary systems [103].FMD seroprevalence in Ethiopia was lower than in other countries in the region, such as Kenya and Eritrea, where higher rates were observed using different diagnostic methods [104,105,106]. They suggested conducting more comprehensive and coordinated seroprevalence studies in Ethiopia to elucidate the molecular epidemiology of FMDV and devise effective control strategies. FMD outbreaks in Ethiopia have increased over time, especially in the Oromia and Amhara regions, where the highest number of outbreaks occurred in 2004, 2007/2008, and 2011/2012 [13, 17, 103]. The high FMD incidence in some regions may be attributed to various factors, such as susceptible animals, animal movement, market contact, and shared resources [76].
Risk factors associated with FMD infection and transmission
FMD infection and transmission can be affected by various factors related to the virus, the host, and the environment. Viral factors include the serotype, genotype, virulence, antigenic variation, and persistence of FMDV in the environment or in carrier animals. Host factors include the species, breed, age, immune status, susceptibility, and density of susceptible animals [7, 107]. The environmental factors include the climate, season, geography, vegetation, wildlife reservoirs, and human activities. Some of the human activities that can raise the risk of FMD infection and transmission are unregulated or illicit movement of animals or animal products across borders or within regions [108, 109]. Mixing of animals at water points, grazing areas, markets, or slaughterhouses; contact with infected wild animals or their products; insufficient biosecurity measures at farms, transport vehicles, or slaughter facilities; and low effectiveness or coverage of vaccination programs [107, 109].
Factors such as production system, geographic location, wildlife contact, seasonality, animal species, and breed influence FMD outbreaks in Ethiopia [2, 17]. FMD incidence is higher in market-oriented systems and central, southern, and southeastern regions than in subsistence systems and northern and western regions [63]. Young animals are more susceptible to FMD than older animals [12, 14]. Wildlife contact can occur through shared resources or fence breaking and can increase FMD transmission risk. The African buffalo, Syncerus caffer, is the only confirmed wildlife reservoir of FMD [7]. FMD outbreaks are more frequent during the dry season than during the wet season [14]. Mixing of different livestock species can enhance FMD transmission [15], as they may have different levels of susceptibility and immunity to the virus. FMD resistance varies among cattle breeds [14]. The genetic diversity of FMDV challenges its diagnosis, surveillance, and vaccination. FMD also has severe socioeconomic impacts on livestock stakeholders in developing countries. Socioeconomic factors affect the prevention and control of FMD by influencing the behavior and awareness of livestock owners and traders.
Molecular epidemiology of FMDV
Molecular epidemiology is essential for understanding FMDV distribution and disease situation and for developing effective control strategies. It uses genetic sequences of FMDV strains, particularly the VP1 gene that encodes the major capsid protein and antigenic determinant of the virus, to infer their origin, spread, and relatedness. VP1 can also help trace virus diffusion associated with animal movements, interspecies transmission events, transcontinental introductions, and the spatiotemporal distribution of viruses involved in FMD outbreaks [110]. However, whole-genome sequencing of FMDVs that cause subclinical infections has been reported in few studies [111], which is a better tool for tracing transmission within and between herds [112].
Molecular epidemiology can also track the emergence and distribution of new FMDV variants that may threaten animal health and trade [26, 113]. The main antigenic sites of the FMDV capsid have been characterized to trace virus strains and their transmission patterns [113]. FMD occurrences are also influenced by interspecies transmission, livestock movements (both formal and informal), and geographic relatedness among FMD virus isolates. Whole genome sequencing can help distinguish between closely related viruses and reconstruct transmission pathways between farms within outbreaks [114]. The high rate of replication errors in the FMD viral RNA replication process leads to high genetic diversity among virus serotypes, which share approximately 86% homology [115]. VP1 is a variable protein that contains the major immunogenic epitopes of FMDV. It varies by 30–50% among the seven serotypes. It is important for FMDV molecular epidemiology research worldwide [110].
Phylogenetic analyses of VP1 nucleotide sequences can reveal variation, genetic relationships, and geographical distributions among different FMDV serotypes [116]. Therefore, the VP1 genomic region is useful for molecular diversity analyses that determine lineages and topotypes. However, full genome sequences can provide more insights into the evolution and virulence of cocirculating strains [117], as they have serotype-specific amino acids that allow for serotype differentiation [118]. The complete genome sequence of FMDV, especially the ORF region, can reveal the amino acid changes that affect the serotype-specific immunogenicity, antigenicity, disease outcome, and transmission of the virus [119]. Similarly, molecular analysis of FMDV capsid proteins can provide detailed information on the antigenic and genomic characteristics of the virus serotype [120]. The presence of a high FMD-susceptible population, vaccine- and infection-induced partial antibody responses in some areas, and animal movement facilitate the establishment and spread of FMDV genetic lineages [121].
The VP1 region of the virus is commonly used for phylogenetic analysis to study the origin and spread of FMD in different regions of the world [110]. Understanding the outbreak pattern and epidemiology is essential for effective control strategies [17]. In Ethiopia, five FMDV serotypes (except Asia-1 and SAT3) were isolated from 1974 to 2007 [99]. Serotypes A and O are the most common and have a high impact on livestock production in Ethiopia [76]. Full genome sequences can provide more insights into the evolution and virulence of cocirculating strains [117], as they have serotype-specific amino acids that enable serotype differentiation [118]. Molecular analysis of the entire virus genome, especially the ORF region, can detect the amino acid sequence changes that indicate FMDV-specific serotype immunogenicity, antigenicity, disease prognosis, and spread [122].
Moreover, molecular examination of FMDV viral capsid proteins can provide comprehensive data on the antigenic and genomic determinants of the virus serotype [120]. The presence of a high FMD-susceptible population, vaccine- and infection-induced partial antibody responses in some areas, and animal movement facilitate the establishment and spread of FMDV genetic lineages [121]. FMDV serotypes A, O and SAT 2 caused most FMD outbreaks in Ethiopia [99]. However, in 2018, FMD outbreaks caused by serotypes O and A were reported, but genomic analysis showed that serotype A was the predominant serotype in the study areas [3], contrary to previous reports that serotype O was the most common, followed by serotype A [13, 123]. One sample had a coinfection of both serotypes. Serotypes O and A were further classified as East Africa and Africa topotypes of genotype IV, respectively [3].
FMDV serotype O is the most prevalent and widespread serotype in Asia, where it causes frequent outbreaks of FMD in susceptible animals. This serotype has 11 topotypes, which are groups of viruses that share genetic similarities in the VP1 gene. The VP1 gene encodes the major capsid protein and antigenic determinant of the virus. The topotypes are named after their geographic regions: Europe-South America (Euro-SA), Middle East-South Asia (ME-SA), Southeast Asia (SEA), Cathay (CHY), West Africa (WA), East Africa 1 (EA-1), East Africa 2 (EA-2), East Africa 3 (EA-3), East Africa 4 (EA-4), Indonesia-1 (ISA-1), and Indonesia-2 (ISA-2) [124, 125].
FMDV serotype Asia 1 has seven genotypes: I to VII. The topotypes and genotypes differ in their geographic distribution, antigenicity, virulence, and host range. In Asia, the dominant topotypes of FMDV serotype O are CHY, ME-SA, and SEA, while the dominant topotype of FMDV serotype Asia 1 is Cathay [126]. The ME-SA topotype viruses were mainly found in Egypt and Libya, reflecting their trade links with the Middle East [125]. The EA topotypes (EA-1 to EA-4) are dominant in Africa. Serotypes A and C were grouped into their African topotypes. Type A was similar to the serotypes from Egypt and Kenya. However, the type C viruses were distinct from but related to the Kenyan vaccine strain (K267/67). Serotype SAT2 topotype outbreaks (SAT2/XIV, SAT2/XIII, and SAT2/IV) were reported in Ethiopia, Sudan, and Kenya, respectively [123].
Phylogenetic analysis of FMDV VP1 coding region sequences from 2008 to 2017 revealed seven different FMD viral clades in Ethiopia: O/East Africa-3 (EA-3), O/East Africa-4 (EA-4), A/AFRICA/G-I, A/AFRICA/G-IV, A/AFRICA/G-VII, SAT2/VII, and SAT2/XIII (Fig. 5) [81]. These strains were related to other East African FMDV strains but also showed some genetic variation within and among serotypes. Molecular characterization of FMDV strains can help monitor the disease epidemiology and select suitable vaccines for FMD control in Ethiopia. However, only FMDV serotype SAT2 (classified into two topotypes: SAT2/VII and SAT2/XIII) was detected for the first time in the three Afar regions in a 12-year retrospective study, while no other FMDV serotypes were detected in this area [15].

FMDV isolates acquired in Ethiopia from 2008 to 2019 were distributed across the country. The map was adapted from https://onlinelibrary.wiley.com/doi/10.1111/tbed.13675
Serotype C of FMDV was first detected in Europe in the 1920s and then spread to other continents. It was less prevalent and less severe than other serotypes. The last outbreaks of serotype C occurred in Brazil and Kenya in 2004 [27]. In Ethiopia, serotype C emerged in 1957 along with serotype O [96] and remained endemic until 1983, coexisting with serotypes O, A, and SAT1 [127]. Paton, et al. [27] reported that no cases of FMDV serotype C have been detected globally since 2004. Therefore, they suggest stopping the production, testing, and use of serotype C vaccines and stocks in worldwide repositories. They also advise restricting the in vitro handling of the virus to situations where risk assessment supports it and biosecurity measures are stringent. Furthermore, they encourage researchers to join global monitoring efforts for serotype C [27].
Molecular epidemiology showed that most of the FMDV type O isolates from Egypt, Eritrea, and Sudan belong to the EA-3 topotype. This topotype was also detected in Ethiopia every year from 2008 to 2019, except for 2016. The O/EA-4 topotype was mostly found in the central and southwest regions of Oromia [81]. It was first detected in Ethiopia in 2005 and has largely remained there, with only a few outbreaks in Kenya in 2010. Currently, EA-2 and EA-3 are the main topotypes responsible for most of the serotype O outbreaks in the region [128]. Serotype A was detected sporadically from 2008 to 2019 and belonged to three different genotypes: A/AFRICA/G-I, A/AFRICA/G-IV, and A/AFRICA/G-VII. Serotype SAT2 was also detected intermittently from 2009 to 2018, belonging to two different topotypes: SAT2/VII and SAT2/XIII [81]. The CHY topotype of FMDV serotype O and the Cathay topotype of FMDV serotype Asia 1 have higher evolutionary rates than the other topotypes, which may indicate their adaptation to different hosts and environments [126].
The economic consequences of FMD
Global impact of FMD
FMD outbreaks affect the entire cattle agribusiness chain, resulting in (1) a reduced cattle population, (2) increased imports of cattle and beef, (3) decreased beef consumption, and (4) huge economic losses [55, 129]. FMD also has a severe social impact on farmers, who may suffer from illness, stress, depression, stroke, divorce, or even suicide. FMD outbreaks in FMD-free countries or zones trigger strict and costly control measures, such as culling, movement restrictions, and trade bans, until the disease is eradicated, and the FMD-free status is regained. For instance, FMD outbreaks in the United Kingdom, Japan, and South Korea have shown the vulnerability of even advanced biosecurity systems to FMD introduction and spread and the consequent losses in domestic and export markets [132, 134]. FMD also hampers the trade potential of low- and middle-income countries, where most of the world's cattle population resides but only a small fraction of global livestock exports originate. For example, Uganda’s endemic FMD situation prevents it from accessing lucrative export markets [133]. Zambia faces an annual loss of $1.6 billion in exports of beef and sable antelopes due to FMD-related import bans from Botswana and South Africa [134].
FMD control requires cooperation and public investment in veterinary services and surveillance systems within and between countries, which can also help control other livestock diseases. The Global FMD Control Strategy aims to help poor countries and animal trade by enhancing FMD control in endemic regions and safeguarding FMD-free regions [74]. FMD control can reduce the direct effects of the disease on animal production and productivity, which can cost USD 6.5–21 billion a year in endemic regions [73, 135]. It can also mitigate the indirect effects of the disease on market access, technology adoption, food security, income, and livelihoods of livestock-dependent households, particularly in low-income countries [55, 73]. Moreover, FMD control can prevent or minimize export losses due to trade restrictions on livestock and livestock products, which can constitute the majority of the total costs of an FMD outbreak, depending on the country and the control measures implemented [136].
Economic losses of FMD in Ethiopia
FMD is a widespread and devastating disease in Ethiopia that affects the well-being and income of livestock farmers and other actors in the sector [53]. The disease not only reduces animal productivity by causing lower milk production, reproductive problems, calf mortality, and early culling but also limits market access and tourism opportunities due to trade barriers, movement restrictions, and negative perceptions [49]. The country lost more than 14 million USD as a result of the export ban between 2005 and 2006 [137]. Similarly, an economic loss of 3,322,269 USD was reported in 2011 due to bull export rejection from an international market [95]. The economic losses due to FMD in northwest Ethiopia varied by production system. The mixed crop-livestock (MCL) system had lower losses of USD 34 per affected herd than the commercial dairy farms, which had losses of USD 459.1 per affected farm [49].
The average losses per herd and per animal in the MCL system were 76 USD and 9.8 USD, respectively. In the pastoral system, the average losses per herd and per animal were 174 USD and 5.3 USD, respectively. The animal-level mortality losses were 129 USD in the MCL system and 151 USD in the pastoral system [138]. In addition to export losses and control costs, an estimated annual economic loss of 1,350 million ETB was incurred. However, production losses were the costliest. Furthermore, an average of 40 million ETB and more than 300 million ETB were lost per year due to FMD export costs and animal death, respectively [76].
Tadesse, et al. [49] analyzed the economic losses due to FMD in commercial dairy farms and MCL production systems. They found that the average economic loss per dead animal was USD 194.45, ranging from USD 30.8 in young stock to USD 388.9 in draught ox. The average daily milk loss per affected lactating cow was 1.85 L (L), with 1.4 L in local cows and 2.9 L in crossbreed cows. FMD affected the milk production of cows differently depending on the production system. The average milk loss per infected cow was USD 26 in the mixed crop-livestock (MCL) system and USD 97.5 in commercial dairy farms [49]. They also found that mortality loss, milk loss and draft loss contributed to the economic loss at the animal level in the MCL production system, while milk loss was the main loss in commercial dairy farms, where no mortality loss was observed. This suggests that commercial dairy farms provided better care for infected animals [49]. Hence, future research should aim to develop and evaluate effective control measures to mitigate the morbidity and economic losses resulting from FMD.
FMD is a highly contagious viral disease that affects many kinds of animals, including livestock and wildlife [140]. To control FMD, countries need to assess their disease risk and implement appropriate measures to reduce it. One framework for doing this is the Progressive Control Pathway for FMD (PCP-FMD), which guides countries to gradually and systematically improve their FMD situation [141]. Some of the measures that countries can use to control FMD are destroying infected and exposed animals, restricting animal and animal product movement, and vaccinating susceptible animals. However, these measures have limitations because the FMD virus has many types and can spread easily [139]. Therefore, it is also important to develop better diagnostic and vaccine technologies, strengthen veterinary services and laboratories, coordinate national and regional actions, and educate and motivate farmers, traders, consumers, and policymakers to adopt good practices [79].
Diagnosis of FMD
Clinical diagnosis
FMD is a viral disease that spreads quickly and easily among cloven-hoofed animals, both domestic and wild. The disease can cause fever, loss of appetite, weight loss, lameness, drooling, and depression in infected animals. Blister-like sores on the mouth, teats and hooves are other characteristics of FMD (Fig. 6) [7]. It can also produce blisters on the mouth, feet, and udder that break and heal over time. The hoof wall may show growth rings due to damage to the skin around the hoof. The clinical signs of FMD vary depending on several factors, such as the type of virus, dose of exposure, age and breed of the animal, host species, and immune status. The OIE Terrestrial Manual (chapter 3.1.8) provides a detailed description of how FMD affects different animal species [54].

Ruptured blister on a cow's tongue (A); foot lesions on the coronet and interdigital area of the hoofs (B); affected animal develops vesicles in the muzzle (C) and on the teats (in lactating animals) (D); source: https://www.cfsph.iastate.edu/diseaseinfo/disease-images/?disease=foot-and-mouth-disease (accessed on 17 July 2023)
FMD diagnosis requires laboratory confirmation, as the clinical signs may be mild or unusual in some cases or may resemble other diseases that cause blisters. Laboratory tests can identify the virus or its antibodies in samples from suspected animals. The analysis of the lesion characteristics can also help to determine the source and duration of the virus infection or circulation. FMD diagnosis is important for effective control and surveillance of the disease [142].
Differential diagnosis
Clinical signs and postmortem findings can indicate FMD, but they are not conclusive because other vesicular diseases [143] can cause similar symptoms. Therefore, other vesicular disorders, such as SVD, vesicular stomatitis, traumatic stomatitis, and vesicular exanthema, should be ruled out when diagnosing FMD. Pigs can develop SVD, vesicular stomatitis, and FMD, while cattle can develop vesicular stomatitis and FMD [65]. Clinical signs are more noticeable in highly susceptible animals; however, in regions where FMD is endemic, clinical signs may be mild or ambiguous due to partial natural immunity or vaccinal immunity [144]. Therefore, laboratory tests are essential to confirm any suspicion of FMD and to stop the disease from spreading further [145].
Laboratory diagnosis
FMD diagnosis is important for effective control and prevention of the disease, as well as for avoiding socioeconomic impacts due to trade barriers imposed by FMD-free countries [145]. This review article provides an overview of some of the current diagnostic methods for FMD, such as ELISA-based methods and molecular methods. FMD diagnosis can be performed by detecting FMD virus or its specific antibodies in various samples from animals with clinical signs or exposure to the virus, such as fluid or tissue from blisters, fluid from the throat, milk, blood, or saliva [54]. Traditional methods of FMD diagnosis include virus isolation (VI), which is the gold standard but is slow and labor intensive and requires high-level biosafety facilities [146]; the virus neutralization test (VNT), which is highly specific and sensitive but also requires live virus and cell culture facilities [147]; the complement fixation test (CFT); and ELISA. In response to an FMD outbreak, prompt actions are typically implemented to ascertain a differential and definitive diagnosis, which is crucial to impede further dissemination of the disease (Fig. 7). Moreover, in conjunction with vaccination and stamping out policies, prompt FMD detection in cloven-hoofed animals utilizing currently available diagnostic instruments has been extensively employed as a strategy to combat this highly scrutinized agent [145].

Laboratory tests for FMD diagnosis based on viral materials and antibodies in different scenarios; adapted from https://www.ncbi.nlm.nih.gov/core/lw/2.0/html/tileshop_pmc/tileshop_pmc_inline.html?title=Click on image to zoom&p = PMC3&id = 7473413_fvets-07–00477-g0001.jpg (Accessed on 18 July 2023). Source: [145]
The complement fixation test (CFT) is a serological method that measures antibodies against FMD virus in serum samples. It is easy and inexpensive, but it has low sensitivity and specificity and can be affected by factors that disrupt complement activity in the samples [148]. Enzyme-linked immunosorbent assay (ELISA) is a common method for FMD diagnosis that can detect either FMD virus antigens or antibodies in different samples. ELISA has several benefits [145]. However, molecular methods have some limitations, such as the need for RNA extraction, the risk of false-positive results due to contamination or nonspecific amplification, and the inability to indicate whether the detected RNA is associated with infectious virus [149]. Molecular methods for FMD diagnosis are based on detecting the genetic material of FMD virus in samples. Some of these methods are conventional PCR, multiplex PCR (m-PCR), real-time PCR (RT‒qPCR), reverse transcription loop-mediated isothermal amplification (RT-LAMP), reverse transcription recombinase polymerase amplification (RT-RPA), reverse transcription insulated isothermal PCR (RT-iiPCR), and reverse transcription droplet digital PCR (RT-ddPCR) [145].
Inactivated vaccines for FMD contain purified FMDV antigen that lacks most of the viral nonstructural proteins (NSPs). These vaccines stimulate antibodies mainly against the viral proteins that form the structure (SP), which help the virus attach and enter the cells. However, when animals are naturally infected with FMDV, they produce antibodies against both SP and NSP, which play a role in virus replication and pathogenesis. Therefore, tests that can detect antibodies against NSPs can distinguish animals that are infected from those that are vaccinated (DIVA test) [150]. DIVA is an important strategy for FMD control and eradication programs that use vaccination as a tool. One of the most widely used NSPs for DIVA testing is the 3ABC polyprotein, which is highly conserved and immunogenic across all FMDV serotypes [151, 152].
Several methods based on ELISA can detect antibodies to the 3ABC polyprotein, a nonstructural protein of FMDV, in serum samples from various animals. These methods include liquid-phase blocking (LPBE-3ABC), solid-phase competition (SPC-3ABC), and direct or indirect sandwich ELISA (ELISA-3ABC). They have high sensitivity and specificity for FMD diagnosis. Immunoblotting can also detect nonstructural proteins and confirm the ELISA results. It can also show which nonstructural proteins are recognized by the antibodies. NSPs can help to monitor the FMDV infection status and vaccination efficacy in endemic regions with active vaccination programs [150]. In Ethiopia, FMD diagnosis is performed by using virus isolation, 3ABC-ELISA, virus neutralization tests, and conventional PCR at the National Veterinary Institute (NVI) and the National Animal Health Diagnostic Center (NAHDIC).
Strategies for control, prevention, and treatment of FMDV
Foot-and-mouth disease (FMD) prevention and control require a lot of resources and efforts, which are mostly provided by low- and lower-middle-income countries. FMD has seven different strains, each of which needs a specific vaccine. Therefore, FMD prevention depends on the ability to detect the disease early, to warn other countries and regions, and to monitor the animal populations for signs of infection [7]. FMDV infection can be prolonged by antibiotics, but they do not clear the virus from the body. Antibiotics can help infected animals survive longer, but they also increase the risk of virus persistence and transmission. The virus can stay in the pharyngeal epithelia of some animals for a long time, making them carriers of the disease. These carriers can infect other animals through their saliva and nasal secretions, either directly or indirectly [34, 153]. Different countries have different strategies to control FMD, depending on their resources and regulations. In developed countries, the main strategy is to cull and dispose of infected or exposed animals and disinfect their premises. This can prevent the spread of the disease and protect the trade of animal products [154, 155].
However, in Ethiopia, where livestock is a vital source of income and food security, culling is not feasible. Instead, infected animals are isolated and treated with supportive care, such as antibiotics, anti-inflammatory drugs, and wound care. This can reduce the death and suffering of the animals, but it does not stop the virus from persisting and transmitting [156]. This allows the virus to remain in the animal population and cause recurrent outbreaks of FMD [157]. Antibiotic maintenance may have multiple impacts on FMD transmission, which are not well understood or documented. It may alter the gut microbiota of animals, which may impair their immune system and increase their susceptibility to FMD infection [158]. It may also promote the development and spread of antibiotic resistance genes, which may be transferred to other microorganisms through horizontal gene transfer. This may lead to the emergence of novel or more virulent FMD virus strains [159].
Furthermore, it may enable the movement and trade of animals across regions or borders, which may expose them to different sources and types of FMD virus. These potential effects of antibiotic maintenance on FMD transmission pose serious challenges and risks for animal health, public health, and food security in low-income countries [160]. Therefore, it is important to promote the rational use of antibiotics and implement appropriate biosecurity measures to prevent and control FMD in low-income countries [161]. We also need more research and evidence to understand how and why these effects happen, and what we can do to stop them. This also requires a One Health approach that considers the health of animals, humans, and the environment together.
Vaccination
FMD control strategies and vaccine development vary by disease status of the country or region. In FMD-free countries, the main strategy is to stamp out infected and in-contact animals, disinfect premises, and vaccinate animals around outbreak areas [61]. In endemic countries, these options are not feasible. Hence, following the OIE/FAO progressive control pathway helps to enhance veterinary services, outbreak preparedness and vaccination programs [68]. In many developing countries, vaccination with an inactivated whole-virus vaccine is the only way to control the epidemic [162]. Some countries or regions have eliminated FMD by using inactivated FMD vaccines as part of mandatory vaccination programs. However, FMD remains endemic in parts of Africa and Asia [6]. One of the limitations of inactivated FMD vaccines is that they confer short-term immunity and require frequent boosting [162].
FMD virus infects a wide range of wild and domesticated cloven-footed mammals and can spread rapidly across borders. FMD outbreaks can have severe economic consequences for the livestock industry due to trade restrictions and reduced animal productivity. An ideal vaccine formulation should meet the following criteria: safety, thermal stability, low cost, multivalency, rapid and long-lasting immunity with a single dose, and compatibility with DIVA (differentiating infected from vaccinated animals) principles (Fig. 8) [139].

Source: source [139]
Desirable features of a perfect FMD vaccine; Adapted from https://link.springer.com/article/10.1007/s00705-019-04216-x/figures/2 (Accessed on 17 August 2023).
Currently, inactivated vaccines are the most widely used to prevent FMD in endemic regions, but they have several limitations, such as the need for high-containment facilities to grow virulent FMDV, the limited cross-protection among different strains and topotypes within the same serotype, the frequent emergence of new variants that may escape vaccine-induced immunity, and the inability to eliminate virus carriers [68].
FMD vaccines mainly consist of inactivated viruses that elicit protective antibodies against virus structural proteins. However, when FMDV infects an animal, it also reproduces and makes the body produce more antibodies against its inner proteins. These antibodies can help to distinguish.
animals that are infected from those that are vaccinated (DIVA), which is important for knowing how widespread or rare the infection is. Additionally, NSP antibody tests can identify FMD infection regardless of the virus serotypes present [150, 164]. However, some new FMDV strains found in Egypt could not be identified by the usual molecular methods for serotyping, suggesting that they are different from the ones that the current FMDV vaccines can protect against [122].
A recent study by [11] reported a high seroprevalence of FMDV and some risk factors (district, age, body condition score, and management system) in Ethiopia, suggesting the need for tailored control and prevention strategies based on the circulating virus serotype. However, determining the circulating virus serotype is difficult in Ethiopia, as FMD outbreaks caused by serotypes A, O, and SAT2 have occurred in different regions [99].The current trivalent inactivated vaccine used in Ethiopia for FMDV serotypes O, A, and SAT2 contains vaccine strains ETH/38/2005, ETH/6/2000, and ETH/64/2009, respectively [165]. These vaccine strains may not match well with the field strains and may provide suboptimal protection. Outbreak reports guide FMD control measures in the country. However, few private livestock owners vaccinate their animals regularly or before FMD outbreaks due to various reasons, such as lack of awareness, cost, or availability of vaccines. Continuous molecular monitoring of circulating FMDV strains is strongly recommended to ensure the selection of the appropriate strain for the preparation of effective vaccines [14]. According to a vaccine-matching study conducted in Ethiopia, the O/ETH/38/2005 vaccine strain can provide protection against outbreaks caused by the O/EA-3 topotype but has lower efficacy for the O/EA-4 topotype [166].
Additionally, the amount of FMD vaccines produced in Ethiopia is insufficient compared to the country’s livestock population, and the vaccines are also expensive, making livestock owners reluctant to pay for them [167]. To reduce the economic losses caused by FMD, the government should implement policies that include vaccination and movement restrictions for livestock. These actions can stop FMD from spreading in the country, which can happen because of poor or missing vaccination, free animal movement, and the market chain[99]. Additionally, not knowing enough about the molecular details of FMD makes it harder to control it [81]. Therefore, a molecular understanding of virus-antibody interactions is required for the development of better vaccines as well as timely assessment of the spread and severity of epidemics [168].
The vaccine development efforts against FMD are not commercially attractive despite the huge market potential because of various technical, regulatory, and economic challenges [169]. Some of these challenges include the costly biosafety facilities required to produce live virus, the need to differentiate infected from vaccinated animals, and the variable local regulatory restrictions to produce and commercialize the vaccine. The high investment and operational costs of FMD vaccine production and delivery, the low profit margin and market share of FMD vaccines, the lack of incentives and subsidies for FMD vaccine research and development, and the competition and conflicts of interest among different stakeholders in FMD control and eradication also hinder the development of novel vaccines that overcome these limitations and provide effective and safe protection against FMD [153, 170,171,172].
Application of nanoliposomes
Although commercially available FMD vaccines are effective, they provide short-term immunity requiring regular boosters. A new FMD vaccine is needed to improve immunization, safety, and long-term immune responses. A synthetic peptide vaccine is one of the safe and important vaccines. Peptide vaccine has low immunogenicity, requiring strong adjuvants. Nanoliposomes can be used as new adjuvants to improve immune response. In the current study, nanoliposomal carriers were selected using Dimyristoylphosphatidylcholine (DMPC), dimyristoyl phosphoglycerol (DMPG), and Cholesterol (Chol) as an adjuvant containing two immunodominant synthetic FMDV peptides. The liposomal formulations were characterized by various physicochemical properties. The size, zeta potential, and encapsulation efficiency were optimized, and the obtained nanoliposome was suitable as a vaccine [162].
Nanoliposomes are tiny vesicles made of lipids that can carry drugs or antigens inside them. They are promising candidates for delivering vaccines against FMD. Nanoliposomes can enhance the immune response to FMD vaccines by protecting the antigens from degradation and targeting them to specific cells. Nanoliposomes can also release antigens slowly and steadily, which may improve the duration and quality of immunity. Lycium barbarum polysaccharides (LBP) are natural compounds that have immunomodulatory effects. They can be encapsulated in nanoliposomes to act as adjuvants for FMD vaccines. LBP-nanoliposomes have been shown to induce higher antibody levels and stronger cellular immunity than conventional adjuvants in mice [173]. Therefore, nanoliposomes are a potential nanotechnology platform for developing more effective and safer FMD vaccines. However, vaccines take several days to induce a response and need a booster dose to sustain herd immunity [174].
Surveillance and biosecurity
The main source of FMD surveillance data in Ethiopia is the passive reporting system, where farmers and veterinarians notify the National Animal Health Diagnostic and Investigation Center (NAHDIC) and the National Veterinary Institute (NVI) of suspected FMD cases. These centers also perform laboratory confirmation and molecular typing of FMDV isolates obtained from outbreak investigations [81, 175]. To prevent and control FMD transmission among livestock, Ethiopia implements various biosecurity measures at different levels, such as restricting animal movements, enforcing quarantine regulations, conducting vaccination campaigns, applying disinfection protocols, disposing of infected carcasses and animal products safely, and raising public awareness and education on FMD prevention [12, 175]. However, these measures may not be very effective in Ethiopia due to several challenges, such as the coexistence of multiple serotypes and subtypes of FMDV, free livestock movement, scarcity of effective and affordable FMD vaccines, lack of regular vaccination campaigns, and inadequate public awareness of FMD prevention.
FMD surveillance and biosecurity in Ethiopia face several challenges, such as lack of adequate resources, infrastructure, personnel, coordination, and data management; low coverage and efficacy of vaccination; high diversity and mobility of livestock; cross-border transmission of FMDV; and socio-economic factors that influence farmers’ compliance and participation. Moreover, FMD surveillance and biosecurity in Ethiopia also offer some opportunities for improvement, such as enhancing the capacity and collaboration of national and regional laboratories; applying novel diagnostic and epidemiological tools; developing effective and tailored vaccines; engaging stakeholders and communities; and harmonizing regional and international efforts to control FMD [12, 81, 175].
FMD control and surveillance require prompt and accurate disease confirmation [176]. FMD can also be controlled by restricting animal movement, isolating infected animals, and disinfecting infected premises [177, 178]. However, FMD vaccine development and selection are challenging due to the antigenic diversity and genetic variation of FMDV serotypes and strains [179]. Antibodies induced by one serotype do not protect against other serotypes and may not cross-protect within the same serotype [80]. Therefore, it is essential to monitor new FMDV strains to prevent their spread and ensure effective vaccination programs. The identification of new FMDV variants will help to update vaccine production and control measures [8, 122]. FMD vaccines are often designed to cover multiple serotypes and strains, but this compromises their efficacy and cost compared to monovalent vaccines [180], making the use of vaccination to control FMD challenging for developing countries with limited resources such as Ethiopia [167].
Epidemiological techniques such as routine disease data collection in the population (monitoring and surveillance) are essential for detecting newly emerged serotypes for vaccine matching studies, which help to maintain effective vaccine production [166]. In Ethiopia, where FMD is endemic, new viral strains may emerge. Hence, the surveillance system needs to implement several actions, such as notifying suspected cases by farmers and veterinarians, investigating suspected and high-risk properties, conducting serological surveys, and monitoring animals in slaughterhouses. The authors also recommend more FMD surveillance and multivalent vaccines and emphasize regular vaccination and control measures for sustainable food security and rural development.
Conclusions
FMD is a major threat to the livestock sector in Ethiopia. The virus, which has multiple serotypes and strains, spreads through transboundary animal movement. Due to the high genetic variation and antigenic diversity of FMDV, infection with one genetic lineage does not provide full protection against another lineage in the same serotype. Different lineages of the same serotype that evolved independently in different geographical regions are called different topotypes, and the current trivalent vaccines in Ethiopia may not cover all of them. FMD affects the domestic and international markets of livestock products, as well as food security and economic development in the country. Therefore, effective control measures are needed to combat FMD. We propose the following recommendations:
-
Develop better and multivalent vaccines that provide longer and wider protection against different FMDV strains.
-
Regulate cattle movement and prevent contact between infected and susceptible herds.
-
Ensure adequate vaccine supplies to endemic areas and encourage regular vaccination among livestock owners.
-
Monitor the antigenic diversity and evolution of FMDV in the country through vaccine matching studies.
-
Understand the spatial and temporal distribution of FMD at the national level through comprehensive surveillance studies.
Availability of data and materials
Not applicable.
Abbreviations
- Abs:
-
Antibodies
- CFT:
-
Complement fixation test
- DIVA:
-
Differentiate infected from vaccinated animals
- EA:
-
East Africa
- ELISA:
-
Enzyme-linked immunosorbent assay
- ETB:
-
Ethiopian Birr
- FMD:
-
Foot-and-mouth disease
- LPBE:
-
Liquid-phase blocking ELISA
- Lpro :
-
Leader proteinase
- MCL:
-
Mixed crop-livestock
- m-PCR:
-
Multiplex-PCR
- NAHDIC:
-
National animal health diagnostic and investigation center
- NSP:
-
Nonstructural protein
- NVI:
-
National Veterinary institute
- ORF:
-
Open reading frame
- PCR:
-
Polymerase chain reaction
- RNA:
-
Ribonucleic acid
- SAT:
-
South African territories
- SP:
-
Structural protein
- SVD:
-
Swine vesicular disease
- USD:
-
United States dollar
- VNT:
-
Virus neutralization test
- VP:
-
Viral protein
References
CSA Agricultural sample survey 2020/21 [2013]. Report on livestock and livestock characteristics. Statistical Bulletin 589. Vol. II. Addis Ababa, Ethiopia.," Ethiopia, 2021, vol. II. [Online]. Available: http://www.statsethiopia.gov.et/wp-content/uploads/2021/05/2013.LIVESTOCK-REPORT.FINAL_.pdf
Dabasa G, Abunna F. Review on Epidemiology of Foot and Mouth Disease (FMD) in Ethiopia. J Trop Dis. 2021;9:269.
Tesfaye Y, Khan F, Gelaye E. Molecular characterization of foot-and-mouth disease viruses collected from Northern and Central Ethiopia during the 2018 outbreak. Vet World. 2020;13(3):542–8. https://doi.org/10.14202/vetworld.2020.542-548.
Zhang Z, Waters R, Li Y. Pathogenesis of non-epithelial foot-and-mouth disease in neonatal animals. Vet Microbiol. 2021;254: 108961. https://doi.org/10.1016/j.vetmic.2020.108961.
Rodríguez-Habibe I, Celis-Giraldo C, Patarroyo ME, Avendaño C, Patarroyo MA. A comprehensive review of the immunological response against foot-and-mouth disease virus infection and its evasion mechanisms. Vaccines (Basel). 2020. https://doi.org/10.3390/vaccines8040764.
Lu Z, et al. Development of foot-and-mouth disease vaccines in recent years. Vaccines (Basel). 2022. https://doi.org/10.3390/vaccines10111817.
WOAH. Foot and mouth disease (FMD). World Organisation for Animal Health. https://www.woah.org/en/disease/foot-and-mouth-disease/ (accessed 21 March 2023
Ward JC, et al. The RNA pseudoknots in foot-and-mouth disease virus are dispensable for genome replication, but essential for the production of infectious virus. PLoS Pathog. 2022;18(6): e1010589. https://doi.org/10.1371/journal.ppat.1010589.
J. L. Martel 1974 Foot-and-mouth disease in Ethiopia. Distribution of serotypes of foot-and-mouth disease virus, Rev Elev Med Vet Pays Trop, 27 2: 169–75, 1974. La fièvre aphteuse en Ethiopie. Distribution des sérotypes de virus aphteux.
Desissa F, Tura D, Mamo B, Rufael T. Epidemiological study on foot and mouth disease in cattle: seroprevalence and risk factor assessment in Kellem Wollega Zone, West Ethiopia. Afr J Agric Res. 2014;9(18):1391–5.
Awel SM, Dilba GM, Abraha B, Zewde D, Wakjira BS, Aliy A. Seroprevalence and molecular detection of foot and mouth disease virus in dairy cattle around Addis Ababa, Central Ethiopia. Vet Med (Auckl). 2021;12:187–97. https://doi.org/10.2147/vmrr.S317103.
Abdela N. Sero-prevalence, risk factors and distribution of foot and mouth disease in Ethiopia. Acta Trop. 2017;169:125–32. https://doi.org/10.1016/j.actatropica.2017.02.017.
Ayelet G, Gelaye E, Negussie H, Asmare K. Study on the epidemiology of foot and mouth disease in Ethiopia. Rev Sci Tech. 2012;31(3):789–98. https://doi.org/10.20506/rst.31.3.2153.
Hemida MG, Rizk El-Ghareeb W, Al-Hizab F, Ibrahim A. Foot-and-mouth disease virus O/ME-SA/Ind 2001 lineage outbreak in vaccinated Holstein Friesian cattle in Saudi Arabia in 2016. Vet Q. 2018;381:88–98. https://doi.org/10.1080/01652176.2018.1539568.
Dubie T, Negash W. Seroprevalence of bovine foot and mouth disease (FMD) and its associated risk factors in selected districts of Afar region, Ethiopia. Vet Med Sci. 2021;7(5):1678–87. https://doi.org/10.1002/vms3.574.
Shurbe M, Simeon B, Seyoum W, Muluneh A, Tora E, Abayneh E. Seroprevalence and associated risk factors for foot and mouth disease virus seropositivity in cattle in selected districts of Gamo zone, Southern Ethiopia. Front Vet Sci. 2022;9: 931643. https://doi.org/10.3389/fvets.2022.931643.
Wubshet AK, Dai J, Li Q, Zhang J. Review on outbreak dynamics, the endemic serotypes, and diversified topotypic profiles of foot and mouth disease virus isolates in Ethiopia from 2008 to 2018. Viruses. 2019;11(11):1076.
Knowles NJ, Samuel AR. "Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 2003;91(1):65–80. https://doi.org/10.1016/s0168-1702(02)00260-5.
Carrillo C, et al. Comparative genomics of foot-and-mouth disease virus. J Virol. 2005;79(10):6487–504. https://doi.org/10.1128/jvi.79.10.6487-6504.2005.
Grubman MJ, Baxt B. Foot-and-mouth disease. Clin Microbiol Rev. 2004;17(2):465–93.
Grubman MJ. The 5’ end of foot-and-mouth disease virion RNA contains a protein covalently linked to the nucleotide pUp. Arch Virol. 1980;63(3–4):311–5. https://doi.org/10.1007/bf01315038.
Gao Y, Sun SQ, Guo HC. Biological function of Foot-and-mouth disease virus non-structural proteins and non-coding elements. Virol J. 2016;13:107. https://doi.org/10.1186/s12985-016-0561-z.
Semkum P, et al. A novel plasmid DNA-based foot and mouth disease virus minigenome for intracytoplasmic mRNA production. Viruses. 2021. https://doi.org/10.3390/v13061047.
Serrano P, Pulido MR, Sáiz M, Martínez-Salas E. The 3’ end of the foot-and-mouth disease virus genome establishes two distinct long-range RNA-RNA interactions with the 5’ end region. J Gen Virol. 2006;87(Pt 10):3013–22. https://doi.org/10.1099/vir.0.82059-0.
Logan G, et al. Deep sequencing of foot-and-mouth disease virus reveals RNA sequences involved in genome packaging. J Virol. 2018. https://doi.org/10.1128/jvi.01159-17.
Brito B, et al. A traditional evolutionary history of foot-and-mouth disease viruses in Southeast Asia challenged by analyses of non-structural protein coding sequences. Sci Rep. 2018;8(1):6472. https://doi.org/10.1038/s41598-018-24870-6.
Paton DJ, et al. The history of foot-and-mouth disease virus serotype C: the first known extinct serotype? Virus Evol. 2021. https://doi.org/10.1093/ve/veab009.
Ekanayaka P, et al. Foot-and-mouth disease virus VP1 target the MAVS to inhibit type-I interferon signaling and VP1 E83K mutation results in virus attenuation. PLoS Pathog. 2020;16(11): e1009057. https://doi.org/10.1371/journal.ppat.1009057.
Zhu Z, et al. Foot-and-mouth disease virus capsid protein VP1 interacts with host ribosomal protein SA To maintain activation of the MAPK signal pathway and promote virus replication. J Virol. 2020. https://doi.org/10.1128/jvi.01350-19.
Childs K, Jackson B, Harvey Y, Seago J. Trans-encapsidation of foot-and-mouth disease virus genomes facilitates escape from neutralizing antibodies. Viruses. 2022. https://doi.org/10.3390/v14061161.
Peng YH, Lin CH, Lin CN, Lo CY, Tsai TL, Wu HY. Characterization of the role of hexamer AGUAAA and Poly(A) tail in coronavirus polyadenylation. PLoS ONE. 2016;11(10): e0165077. https://doi.org/10.1371/journal.pone.0165077.
Spagnolo JF, Hogue BG. Requirement of the poly(A) tail in coronavirus genome replication. Adv Exp Med Biol. 2001;494:467–74. https://doi.org/10.1007/978-1-4615-1325-4_68.
Mitton-Fry RM, DeGregorio SJ, Wang J, Steitz TA, Steitz JA. Poly(A) tail recognition by a viral RNA element through assembly of a triple helix. Science. 2010;330(6008):1244–7. https://doi.org/10.1126/science.1195858.
Li K, Wang C, Yang F, Cao W, Zhu Z, Zheng H. Virus-Host interactions in foot-and-mouth disease virus infection. Front Immunol. 2021;12: 571509. https://doi.org/10.3389/fimmu.2021.571509.
Valdazo-González B, Timina A, Scherbakov A, Abdul-Hamid NF, Knowles NJ, King DP. Multiple introductions of serotype O foot-and-mouth disease viruses into East Asia in 2010–2011. Vet Res. 2013;44(1):76. https://doi.org/10.1186/1297-9716-44-76.
Li G, et al. Antigenicity and immunogenicity analysis of the E. coli expressed FMDV structural proteins; VP1, VP0, VP3 of the South African territories type 2 virus. Viruses. 2021. https://doi.org/10.3390/v13061005.
Dong H, et al. Structural and molecular basis for foot-and-mouth disease virus neutralization by two potent protective antibodies. Protein Cell. 2022;13(6):446–53. https://doi.org/10.1007/s13238-021-00828-9.
Kristensen T, Belsham GJ. Identification of plasticity and interactions of a highly conserved motif within a picornavirus capsid precursor required for virus infectivity. Sci Rep. 2019;9(1):11747. https://doi.org/10.1038/s41598-019-48170-9.
Zhang K, et al. Foot-and-mouth disease virus structural protein VP1 destroys the stability of TPL2 trimer by degradation TPL2 to evade host antiviral immunity. J Virol. 2020. https://doi.org/10.1128/jvi.02149-20.
Mishu ID, Akter S, Alam A, Hossain MA, Sultana M. In silico evolutionary divergence analysis suggests the potentiality of capsid protein VP2 in serotype-independent foot-and-mouth disease virus detection. Front Vet Sci. 2020;7:592. https://doi.org/10.3389/fvets.2020.00592.
Dill V, Eschbaumer M. Cell culture propagation of foot-and-mouth disease virus: adaptive amino acid substitutions in structural proteins and their functional implications. Virus Genes. 2020;56(1):1–15. https://doi.org/10.1007/s11262-019-01714-7.
Chitray M, et al. Diagnostic and epitope mapping potential of single-chain antibody fragments against foot-and-mouth disease virus serotypes A, SAT1, and SAT3. Front Vet Sci. 2020;7:475. https://doi.org/10.3389/fvets.2020.00475.
Azzinaro PA, et al. Mutation of FMDV L(pro) H138 residue drives viral attenuation in cell culture and in vivo in swine. Front Vet Sci. 2022;9:1028077. https://doi.org/10.3389/fvets.2022.1028077.
Zell R, et al. ICTV virus taxonomy profile: picornaviridae. J Gen Virol. 2017;98(10):2421–2. https://doi.org/10.1099/jgv.0.000911.
Belsham GJ, Kristensen T, Jackson T. Foot-and-mouth disease virus: Prospects for using knowledge of virus biology to improve control of this continuing global threat. Virus Res. 2020;281: 197909. https://doi.org/10.1016/j.virusres.2020.197909.
Brito BP, Rodriguez LL, Hammond JM, Pinto J, Perez AM. Review of the global distribution of foot-and-mouth disease Virus from 2007 to 2014. Transbound Emerg Dis. 2017;64(2):316–32. https://doi.org/10.1111/tbed.12373.
OIE foot-and-mouth-disease-1.pdf in OIE Technical Disease Card: Foot and mouth disease, 2021, pp. 1–6
Jemberu WT, Molla W, Fentie T. A randomized controlled field trial assessing foot and mouth disease vaccine effectiveness in Gondar Zuria district, Northwest Ethiopia. Prev Vet Med. 2020;183: 105136. https://doi.org/10.1016/j.prevetmed.2020.105136.
Tadesse B, Tesfahun A, Molla W, Demisse E, Jemberu WT. Foot and mouth disease outbreak investigation and estimation of its economic impact in selected districts in northwest Ethiopia. Vet Med Sci. 2020;6(1):122–32. https://doi.org/10.1002/vms3.208.
Muthukrishnan M, Singanallur Balasubramanian N, Villuppanoor Alwar S. Experimental infection of foot and mouth disease in Indian sheep and goats. Front Vet Sci. 2020;7:356. https://doi.org/10.3389/fvets.2020.00356.
Stenfeldt C, Pacheco JM, Rodriguez LL, Arzt J. “Early events in the pathogenesis of foot-and-mouth disease in pigs; identification of oropharyngeal tonsils as sites of primary and sustained viral replication,” (in eng). PLoS ONE. 2014;9(9): e106859. https://doi.org/10.1371/journal.pone.0106859.
Graham B, Anette B, Louise L Foot-and-Mouth Disease in Animals. https://www.msdvetmanual.com/generalized-conditions/foot-and-mouth-disease/foot-and-mouth-disease-in-animals (accessed 07 June, 2023).
Yang B, et al. Molecular mechanisms of immune escape for foot-and-mouth disease virus. Pathogens. 2020. https://doi.org/10.3390/pathogens9090729.
OIE Foot-and-Mouth Disease (Infection with Foot and Mouth disease viruses) " in OIE Terrestrial Manual 2022 2022, ch. (Chapter 3.1.8), pp 1–34
Knight-Jones TJD, McLaws M, Rushton J. Foot-and-mouth disease impact on smallholders - what do we know, what don’t we know and how can we find out more? Transbound Emerg Dis. 2017;64(4):1079–94. https://doi.org/10.1111/tbed.12507.
Wang Y, et al. Inability of FMDV replication in equine kidney epithelial cells is independent of integrin αvβ3 and αvβ6. Virology. 2016;492:251–8. https://doi.org/10.1016/j.virol.2016.01.025.
WOAH. Foot and Mouth disease (FMD)." WOrld Organization for Animal Health. https://www.woah.org/en/disease/foot-and-mouth-disease/ (accessed 21 March 2023, 2023)
FAO. The global foot and mouth disease control strategy." Food and Agriculture Organization of the United Nations. https://www.fao.org/in-action/kore/good-practices/good-practices-details/en/c/1137339/ (accessed 29 Nov 2023, 2023)
Brown E, Nelson N, Gubbins S, Colenutt C. Airborne transmission of foot-and-mouth disease virus: a review of past and present perspectives. Viruses. 2022. https://doi.org/10.3390/v14051009.
Souley Kouato B, et al. Outbreak investigations and molecular characterization of foot-and-mouth disease viruses circulating in south-west Niger. Transbound Emerg Dis. 2018;65(1):146–57. https://doi.org/10.1111/tbed.12642.
Colenutt C, et al. Quantifying the transmission of foot-and-mouth disease virus in cattle via a contaminated environment. MBio. 2020. https://doi.org/10.1128/mBio.00381-20.
Colenutt C, et al. Environmental sampling as a low-technology method for surveillance of foot-and-mouth disease virus in an area of endemicity. Appl Environ Microbiol. 2018. https://doi.org/10.1128/aem.00686-18.
Jemberu WT, Mourits MC, Sahle M, Siraw B, Vernooij JC, Hogeveen H. Epidemiology of foot and mouth disease in Ethiopia: a retrospective analysis of district level outbreaks, 2007–2012. Transbound Emerg Dis. 2016;63(6):e246–59. https://doi.org/10.1111/tbed.12338.
Gortázar C, Barroso P, Nova R, Cáceres G. The role of wildlife in the epidemiology and control of Foot-and-mouth-disease And Similar Transboundary (FAST) animal diseases: a review. Transbound Emerg Dis. 2022;69(5):2462–73. https://doi.org/10.1111/tbed.14235.
Jamal SM, Belsham GJ. Foot-and-mouth disease: past, present and future. Vet Res. 2013;44(1):116. https://doi.org/10.1186/1297-9716-44-116.
Wang Y, Liu M, The Causative Agent of FMD Disease (Some RNA Viruses). 2020
Aiewsakun P, Pamornchainavakul N, Inchaisri C. Early origin and global colonisation of foot-and-mouth disease virus. Sci Rep. 2020;10(1):15268. https://doi.org/10.1038/s41598-020-72246-6.
Hammond JM, Maulidi B, Henning N. Targeted FMD vaccines for Eastern Africa: the AgResults foot and mouth disease vaccine challenge project. Viruses. 2021. https://doi.org/10.3390/v13091830.
OIE. South-East Asia, China and Mongolia Foot and Mouth Disease (SEACFMD) Campaign." World Organisation for Animal Health. https://rr-asia.woah.org/en/our-mission/our-mission-regional-strategies/the-south-east-asia-and-china-foot-and-mouth-disease-seacfmd-campaign/ (accessed 02 December 2023, 2023).
Chen J, et al. Retrospect and risk analysis of foot-and-mouth disease in China based on integrated surveillance and spatial analysis tools. Front Vet Sci. 2019;6:511. https://doi.org/10.3389/fvets.2019.00511.
Haoran W, et al. Assessment of foot-and-mouth disease risk areas in mainland China based spatial multi-criteria decision analysis. BMC Vet Res. 2021;17(1):374. https://doi.org/10.1186/s12917-021-03084-5.
Donaldson AI, Alexandersen S, Sørensen JH, Mikkelsen T. Relative risks of the uncontrollable (airborne) spread of FMD by different species. Vet Rec. 2001;148(19):602–4. https://doi.org/10.1136/vr.148.19.602.
Knight-Jones TJ, Rushton J. The economic impacts of foot and mouth disease - what are they, how big are they and where do they occur? Prev Vet Med. 2013;112(3–4):161–73. https://doi.org/10.1016/j.prevetmed.2013.07.013.
FAO. "The global foot and mouth disease control strategy." FAO. https://www.fao.org/3/I9857EN/i9857en.PDF (accessed 14 June 2023).
Marqués FJ, Battistessa EI, Peek SF, Raabis SM, Darien BJ. The effect of foot-and-mouth disease vaccination on early pregnancy loss in beef heifers in Argentina. Prev Vet Med. 2019;170:104716. https://doi.org/10.1016/j.prevetmed.2019.104716.
Jemberu WT, Mourits M, Rushton J, Hogeveen H. Cost-benefit analysis of foot and mouth disease control in Ethiopia. Prev Vet Med. 2016;132:67–82. https://doi.org/10.1016/j.prevetmed.2016.08.008.
Domingo E, et al. Foot-and-mouth disease virus evolution: exploring pathways towards virus extinction. Curr Top Microbiol Immunol. 2005;288:149–73. https://doi.org/10.1007/3-540-27109-0_7.
Knowles N, Samuel A. Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 2003;91(1):65–80.
Paton DJ, Sumption KJ, Charleston B. Options for control of foot-and-mouth disease: knowledge, capability and policy. Philos Trans R Soc Lond B Biol Sci. 2009;364(1530):2657–67. https://doi.org/10.1098/rstb.2009.0100.
Yang M, Holland H, Clavijo A. Production of monoclonal antibodies against whole virus particles of foot-and-mouth disease virus serotype O and A and their potential use in quantification of intact virus for vaccine manufacture. Vaccine. 2008;26(27–28):3377–82. https://doi.org/10.1016/j.vaccine.2008.04.062.
Gizaw D, et al. Molecular characterization of foot-and-mouth disease viruses circulating in Ethiopia between 2008 and 2019. Transbound Emerg Dis. 2020;67(6):2983–92. https://doi.org/10.1111/tbed.13675.
Tesfaye A, Sehale M, Abebe A, Muluneh A, Gizaw D. Sero-prevalence of foot and mouth disease in cattle in Borena Zone, Oromia regional state, Ethiopia. Ethiopian Veterinary Journal. 2016;20(1):55–66.
Woldemariyam, et al. Temporal and spatial patterns and a space-time cluster analysis of foot-and-mouth disease outbreaks in Ethiopia from 2010 to 2019. Viruses. 2022. https://doi.org/10.3390/v14071558.
Roeder PL, Abraham G, Mebratu GY, Kitching RP. Foot-and-mouth disease in Ethiopia from 1988 to 1991. Trop Anim Health Prod. 1994;26(3):163–7. https://doi.org/10.1007/bf02241075.
Legesse Y, Asfaw Y, Sahle M, Ayelet G, Jenberie S, Negussie H. First confirmation of foot and mouth disease virus serotype SAT-1 in cattle and small ruminants in Ethiopia in 2007/08. Trop Anim Health Prod. 2013;45(5):1265–7. https://doi.org/10.1007/s11250-012-0339-2.
Asfaw W, Sintaro T. The status of FMD in Ethiopia, A growing concern. Ethiop Vet Epi News Lett. 2000;1:1–5.
Megersa B, Beyene B, Abunna F, Regassa A, Amenu K, Rufael T. Risk factors for foot and mouth disease seroprevalence in indigenous cattle in Southern Ethiopia: the effect of production system. Trop Anim Health Prod. 2009;41(6):891–8. https://doi.org/10.1007/s11250-008-9276-5.
Rweyemamu M, Roeder P, MacKay D, Sumption K, Brownlie J, Leforban Y. Planning for the progressive control of foot-and-mouth disease worldwide. Transbound Emerg Dis. 2008;55(1):73–87. https://doi.org/10.1111/j.1865-1682.2007.01016.x.
Tesfaye A, Mengistu A, Rufael T. Sero-prevalence status of foot and mouth disease in the North Western Amhara Regional State, Ethiopia. Ethiop Vet J. 2016;20(2):43–53.
Bahiru A, Assefa A. Seroepidemiological investigation of foot and mouth disease (FMD) in Northern Amhara, Ethiopia. Sci Afr. 2022;16: e01267.
Gezahegn A, Girma Z, Berhanu A. Seroprevalence of foot and mouth disease (FMD) and associated economic impact on Central Ethiopian cattle feedlots. J Vet Med Anim Health. 2014;6(5):154–8.
Mohamoud A, Tessema E, Degefu H. Seroprevalence of bovine foot and mouth disease (FMD) in Awbere and Babille districts of Jijiga zone, Somalia Regional State, Eastern Ethiopia. Afr J Microbiol Res. 2011;5(21):3559–63.
Mesfine M, Nigatu S, Belayneh N, Jemberu WT. Sero-epidemiology of foot and mouth disease in domestic ruminants in Amhara Region, Ethiopia. Frontiers in Veterinary Science. 2019;6:130.
Ahmed B, Megersa L, Mulatu G, Siraj M, Boneya G. Seroprevalence and associated risk factors of foot and mouth disease in Cattle in West Shewa Zone, Ethiopia. Vet Med Int. 2020;2020:6821809. https://doi.org/10.1155/2020/6821809.
Alemayehu G, Zewde G, Admassu B. Seroprevalence of foot and mouth disease (FMD) and associated economic impact on Central Ethiopian cattle feedlots. J Vet Med Anim Health. 2014;6(5):154–8.
Molla B, Ayelet G, Asfaw Y, Jibril Y, Ganga G, Gelaye E. Epidemiological study on foot-and-mouth disease in cattle: seroprevalence and risk factor assessment in South Omo zone, south-western Ethiopia. Transbound Emerg Dis. 2010;57(5):340–7.
Abunna F, Fikru S, Rufael T. Seroprevalence of foot and mouth disease (FMD) at dire dawa and its surroundings, Eastern Ethiopia. Global Veterinaria. 2013;11(5):575–8.
Belina D, Girma B, Mengistu S. Sero-prevalence of bovine foot and mouth disease in selected districts of eastern showa zone, Oromia regional state, Ethiopia. Glob J Sci Front Res. 2016;16(4):79–84.
Sulayeman M, Dawo F, Mammo B, Gizaw D, Shegu D. Isolation, molecular characterization and sero-prevalence study of foot-and-mouth disease virus circulating in central Ethiopia. BMC Vet Res. 2018;14(1):110. https://doi.org/10.1186/s12917-018-1429-9.
Jenbere TS, Manyahilishal E, Haileluel N. Study on the risk factors of foot and mouth disease in selected districts of Afar pastoral area, northeast Ethiopia. J Anim Vet Adv. 2011;10(11):1368–72.
Mesfine M, Nigatu S, Belayneh N, Jemberu WT. Sero-Epidemiology of foot and mouth disease in domestic ruminants in Amhara Region, Ethiopia. Front Vet Sci. 2019;6:130. https://doi.org/10.3389/fvets.2019.00130.
Govindaraj G, et al. Foot and Mouth Disease (FMD) incidence in cattle and buffaloes and its associated farm-level economic costs in endemic India. Prev Vet Med. 2021;190:105318. https://doi.org/10.1016/j.prevetmed.2021.105318.
Gelaye E, Ayelet G, Abera T, Asmare K. Seroprevalence of foot and mouth disease in Bench Maji zone, Southwestern Ethiopia. J Vet Med Anim Health. 2009;1(1):005–10.
Kibore B, Gitao C, Sangula A, Kitala P. Foot and mouth disease sero-prevalence in cattle in Kenya. J Vet Med Anim Healthcare (Basel). 2014. https://doi.org/10.5897/JVMAH2013.0220.
Tekleghiorghis T, Weerdmeester K, van Hemert-Kluitenberg F, Moormann RJM, Dekker A. Foot-and-mouth disease seroprevalence in cattle in Eritrea. Transbound Emerg Dis. 2017;64(3):754–63. https://doi.org/10.1111/tbed.12434.
Chepkwony EC, Gitao GC, Muchemi GM, Sangula AK, Kairu-Wanyoike SW. Epidemiological study on foot-and-mouth disease in small ruminants: sero-prevalence and risk factor assessment in Kenya. PLoS ONE. 2021;16(8): e0234286. https://doi.org/10.1371/journal.pone.0234286.
Auty H, Mellor D, Gunn G, Boden LA. The risk of foot and mouth disease transmission posed by public access to the countryside during an outbreak. Front Vet Sci. 2019;6:381. https://doi.org/10.3389/fvets.2019.00381.
K. Ilbeigi, S. Bokaie, S. Aghasharif, R. J. Soares Magalhães, and M. Rashtibaf, "Risk factors for recurrence of FMD outbreaks in Iran: a case-control study in a highly endemic area," BMC Veterinary Research, vol. 14, no. 1, p. 253, 2018/08/29 2018, doi: https://doi.org/10.1186/s12917-018-1580-3.
Souley Kouato B, et al. Review of epidemiological risk models for foot-and-mouth disease: implications for prevention strategies with a focus on Africa. PLoS ONE. 2018;13(12):e0208296. https://doi.org/10.1371/journal.pone.0208296.
Ularamu HG, et al. Characterization of foot-and-mouth disease viruses collected in Nigeria Between 2007 and 2014: evidence for epidemiological links between West and East Africa. Transbound Emerg Dis. 2017;64(6):1867–76. https://doi.org/10.1111/tbed.12584.
Ramirez-Carvajal L, et al. Genetic stability of foot-and-mouth disease virus during long-term infections in natural hosts. PLoS ONE. 2018;13(2): e0190977. https://doi.org/10.1371/journal.pone.0190977.
Abdul-Hamid NF, Fırat-Saraç M, Radford AD, Knowles NJ, King DP. Comparative sequence analysis of representative foot-and-mouth disease virus genomes from Southeast Asia. Virus Genes. 2011;43(1):41–5. https://doi.org/10.1007/s11262-011-0599-3.
Kassaw K, Afera B, Amasalu K, Hussien D. Serotype identification and molecular characterization of foot and mouth disease in and around Mekelle, Tigray region. Vet. 2013;11:390–4.
Cottam EM, et al. Transmission pathways of foot-and-mouth disease virus in the United Kingdom in 2007. PLoS Pathog. 2008;4:e1000050. https://doi.org/10.1371/journal.ppat.1000050.
Belsham GJ. Towards improvements in foot-and-mouth disease vaccine performance. Acta Vet Scand. 2020;62(1):20. https://doi.org/10.1186/s13028-020-00519-1.
Pybus OG, Rambaut A. Evolutionary analysis of the dynamics of viral infectious disease. Nat Rev Genet. 2009;10(8):540–50. https://doi.org/10.1038/nrg2583.
Naqvi SS, et al. Evolutionary dynamics of foot and mouth disease virus serotype a and its endemic sub-lineage A/ASIA/Iran-05/SIS-13 in Pakistan. Viruses. 2022. https://doi.org/10.3390/v14081634.
Knowles NJ, Wadsworth J, Bachanek-Bankowska K, King DP. VP1 sequencing protocol for foot and mouth disease virus molecular epidemiology. Rev Sci Tech. 2016;35(3):741–55. https://doi.org/10.20506/rst.35.3.2565.
Hassan AM, et al. Molecular characterization of newly emerging foot-and-mouth disease virus serotype SAT 2 of Lib-12 Lineage Isolated from Egypt. Virus Res. 2022;311:198651.
Hagag NM, et al. Molecular detection and phylogenetic analysis of newly emerging foot-and-mouth disease virus type A, Lineage EURO-SA in Egypt in 2022. Virus Res. 2022;323:198960. https://doi.org/10.1016/j.virusres.2022.198960.
Subramaniam S, et al. Foot-and-mouth disease status in India during the second decade of the twenty-first century (2011–2020). Vet Res Commun. 2022;46(4):1011–22. https://doi.org/10.1007/s11259-022-10010-z.
Hassan AM, et al. Molecular detection, phylogenetic analysis and genetic diversity of recently isolated foot-and-mouth disease virus serotype A African topotype Genotype IV. Virol J. 2022;19(1):1. https://doi.org/10.1186/s12985-021-01693-y.
Ayelet G, et al. Genetic characterization of foot-and-mouth disease viruses, Ethiopia, 1981–2007. Emerg Infect Dis. 2009;15(9):1409–17. https://doi.org/10.3201/eid1509.090091.
Ranaweera LT, et al. Characterization of the FMDV-serotype-O isolates collected during 1962 and 1997 discloses new topotypes, CEY-1 and WCSA-1, and six new lineages. Sci Rep. 2019;9(1):14526. https://doi.org/10.1038/s41598-019-51120-0.
Le VP, Vu TT, Duong HQ, Than VT, Song D. Evolutionary phylodynamics of foot-and-mouth disease virus serotypes O and A circulating in Vietnam. BMC Vet Res. 2016;12(1):269. https://doi.org/10.1186/s12917-016-0896-0.
Li F, et al. Molecular evolution, diversity, and adaptation of foot-and-mouth disease virus serotype O in Asia. Front Microbiol. 2023;14:1147652. https://doi.org/10.3389/fmicb.2023.1147652.
Aman E, Molla W, Gebreegizabher Z, Jemberu WT. Spatial and temporal distribution of foot and mouth disease outbreaks in Amhara region of Ethiopia in the period 1999 to 2016. BMC Vet Res. 2020;16(1):185. https://doi.org/10.1186/s12917-020-02411-6.
Lloyd-Jones K, et al. Genetic and antigenic characterization of serotype O FMD viruses from East Africa for the selection of suitable vaccine strain. Vaccine. 2017;35(49):6842–9. https://doi.org/10.1016/j.vaccine.2017.10.040.
Benjamin B et al. Direct economic impacts of a foot-and-mouth (FMD) disease incursion in Australia, An update of ABARES 2013 estimate. ABARES, https://www.agriculture.gov.au/abares/research-topics/biosecurity/biosecurity-economics/fmd-update-of-2013-estimate (accessed 30 November 2023, 2023)
Thompson D, et al. Economic costs of the foot and mouth disease outbreak in the United Kingdom in 2001. Rev Sci Tech. 2002;21(3):675–87. https://doi.org/10.20506/rst.21.3.1353.
Blake A, Sinclair MT, Sugiyarto G. Quantifying the impact of foot and mouth disease on tourism and the UK economy. Tour Econ. 2003;9(4):449–65.
Pendell DL, Cho C. Stock market reactions to contagious animal disease outbreaks: an event study in Korean foot-and-mouth disease outbreaks. Agribusiness. 2013;29(4):455–68.
Baluka SA. Economic effects of foot and mouth disease outbreaks along the cattle marketing chain in Uganda. Vet World. 2016;9(6):544–53. https://doi.org/10.14202/vetworld.2016.544-553.
Sinkala Y, et al. Challenges and economic implications in the control of foot and mouth disease in sub-Saharan Africa: lessons from the Zambian experience. Vet Med Int. 2014. https://doi.org/10.1155/2014/373921.
Abao LN, Kono H, Gunarathne A, Promentilla RR, Gaerlan MZ. Impact of foot-and-mouth disease on pork and chicken prices in Central Luzon, Philippines. Prev Vet Med. 2014;113(4):398–406. https://doi.org/10.1016/j.prevetmed.2013.12.005.
Marschik T, et al. The epidemiological and economic impact of a potential foot-and-mouth disease outbreak in Austria. Front Vet Sci. 2020;7: 594753. https://doi.org/10.3389/fvets.2020.594753.
Pal M. Foot and mouth disease: a highly infectious viral zoonosis of global importance. J Appl Microbiol Biochem. 2018;2(3):12.
Jemberu WT, Mourits MC, Woldehanna T, Hogeveen H. Economic impact of foot and mouth disease outbreaks on smallholder farmers in Ethiopia. Prev Vet Med. 2014;116(1–2):26–36. https://doi.org/10.1016/j.prevetmed.2014.06.004.
Kamel M, El-Sayed A, Castañeda Vazquez H. Foot-and-mouth disease vaccines: recent updates and future perspectives. Arch Virol. 2019;164(6):1501–13. https://doi.org/10.1007/s00705-019-04216-x.
Paton DJ, Sumption KJ, Charleston B. Options for control of foot-and-mouth disease: knowledge, capability and policy. Philos Trans Royal Soc B Biol Sci. 2009;364(1530):2657–67.
FAO. The Global Foot and Mouth Disease Control Strategy. OIE and FAO. https://www.fao.org/3/an390e/an390e.pdf (accessed 30 November 2023, 2023).
El Bagoury GF, Elhabashy R, Mahmoud AH, Hagag NM, El Zowalaty ME. Development and evaluation of one-step real-time RT-PCR assay for improved detection of foot-and-mouth disease virus serotypes circulating in Egypt. J Virol Methods. 2022;306: 114525. https://doi.org/10.1016/j.jviromet.2022.114525.
Radostits OM, Gay C, Hinchcliff KW, Constable PD. A textbook of the diseases of cattle, horses, sheep, pigs and goats. Vet Med. 2007;10:2045–50.
Upadhayay U, Ewam P. Epidemiology and diagnosis of foot-and-mouth disease: a review. Indian J Anim Sci. 2012;6:543–51.
Wong CL, Yong CY, Ong HK, Ho KL, Tan WS. Advances in the diagnosis of foot-and-mouth disease. Front Vet Sci. 2020;7:477. https://doi.org/10.3389/fvets.2020.00477.
Admassu B, Getnet K, Shite A, Mohammed S. Review on foot and mouth disease: distribution and economic significance. Acad J Anim Dis. 2015. https://doi.org/10.5829/idosi.ajad.2015.4.3.95165.
Mahmoud M, Ghazy AA, Shaapan RM. Diagnosis and control of foot and mouth disease (FMD) in dairy small ruminants; sheep and goats. Int J Dairy Sci. 2019;14:45–52.
Souf S. Recent advances in diagnostic testing for viral infections Bioscience Horizons. Int J Stud Res. 2016;9:hzw010.
Arzt J, Juleff N, Zhang Z, Rodriguez LL. The pathogenesis of foot-and-mouth disease I: viral pathways in cattle. Transbound Emerg Dis. 2011;58(4):291–304. https://doi.org/10.1111/j.1865-1682.2011.01204.x.
Tewari A, et al. Development and validation of confirmatory foot-and-mouth disease virus antibody ELISAs to identify infected animals in vaccinated populations. Viruses. 2021. https://doi.org/10.3390/v13050914.
Brocchi E, et al. Comparative evaluation of six ELISAs for the detection of antibodies to the non-structural proteins of foot-and-mouth disease virus. Vaccine. 2006;24(47–48):6966–79. https://doi.org/10.1016/j.vaccine.2006.04.050.
de Sousa Lacerda CM, Almeida NBF, Dos Santos VCF, Plentz F, de Andrade ASR. Foot-and-mouth disease virus: DNA aptamer selection for the 3ABC protein. Virus Res. 2022;323:199008. https://doi.org/10.1016/j.virusres.2022.199008.
Mignaqui AC, et al. Foot-and-mouth disease: optimization, reproducibility, and scalability of high-yield production of virus-like particles for a next-generation vaccine. Front Vet Sci. 2020;7:601. https://doi.org/10.3389/fvets.2020.00601.
Stenfeldt C, Arzt J. The carrier conundrum; A review of recent advances and persistent gaps regarding the carrier state of foot-and-mouth disease virus. Pathogens. 2020. https://doi.org/10.3390/pathogens9030167.
Zhu JJ, et al. Inferred causal mechanisms of Persistent FMDV infection in cattle from differential gene expression in the nasopharyngeal mucosa. Pathogens. 2022. https://doi.org/10.3390/pathogens11080822.
Han L, et al. Cellular response to persistent foot-and-mouth disease virus infection is linked to specific types of alterations in the host cell transcriptome. Sci Rep. 2018;8(1):5074. https://doi.org/10.1038/s41598-018-23478-0.
Seyoum W, Tora E. Foot and mouth disease in Ethiopia: a systematic review and meta-analysis in the last fifteen years (2007–2021). Bull Nat Res Centre. 2023;47(1):1–14.
OIE. "Manual: Risk analysis for Foot and Mouth Disease." World Organisation for Animal Health. https://rr-asia.woah.org/wp-content/uploads/2020/02/seacfmd-manual-1.pdf (accessed 01 December 2023, 2023)
Iriarte MV, Gonzáles JL, de Freitas Costa E, Gil AD, de Jong MCM. Main factors associated with foot-and-mouth disease virus infection during the 2001 FMD epidemic in Uruguay. Front Vet Sci. 2023;10:1070188. https://doi.org/10.3389/fvets.2023.1070188.
Meng M, Li Y, Yao H. Plasmid-mediated transfer of antibiotic resistance genes in soil. Antibiotics (Basel). 2022. https://doi.org/10.3390/antibiotics11040525.
Alexandersen S, Mowat N. Foot-and-mouth disease: host range and pathogenesis. Curr Top Microbiol Immunol. 2005;288:9–42. https://doi.org/10.1007/3-540-27109-0_2.
Heshmati L, et al. Immunity evaluation of an experimental designed nanoliposomal vaccine containing FMDV Immunodominant peptides. Arch Razi Inst. 2021;76(5):1183–90. https://doi.org/10.22092/ari.2021.352498.1566.
Peta FRM, et al. Evaluation of potency and duration of immunity elicited by a multivalent FMD vaccine for use in South Africa. Front Vet Sci. 2021;8: 750223. https://doi.org/10.3389/fvets.2021.750223.
Ranjitha HB, et al. Thermostable negative-marker foot-and-mouth disease virus serotype O induces protective immunity in guinea pigs. Appl Microbiol Biotechnol. 2023;107(4):1285–97. https://doi.org/10.1007/s00253-023-12359-w.
Ayelet G, et al. FMD virus isolates: the candidate strains for polyvalent vaccine development in Ethiopia. Acta Trop. 2013;126(3):244–8. https://doi.org/10.1016/j.actatropica.2013.02.005.
Tesfaye Y, et al. A vaccine-matching assessment of different genetic variants of serotype O foot-and-mouth disease virus isolated in Ethiopia between 2011 and 2014. Arch Virol. 2020;165(8):1749–57. https://doi.org/10.1007/s00705-020-04662-y.
Jemberu WT, Molla W, Dagnew T, Rushton J, Hogeveen H. Farmers’ willingness to pay for foot and mouth disease vaccine in different cattle production systems in Amhara region of Ethiopia. PLoS ONE. 2020;15(10): e0239829. https://doi.org/10.1371/journal.pone.0239829.
Marrero R, Seki C, Mattion NM, König GA. Functional and in silico characterization of neutralizing interactions between antibodies and the foot-and-mouth disease virus immunodominant antigenic site. Front Vet Sci. 2021;8: 554383. https://doi.org/10.3389/fvets.2021.554383.
GFRA, EUFMD, Oa FAO. "Vaccination Against FMD – Principles and Practice. Joint virtual workshop for Asia and South East Asia on vaccination against FMD." https://www.ars.usda.gov/GFRA/events/2020%20EUFMD/REPORT%20SEA%20EUFMD%20GFRA%20MEETING%202020.pdf (accessed 01 December 2023, 2023).
Capozzo AV, Vosloo W, de Los Santos T, Pérez AM, Pérez-Filgueira M. Editorial: Foot-and-mouth disease epidemiology, vaccines and vaccination: moving forward. Front Vet Sci. 2023;10:1231005. https://doi.org/10.3389/fvets.2023.1231005.
Hardham JM, et al. Novel foot-and-mouth disease vaccine platform: formulations for safe and DIVA-compatible FMD vaccines with improved potency. Front Vet Sci. 2020;7: 554305. https://doi.org/10.3389/fvets.2020.554305.
Ozturk N, Kocak O, Vosough Ahmadi B. Economic Analysis of increasing foot-and-mouth disease vaccination frequency: the case of the biannual mass vaccination strategy. Front Vet Sci. 2020;7:557190. https://doi.org/10.3389/fvets.2020.557190.
Li J, Chang Y, Yang S, Zhou G, Wei Y. Formulation enhanced the stability of Foot-and-mouth virus and prolonged vaccine storage. Virol J. 2022;19(1):207. https://doi.org/10.1186/s12985-022-01928-6.
Kamel M, El-Sayed A, Castañeda Vazquez H. Foot-and-mouth disease vaccines: recent updates and future perspectives. Arch Virol. 2019;164:1501–13. https://doi.org/10.1007/s00705-019-04216-x.pdf.
Kompas T, Ha PV, Nguyen HT, Garner G, Roche S, East I. "Optimal surveillance against foot-and-mouth disease: A sample average approximation approach. PLoS ONE. 2020;15(7): e0235969. https://doi.org/10.1371/journal.pone.0235969.
Howson ELA, et al. Direct detection and characterization of foot-and-mouth disease virus in East Africa using a field-ready real-time PCR platform. Transbound Emerg Dis. 2018;65(1):221–31. https://doi.org/10.1111/tbed.12684.
Lyons NA, et al. Considerations for design and implementation of vaccine field trials for novel foot-and-mouth disease vaccines. Vaccine. 2019;37(8):1007–15. https://doi.org/10.1016/j.vaccine.2018.12.064.
Park SY, et al. Efficient removal of non-structural protein using chloroform for foot-and-mouth disease vaccine production. Vaccines (Basel). 2020. https://doi.org/10.3390/vaccines8030483.
He Y, et al. Structures of foot-and-mouth disease virus with bovine neutralizing antibodies reveal the determinant of intraserotype cross-neutralization. J Virol. 2021;95(24):e0130821. https://doi.org/10.1128/jvi.01308-21.
Domenech J, Ferrari G, Leboucq N, Todeschini B, Metwally S, De Leeuw P The global foot and mouth disease control strategy. In: The FAO/OIE Global Conference on Foot and Mouth Disease Control, 2012, p 211
Acknowledgements
Nothing to declare.
Funding
This review did not receive any funding.
Author information
Authors and Affiliations
Contributions
GZ, MA, WT, HB, GD, MZ, and YT contributed to the review and writing of the paper. GZ wrote the first draft, and all authors gave feedback on subsequent versions. All authors also participated in the study design and manuscript revision.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
No animal experiments were involved in this article.
Consent for publication
Not applicable.
Competing interests
The authors have no financial or personal conflicts that could bias the review in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

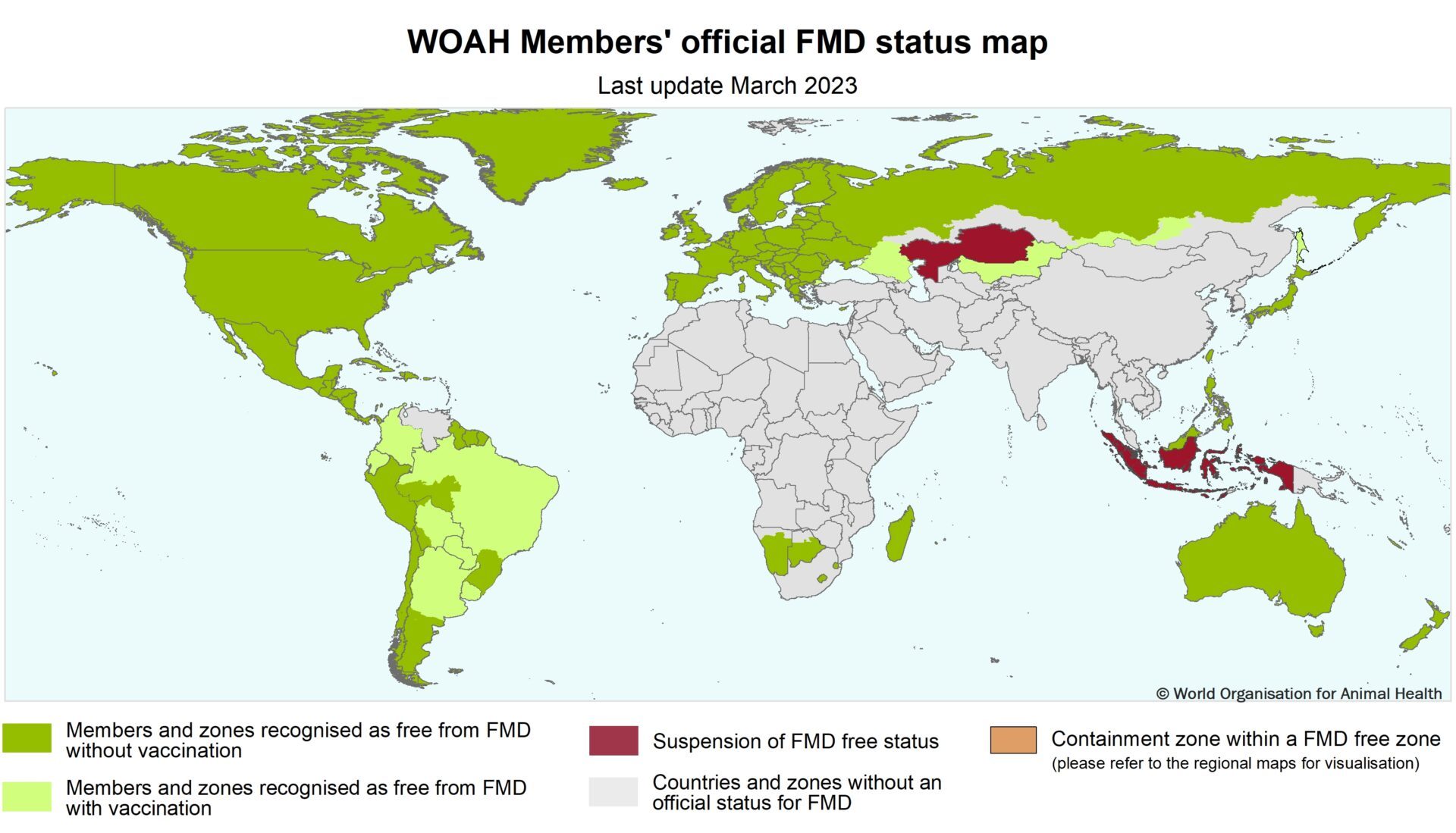{kind=link}
{kind=link}
Cite this article
Zewdie, G., Akalu, M., Tolossa, W. et al. A review of foot-and-mouth disease in Ethiopia: epidemiological aspects, economic implications, and control strategies. Virol J 20, 299 (2023). https://doi.org/10.1186/s12985-023-02263-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12985-023-02263-0