
Abstract
At first glance, longevity and immunity appear to be different traits that have not much in common except the fact that the immune system promotes survival upon pathogenic infection. Substantial evidence however points to a molecularly intertwined relationship between the immune system and ageing. Although this link is well-known throughout the animal kingdom, its genetic basis is complex and still poorly understood. To address this question, we here provide a compilation of all genes concomitantly known to be involved in immunity and ageing in humans and three well-studied model organisms, the nematode worm Caenorhabditis elegans, the fruit fly Drosophila melanogaster, and the house mouse Mus musculus. By analysing human orthologs among these species, we identified 7 evolutionarily conserved signalling cascades, the insulin/TOR network, three MAPK (ERK, p38, JNK), JAK/STAT, TGF-β, and Nf-κB pathways that act pleiotropically on ageing and immunity. We review current evidence for these pathways linking immunity and lifespan, and their role in the detrimental dysregulation of the immune system with age, known as immunosenescence. We argue that the phenotypic effects of these pathways are often context-dependent and vary, for example, between tissues, sexes, and types of pathogenic infection. Future research therefore needs to explore a higher temporal, spatial and environmental resolution to fully comprehend the connection between ageing and immunity.
Similar content being viewed by others
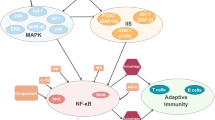

Background
Organisms are constantly challenged by various types of pathogens, which inflict numerous negative effects on health and fitness of infected hosts. A speedy resolution of pathogenic infections thus limits their deleterious consequences and is consequently beneficial for survival, providing an adaptive advantage. Hosts achieve immunity through a variety of mechanisms. Foremost, the innate and adaptive immune systems represent the two major routes of promoting resistance to infectious organisms. Immune system activation generally triggers the expression of cytotoxic molecules, such as antimicrobial peptides (AMPs) or reactive oxygen species (ROS), and the recruitment of specialized immune cells to the infected tissue that inhibit and destroy microbial intruders. In addition, hosts alter their own physiological state upon infection to create an undesirable environment for pathogens. Although these defence mechanisms support survival in the presence of pathogens, maintaining and mounting an immune response can also be associated with two types of costs. First, physiological trade-offs between immunity and other metabolically expensive traits, in particular growth and reproduction, have been identified across the tree of life, including plants [1], insects [2], and vertebrates [3]. Second, because immune mechanisms are not entirely specific to pathogens, they cause adverse side-effects, for instance, by harming the host tissue. Organisms thus need to find a delicate balance between allocation of limited resources and the intensity of the immune response to optimize their evolutionary fitness.
The adverse properties of immunity become particularly apparent as organisms age. The immune system and the inflammatory response are subject to strong age-dependent loss of homeostasis, resulting in age-related immunopathology known as immunosenescence – a term first coined by Roy Walford in the 1960s [4]. The decline in normal immune function manifests in a multitude of effects detrimental for health and longevity, including increased susceptibility to pathogens, decreased vaccine-response, chronic inflammation or ‘inflammaging’, impaired wound healing, and a higher incidence of cancer [5, 6]. Age-associated increases in auto-immunity factors, such as self-reacting auto-antibodies, are also common but do not necessarily cause a higher incidence of auto-immune diseases in elderly, potentially due to enhanced protective mechanisms which occur in parallel [7]. Interestingly, almost all species including vertebrates and invertebrates are subject to immunosenescence, even though they vary vastly in their immune systems, life history, lifespans, and ecological niches. Figure 1 summarises the components and pathways involved in the immune system of the four different organisms. It is well known that survival upon pathogen exposure declines with age at infection in the nematode worm Caenorhabditis elegans [8, 9], the fruit fly Drosophila melanogaster [10, 11], and several mammals [12,13,14], demonstrating that immunosenescence affects both the innate and adaptive immune system. Counterintuitively, the reduced pathogen resistance is accompanied by an increase in basal expression levels of several innate immunity and inflammatory genes, such as antimicrobial peptides, cytokines, and complement system factors in flies [15, 16], mice [17, 18], and humans [6, 19], although this has not been reported in C. elegans to our knowledge. Thus, even though baseline immunity is higher in older individuals, realized immunity, in terms of pathogen defence, depreciates. Moreover, this ramping up of baseline immunity with age is thought to be the main reason for the deleterious, pro-inflammatory state of inflammaging [5, 6]. In parallel to this, innate immunity cells decline in phagocytic ability with age in D. melanogaster [20] and mammals [6], while changes in the adaptive immune system are characterized by a depletion in naïve cells and a rise in memory cells [5, 6], theoretically making the organism less adaptable to novel antigens. The exhaustion in naïve immune cells with age makes the host reliant on memory cells, which are specific to pathogens encountered in the past, while defence against novel antigens is weakened. This depletion is thought to be driven by two major factors: (1) the genetically programmed decline in mass of the thymus where T-cells mature, known as thymic involution, and (2) life-long accumulated antigenic exposure diminishing naïve immune cells [5, 6].

Major differences in immunity of C. elegans, D. melanogaster and mammals. Invertebrates, such as nematodes and insects, represented by C. elegans and D. melanogaster respectively, completely rely on innate immunity upon infection, while vertebrates (represented by mammals) have also evolved an adaptive immune system. Cellular immunity occurs in fruit flies as hemocytes (immune cells in the hemolymph), and as different types of leukocytes in mammals, but is absent in worms. In contrast to this, the germ-line encoded complement system is unique to mammals. Moreover, pattern recognition receptors (PRRs) of the innate immune system – most prominently the Toll- and Toll-like receptors, are incremental for pathogen-recognition in fruit flies and mammals, but whether the single Toll-like receptor homolog in C. elegans fulfils the same function is still under debate. Finally, Nf-κB transcription factors, which regulate the expression of immune-related genes, are central to D. melanogaster and mammalian immunity, but have not been identified in C. elegans. The well-known cross-talk between immune and JNK MAPK signalling is indicated, but notably several other pathway interactions do also exist. Immunity genes and protein complexes also associated with lifespan and ageing are marked in green and bold
It has also become apparent that many age-associated diseases, such as Alzheimer’s, Parkinson’s, diabetes, cancer, and atherosclerosis, exhibit a significant immunological and inflammatory component [21,22,23,24], as further corroborated by studies in model organisms (e.g. [25,26,27]). Identifying the interlinked genes and pathways in different species, which contribute pleiotropically to phenotypes associated with ageing, lifespan and immunity, could help to identify common molecular mechanisms of the immunity-ageing cross-talk.
In this review, we utilise public databases and inhouse manual curation to assemble a resource of genes known to affect both immunity and ageing (from here on called ‘immuno-ageing’ factors) in two invertebrates, C. elegans and D. melanogaster, and two mammalian species, the house mouse (Mus musculus) and humans (Homo sapiens). We then identify conserved genes between these species based on human orthologs and associate them to core immuno-ageing pathways, which we discuss in this review. We finish by outlining open questions and necessary experiments to further understand the complex relationship between ageing and immunity.
Evolutionarily conserved immuno-ageing factors
To classify pathways likely mediating the cross-talk between ageing and immunity, we first combined all genes within the Gene Ontology (GO) and KEGG terms related to immunity or ageing, and further included annotations from two ageing (GenAge [28], AgeFactDB [29]) and three immunity databases (insect innate immunity database IIIDB [30], InnateDB [31], immunome knowledge base IKB [32]), resulting in an ‘ageing’ and ‘immunity’ gene list for each species. The definition of ageing and immunity genes varies among these databases, but broadly, the function of the genes in our lists are derived from experimental evidence and computational inference. Next, we identified pleiotropic immuno-ageing genes by intersecting the ageing and immunity gene lists of each species (Fig. 2). Details on gene list construction, number of genes per database, and lists of identified ageing, immunity and immuno-ageing genes across the four species are available in Table S1. We obtained different numbers of ageing, immunity and immuno-ageing genes across the four species, which could be driven by biological differences, such as varying number of genes modulating ageing and immunity between the species or the total number of genes in each species. From a technical point of view, two major caveats of our approach, that also influence these differences, are study-preferences for either ageing or immune function in a particular model, and researcher-biases in studying specific genes and pathways considered more important, which has previously been shown to influence the analyses of ageing-related genes [33].

Overlap in 437 Human immuno-ageing orthologs between C. elegans (blue), D. melanogaster (green), M. musculus (orange), and H. sapiens (red). Orthologs were obtained using DIOPT. Details on shared orthologs, original gene names and methods are given in Table S2
Next, we asked if there are any evolutionarily conserved immuno-ageing genes, which might be interesting candidates for modulating ageing, lifespan and immunity across evolutionarily distant species, and identified in total 437 orthologs using DIOPT [34] based on human genes (Fig. 2 and Table S2). To further identify the most conserved genes, we overlapped orthologs across the four species. As expected, the overlap between mammals and invertebrates is low, reflecting in part the lack of adaptive immunity in flies and worms. Only ten highly conserved immuno-ageing genes were common in all four species. Of these, the six genes akt-1/Akt1/AKT2, age-1/Pi3K92E/PIK3CD, daf-2/InR/IGF1R, daf-16/foxo/FOXO3, let-363/Tor/MTOR, and rsks-1/S6k/RPS6KB2 (order of gene names: C. elegans / D. melanogaster / mammals) are members of the insulin/insulin-like growth factor signalling (IIS) and the target-of-rapamycin (TOR) pathways and are well-known for their conserved roles in ageing (Fig. 3). The other four genes in this category act within the ERK and p38 MAPK pathways and include the mpk-1/rl/MAPK1, pmk-1/p38a and b/MAPK14, mek-2/Dsor1/MAP2K1, and let-60/Ras85D/HRAS (Fig. 4). While the ERK MAPK pathway mainly gets activated through IIS and growth factors and promotes proliferation, the p38 MAPK pathway is mostly known for its response to environmental stresses [35]. Inspection of triple and double overlaps between invertebrates and mammals included further IIS/TOR network components, as well as genes related to the JNK MAPK pathway (Fig. 4), which performs similar downstream functions as the p38 cascade [35]. Moreover, we found genes related to the immuno-supporting JAK/STAT [36, 37], and the TGF-β pathway, which is thought to have a great diversity of context-dependent roles in cellular and physiological function, including immunity [38, 39] (Fig. 4). In addition, the Nf-κB signalling components were shared between fruit flies and mammals, since this pathway is thought to be absent in C. elegans (Fig. 1). In general, the overlaps between invertebrates and mammals comprised genes acting broadly across the signalling transduction, such as ligands, receptors and transcription factors. We also found some noteworthy genes partially controlled by Foxo transcription factors, including sod-3/Sod2/SOD2 and ctl-1/Cat/CAT which are involved in the clearance of oxidative stress, atm-1/tefu/ATM that functions in DNA repair and telomere maintenance, and the antiviral gene dcr-1/Dcr-2/DICER1 (see discussion below).

Simplified overview of the insulin-insulin-like growth factor and TOR signalling (IIS/TOR) network. Activation of IIS/TOR network signalling is induced by insulin and insulin-like peptides, environmental cues and nutrients, and leads to a signalling cascade promoting growth and reducing lifespan. In absence of IIS/TOR signalling, such as under dietary restriction, Foxo transcription factors and the 4E-BP inhibitor of translation inhibit growth and promote longevity. Factors stimulating IIS/TOR signalling and thought to reduce lifespan are shown in red, those generally beneficial for lifespan in blue. Gene names are given in the white boxes in the order of C. elegans, D. melanogaster and H. sapiens (overall, equal to M. musculus). Genes with pleiotropic effects on immunity are marked in green and bold

MAPK, JAK/STAT and TGF-β pathways regulate ageing and immunity. Multiple signalling cascades in addition to the IIS and TOR pathways contribute to ageing, longevity and immunity. Three MAPK pathways are conserved in worms, flies and mammals. The ERK-MAPK pathway is activated by growth and other factors through binding to receptors, including EGFR, and can further be triggered by the IIS pathway. Activity leads to transcription factors mainly promoting growth, cell division and differentiation of cells. In contrast, p38 and JNK MAPK pathways are classically considered to respond to stress including infections, while the JNK cascade is also mediated by Nf-κB signalling (Fig. 1). The multiple transcription factors downstream of p38 and JNK stimulate inflammation, immunity, longevity and other functions. JAK/STAT signalling is triggered by upd cytokines in D. melanogaster, and interferons, multiple cytokines and other factors in mammals. Upon receptor binding, the JAK/STAT signalling cascade leads to activation of STAT transcription factors, which are best known for fostering inflammation and immunity, particularly against viral infections. Notably, JAK/STAT in C. elegans only includes the two STAT transcription factors, sta-1 and sta-2, but does not share the same upstream factors as fruit flies and mammals. Finally, the branches of TGF-β pathways are activated by BMPs, activins and other factors. The signalling cascade regulates multiple transcription factors, leading to immunosuppression, anti-inflammatory responses and several other physiological effects. Notably, the pathways show various amounts of cross-regulation among each other but also with the IIS/TOR network and the Nf-κB cascade, and for simplicity, only well-known cross-talk is indicated. Gene names are given in the white boxes in the order of C. elegans, D. melanogaster and H. sapiens (overall, equal to M. musculus). Genes and factors concomitantly involved in lifespan, ageing, and immunity are marked in green and bold
In summary, our analysis suggests that the IIS/TOR network together with ERK/p38/JNK MAPK, JAK/STAT, TGF-β, and the Nf-κB pathways have highly conserved mechanisms in immunity, lifespan and ageing. In the remainder of the review, we shall explore the roles of each of these pathways on the immunity-lifespan cross-talk and immunosenescence as revealed in studies on the different organisms.
The IIS/TOR network is a key regulator of ageing and immunity
Invertebrate IIS/TOR network
The nutrient-sensing IIS/TOR signalling network is among the best understood molecular determinants of ageing and lifespan. Reducing the activity of IIS and/or TOR signalling through genetic or nutritional interventions (e.g. dietary restriction) has long been established to prolong lifespan in yeast, nematodes, insects, and mammals [40,41,42].
Pioneered by studies in C. elegans, a major role of the IIS/TOR network in immunity has been demonstrated in addition to regulating lifespan. A seminal discovery establishing this connection is that long-lived loss-of-function mutants of daf-2, encoding the insulin-like growth factor 1 (IGF-1) receptor, are more resistant to gram-positive and gram-negative bacteria [43,44,45] (Fig. 3). These results are further supported by the observation that bacterial infection of the pharynx, thought to be a major cause of early death in C. elegans [46], was less severe in daf-2 loss-of-function mutants during ageing than in wildtype worms [47]. Increased pathogen resistance has also been shown for phosphatidyl-inositol 3 kinase (PI3K) AGE-1 null mutants [43, 45, 48], while the downstream forkhead transcription factor DAF-16, which is suppressed upon IIS activation, is usually conferring pathogen resistance [43, 45, 49, 50].
Comparable to worms, downregulating insulin signalling through loss-of-function of the insulin receptor substrate gene chico in D. melanogaster (Fig. 3) is beneficial for longevity and increases resistance to pathogenic bacteria, although effects vary dependent on the pathogens or the chico null allele used [51,52,53]. Surprisingly, AMP expression in chico mutants is generally decreased or equal to that in wildtype flies, which represents a poorly understood mismatch between potential and realized immunity [52, 53]. Moreover, similar to worms, lowered foxo expression has negative consequences on survival upon infection potentially because of a concurrent downregulation in AMPs [54,55,56].
The immunity modulating features further extend to the TOR pathway. Reduced TOR pathway activity has been associated with improved lifespan and enhanced immunity by promoting autophagy in C. elegans [57,58,59] and inhibiting protein translation in D. melanogaster [60, 61]. However, the effects of TOR in fruit flies are not always clear-cut. While induction of AMP expression independent of the Nf-κB pathway and improved phagocytic ability of hemocytes have been observed upon lowered TOR pathway activity in D. melanogaster, it may deter bacterial clearance at the same time [62, 63].
The similar links between IIS/TOR signalling and pathogen defence in D. melanogaster and C. elegans might even extend to immunity against viruses. In invertebrates, the RNAi pathway confers immunity against viruses, and in worms further contributes to bacterial infection [64,65,66,67,68] (Fig. 5). Several studies suggest that in C. elegans, reduced activity of insulin signalling causes enhancement of RNAi-mediated immunity, including transposable element silencing [69,70,71], which is also thought to be beneficial for longevity in fruit flies and mammals likely due to their neurodegenerative and inflammatory properties [72,73,74,75,76]. In parallel, it has been shown in arthropods that IIS/TOR activation acts pro-viral, whereas FOXO activity positively regulates the expression of antiviral RNAi genes, thereby increasing survival upon viral infection [77, 78]. Reflecting these findings, inhibition of multiple proteins of the IIS/TOR network was implicated in viral replication in mammals [79,80,81], but the role of RNAi in mammalian immunity has been controversial [82, 83]. Further supporting a pro-immunity and pro-longevity role of the RNAi pathway, RNAi loss-of-function mutants have a lower lifespan in C. elegans [84] and mice [85], and reduced lifespan, stress resistance and antiviral immunity in D. melanogaster [75, 86, 87]. In turn, a ubiquitously active RNAi pathway can have detrimental effects on lifespan [88], so that deviations from its wildtype homeostasis may be generally deleterious for ageing.

Antiviral RNAi pathway. The antiviral RNA interference (RNAi) pathway shows genetic and functional conservation across D. melanogaster, C. elegans and mammals. In both, C. elegans and D. melanogaster, antiviral RNAi is thought to be a major innate defence against viruses. However, even though antiviral RNAi exists in mammals, its importance is currently under debate as they contain other potent mechanisms of virus defence, such as the interferon response and adaptive immunity. Components also involved in lifespan in at least one of the represented species are marked in green and bold
Altogether, evidence so far converges on the notion that downregulation of the IIS/TOR network has not only positive effects on lifespan but also improves immune function in addition to its metabolic roles. Recent research in Lepidoptera species and mosquitoes shows that the connection between the IIS/TOR network and immunity is not confined to well-established model organisms [89,90,91]. Moreover, while the three main innate immunity pathways in fruit flies (Toll, IMD, and the antiviral RNAi pathway) are thought to confer resistance to certain types of pathogens, immunity through FOXO regulated genes appears to be general. The effect of the IIS/TOR network on lifespan, however, appears to be slightly more consistent throughout the literature than that for immunity, suggesting that the link might not be omnipresent. Improving lifespan through IIS/TOR may frequently, but not always, cause improved immunity rather than the other way round. Future studies analysing the effect of the IIS/TOR network on lifespan and immunity together in controlled experiments would be crucial to further disentangle the ‘cause-effect’ relationship between both traits.
Mammalian IIS/TOR network
Are the striking similarities of the IIS-TOR-immunity interplay found in C. elegans and insects found in organisms with an adaptive immune system? In parallel to invertebrates, reductions in growth hormone and the insulin signalling pathway were associated with enhanced lifespan and slowed immunosenescence through maintaining young T-cell profiles [92, 93] and decreasing memory relative to naïve T-cells [94, 95]. Indeed, mammalian Foxo transcription factors downstream of IIS/TOR are likely among the key components driving the immuno-ageing cross-talk. Analogous to C. elegans and D. melanogaster, one of the mammalian Foxo homologs Foxo3 is a pro-longevity factor, yet is required for survival upon chronic pathogenic infections, expression of antimicrobial peptides, and likely acts anti-inflammatory [96,97,98]. Although FOXO transcription factors are important mediators of ageing and immunity in mammals, their effects may vary dependent on the FOXO gene and cell type [99,100,101].
Similarly, TOR signalling is found to be necessary for the innate immune response in mammals, where the functions of the two conserved TOR protein complexes have been extensively studied. In agreement with observations in invertebrate models, inhibiting the mTORC2 complex leads to improved survival upon infection and facilitates expression of pro-inflammatory cytokines mediated by FOXO1 [102, 103]. In contrast, mTORC1 activity was linked to the expression of pro-inflammatory cytokines and type I IFN-γ proteins, which are essential molecules in pathogen resistance [104, 105]. Remarkably, opposing findings in D. melanogaster, knockout of two (of three) mammalian inhibitors of translation 4E-BP1 and 4E-BP2, which are inactivated by mTORC1 signalling, increased resistance to viruses by fostering cytokine production [81]. The fact that mTORC1 promotes innate immunity in mammals perhaps explains these contradictory results. Furthermore, there is a large body of work demonstrating the necessity of the TOR pathway in adaptive immunity, where it contributes to antigen presentation, immune cell activation, differentiation, and memory formation (as reviewed by [106]). For example, treating mice with the immunosuppressant drug rapamycin, which only inhibits the mTORC1 complex [107], increased their lifespan and the regenerative ability of hematopoietic stem cells, leading to higher numbers of B-lymphocytes and better survival upon Influenza infection in old individuals [108]. In agreement with this, mTOR inhibition in elderly humans enhanced influenza antibody titres post vaccination, which was concurrent with a reduction in PD-1-positive T-cells that are known to accumulate with age [109]. The fundamental links between the IIS/TOR pathway and immunosenescence are further supported by a tremendous amount of evidence that insulin-like peptide mediated signalling, PI3K activation, and several associated genes function not only in regulation of metabolism and ageing, but are also necessary for innate and adaptive immunity [110,111,112,113,114].
Due to the complexity of the mammalian IIS/TOR network, it is more problematic to generalize its exact effects on immunity and lifespan compared to invertebrates. This is nicely delineated by the different immunological functions of the two mTOR complexes explained above, but also encompasses other components, as demonstrated by the fact that FOXO1 and FOXO3 can have opposing properties supporting survival and apoptosis of T-cells, respectively [99]. A likely explanation for this complexity is repeated gene duplication patterns followed by functional divergence, the fast evolutionary rates of some IIS/TOR genes, and diversifying natural selection [115,116,117]. An alternative explanation might be that C. elegans and D. melanogaster were not studied as extensively with regards to the function of IIS/TOR on ageing and immunity in different tissues and treatments. Hypothetically, invertebrates might exhibit a similar functional complexity as mammals do, but it has not yet been detected. Besides nematodes, insects, mice and humans, not many other species have been investigated for an immuno-ageing interplay, although recent studies in fish revealed a cross-talk between immunity and metabolism through insulin-like peptide hormones [118, 119].
MAPK pathways variably affect ageing and immunity
Invertebrate ERK MAPK pathways
The ERK MAPK pathway is, like the IIS/TOR network, a central regulator of proliferation and cellular processes (Fig. 4). Thus, its activation might comparably mediate resource allocation to growth at the cost of lifespan and immunity. In contrast to this expectation, the ERK-MAPK pathway appears to promote lifespan and immunity in worms. It has been demonstrated that loss of ERK-MAPK activity resulted in reduced lifespan of C. elegans [120], while several genes along the ERK MAPK cascade were shown to promote defence against a bacterial pathogen [121,122,123,124]. Interestingly, this suggests that, in the presence of nutrients, activation of the ERK MAPK pathway in C. elegans antagonizes the detrimental effects of IIS/TOR signalling on lifespan and immunity.
Opposing findings in worms, ERK MAPK activation demonstrably reduces lifespan in fruit flies and leads to inhibition of the IMD/Nf-κB immunity pathway with reduced survival upon bacterial infection as a consequence [125, 126]. Nevertheless, the regulation of immunity conferred by this pathway could depend on the pathogen. Additional reports indicate that ERK MAPK activation restricts replication of viruses after oral infection in D. melanogaster, and this function could be conserved in Aedes mosquitos [127, 128].
Invertebrate p38 and JNK MAPK pathways
C. elegans further relies on the p38 MAPK cascade to mount an antimicrobial peptide response and for a functional immune system (Fig. 4) [120, 129, 130]. Similar to the ERK MAPK pathway, activation of the p38 MAPK signalling has positive effects on both pathogen resistance and lifespan in worms [120, 122, 124, 131,132,133,134,135]. However, its impact on lifespan is possibly context-dependent, as demonstrated by studies reporting that p38 MAPK signalling does either not affect lifespan [120, 130] or reduces it, possibly dependent on temperature or the mutant background [132, 136]. Supporting a complicated role in C. elegans lifespan, the p38 MAPK pathway is further required for lifespan extension by dietary restriction and reduced insulin signalling, while both of these treatments simultaneously reduce its activity [137].
Same as in C. elegans, the p38 MAPK pathway in D. melanogaster is involved in stress response and might be required for downregulation of immunity gene expression after infections [138]. In support of its role in immunity, p38 MAPK is important for resistance against bacteria, fungi, and DNA viruses, and further contributes to regulation of the immune system [139,140,141]. Importantly, p38 MAPK also controls fat and glycogen metabolism in the fat body, which is functionally similar to the mammalian liver, but also AMP expression under bacterial infection, thereby illustrating a case of immune-metabolic cross-regulation [142]. Still, how this pathway affects lifespan is not fully resolved, although recent results suggest a pro-longevity effect in muscle cells, while no effects were observed in neurons [143, 144]. In general, the trend implies similar pro-longevity and pro-immunity roles of the p38 MAPK pathway in C. elegans and D. melanogaster.
Analogously, the JNK-MAPK signalling pathway, known to stimulate stress resistance (Fig. 4), also promotes longevity and survival to bacterial infections in C. elegans, and this is at least partly caused by cross-talk with IIS/TOR and activation of DAF-16 [145,146,147]. This, however, only partly extends to fruit flies, where JNK activation generally extends lifespan although possibly dependent on the tissue [148,149,150]. Moreover, JNK is cross-talking with IMD and is required for the activation of antimicrobial peptides [151]. Despite this, reduced JNK signalling activity has been shown to increase survival upon P. entomophila infection [152]. This is akin of recent findings in Anopheles stephensi mosquitos, where reduced JNK activity led to elevated Plasmodium resistance [153].
Mammalian MAPK pathways
Mammals resemble the MAPK-driven regulation of immunity and ageing observed in C. elegans and D. melanogaster. Parallel to findings in fruit flies, knockout of the ERK pathway component gene RasGfr1 in mice had beneficial effects on lifespan, suggesting that ERK activation is limiting lifespan in mammals [154]. In line with this, long-lived dwarf mouse models have reductions in ERK and p38 MAPK signalling along with alleviated immunosenescence [92, 155, 156]. Indeed, p38 MAPK signalling can promote ageing of gut stem cells after activation through mTORC1 [157]. Yet, cardiac-specific knockout of p38⍺ (Mpk14) MAPK [158] and two MAPK pathway activators did not impact lifespan in mice [159], suggesting that other functionally redundant genes can compensate for the loss-of-function in this kind of studies.
Mammalian MAPK pathways are further not just involved in innate immunity but also contribute to T- and B-cell survival and activation, as well as inflammation and Nf-κB mediated transcription [160,161,162]. Yet, their impact on immunity is complex, as demonstrated by the negative effects of JNK1 (Mpk8) activation on survival upon fungal infection in mice, which was explained by a reduced production of nitric oxide defence molecules [163].
Similarly, inhibition of the p38 MAPK pathway increased the survival time of influenza infected mice [164] and comparably improved immunity to varicella zoster virus in the human skin [165], likely through reducing pro-inflammatory cytokine production and preventing an overreaction of the immune system also known as a ‘cytokine storm’. Consistent with these results, the ability to resolve inflammation is reduced by increasing p38 MAPK activity in mononuclear phagocytes of elderly humans, but can be restored upon pharmacological inhibition of p38 [166]. Moreover, deficiencies in DUSP genes, that inhibit MAPK pathways in a negative feedback loop, result in deregulation of the inflammatory response (as discussed by Arthur & Ley [167]).
Context-dependent effects of the JAK/STAT pathway on ageing and immunity
A further conserved immuno-ageing pathway is the JAK/STAT signalling cascade, which regulates numerous cellular processes, but is also central to immunity and the expression of cytokines [36, 168] (Fig. 4). Despite C. elegans lacking a homolog of JAK, it retains two functional STAT orthologs that play a role in the defence response to viruses and fungi and reduce lifespan upon knockout [168, 169]. Interestingly, the worm STAT orthologs possibly have opposing roles in immunity, and either promote infection response genes, such as AMPs, dependent on the p38 MAPK and TGF-β pathways [168] or repress them [169].
Comparable to C. elegans, JAK/STAT signalling regulates the expression of genes fighting pathogenic infections in D. melanogaster, with the difference that the pathway is activated by cytokine-like proteins [37]. Several lines of evidence suggest that activating JAK/STAT signalling is detrimental for lifespan, and the effect size might depend on diet and tissue [170,171,172]. This implies that JAK/STAT signalling possibly represents a pathway mediating a trade-off between immunity and lifespan, which is akin to the opposite effects of that STAT ortholog sta-1 on lifespan and immunity in C. elegans [169].
Further supporting a conserved role of JAK/STAT in immunity and ageing, loss-of-activity of JAK/STAT genes in mice has negative consequences on immunity, and causes defects in interferon signalling and increased susceptibility to viral infection [173]. In line with a trade-off between immunity and lifespan, administration of a JAK pathway inhibitor reduced inflammation and alleviated cellular senescence [174]. In addition, loss of expression of the JAK/STAT inhibitor Socs2 was associated with decreased lifespan in mice [175]. Opposite to this, the downstream transcription factor STAT3 was found to be protective against inflammation-induced heart damage, highlighting context-dependency of the immuno-ageing properties of JAK/STAT [176].
TGF-β signalling affects ageing and immunity dependent on context
The conserved TGF-β pathway acts as a key regulator of physiological homeostasis, as it controls a huge number of cellular functions including growth, differentiation, and apoptosis, often in a context-specific manner [177] (Fig. 4). Perhaps unsurprisingly, considering its diverse roles, it also helps to mediate the relationship between lifespan, ageing and immunity. In C. elegans, loss-of-function of the TGF-β homolog dbl-1 has negative effects on survival upon infection and longevity [178], suggesting positive effects on immunity and ageing. In contrast, TGF-β signalling activation in D. melanogaster is associated with a negative regulation of the immune response mediated by the infection/wounding-regulated genes dpp and daw [179]. These results might again be context-specific: upon infection with pathogenic nematodes, both of these genes were thought to promote survival, but the exact factors involved might vary between adults and larvae [180, 181]. While dpp and its downstream transcription factor Mad (BMP signalling) are considered to have anti-ageing functions, lifespan effects for daw, its receptor babo, and downstream factor Smox (Activin signalling) are variable [182, 183]. Nevertheless, TGF-β signalling might help maintaining protein homeostasis, suggested to be a central hallmark of ageing [184].
In mammals, TGF-β signalling is also known to activate MAPK pathways and is similarly crucial for regulating a vast amount of cellular processes, including survival and immunity [38, 39]. Apart of its multifaceted biological roles, TGF-β knockout mice exhibited higher inflammation, auto-immunity and reduced lifespan, and the detrimental effects were alleviated by further removing MHC class I functionality necessary for CD8+ T cell development [185]. As before, context-dependency of TGF-β exists because lifespan was reduced in females, but not in males in mice with impaired TGF-β in salivary glands [186].
Nf-κB signalling mediates a trade-off between immunity and ageing
While the immuno-ageing pathways outlined above are conserved between D. melanogaster and C. elegans, central parts of fruit fly and mammalian immunity signalling missing in worms are Nf-κB transcription factors (Fig. 1). Nf-κB signalling has central functions in innate and adaptive immunity, and is triggered upon activation of several immune receptors, most prominently Toll-like receptors, during pathogenic infection. Downstream, it results in the expression of pro-inflammatory genes, antimicrobial peptides, and regulates the activation, differentiation and survival of immunity cells [187]. However, Nf-κB signalling having an influential role in ageing and lifespan is less established. In Drosophila, overexpression of two immuno-stimulating pattern recognition receptors, the intracellular PGRP-LE and the transmembrane PGRP-LC, both upstream of Toll and IMD confer an inflammatory state with detrimental effects on lifespan [188, 189]. In line with this, genetic manipulation of immune-suppressive PGRP genes implies that activation of the immune system is unfavourable for longevity [190, 191]. As expected, the activity of IMD and Toll pathway is overall detrimental for lifespan, however, might also partially depend on tissue, sex, the gut microbiome, and type of genetic intervention [26, 192,193,194,195,196,197,198,199,200,201,202,203,204]. The effect on lifespan might be dependent on the expression of downstream AMPs, which can take both pro- and anti-longevity roles, with some study-specificity regarding their effect on lifespan [193, 196, 204]. As such, the increased immune system activity associated with ageing is perhaps harmful for lifespan because detrimental components outweigh beneficial factors.
One emerging mechanism explaining lifespan regulation by immune signalling pathways in Drosophila is through cross-talk with the IIS/TOR network. Innate immune signalling has been found to reduce insulin and TOR pathway activity, where this might be partially directed by cross-talk with the JNK pathway [148, 149, 194, 205,206,207,208]. In contrast to this, IMD can also activate insulin signalling in enteroendocrine cells, and plays a role in lipid metabolism and development, therefore highlighting tissue-specific effects and the difficulty to formulate general system-wide conclusions [209]. These findings pose a challenging molecular conundrum: although Nf-κB inhibits the IIS/TOR signalling network in certain conditions, which usually results in increased lifespan, it concomitantly has negative effects on lifespan.
Equivalent to findings in Drosophila, there is increasing evidence that in mammals, Nf-κB signalling stimulates pro-inflammatory responses at the cost of reduced lifespan [210, 211] and has been associated with several age-related pathologies [212, 213]. Indeed, long-lived dwarf mice have decreased levels of inflammatory markers in neurological and other tissues, possibly attributable to lowered JNK and Nf-κB activity, further confirming that these mice suffer less from chronic inflammation with age [156, 214]. Thus, while Nf-κB activity is required for pathogen resistance and inflammation, it is generally detrimental for lifespan in both fruit flies and mammals, representing a conserved trade-off between immunity and ageing.
Conclusions
In this review, we combined curation and analysis of orthologs between D. melanogaster, C. elegans, mice and humans to reveal that genes currently known to be pleiotropically involved in immunity, lifespan, and ageing reside in a few core pathways mediating the immuno-ageing interplay (Fig. 2, Table S1, Table S2). Remarkably, several of the most conserved immuno-ageing pathways we found are historically considered to function either in metabolism and longevity or in immunity. However, we highlight that these pathways not only cross-talk, but also clearly act pleiotropically to regulate pathogen resistance, lifespan, and ageing among many other physiological processes such as metabolism and stress resistance.
We also identified several cases where the effects of these pathways were not consistent, but varied between tissues, experimental conditions, sexes and pathogens. Classifications of genes into ‘pro’ and ‘anti’ longevity/immunity might therefore present a wrong or incomplete picture in many cases as context-dependency needs to be considered. Irrespective of that, we attempted to generalize the effect of each discussed pathway on lifespan and immunity (see Table 1) to discover trade-offs and functional conservation. Based on Table 1, we hypothesize that IIS/TOR, RNAi, and Nf-κB tend to be more functionally conserved than other pathways. Among these pathways, only the Nf-κB pathway clearly modulates a lifespan-immunity trade-off, as its activation facilitates pathogen defence but reduces lifespan. A good candidate mechanism explaining this is that triggering the Nf-κB cascades drives inflammatory and antimicrobial processes, which are detrimental for pathogens and the infected host at the same time. In contrast, activation of IIS/TOR mostly promotes growth in the presence of nutrients at the cost of both immunity and lifespan. The reallocation of resources between these phenotypes is mediated by Foxo transcription factors that transcribe longevity-conferring genes but concurrently upregulate immunity and stress-response genes. Finally, the RNAi pathway, which is also influenced by the IIS/TOR network, generally facilitates longevity and pathogen resistance, that could be resulting from regulation of gene expression and silencing of transposable elements in addition to its antiviral function.
Notably, most of the immuno-ageing pathways were identified through loss-of-function and gene knockdown assays, while insights from studies on population-wide genetic variation are limited. Most evidence for how alleles in these pathways affect ageing and immunity comes from Drosophila and humans. For instance, variants in Nf-κB related immunity genes were associated with longevity and improved immunosenescence in Drosophila [195]. In humans, FOXO3 and the cytokine interleukin 6 (IL6), which induces JAK/STAT, ERK-MAPK and PI3K signalling, are two prominent examples for which genetic variability has been linked with variation in lifespan and inflammation [98, 215,216,217]. This highlights that methods connecting genomic and phenotypic variation, such as genome-wide association studies, are powerful tools that will be further utilized to understand immuno-ageing genes and pathways.
Our review demonstrates that loss of immune homeostasis is a central determinant of ageing across diverse phyla. Yet, whether immunosenescence and the age-associated decline in other traits is a cause or result of ageing remains a fundamental problem difficult to resolve. Knowing the exact time and place of changes related to immunity and ageing would be a huge step in answering this question. Moreover, how environmental effects, including life-long pathogenic challenges, variation in the microbiome, or nutrition affect age-related changes in immunity is poorly understood. To date most studies are restricted in resolution, particularly in terms of analysed tissues, time points, phenotypes and experimental conditions. Cutting-edge technologies such as single-cell sequencing can be useful in that respect and could be utilized to characterize molecular changes during ageing and infection in specific cell types. In combination with genome-wide CRISPR knockout screens, new immuno-ageing genes can be discovered and the cross-talk between immunity and ageing further deciphered. Currently, the level of detail needed to solve the causality enigma of ageing is likely not achievable in humans but may be addressed in shorter lived model organisms that are easier to manipulate. Once we understood ageing at this unprecedented level, it will be possible to optimize lifestyle factors and emerging drug therapies treating senescence to facilitate healthy ageing and extend lifespan.
Abbreviations
- AMP:
-
Antimicrobial peptides
- EGFR:
-
Epidermal growth factor receptor
- ERK:
-
Extracellular-signal-regulated kinase
- GO:
-
Gene Ontology
- IFN:
-
Interferon
- IGF:
-
Insulin-like growth factor
- IIS:
-
Insulin-insulin-like growth factor signalling
- JAK/STAT:
-
Janus kinase / signal transducer and activator of transcription
- JNK:
-
c-Jun N-terminal kinase
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- MAPK:
-
Mitogen-activated protein kinase
- MHC:
-
Major histocompatibility complex
- mTORC1/2:
-
Mammalian target of rapamycin complex 1/2
- Nf-κB:
-
Nuclear factor kappa-light-chain-enhancer of activated B cells
- PI3K:
-
Phosphatidyl-inositol 3 kinase
- PGRP:
-
Peptidoglycan recognition proteins
- PRR:
-
Pattern recognition receptors
- RNAi:
-
RNA interference
- ROS:
-
Reactive oxygen species
- TGF-β:
-
Transforming growth factor beta
- TOR:
-
Target of rapamycin
References
Lozano-Durán R, Zipfel C. Trade-off between growth and immunity: role of brassinosteroids. Trends Plant Sci. 2015;20(1):12–9.
Schwenke RA, Lazzaro BP, Wolfner MF. Reproduction–Immunity Trade-Offs in Insects. Annu Rev Entomol. 2016;61:239–56.
Lochmiller RL, Deerenberg C. Trade-offs in evolutionary immunology: just what is the cost of immunity? Oikos. 2000;88(1):87–98.
Effros RB. Roy Walford and the immunologic theory of aging. Immun Ageing. 2005;2:7.
Sadighi Akha AA. Aging and the immune system: an overview. J Immunol Methods. 2018;463:21–6.
Ventura MT, Casciaro M, Gangemi S, Buquicchio R. Immunosenescence in aging: between immune cells depletion and cytokines up-regulation. Clin Mol Allergy. 2017;15:21.
Watad A, Bragazzi NL, Adawi M, Amital H, Toubi E, Porat BS, et al. Autoimmunity in the elderly: insights from basic science and clinics - a mini-review. Gerontology. 2017;63:515–23.
Laws TR, Harding SV, Smith MP, Atkins TP, Titball RW. Age influences resistance of Caenorhabditis elegans to killing by pathogenic bacteria. FEMS Microbiol Lett. 2004;234(2):281–7.
Youngman MJ, Rogers ZN, Kim DH. A decline in p38 MAPK signaling underlies immunosenescence in caenorhabditis elegans. PLoS Genet. 2011;7(5):e1002082.
Ramsden S, Cheung YY, Seroude L. Functional analysis of the Drosophila immune response during aging. Aging Cell. 2008;7(2):225–36. https://doi.org/10.1111/j.1474-9726.2008.00370.x.
Kubiak M, Tinsley MC. Sex-Specific Routes To Immune Senescence In Drosophila melanogaster. Sci Rep. 2017;7(1):10417.
Darby SC, Ewart DW, Giangrande PLF, Spooner RJD, Rizza CR. Importance of age at infection with HIV-1 for survival and development of AIDS in UK haemophilia population. Lancet. 1996;347(9015):1573–9.
Starr ME, Steele AM, Saito M, Hacker BJ, Evers BM, Saito H, et al. A new cecal slurry preparation protocol with improved long-term reproducibility for animal models of sepsis. PLoS One. 2014;9(12):e115705.
Xin L, Jiang TT, Kinder JM, Ertelt JM, Way SS. Infection susceptibility and immune senescence with advancing age replicated in accelerated aging Lmna(Dhe) mice. Aging Cell. 2015;14(6):1122–6.
Seroude L, Brummel T, Kapahi P, Benzer S. Spatio-temporal analysis of gene expression during aging in Drosophila melanogaster. Aging Cell. 2002;1(1):47–56.
Pletcher SD, Macdonald SJ, Marguerie R, Certa U, Stearns SC, Goldstein DB, et al. Genome-wide transcript profiles in aging and calorically restricted Drosophila melanogaster. Curr Biol. 2002;12(9):712–23.
Lee CK, Weindruch R, Prolla TA. Gene-expression profile of the ageing brain in mice. Nat Genet. 2000;25(3):294–7.
Park SK, Kim K, Page GP, Allison DB, Weindruch R, Prolla TA. Gene expression profiling of aging in multiple mouse strains: identification of aging biomarkers and impact of dietary antioxidants. Aging Cell. 2009;8(4):484–95. https://doi.org/10.1111/j.1474-9726.2009.00496.x.
Jia K, Cui C, Gao Y, Zhou Y, Cui Q. An analysis of aging-related genes derived from the genotype-tissue expression project (GTEx). Cell Death Discov. 2018;4:26.
Horn L, Leips J, Starz-Gaiano M. Phagocytic ability declines with age in adult Drosophila hemocytes. Aging Cell. 2014;13:719–28.
Kannarkat GT, Boss JM, Tansey MG. The role of innate and adaptive immunity in parkinson’s disease. J Parkinsons Dis. 2013;3(4):493–514.
Isobe K, Nishio N, Hasegawa T. Immunological aspects of age-related diseases. World J Biol Chem. 2017;8(2):129–37.
Heppner FL, Ransohoff RM, Becher B. Immune attack: the role of inflammation in Alzheimer disease. Nat Rev Neurosci. 2015;16(6):358–72.
Franceschi C, Garagnani P, Parini P, Giuliani C, Santoro A. Inflammaging: a new immune–metabolic viewpoint for age-related diseases. Nat Rev Endocrinol. 2018;14(10):576–90. https://doi.org/10.1038/s41574-018-0059-4.
Wu SC, Cao ZS, Chang KM, Juang JL. Intestinal microbial dysbiosis aggravates the progression of Alzheimer’s disease in Drosophila. Nat Commun. 2017;8(1):24.
Cao Y, Chtarbanova S, Petersen AJ, Ganetzky B. Dnr1 mutations cause neurodegeneration in Drosophila by activating the innate immune response in the brain. Proc Natl Acad Sci U S A. 2013;110(19):E1752–60.
Matarin M, Salih DA, Yasvoina M, Cummings DM, Guelfi S, Liu W, et al. A genome-wide gene-expression analysis and database in transgenic mice during development of amyloid or tau pathology. Cell Rep. 2015;10:633–45 Available from: https://doi.org/10.1016/j.celrep.2014.12.041.
De Magalhães JP, Toussaint O. GenAge: A genomic and proteomic network map of human ageing. FEBS Lett. 2004;571(1-3):243–7.
Hühne R, Thalheim T, Sühnel J. AgeFactDB - The JenAge Ageing Factor Database - Towards data integration in ageing research. Nucleic Acids Res. 2014;42(Database issue):D892–6.
Brucker RM, Funkhouser LJ, Setia S, Pauly R, Bordenstein SR. Insect Innate Immunity Database (IIID): An Annotation Tool for Identifying Immune Genes in Insect Genomes. PLoS One. 2012;7(9):e45125.
Breuer K, Foroushani AK, Laird MR, Chen C, Sribnaia A, Lo R, et al. InnateDB: Systems biology of innate immunity and beyond - Recent updates and continuing curation. Nucleic Acids Res. 2013;41(Database issue):D1228–33.
Ortutay C, Vihinen M. Immunome Knowledge Base (IKB): An integrated service for immunome research. BMC Immunol. 2009;10:3.
Fernandes M, Wan C, Tacutu R, Barardo D, Rajput A, Wang J, et al. Systematic analysis of the gerontome reveals links between aging and age-related diseases. Hum Mol Genet. 2016;25(21):4804–18. https://doi.org/10.1093/hmg/ddw307.
Hu Y, Flockhart I, Vinayagam A, Bergwitz C, Berger B, Perrimon N, et al. An integrative approach to ortholog prediction for disease-focused and other functional studies. BMC Bioinformatics. 2011;12:357.
Morrison D. MAP kinase pathways. Cold Spring Harb Perspect Biol. 2012;4:a011254.
Rawlings JS. The JAK/STAT signaling pathway. J Cell Sci. 2004;117(8):1281–3. https://doi.org/10.1242/jcs.00963.
Myllymäki H, Rämet M. JAK/STAT pathway in Drosophila immunity. Scand J Immunol. 2014;79:377–85.
Travis MA, Sheppard D. TGF-β Activation and Function in Immunity. Annu Rev Immunol. 2014;32:51–82.
Kubiczkova L, Sedlarikova L, Hajek R, Sevcikova S. TGF-β - an excellent servant but a bad master. J Transl Med. 2012;10:183.
Fontana L, Partridge L, Longo VD. Extending healthy life span-from yeast to humans. Science. 2010;328:321–6.
Piper MDW, Selman C, McElwee JJ, Partridge L. Separating cause from effect: how does insulin/IGF signalling control lifespan in worms, flies and mice? J Intern Med. 2008;263:179–91.
Pan H, Finkel T. Key proteins and pathways that regulate lifespan. J Biol Chem. 2017;292:6452–60.
Garsin DA, Villanueva JM, Begun J, Kim DH, Sifri CD, Calderwood SB, et al. Long-lived C.elegans daf-2 mutants are resistant to bacterial pathogens. Science. 2003;300:1921.
Evans EA, Kawli T, Tan MW. Pseudomonas aeruginosa suppresses host immunity by activating the DAF-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008;4(10):e1000175.
Anyanful A, Dolan-Livengood JM, Lewis T, Sheth S, MN DZ, Sherman MA, et al. Paralysis and killing of Caenorhabditis elegans by enteropathogenic Escherichia coli requires the bacterial tryptophanase gene. Mol Microbiol. 2005;57(4):988–1007.
Zhao Y, Gilliat AF, Ziehm M, Turmaine M, Wang H, Ezcurra M, et al. Two forms of death in ageing Caenorhabditis elegans. Nat Commun. 2017;8:15458.
Garigan D, Hsu AL, Fraser AG, Kamath RS, Abringet J, Kenyon C. Genetic analysis of tissue aging in Caenorhabditis elegans: a role for heat-shock factor and bacterial proliferation. Genetics. 2002;161(3):1101–12.
Finch CE, Ruvkun G. The Genetics of Aging. Annu Rev Genomics Hum Genet. 2001;2:435–62.
Zou CG, Tu Q, Niu J, Ji XL, Zhang KQ. The DAF-16/FOXO transcription factor functions as a regulator of epidermal innate immunity. PLoS Pathog. 2013;9(10):e1003660.
Evans EA, Chen WC, Tan MW. The DAF-2 insulin-like signaling pathway independently regulates aging and immunity in C. elegans. Aging Cell. 2008;7(6):879–93.
Clancy DJ, Gems D, Harshman LG, Oldham S, Stocker H, Hafen E, et al. Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein. Science. 2001;292(5514):104–6.
McCormack S, Yadav S, Shokal U, Kenney E, Cooper D, Eleftherianos I. The insulin receptor substrate Chico regulates antibacterial immune function in Drosophila. Immun Ageing. 2016;13:1–11.
Libert S, Chao Y, Zwiener J, Pletcher SD. Realized immune response is enhanced in long-lived puc and chico mutants but is unaffected by dietary restriction. Mol Immunol. 2008;45(3):810–7.
Becker T, Loch G, Beyer M, Zinke I, Aschenbrenner AC, Carrera P, et al. FOXO-dependent regulation of innate immune homeostasis. Nature. 2010;463:369–73.
Dionne MS, Pham LN, Shirasu-Hiza M, Schneider DS. Akt and foxo Dysregulation Contribute to Infection-Induced Wasting in Drosophila. Curr Biol. 2006;16(20):1977–85.
Fink C, Hoffmann J, Knop M, Li Y, Isermann K, Roeder T. Intestinal FoxO signaling is required to survive oral infection in Drosophila. Mucosal Immunol. 2016;9(4):927–36.
Hansen M, Taubert S, Crawford D, Libina N, Lee SJ, Kenyon C. Lifespan extension by conditions that inhibit translation in Caenorhabditis elegans. Aging Cell. 2007;6(1):95–110.
Bakowski MA, Desjardins CA, Smelkinson MG, Dunbar TA, Lopez-Moyado IF, Rifkin SA, et al. Ubiquitin-Mediated Response to Microsporidia and Virus Infection in C. elegans. PLoS Pathog. 2014;10(6):e1004200.
Seo K, Choi E, Lee D, Jeong DE, Jang SK, Lee SJ. Heat shock factor 1 mediates the longevity conferred by inhibition of TOR and insulin/IGF-1 signaling pathways in C. elegans. Aging Cell. 2013;12(6):1073–81.
Bernal A, Kimbrell DA. Drosophila Thor participates in host immune defense and connects a translational regulator with innate immunity. Proc Natl Acad Sci U S A. 2000;97(11):6019–24. https://doi.org/10.1073/pnas.100391597.
Levitin A, Marcil A, Tettweiler G, Laforest MJ, Oberholzer U, Alarco AM, et al. Drosophila melanogaster Thor and response to Candida albicans infection. Eukaryot Cell. 2007;6(4):658–63.
Varma D, Bülow MH, Pesch YY, Loch G, Hoch M. Forkhead, a new cross regulator of metabolism and innate immunity downstream of TOR in Drosophila. J Insect Physiol. 2014;69:80–8.
Cho I, Horn L, Felix TM, Foster L, Gregory G, Starz-gaiano M, et al. Age and diet specific effects of variation at S6 kinase on life history, metabolic, and immune response traits in Drosophila melanogaster. DNA Cell Biol. 2010;29(9):473–85.
Yuen GJ, Ausubel FM. Both live and dead Enterococci activate Caenorhabditis elegans host defense via immune and stress pathways. Virulence. 2018;9(1):683–99.
Iatsenko I, Sinha A, Rödelsperger C, Sommer RJ. New role for DCR-1/dicer in caenorhabditis elegans innate immunity against the highly virulent bacterium bacillus thuringiensis DB27. Infect Immun. 2013;81(10):3942–57. https://doi.org/10.1128/IAI.00700-13.
Lu R, Maduro M, Li F, Li HW, Broitman-Maduro G, Li WX, et al. Animal virus replication and RNAi-mediated antiviral silencing in Caenorhabditis elegans. Nature. 2005;436(7053):1040–3.
Gammon DB, Ishidate T, Li L, Gu W, Silverman N, Mello CC. The Antiviral RNA Interference Response Provides Resistance to Lethal Arbovirus Infection and Vertical Transmission in Caenorhabditis elegans. Curr Biol. 2017;27(6):795–806.
Karlikow M, Goic B, Saleh MC. RNAi and antiviral defense in Drosophila: setting up a systemic immune response. Dev comp Immunol. 2014;42:85–92.
Wang D, Ruvkun G. Regulation of Caenorhabditis elegans RNA interference by the daf-2 insulin stress and longevity signaling pathway. Cold Spring Harb Symp Quant Biol. 2004;69:429–31.
Simon M, Sarkies P, Ikegami K, Doebley AL, Goldstein LD, Mitchell J, et al. Reduced insulin/IGF-1 signaling restores germ cell immortality to Caenorhabditis elegans Piwi mutants. Cell Rep. 2014;7(3):762–73.
Simon M, Spichal M, Heestand B, Frenk S, Hedges A, Godwin M, et al. DAF-16/Foxo suppresses the transgenerational sterility of prg-1 piRNA mutants via a systemic small RNA pathway. unpublished data. 2018.
Krug L, Chatterjee N, Borges-Monroy R, Hearn S, Liao WW, Morrill K, et al. Retrotransposon activation contributes to neurodegeneration in a Drosophila TDP-43 model of ALS. PLoS Genet. 2017;13:e1006635.
De Cecco M, Criscione SW, Peterson AL, Neretti N, Sedivy JM, Kreiling JA. Transposable elements become active and mobile in the genomes of aging mammalian somatic tissues. Aging (Albany NY). 2013;5(12):867–83.
De Cecco M, Ito T, Petrashen AP, Elias AE, Skvir NJ, Criscione SW, et al. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature. 2019;566(7742):73–8. https://doi.org/10.1038/s41586-018-0784-9.
Li W, Prazak L, Chatterjee N, Grüninger S, Krug L, Theodorou D, et al. Activation of transposable elements during aging and neuronal decline in Drosophila. Nat Neurosci. 2013;16:529–31.
Fabian DK, Donertas H, Fuentealba M, Partridge L, Thornton J. Transposable element landscape in Drosophila populations selected for longevity. Genome Biol Evol. 2021;13(4):evab031.
Patel RK, Hardy RW. Role for the phosphatidylinositol 3-kinase-Akt-TOR pathway during Sindbis virus replication in arthropods. J Virol. 2012;86(7):3595–604.
Spellberg MJ, Marr MT. FOXO regulates RNA interference in Drosophila and protects from RNA virus infection. Proc Natl Acad Sci U S A. 2015;112:14587–92 Available from: https://www.pnas.org/lookup/doi/10.1073/pnas.1517124112.
Saeed MF, Kolokoltsov AA, Freiberg AN, Holbrook MR, Davey RA. Phosphoinositide-3 kinase-akt pathway controls cellular entry of ebola virus. PLoS Pathog. 2008;4(8):e1000141. https://doi.org/10.1371/journal.ppat.1000141.
Mannova P, Beretta L. Activation of the N-Ras-PI3K-Akt-mTOR pathway by hepatitis C virus: control of cell survival and viral replication. J Virol. 2005;79(14):8742–9.
Nehdi A, Sean P, Linares I, Colina R, Jaramillo M, Alain T, et al. Deficiency in either 4E-BP1 or 4E-BP2 augments innate antiviral immune responses. PLoS One. 2014;9(12):e114854.
Ding SW, Han Q, Wang J, Li WX. Antiviral RNA interference in mammals. Curr Opin Immunol. 2018;54:109–14. https://doi.org/10.1016/j.coi.2018.06.010.
Jeang KT. RNAi in the regulation of mammalian viral infections. BMC Biol. 2012;10:58.
Mori MA, Raghavan P, Thomou T, Boucher J, Robida-Stubbs S, MacOtela Y, et al. Role of microRNA processing in adipose tissue in stress defense and longevity. Cell Metab. 2012;16:336–3347.
Frezzetti D, Reale C, Calì G, Nitsch L, Fagman H, Nilsson O, et al. The microRNA-processing enzyme dicer is essential for thyroid function. PLoS One. 2011;6(11):e27648.
Lim DH, Oh CT, Lee L, Hong JS, Noh SH, Hwang S. et al, The endogenous siRNA pathway in Drosophila impacts stress resistance and lifespan by regulating metabolic homeostasis. FEBS Lett. 2011;585(19):3079–85.
Zhou R, Rana TM. RNA-based mechanisms regulating host-virus interactions. Immunol Rev. 2013;253(1):97–111.
Alic N, Hoddinott MP, Foley A, Slack C, Piper MDW, Partridge L. Detrimental effects of RNAi: a cautionary note on its use in Drosophila ageing studies. PLoS One. 2012;7:1–5.
Zhang J, Yang W, Xu J, Yang W, Li Q, Zhong Y. Regulation of antimicrobial peptide genes via insulin-like signaling pathway in the silkworm Bombyx mori. Insect Biochem Mol Biol. 2018;103:12–21.
Zhong X, Chowdhury M, Li CF, Yu XQ. Transcription factor forkhead regulates expression of antimicrobial peptides in the tobacco hornworm, manduca sexta. Sci Rep. 2017;7(1):2688.
Pakpour N, Corby-Harris V, Green GP, Smithers HM, Cheung KW, Riehle MA, et al. Ingested human insulin inhibits the mosquito NF-κB-dependent immune response to Plasmodium falciparum. Infect Immun. 2012;80(6):2141–9.
Flurkey K, Papaconstantinou J, Miller RA, Harrison DE. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proc Natl Acad Sci U S A. 2001;98(12):6736–41.
Brown-Borg HM, Borg KE, Meliska CJ, Bartke A. Dwarf mice and the ageing process. Nature. 1996;384(6604):33.
Selman C, Lingard S, Choudhury AI, Batterham RL, Claret M, Clements M, et al. Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice. FASEB J. 2007;22(3):807–18.
Selman C, Partridge L, Withers DJ. Replication of extended lifespan phenotype in mice with deletion of insulin receptor substrate 1. PLoS One. 2011;6(1):e16144.
Shimokawa I, Komatsu T, Hayashi N, Kim SE, Kawata T, Park S, et al. The life-extending effect of dietary restriction requires Foxo3 in mice. Aging Cell. 2015;14(4):707–9.
Joseph J, Ametepe ES, Haribabu N, Agbayani G, Krishnan L, Blais A, et al. Inhibition of ROS and upregulation of inflammatory cytokines by FoxO3a promotes survival against Salmonella typhimurium. Nat Commun. 2016;7:12748.
Morris BJ, Willcox DC, Donlon TA, Willcox BJ. FOXO3: a major gene for human longevity - a mini-review. Gerontology. 2015;61:515–25.
Cabrera-Ortega AA, Feinberg D, Liang Y, Rossa C, Graves DT. The role of Forkhead box 1 (FOXO1) in the immune system: dendritic cells, T cells, B cells, and hematopoietic stem cells. Crit Rev Immunol. 2017;37(1):1–13.
Hedrick SM, Michelini RH, Doedens AL, Goldrath AW, Stone EL. FOXO transcription factors throughout T cell biology. Nat Rev Immunol. 2012;12(9):649–61.
Martins R, Lithgow GJ, Link W. Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Aging Cell. 2016;15(2):196–207.
Weichhart T, Costantino G, Poglitsch M, Rosner M, Zeyda M, Stuhlmeier KM, et al. The TSC-mTOR Signaling Pathway Regulates the Innate Inflammatory Response. Immunity. 2008;29(4):565–77.
Brown J, Wang H, Suttles J, Graves DT, Martin M. Mammalian target of rapamycin complex 2 (mTORC2) negatively regulates toll-like receptor 4-mediated inflammatory response via FoxO1. J Biol Chem. 2011;286(52):44295–305.
Schmitz F, Heit A, Dreher S, Eisenächer K, Mages J, Haas T, et al. Mammalian target of rapamycin (mTOR) orchestrates the defense program of innate immune cells. Eur J Immunol. 2008;38(11):2981–92.
Kusaba H, Ghosh P, Derin R, Buchholz M, Sasaki C, Madara K, et al. Interleukin-12-induced interferon-gamma production by human peripheral blood T cells is regulated by mammalian target of rapamycin (mTOR). J Biol Chem. 2005;280(2):1037–43.
Powell JD, Pollizzi KN, Heikamp EB, Horton MR. Regulation of Immune Responses by mTOR. Annu Rev Immunol. 2012;30:39–68.
Gaubitz C, Prouteau M, Kusmider B, Loewith R. TORC2 structure and function. Trends Biochem Sci. 2016;41(6):532–45.
Chen C, Liu Y, Liu Y, Zheng P. MTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci Signal. 2009;2(98):ra75.
Mannick JB, Del Giudice G, Lattanzi M, Valiante NM, Praestgaard J, Huang B, et al. mTOR inhibition improves immune function in the elderly. Sci Transl Med. 2014;6(268):268ra179.
Fruman DA, Chiu H, Hopkins BD, Bagrodia S, Cantley LC, Abraham RT. The PI3K pathway in human disease. Cell. 2017;170(4):605–35.
Smith TJ. Insulin-like growth factor-I regulation of immune function: a potential therapeutic target in autoimmune diseases? Pharmacol Rev. 2010;62(2):199–236.
Koyasu S. The role of P13K in immune cells. Nat Immunol. 2003;4(4):313–9.
Fischer HJ, Sie C, Schumann E, Witte A-K, Dressel R, van den Brandt J, et al. The insulin receptor plays a critical role in T cell function and adaptive immunity. J Immunol. 2017;198(5):1910–20.
Tsai S, Clemente-Casares X, Zhou AC, Lei H, Ahn JJ, Chan YT, et al. Insulin Receptor-Mediated Stimulation Boosts T Cell Immunity during Inflammation and Infection. Cell Metab. 2018;28(6):922–934.e4.
McGaugh SE, Bronikowski AM, Kuo C-H, Reding DM, Addis EA, Flagel LE, et al. Rapid molecular evolution across amniotes of the IIS/TOR network. Proc Natl Acad Sci U S A. 2015;112(22):7055–60.
Wang M, Zhang X, Zhao H, Wang Q, Pan Y. FoxO gene family evolution in vertebrates. BMC Evol Biol. 2009;9:222.
Philippon H, Brochier-Armanet C, Perrière G. Evolutionary history of phosphatidylinositol- 3-kinases: ancestral origin in eukaryotes and complex duplication patterns. BMC Evol Biol. 2015;15(1):226. https://doi.org/10.1186/s12862-015-0498-7.
Yada T. Growth hormone and fish immune system. Gen Comp Endocrinol. 2007;152(2-3):353–8.
Alzaid A, Castro R, Wang T, Secombes CJ, Boudinot P, Macqueen DJ, et al. Cross talk between growth and immunity: coupling of the igf axis to conserved cytokine pathways in rainbow trout. Endocrinology. 2016;157(5):1942–55.
Okuyama T, Inoue H, Ookuma S, Satoh T, Kano K, Honjoh S, et al. The ERK-MAPK pathway regulates longevity through SKN-1 and insulin-like signaling in Caenorhabditis elegans. J Biol Chem. 2010;285(39):30274–81. https://doi.org/10.1074/jbc.M110.146274.
Nicholas HR, Hodgkin J. The ERK MAP kinase cascade mediates tail swelling and a protective response to rectal infection in C. elegans. Curr Biol. 2004;14(14):1256–61.
Papp D, Csermely P, Soti C. A role for SKN-1/Nrf in pathogen resistance and immunosenescence in caenorhabditis elegans. PLoS Pathog. 2012;8(4):e1002673. https://doi.org/10.1371/journal.ppat.1002673.
Inoue H, Hisamoto N, Jae HA, Oliveira RP, Nishida E, Blackwell TK, et al. The C. elegans p38 MAPK pathway regulates nuclear localization of the transcription factor SKN-1 in oxidative stress response. Genes Dev. 2005;19(19):2278–83.
Zhi L, Yu Y, Li X, Wang D, Wang D. Molecular control of innate immune response to Pseudomonas aeruginosa infection by intestinal let-7 in Caenorhabditis elegans. PLoS Pathog. 2017;13(1):e1006152.
Slack C, Alic N, Foley A, Cabecinha M, Hoddinott MP, Partridge L. The Ras-Erk-ETS-Signaling Pathway Is a Drug Target for Longevity. Cell. 2015;162(1):72–83.
Ragab A, Buechling T, Gesellchen V, Spirohn K, Boettcher AL, Boutros M. Drosophila Ras/MAPK signalling regulates innate immune responses in immune and intestinal stem cells. EMBO J. 2011;30(6):1123–36.
Xu J, Hopkins K, Sabin L, Yasunaga A, Subramanian H, Lamborn I, et al. ERK signaling couples nutrient status to antiviral defense in the insect gut. Proc Natl Acad Sci U S A. 2013;110(37):15025–30.
Liu W-Q, Chen S-Q, Bai H-Q, Wei Q-M, Zhang S-N, Chen C, et al. The Ras/ERK signaling pathway couples antimicrobial peptides to mediate resistance to dengue virus in Aedes mosquitoes. PLoS Negl Trop Dis. 2020;14(8):e0008660.
Irazoqui JE, Urbach JM, Ausubel FM. Evolution of host innate defence: insights from Caenorhabditis elegans and primitive invertebrates. Nat Rev Immunol. 2010;10(1):47–58. https://doi.org/10.1038/nri2689.
Liberati NT, Fitzgerald KA, Kim DH, Feinbaum R, Golenbock DT, Ausubel FM. Requirement for a conserved toll/interleukin-1 resistance domain protein in the Caenorhabditis elegans immune response. Proc Natl Acad Sci U S A. 2004;101(17):6593–8.
McEwan DL, Feinbaum RL, Stroustrup N, Haas W, Conery AL, Anselmo A, et al. Tribbles ortholog NIPI-3 and bZIP transcription factor CEBP-1 regulate a Caenorhabditis elegans intestinal immune surveillance pathway. BMC Biol. 2016;14(1):105. https://doi.org/10.1186/s12915-016-0334-6.
Pujol N, Cypowyj S, Ziegler K, Millet A, Astrain A, Goncharov A, et al. Distinct Innate Immune Responses to Infection and Wounding in the C. elegans Epidermis. Curr Biol. 2008;18(7):481–9.
Kim KW, Thakur N, Piggott CA, Omi S, Polanowska J, Jin Y, et al. Coordinated inhibition of C/EBP by Tribbles in multiple tissues is essential for Caenorhabditis elegans development. BMC Biol. 2016;14(1):104.
Shivers RP, Pagano DJ, Kooistra T, Richardson CE, Reddy KC, Whitney JK, et al. Phosphorylation of the conserved transcription factor ATF-7 by PMK-1 p38 MAPK regulates innate immunity in Caenorhabditis elegans. PLoS Genet. 2010;6(4):e1000892.
Yunger E, Safra M, Levi-Ferber M, Haviv-Chesner A, Henis-Korenblit S. Innate immunity mediated longevity and longevity induced by germ cell removal converge on the C-type lectin domain protein IRG-7. PLoS Genet. 2017;13(2):e1006577.
Troemel ER, Chu SW, Reinke V, Lee SS, Ausubel FM, Kim DH. p38 MAPK regulates expression of immune response genes and contributes to longevity in C. elegans. PLoS Genet. 2006;2(11):e183.
Wu Z, Isik M, Moroz N, Steinbaugh MJ, Zhang P, Blackwell TK. Dietary Restriction Extends Lifespan through Metabolic Regulation of Innate Immunity. Cell Metab. 2019; 29: 1192–1205.e8.
Han Z, Enslen H, Hu X, Meng X, Wu I, Barrett T, et al. A conserved p38 mitogen-activated protein kinase pathway regulates Drosophila immunity gene expression. Mol Cell Biol. 1998;18(6):3527–39.
Davis MM, Primrose DA, Hodgetts RB. A Member of the p38 Mitogen-Activated Protein Kinase Family Is Responsible for Transcriptional Induction of Dopa decarboxylase in the Epidermis of Drosophila melanogaster during the Innate Immune Response. Mol Cell Biol. 2008;28:4883–95.
Chen J, Xie C, Tian L, Hong L, Wu X, Han J. Participation of the p38 pathway in Drosophila host defense against pathogenic bacteria and fungi. Proc Natl Acad Sci U S A. 2010;107(48):20774–9. https://doi.org/10.1073/pnas.1009223107.
West C, Silverman N. p38b and JAK-STAT signaling protect against invertebrate iridescent virus 6 infection in Drosophila. PLoS Pathog. 2018;14(5):e1007020. https://doi.org/10.1371/journal.ppat.1007020.
Clark RI, Tan SWS, Pean CB, Roostalu U, Vivancos V, Bronda K, et al. MEF2 is an in vivo immune-metabolic switch. Cell. 2013;155(2):435–47.
Vrailas-Mortimer A, del Rivero T, Mukherjee S, Nag S, Gaitanidis A, Kadas D, et al. A muscle-specific p38 MAPK/Mef2/MnSOD pathway regulates stress, motor function, and life span in Drosophila. Dev Cell. 2011;21(4):783–95.
Ryan SM, Almassey M, Burch AM, Ngo G, Barbee S, Mortimer NT, et al. Drosophila p38 MAPK Interacts with BAG-3/starvin to Regulate Age-dependent Protein Homeostasis. unpublished data. 2019.
Oh SW, Mukhopadhyay A, Svrzikapa N, Jiang F, Davis RJ, Tissenbaum HA. JNK regulates lifespan in Caenorhabditis elegans by modulating nuclear translocation of forkhead transcription factor/DAF-16. Proc Natl Acad Sci U S A. 2005;102(12):4494–9.
Marudhupandiyan S, Balamurugan K. Intrinsic JNK-MAPK pathway involvement requires daf-16-mediated immune response during Shigella flexneri infection in C. elegans. Immunol Res. 2017;65(3):609–21.
Neumann-Haefelin E, Qi W, Finkbeiner E, Walz G, Baumeister R, Hertweck M. SHC-1/p52Shc targets the insulin/IGF-1 and JNK signaling pathways to modulate life span and stress response in C. elegans. Genes Dev. 2008;22(19):2721–35.
Biteau B, Karpac J, Supoyo S, DeGennaro M, Lehmann R, Jasper H. Lifespan extension by preserving proliferative homeostasis in Drosophila. PLoS Genet. 2010;6(10):e1001159.
Wang MC, Bohmann D, Jasper H. JNK extends life span and limits growth by antagonizing cellular and organism-wide responses to insulin signaling. Cell. 2005;121(1):115–25.
Biteau B, Karpac J, Hwangbo DS, Jasper H. Regulation of Drosophila lifespan by JNK signaling. Exp Gerontol. 2011;46:349–54.
Delaney JR, Stöven S, Uvell H, Anderson KV, Engström Y, Mlodzik M. Cooperative control of Drosophila immune responses by the JNK and NF-κB signaling pathways. EMBO J. 2006;25(13):3068–77. https://doi.org/10.1038/sj.emboj.7601182.
Zhou J, Edgar BA, Boutros M. ATF3 acts as a rheostat to control JNK signalling during intestinal regeneration. Nat Commun. 2017;8:14289.
Souvannaseng L, Hun LV, Baker H, Klyver JM, Wang B, Pakpour N, et al. Inhibition of JNK signaling in the Asian malaria vector Anopheles stephensi extends mosquito longevity and improves resistance to Plasmodium falciparum infection. PLoS Pathog. 2018;14(11):e1007418.
Borrás C, Monleón D, López-Grueso R, Gambini J, Orlando L, Pallardó F V., et al. RasGrf1 deficiency delays dging in mice. Aging (Albany NY). 2011;3(3):262–76.
Hsieh CC, Papaconstantinou J. Akt/PKB and p38 MAPK signaling, translational initiation and longevity in Snell dwarf mouse livers. Mech Ageing Dev. 2004;125(10-11):785–98.
Sun LY, Fang Y, Patki A, Koopman JJ, Allison DB, Hill CM, et al. Longevity is impacted by growth hormone action during early postnatal period. Elife. 2017;6:e24059.
He D, Wu H, Xiang J, Ruan X, Peng P, Ruan Y, et al. Gut stem cell aging is driven by mTORC1 via a p38 MAPK-p53 pathway. Nat Commun. 2020;11(1):37. https://doi.org/10.1038/s41467-019-13911-x.
Nishida K, Yamaguchi O, Hirotani S, Hikoso S, Higuchi Y, Watanabe T, et al. p38alpha mitogen-activated protein kinase plays a critical role in Cardiomyocyte survival but not in cardiac hypertrophic growth in response to pressure overload. Mol Cell Biol. 2004;24(24):10611–20.
Bisson N, Tremblay M, Robinson F, Kaplan DR, Trusko SP, Moss T. Mice lacking both mixed-lineage kinase genes Mlk1 and Mlk2 retain a wild type phenotype. Cell Cycle. 2008;7(7):909–16. https://doi.org/10.4161/cc.7.7.5610.
Huang G, Shi LZ, Chi H. Regulation of JNK and p38 MAPK in the immune system: signal integration, propagation and termination. Cytokine. 2009;48(3):161–9.
Cuenda A, Rousseau S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta - Mol. Cell Res. 2007;1773(8):1358–75.
Lee JC, Laydon JT, McDonnell PC, Gallagher TF, Kumar S, Green D, et al. A protein kinase involved in the regulation of inflammatory cytokine biosynthesis. Nature. 1994;372(6508):739–46.
Zhao X, Guo Y, Jiang C, Chang Q, Zhang S, Luo T, et al. JNK1 negatively controls antifungal innate immunity by suppressing CD23 expression. Nat Med. 2017;23(3):337–46.
Börgeling Y, Schmolke M, Viemann D, Nordhoff C, Roth J, Ludwig S. Inhibition of p38 mitogen-activated protein kinase impairs influenza virus-induced primary and secondary host gene responses and protects mice from lethal H5N1 infection. J Biol Chem. 2014;289(1):13–27.
Vukmanovic-Stejic M, Chambers ES, Suárez-Fariñas M, Sandhu D, Fuentes-Duculan J, Patel N, et al. Enhancement of cutaneous immunity during aging by blocking p38 mitogen-activated protein (MAP) kinase–induced inflammation. J Allergy Clin Immunol. 2018;142:844–56.
De Maeyer RPH, van de Merwe RC, Louie R, Bracken OV, Devine OP, Goldstein DR, et al. Blocking elevated p38 MAPK restores efferocytosis and inflammatory resolution in the elderly. Nat Immunol. 2020;21:615–25.
Arthur JSC, Ley SC. Mitogen-activated protein kinases in innate immunity. Nat Rev Immunol. 2013;13(9):679–92.
Dierking K, Polanowska J, Omi S, Engelmann I, Gut M, Lembo F, et al. Unusual regulation of a STAT protein by an SLC6 family transporter in C. elegans epidermal innate immunity. Cell Host Microbe. 2011;9(5):425–35.
Tanguy M, Véron L, Stempor P, Ahringer J, Sarkies P, Miska EA. An alternative STAT signaling pathway acts in viral immunity in Caenorhabditis elegans. MBio. 2017;8(5):e00924–17.
Li H, Qi Y, Jasper H. Preventing Age-Related Decline of Gut Compartmentalization Limits Microbiota Dysbiosis and Extends Lifespan. Cell Host Microbe. 2016;19(2):240–53.
Woodcock KJ, Kierdorf K, Pouchelon CA, Vivancos V, Dionne MS, Geissmann F. Macrophage-derived upd3 cytokine causes impaired glucose homeostasis and reduced lifespan in Drosophila fed a lipid-rich diet. Immunity. 2015;42(1):133–44. https://doi.org/10.1016/j.immuni.2014.12.023.
Larson K, Yan SJ, Tsurumi A, Liu J, Zhou J, Gaur K, et al. Heterochromatin formation promotes longevity and represses ribosomal RNA synthesis. PLoS Genet. 2012;8(1):e1002473.
Igaz P, Tóth S, Falus A. Biological and clinical significance of the JAK-STAT pathway; lessons from knockout mice. Inflamm Res. 2001;50(9):435–41. https://doi.org/10.1007/PL00000267.
Xu M, Tchkonia T, Ding H, Ogrodnik M, Lubbers ER, Pirtskhalava T, et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proc Natl Acad Sci U S A. 2015;112(46):E6301–10.
Casellas J, Medrano JF. Lack of Socs2 expression reduces lifespan in high-growth mice. Age (Omaha). 2008;30(4):245–9.
Jacoby JJ, Kalinowski A, Liu M-G, Zhang SS-M, Gao Q, Chai G-X, et al. Cardiomyocyte-restricted knockout of STAT3 results in higher sensitivity to inflammation, cardiac fibrosis, and heart failure with advanced age. Proc Natl Acad Sci U S A. 2003;100(22):12929–34.
Morikawa M, Derynck R, Miyazono K. TGF- β and the TGF-β family: context-dependent roles in cell and tissue physiology. Cold Spring Harb Perspect Biol. 2016;8:a021873.
Mallo GV, Kurz CL, Couillault C, Pujol N, Granjeaud S, Kohara Y, et al. Inducible antibacterial defense system in C. elegans. Curr Biol. 2002;12(14):1209–14.
Clark RI, Woodcock KJ, Geissmann F, Trouillet C, Dionne MS. Multiple TGF-β superfamily signals modulate the adult drosophila immune response. Curr Biol. 2011;21(19):1672–7.
Eleftherianos I, Castillo JC, Patrnogic J. TGF-β signaling regulates resistance to parasitic nematode infection in Drosophila melanogaster. Immunobiology. 2016;221(12):1362–8.
Patrnogic J, Heryanto C, Eleftherianos I. Transcriptional up-regulation of the TGF-β intracellular signaling transducer Mad of Drosophila larvae in response to parasitic nematode infection. Innate Immun. 2018;24(6):349–56.
Bai H, Kang P, Hernandez AM, Tatar M. Activin signaling targeted by insulin/dFOXO regulates aging and muscle Proteostasis in Drosophila. PLoS Genet. 2013;9(11):e1003941.
Langerak S, Kim M-J, Lamberg H, Godinez M, Main M, Winslow L, et al. The Drosophila TGF-beta/Activin-like ligands dawdle and Myoglianin appear to modulate adult lifespan through regulation of 26S proteasome function in adult muscle. Biol Open. 2018;7(4):bio029454.
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–217. https://doi.org/10.1016/j.cell.2013.05.039.
Kobayashi S, Yoshida K, Ward JM, Letterio JJ, Longenecker G, Yaswen L, et al. Beta 2-microglobulin-deficient background ameliorates lethal phenotype of the TGF-beta 1 null mouse. J Immunol. 1999;163(7):4013–9.
Nandula SR, Amarnath S, Molinolo A, Bandyopadhyay BC, Hall B, Goldsmith CM, et al. Female mice are more susceptible to developing inflammatory disorders due to impaired transforming growth factor β signaling in salivary glands. Arthritis Rheum. 2007;56(6):1798–805.
Liu T, Zhang L, Joo D, Sun SC. NF-κB signaling in inflammation. Signal Transduct Target Ther. 2017;2(1):17023. https://doi.org/10.1038/sigtrans.2017.23.
DeVeale B, Brummel T, Seroude L. Immunity and aging: the enemy within? Aging Cell. 2004;3:195–208.
Libert S, Chao Y, Chu X, Pletcher SD. Trade-offs between longevity and pathogen resistance in Drosophila melanogaster are mediated by NFκB signaling. Aging Cell. 2006;5:533–43.
Paredes JC, Welchman DP, Poidevin M, Lemaitre B. Negative regulation by Amidase PGRPs shapes the drosophila antibacterial response and protects the Fly from innocuous infection. Immunity. 2011;35:770–9.
Paik D, Jang YG, Lee YE, Lee YN, Yamamoto R, Gee HY, et al. Misexpression screen delineates novel genes controlling Drosophila lifespan. Mech Ageing Dev. 2012;133(5):234–45.
Ryu JH, Kim SH, Lee HY, Jin YB, Do NY, Bae JW, et al. Innate immune homeostasis by the homeobox gene Caudal and commensal-gut mutualism in Drosophila. Science. 2008;319(5864):777–82.
Lin YR, Parikh H, Park Y. Stress resistance and lifespan enhanced by downregulation of antimicrobial peptide genes in the Imd pathway. Aging (Albany NY). 2018;10:622–31.
Davoodi S, Galenza A, Panteluk A, Deshpande R, Ferguson M, Grewal S, et al. The immune deficiency pathway regulates metabolic homeostasis in Drosophila. J Immunol. 2019;202(9):2747–59.
Fabian DK, Garschall K, Klepsatel P, Santos-Matos G, Sucena É, Kapun M, et al. Evolution of longevity improves immunity in Drosophila. Evol Lett. 2018;2(6):567–79. https://doi.org/10.1002/evl3.89.
Loch G, Zinke I, Mori T, Carrera P, Schroer J, Takeyama H, et al. Antimicrobial peptides extend lifespan in Drosophila. PLoS One. 2017;12(5):e0176689.
Fernando MDA, Kounatidis I, Ligoxygakis P. Loss of Trabid, a new negative regulator of the Drosophila immune-deficiency pathway at the level of TAK1, Reduces Life Span. PLoS Genet. 2014;10(2):e1004117.
Kounatidis I, Chtarbanova S, Cao Y, Hayne M, Jayanth D, Ganetzky B, et al. NF-κB Immunity in the Brain Determines Fly Lifespan in Healthy Aging and Age-Related Neurodegeneration. Cell Rep. 2017;19(4):836–48.
Morris O, Liu X, Domingues C, Runchel C, Chai A, Basith S, et al. Signal Integration by the IκB Protein Pickle Shapes Drosophila Innate Host Defense. Cell Host Microbe. 2016;20(3):283–95.
Petersen AJ, Rimkus SA, Wassarman DA. ATM kinase inhibition in glial cells activates the innate immune response and causes neurodegeneration in Drosophila. Proc Natl Acad Sci U S A. 2012;109(11):E656–64. https://doi.org/10.1073/pnas.1110470109.
Moskalev A, Shaposhnikov M. Pharmacological inhibition of NF-kB prolongs lifespan of Drosophila melanogaster. Aging (Albany NY). 2011;3:391–4.
Le Bourg É, Malod K, Massou I. The NF-κB-like factor DIF could explain some positive effects of a mild stress on longevity, behavioral aging, and resistance to strong stresses in Drosophila melanogaster. Biogerontology. 2012;13:445–55.
Petersen AJ, Katzenberger RJ, Wassarman DA. The innate immune response transcription factor relish is necessary for neurodegeneration in a Drosophila model of ataxia-telangiectasia. Genetics. 2013;194(1):133–42.
Badinloo M, Nguyen E, Suh W, Alzahrani F, Castellanos J, Klichko VI, et al. Overexpression of antimicrobial peptides contributes to aging through cytotoxic effects in Drosophila tissues. Arch Insect Biochem Physiol. 2018;98(4):e21464.
DiAngelo JR, Bland ML, Bambina S, Cherry S, Birnbaum MJ. The immune response attenuates growth and nutrient storage in Drosophila by reducing insulin signaling. Proc Natl Acad Sci U S A. 2009;106(49):20853–8.
Roth SW, Bitterman MD, Birnbaum MJ, Bland ML. Innate immune signaling in Drosophila blocks insulin signaling by uncoupling PI(3,4,5)P3 production and Akt activation. Cell Rep. 2018;22(10):2550–6.
Musselman LP, Fink JL, Grant AR, Tuthill BF, Baranski TJ. A Complex Relationship between Immunity and Metabolism in Drosophila Diet-Induced Insulin Resistance. Mol Cell Biol. 2018;38:e00259–17.
Atilano M, Glittenberg M, Bahuguna S, Wang L, Ligoxygakis P. Drosophila Toll links systemic immunity to long-term intestinal epithelial integrity. unpublished data. 2018.
Kamareddine L, Robins WP, Berkey CD, Mekalanos JJ, Watnick PI. The Drosophila Immune Deficiency Pathway Modulates Enteroendocrine Function and Host Metabolism. Cell Metab. 2018;28(3):449–62.e5.
Zhang G, Li J, Purkayastha S, Tang Y, Zhang H, Yin Y, et al. Hypothalamic programming of systemic ageing involving IKK-β, NF-κB and GnRH. Nature. 2013;497(7448):211–6.
Jurk D, Wilson C, Passos JF, Oakley F, Correia-Melo C, Greaves L, et al. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice. Nat Commun. 2014;2:4172.
Tilstra JS, Clauson CL, Niedernhofer LJ, Robbins PD. NF-κB in Aging and Disease. Aging Dis. 2011;2(6):449–65.
Tilstra JS, Robinson AR, Wang J, Gregg SQ, Clauson CL, Reay DP, et al. NF-κB inhibition delays DNA damage - Induced senescence and aging in mice. J Clin Invest. 2012;122(7):2601–12.
Sadagurski M, Landeryou T, Cady G, Kopchick JJ, List EO, Berryman DE, et al. Growth hormone modulates hypothalamic inflammation in long-lived pituitary dwarf mice. Aging Cell. 2015;14(6):1045–54.
Scheller J, Chalaris A, Schmidt-Arras D, Rose-John S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta - Mol. Cell Res. 2011;1813:878–88.
Akbaraly TN, Hamer M, Ferrie JE, Lowe G, Batty GD, Hagger-Johnson G, et al. Chronic inflammation as a determinant of future aging phenotypes. Cmaj. 2013;185(16):E763–70. https://doi.org/10.1503/cmaj.122072.
Bonafe M, Olivieri F, Cavallone L, Giovagnetti S, Marchegiani F, Cardelli M, et al. A gender – dependent genetic predisposition to produce high levels of IL-6 is detrimental for longevity. Eur J Immunol. 2001;31(8):2357–61.
Acknowledgements
We are grateful to Joao Pedro de Magalhaes and one anonymous reviewer for thoughtful comments on our manuscript. Figures were created with BioRender.com.
Funding
This research was funded in whole, or in part, by the Wellcome Trust [WT098565/Z/12/Z]. For the purpose of Open Access, the author has applied a CC BY public copyright licence to any Author Accepted Manuscript version arising from this submission. The grant holders are JMT and LP. The work was further supported by EMBL (HMD and JMT), and Comisión Nacional de Investigación Científica y Tecnológica – Government of Chile (CONICYT scholarship to MF).
Author information
Authors and Affiliations
Contributions
DKF, JMT, and LP conceptualized the manuscript. DKF wrote the manuscript, designed all Figures, and curated gene lists. HMD, MF, JMT, and LP contributed critical comments and reviewed the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Immuno-ageing, longevity/ageing, and immunity genes. To define immuno-ageing genes (‘ImmAg’ sheets) we assembled all currently known immunity and longevity/ageing genes (‘Imm’ and ‘Ag’ sheets) from various databases and identified genes occurring in both traits (see ‘Summary’ tab). This was done for two invertebrates, C. elegans and D. melanogaster (‘Cele’ and ‘Dmel’ sheets), and two mammals, M. musculus and H. sapiens (‘Mouse’ and ‘Human’ sheets). Ageing genes were obtained from the GO term ‘aging’ (GO:0007568), the KEGG pathway annotation ‘Longevity regulating pathway’ (KEGG id: 04213), and two ageing databases GenAge and AgeFactDB. Immunity genes were compiled from the GO term ‘immune system process‘ (GO:0002376), multiple immune-related KEGG pathways (for D. melanogaster only available KEGG id: 04624; while for mammals we used: 04640, 04610, 04611, 04620, 04621, 04622, 04623, 04625, 04650, 04612, 04660, 04658, 04659, 04657, 04662, 04664, 04666, 04670, 04672, and 04062), and the three immunity databases insect innate immunity database IIIDB, InnateDB, and immunome knowledge base (IKB). Genes from GO and KEGG were obtained using the biomaRt package in R, while genes from databases were downloaded from the corresponding websites. We added additional annotations (such as IDs from multiple databases) using biomaRt, and the species-specific browsers WormBase for C. elegans, FlyBase for D. melanogaster, MGI for M. musculus, and HGNC for H. sapiens. For AgeFactDB, we excluded all genes for which the experimental evidence was annotated as ‘putative’ or ‘no’. Annotations were obtained in January 2019 for IIIDB and Immunome, and August 2020 for all others. Additional genes with clear effects on immunity and ageing, which we found reviewing the literature, were manually added for C. elegans and D. melanogaster (‘handcurated’ column; references below). The number of genes obtained from each database are given in the ‘Summary’ sheet. The column ‘Longevity’ in the gene lists indicates whether a gene is beneficial (pro) or detrimental (anti) for lifespan or had no effect (none) as defined by the GenAge database.
Additional file 2: Table S2.
Shared immuno-ageing orthologs. The table gives details to the 437 human orthologs/genes in the partitions of the cross-species VennDiagram in Fig. 2. Human orthologs of immuno-ageing genes of all species were obtained using DIOPT (Option: Return only best match when there is more than one match per input gene or protein). The species and count of species sharing a human immuno-ageing ortholog are given in the column “Overlap” and “SpeciesCount”, respectively.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Fabian, D.K., Fuentealba, M., Dönertaş, H.M. et al. Functional conservation in genes and pathways linking ageing and immunity. Immun Ageing 18, 23 (2021). https://doi.org/10.1186/s12979-021-00232-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12979-021-00232-1