
Abstract
The liver, the largest organ in the human body, plays a multifaceted role in digestion, coagulation, synthesis, metabolism, detoxification, and immune defense. Changes in liver function often coincide with disruptions in both the central and peripheral nervous systems. The intricate interplay between the nervous and immune systems is vital for maintaining tissue balance and combating diseases. Signaling molecules and pathways, including cytokines, inflammatory mediators, neuropeptides, neurotransmitters, chemoreceptors, and neural pathways, facilitate this complex communication. They establish feedback loops among diverse immune cell populations and the central, peripheral, sympathetic, parasympathetic, and enteric nervous systems within the liver. In this concise review, we provide an overview of the structural and compositional aspects of the hepatic neural and immune systems. We further explore the molecular mechanisms and pathways that govern neuroimmune communication, highlighting their significance in liver pathology. Finally, we summarize the current clinical implications of therapeutic approaches targeting neuroimmune interactions and present prospects for future research in this area.
Similar content being viewed by others


Introduction
The nervous and immune systems are crucial for survival. Traditionally, these systems were believed to function independently; however, accumulating scientific evidence suggests an active and beneficial dialogue between them [1]. The liver, innervated by autonomic and sensory fibers of the sympathetic nervous system (SNS) and parasympathetic nervous system (PSNS), plays a pivotal role in various life-sustaining functions, including digestion, coagulation, synthesis, metabolism, detoxification, and immune defense [2, 3]. Liver injury, responsible for approximately 2 million deaths worldwide annually, can result from toxins, metabolic disorders, infections, and genetic diseases [4]. The initial presentation of most chronic and many non-fulminant acute liver diseases is often asymptomatic or involves nonspecific signs and symptoms, such as fatigue, anorexia, and weight loss [5,6,7]. It is only when the liver capsule or intrahepatic bile duct is involved in inflammation, such as in Fitz-Hugh-Curtis syndrome [8] and hepatolithiasis, [9] that patients complain of right upper quadrant pain. Liver transplantation leads to effective liver denervation, which may contribute to higher rates of obesity, dyslipidemia, and diabetes in liver transplant recipients [10,11,12,13]. These observations suggest a strong communication between the liver (including the liver capsule, hilar region, portal region, and liver parenchyma) and the nervous system.
Although the role of the hepatic nervous system in liver homeostasis is increasingly appreciated, much remains unknown about the specific mechanisms by which hepatic nerves influence and are influenced by liver diseases. Both human and rodent livers are innervated by the autonomic nervous system (ANS). While the human liver is innervated by the SNS and PSNS, the rodent liver may be innervated only by the former [14]. The liver possesses a finely tuned cellular immune system, including innate immune cells such as Kupffer cells (KCs), monocyte-derived macrophages, dendritic cells (DCs), natural killer (NK) cells, natural killer T cells, and adaptive immune cells, such as T and B cells. Disruption of the hepatic immune system can cause autoimmune hepatitis, steatohepatitis, cirrhosis, and liver cancer [15]. Although the positional relationships between the nerves and immune cells in the liver are unclear, the role of neuroimmunology in liver diseases has been explored through surgical and genetic techniques. Given the fundamental connection between the nervous system and liver, understanding the role of neuroimmune innervation in liver dysfunction holds promise for developing novel therapeutic approaches for liver diseases.
In this concise review, we provide a brief description of the structure and composition of the hepatic neural and immune systems. Subsequently, we introduce the molecular mechanisms and pathways that mediate neuroimmune communications and discuss their significance in liver ailments. Lastly, we summarize the current clinical implications of therapeutic strategies targeting neuroimmune interactions and propose future research directions.
Hepatic nervous and immune systems
Hepatic neuroanatomy
The neural circuitry connecting the nervous system and liver is intricate. The centrifugal neural circuits are divided into the central nervous system (CNS), which includes the brain and spinal cord, and the peripheral nervous system (PNS), comprising somatic and autonomic nerves [16]. The somatic nerves, originating from the CNS, regulate skeletal muscles and elicit voluntary movements [17]. The ANS consists of the SNS, PSNS, and enteric nervous system (Fig. 1) [18]. The afferent neural circuits are broadly divided into afferent vagus and spinal sensory nerves [19]. The cell bodies of the afferent vagus nerves are situated in two nodose ganglia on either side of the neck, with the left and right vagus nerves innervating visceral organs asymmetrically [20].

The classification of the nervous system and the summary of peripheral nervous system (PNS). A Classification of the nervous system. B Summary of efferent pathways in the PNS. The autonomic nervous system is organized in three anatomical and biochemical distinct systems, the sympathetic nervous system (SNS, in red), the parasympathetic nervous system (PNS, in blue) and the enteric nervous system (ENS, Fig. 1D), respectively. C Summary of the afferent pathways in the PNS (in the liver and gut). D Schematic diagram of the ENS. The ENS is located within intestinal tissues and has a characteristic architecture. CNS central nervous system, ENS enteric nervous system, PNS peripheral nervous system, PSNS parasympathetic nerve system, SNS sympathetic nerve system, III the third cranial nerve, VII the seventh cranial nerve, IX the ninth cranial nerve, X the tenth cranial nerve
Autonomic nerves, both afferent and efferent, innervate various parts of the liver, including the portal vein, hepatic artery, bile ducts, and liver hilum [21]. The anterior plexus, originating from the left portion of the celiac plexus and the right abdominal branch of the vagus, forms a network of nerves surrounding the hepatic artery. The posterior plexus, derived from the right portion of the celiac plexus, surrounds the portal vein, occasionally extending innervations accompanying the hepatic vein (Fig. 2A). The distribution of nerves within the liver is highly species-specific, with most species exhibiting innervation of the portal tract [13]. For instance, while the human liver exhibits both intra-acinar and portal tract innervation, mice and rats primarily display portal tract innervation (Fig. 2B) [14]. The distribution of nerves within the liver in various species, such as rats, rabbits, monkeys, guinea pigs, pigeons, quail, golden hamsters, and turtles, has been thoroughly reviewed in previous studies [13].

Hepatic neuroanatomy. A Origin and concomitant of hepatic plexus. The anterior plexus, originating from the left portion of the celiac plexus and the right abdominal branch of the vagus, forms a network of nerves surrounding the hepatic artery. Meanwhile, the posterior plexus, derived from the right portion of the celiac plexus, is located around the portal vein with occasional innervations accompanying the hepatic vein. B Anatomy of the intrinsic sympathetic and parasympathetic nerve fibers. In all mammalian species, sympathetic and parasympathetic fibers surround the portal are an humans and guinea pigs but not rats. sympathetic fibers course into liver sinusoids. C Innervation of the human liver. Within the liver, nerves are found around the hepatic artery, portal vein, and bile ducts. Sympathetic nerves, but not parasympathetic nerves, synapse on hepatocytes as well. LSEC liver sinusoids endothelial cell
Hepatic immune system
The mammalian liver is a multifaceted organ with diverse cell types that perform numerous physiological functions, [22,23,24] including digestion, synthesis, metabolism, and detoxification. Additionally, the liver plays a vital role in immune processes, such as anti-infection, maintaining autoimmune stability, and supporting antitumor activities (Fig. 3A) [25,26,27]. One of the liver's unique immunological advantages stems from its parenchymal and nonparenchymal cells, which actively contribute to immunoregulation and the maintenance of overall homeostasis. As a fundamental immune organ, the liver receives blood directly through the portal vein, which drains not only the peritoneum but also the gastrointestinal (GI) tract, pancreas, and spleen [28]. This direct blood supply bypasses classic immune surveillance sites such as lymph nodes [29]. Notably, the liver is more exposed to microorganisms and endotoxins than other organs, given its distinctive anatomical location and unique blood composition (mainly derived from the portal vein and enriched with antigens from the GI tract) [30]. Consequently, the liver typically maintains a tolerogenic immune state to prevent unnecessary immune activation and excessive autoimmune responses [31, 32]. However, during acute and chronic infections, the liver transitions from a tolerant state to an active immune state, constituting a crucial line of defense against invading microorganisms [33,34,35,36]. Hepatic defenses depend on a complex network of immune cells, including KCs, DCs, neutrophils, NK cells, and B and T lymphocytes [37]. Furthermore, non-immune cells, such as hepatocytes, cholangiocytes, hepatic stellate cells (HSCs), and liver sinusoidal endothelial cells play pivotal roles in recruiting proinflammatory immune cells and concurrently producing inflammatory cytokines, including acute-phase proteins, complement factors, and cell adhesion molecules (Fig. 3B) [38,39,40].

Hepatic immune system and functions. A Functions of the liver. The mammalian liver is a complex organ consisting of diverse cell types that perform multiple physiological functions including digestion, detoxication, metabolism, biotransformation, and immune. B Hepatic immune system. The liver’s immune defense mechanism primarily depends on a complex network of immune cells and non-immune cells, the latter of which play immune roles by recruiting proinflammatory immune cells and simultaneously producing inflammatory cytokines
Mechanisms of neuroimmune interaction in the liver
The interactions between neurons and immune cells are intricate and multifaceted, necessitating a sophisticated infrastructure to enable signal propagation between these two systems. In the following sections, we delve into the principal pathways through which the nervous and peripheral immune systems communicate and influence each other (Fig. 4A). Each pathway serves as a distinct conduit for transmitting specific information, employing unique mechanisms to convey messages.

Neuronal regulation of immune activation. A Outline. An overview of inter-regulation of the nervous system and the immune system. B Endocrine pathways. The endocrine signals enable the synchronization of complex physiological processes and immune activity. C Neuronal efferent pathways. Noradrenaline (recognized by α-adrenergic and β-adrenergic receptors) is the primary neurotransmitter of the SNS, while acetylcholine (ACh; recognized by nicotinic and muscarinic ACh receptors) is the primary PSNS neurotransmitter. Both of which can affect activity of immune organs and immune cells. D Meningeal lymphatic vessels. The meningeal lymphatic system can impact both the central and peripheral immune response by transporting immune cells and antigens from the brain to the peripheral immune system. ACTH adrenocorticotropic hormone, ANS autonomic nervous system, Ach acetylcholine, AI Alzheimer’s disease, AVP arginine vasopressin, CNS central nervous system, CCL21 C–C motif chemokine ligand 21, Epi epinephrine, FSH follicle-stimulating hormone, GH growth hormone, LH luteinizing hormone, M-R muscarinic cholinergic receptor, MS multiple sclerosis, N1-R nicotinic cholinergic receptor type 1, NA norepinephrine, OXT oxytocin, PVN paraventricular nucleus, PRL prolactin, PNS peripheral nervous system, PSNS parasympathetic nerve system, PROX1 prospero homeobox 1, SON supraoptic nucleus, TSH thyroid stimulating hormone, VEGFR3 also known as “FLT4,” fms related receptor tyrosine kinase 4, α/ꞵ-R adrenergic cholinergic receptor
Endocrine pathways
The endocrine system is a powerful tool at the brain’s disposal, regulating many physiological processes. Hormones, controlled by the brain, are released into the bloodstream and transported to target tissues and immune cells. This rapid and efficient pathway simultaneously delivers information to various organs and synchronizes physiological processes. At the center of this system is the hypothalamus, which communicates extensively with other brain regions and governs vital processes, such as hunger, thirst, body temperature, circadian rhythm, and sleep.
The hypothalamus regulates two primary endocrine pathways: the hypothalamic–neurohypophyseal system and the hypothalamic–hypophyseal portal system. In the hypothalamic–neurohypophyseal system, hormones, including oxytocin [41] and arginine-vasopressin (AVP), [42] are produced and secreted. Oxytocin suppresses proinflammatory cytokines [43, 44] and promotes wound healing, [45] while AVP exerts anti-inflammatory effects during sepsis [46]. The activation of AVP-producing neurons by inflammatory cytokines indicates that this endocrine pathway can respond to changes in immune system activity [47].
The hypothalamic–hypophyseal portal system secretes hypothalamic hormones into the bloodstream to ultimately reach the target organ, inducing the release of the final effector hormone. There are five main axes, [1] including the (1) hypothalamic–pituitary–adrenal axis; (2) hypothalamic–pituitary–thyroid axis; (3) hypothalamic–pituitary–gonadal axis; (4) hypothalamic–pituitary–somatotropic axis; and (5) hypothalamic–pituitary–prolactin axis. These endocrine pathways coordinate physiological and developmental events that necessitate adjustments in immune activity, thus influencing the immune response. For instance, sex hormones play a role in the programming of the immune system [48, 49]. Testosterone generally has an immunosuppressive effect, whereas estrogen enhances immune responses [50]. The immunoenhancing effects of estrogens include promotion of neutrophil infiltration, elevation of macrophage phagocytic activity, enhancement of DC antigen-presenting function, promotion of Th2 differentiation, and contributions from T-lymphocytes, B-lymphocytes, cytotoxic CD8 cells, NK cells, and ILC-2 cells [51,52,53]. Consequently, females exhibit increased antibody production [54, 55] and are less susceptible to viral infections but more prone to autoimmune diseases [56, 57]. Androgens suppress the immune response through mechanisms such as influencing neutrophil maturation and function, decreasing TNF-α and IL-1β expression and secretion in macrophages, negatively regulating B- and T-cell development, reducing toll-like receptor expression, and decreasing cytokine production [51, 58]. These mechanisms partly explain the increased susceptibility of males to various parasitic, [59] bacterial, [60] and viral infections [61, 62]. Similarly, androgen-mediated suppression of immune reactivity and inflammation lowers the threshold for cancer, making men more susceptible to cancer than women [63]. Diseases such as primary biliary cirrhosis (PBC) and autoimmune hepatitis (AIH) have a strong female predominance, whereas primary sclerosing cholangitis (PSC) has a higher incidence in males with a more severe course [64]. The strong association between sex and the development and course of autoimmune liver diseases could be partially explained by differences in sex hormones [65]. Another example is the hypothalamic–pituitary–thyroid axis, which boosts metabolic activity and is linked to immune activation [66]. Thyroid hormones induce lymphocyte proliferation [67]. Consequently, the immune response is suppressed by thyroidectomy, [68] and patients with hypothyroidism are more susceptible to infection [69, 70]. Thus, endocrine signals synchronize complex physiological processes and immune activity (Fig. 4B).
Neuronal efferent pathways
The human CNS communicates rapidly and directly with peripheral tissues through the PNS, comprising 12 cranial nerves, 31 spinal nerves, and the ANS [71]. The ANS regulates functions not under conscious control, such as blood pressure, heart rate, and GI motility and function. The SNS and PSNS have opposing effects on various physiological processes; the SNS regulates the “fight or flight” response, while the PSNS regulates the “rest and digest” state. Many organs, including the bone marrow, lymphatic tissues and organs, joints, blood vessels, lungs, GI tract, liver, kidneys, and spleen, are innervated by the ANS [17].
Noradrenaline, acting through α-adrenergic and β-adrenergic receptors, is the primary neurotransmitter of the SNS, whereas acetylcholine (ACh), acting through nicotinic and muscarinic receptors, is the primary neurotransmitter of the PSNS. Although both systems can influence immune activity, immune organs such as the bone marrow, thymus, lymph nodes, and spleen are predominantly innervated by the SNS [72]. Immune cells, such as NK cells, DCs, T cells, and macrophages, express receptors for neurotransmitters and neuropeptides secreted by the ANS [73, 74]. Notably, immune cells themselves can also secrete various neuronal factors, [74,75,76] revealing an intricate interplay between the nervous system and immunity (Fig. 4C).
Meningeal lymphatic vessels
The brain can regulate immune responses by introducing brain-specific antigens to the peripheral immune system. Immune cells continually monitor all tissues, residing or patrolling within them to gather information about the tissue's condition and potential threats. These cells then travel through lymphatic vessels to the lymph nodes, where they present antigens and trigger a relevant immune response [77]. Recent research has identified dural lymphatic vessels that transport antigens and immune cells from the brain to the lymph nodes and express lymphatic endothelial markers, such as PROX1, CCL21, and VEGFR3 [78]. The lymphatic system envelops the brain, facilitating the drainage of excess fluid, proteins, and immune cells from the brain tissue to peripheral lymph nodes [79]. In a mouse model of multiple sclerosis, the removal of this meningeal lymphatic system led to a reduction in CNS disease and a decrease in T cell inflammatory responses [79]. Similarly, in a transgenic mouse model of Alzheimer's disease, disruptions in meningeal lymphatic vessels promoted meningeal amyloid-β deposition and increased parenchymal amyloid-β accumulation (Fig. 4D) [80]. Therefore, by transporting immune cells and antigens from the brain to the periphery, the meningeal lymphatic system can impact both central and peripheral immune responses.
Immune regulations of neuronal function
Cytokine pathways
The recognition of immune influences on the nervous system has become more evident, though it initially lagged behind our understanding of how the nervous system regulates the immune system. This delay was primarily due to the recent discovery of the molecular mechanisms governing immune communication signals and pathways, particularly cytokines [81]. Cytokines, primarily released by immune cells, notably T cells and myeloid cells, play a pivotal role in active immune responses [81]. Afferent fibers of the ANS can detect cytokines and transmit this information to the brain, forming a "neural" communication pathway [82]. For instance, in adult mice, the intraperitoneal administration of tumor necrosis factor (TNF) or interleukin-1β significantly alters the action potentials originating from the cervical vagus nerve [83]. Cytokines can also directly activate and sensitize neurons, leading to the generation of action potentials that convey information to the brain (Fig. 5) [84,85,86,87]. Under inflammatory conditions, cytokines may directly reach parenchymal brain centers via a “humoral” route, particularly when the blood–brain barrier (BBB) exhibits increased permeability [88].

Immune regulation of neuronal function. The pathways in which immune influences on the nervous system consist of cytokines, afferent neural inputs, and chemoreceptors (including osmoreceptors, ion receptors, and metabolic substance receptors). DAPMs damage-associated molecule patterns, GLUT2 glucose transporter 2, NKCC1 Na +-K+-2Cl− cotransporter, PKC protein kinase C, PAMPs pathogen-associated molecular patterns, TRPV4 transient receptor potential vanilloid 4
Afferent neural inputs
Peripheral primary afferent neurons express receptors for microbial constituents and inflammatory products, indicating their role as a hardwired neural connection that signals inflammation or infection from the periphery to the CNS (Fig. 5). The sensory nervous system is a crucial component of afferent neural circuits, transmitting signals from peripheral tissues to the brain and spinal cord [89]. Various pathogen-associated molecular patterns, such as unmethylated cytosine and guanine and lipopolysaccharide (LPS), as well as danger-associated molecular patterns including DNA, histones, HMGB1, adenosine triphosphate, uric acid, and cytokines, can be detected by sensory nerves [90]. In regions of infection or injury, the activation of peripheral immune cells and the release of cytokines and other inflammatory molecules influence sensory neurons, either triggering or altering signals sent to the spinal cord and brain [91].
Furthermore, sensory neurons in the liver play a role in detecting nutritional and absorptive hormones known as incretins. One prominent incretin, glucagon-like peptide 1 (GLP-1), is sensed by afferent nerves in the portal vein. Activation of GLP-1 receptors stimulates hepatic vagal afferent nerves, leading to changes in pancreatic insulin release through efferent sympathetic nerve signals originating in the brain [92, 93]. Hepatic afferent fibers also express receptors for several hormones secreted by the GI tract, including somatostatin, [94] leptin, [95] and cholecystokinin [96].
In addition to their primary function of transmitting information to the brain, these neurons locally release neuropeptides. Sensory fibers have been extensively studied for their ability to communicate with immune cells, [97] particularly in barrier tissues, such as the skin, [98, 99] lungs, [100] and gut, [101, 102] where immune cells are abundant. These sensory innervations directly affect immune cell activity, providing protection against endotoxemia [103] and sepsis [104]. Therefore, the sensory nervous system has both systemic and local effects on immunity, conveying sensory inputs to the brain to regulate the peripheral immune response and secreting neuropeptides directly into the tissues [104].
Chemoreceptors
The liver serves as an ideal sensor for monitoring osmolality, glucose concentration, and fatty acid levels due to its close connection with the GI tract. Following the absorption of macronutrients from the GI tract, they travel through the portal vein to reach the liver. Within the liver, numerous chemoreceptors play a critical role in sensing ions and nutrients and subsequently transmit signals to the brain. Additionally, several chemoreceptors are multifacetedly correlated with immune response or even neuro-immune communication. These chemoreceptors encompass osmoreceptors, ion receptors, and receptors for various metabolic substances (Fig. 5) [14].
Osmoreceptors
While the brainstem is responsible for detecting disturbances in osmolarity, the liver harbors peripheral osmoreceptors that are quite effective at detecting changes in blood osmolality. This is particularly relevant because the liver can detect changes in water intake before these changes affect systemic blood osmolality [105]. Neurons adjacent to hepatic blood vessels are responsible for detecting and responding to shifts in osmolality within the liver, [105] relying on osmotically-activated ion channels such as transient receptor potential vanilloid 4 (TRPV4) [105]. TRP channels potentiate the inflammatory response following hepatocellular injury by controlling the functions of Kupffer cells, monocytes, and neutrophils, including cytokine and chemokine production, response to chemokines, adhesion, and migration from the bloodstream to injured hepatic sites [106]. Impaired osmoregulation of vasopressin also occurs in the acute phase of severe sepsis, resulting from a depolarization of osmoreceptors, which leads to sick behaviors caused by brain-immune crosstalk dysfunction in response to stress, indicating a crucial role of osmoreceptors in neuro-immune communication [107]. Activation of hepatic osmoreceptors is linked to various physiological and clinical parameters. For instance, water consumption has been associated with outcomes such as weight loss, [108] hepatic denervation, [109] and orthostatic and postprandial hypotension [105].
Ion sensation
The liver possesses receptors that monitor fluid homeostasis by detecting key physiological ions in the portal blood system. Similar to osmoreception, the hepatic nervous system identifies changes in ion concentrations before they impact the entire system [110]. The liver holds a distinct advantage in ion detection compared to other organs, given that the concentration of absorbed substances in the portal vein is 4–5 times higher than what is observed in the systemic blood [111]. Consequently, the liver serves as an ideal "predictor" of systemic blood ion concentrations, allowing for the anticipation of abnormal fluid homeostasis. Patients with cirrhosis commonly exhibit ion imbalances such as hyponatremia or hypochloremia, stemming from the loss of the hepatorenal reflex. Ions serve as critical neurotransmitters in the neuroimmune network in various organs such as the brain and kidneys. Calcium sensing via microglial UDP-specific type 6 purinergic (P2Y6) signaling pathways was reported to promote microglia phagocytosis and shape neuroimmune responses in epileptogenesis [112]. However, the role of ions in liver neuroimmune communication needs further investigation.
Metabolic sensing
Hepatic afferent fibers establish a negative feedback loop involving feeding behavior and metabolite level alterations. Peripheral glucose sensors are distributed in various locations, including the taste buds, intestines, carotid bodies, and liver [113,114,115]. Emerging evidence points to the wall of the portal vein as a probable site for glucose sensing [116]. In the liver, GLUT2 serves as a hepatoportal glucose sensor with a role in hyperglycemia detection [35]. A Glut2–/– knockout mouse model demonstrated a failed hypoglycemic response to portal glucose infusion [35]. However, the exact molecular pathway responsible for hepatic lipid sensing remains undefined. One potential pathway involves the activation of protein kinase C (PKC) δ, θ, and ε [117]. Notably, lipid activation of PKCs is not limited to the liver, as PKC-δ is activated during lipid sensing in duodenal cells as well [118]. This pathway’s activation regulates glucose production through a gut–brain–liver mechanism. A study by Seillet et al. elegantly demonstrated the interdependence of the neuro-immune-metabolic network in steady-state conditions with respect to lipid metabolism [119]. The key finding is that intrinsic cellular rhythms acted in synergy with the cyclic patterns of food intake to drive the production of IL-22 and synchronize the protection of the intestinal epithelium through a vasoactive intestinal peptide (VIP)-vasoactive intestinal peptide receptor 2 (VIPR2) pathway in group 3 innate lymphoid cells (ILC3s). IL-22 released by the ILC3s improves insulin sensitivity and can regulate lipid metabolism in the liver and adipose tissue [120]. The combined input of afferent signals from the gut and liver may trigger hypothalamic activity during food intake [121]. Ongoing research should further investigate the potential intersections of lipid- or other metabolite-induced signaling in the brain, liver, gut, and other organs.
Interaction between liver pathology and neuroimmunity
The interplay between liver pathology and neuroimmune disorders is complex. Although the exact connection between nerves and immune cells within the liver remains elusive, the field of neuroimmunology has investigated liver diseases through surgical and genetic techniques. Recent findings indicate that the nervous system plays a pivotal role in the onset and progression of various liver diseases. Liver diseases frequently coincide with dysfunction in both the CNS and PNS. Chronic inflammatory liver diseases and severe decompensated cirrhosis are often accompanied by behavioral alterations, including fatigue, mood disorders, cognitive dysfunction, and sleep disturbances. In this section, we review several diseases indicative of the interaction between liver pathology and neuroimmunity.
Neuroimmune crosstalk in liver pathology
Liver injury
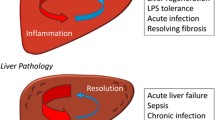
The liver is susceptible to various internal and external stressors, often resulting in a cascade of events including inflammation, repair, regeneration, and fibrosis. These processes are typically driven by a chronic inflammatory microenvironment. In studies involving mice, sympathetic nerve ablation using 6-hydroxydopamine (6-OHDA) prior to carbon tetrachloride (CCl4) treatment has been shown to reduce the expression of proinflammatory cytokines, steatosis, and necrosis [122]. Hypertensive rats with increased peripheral sympathetic activity [123] are more susceptible to CCl4-induced liver toxicity, developing cirrhosis after just 4 weeks of CCl4 administration, whereas control rats display less severe bridging fibrosis [124]. When hepatocytes are damaged by CCl4 exposure, they undergo apoptosis, and sympathetic denervation increases hepatocyte apoptosis while reducing hepatocyte proliferation [125]. Preventing apoptosis through sympathetic signaling could lead to the accumulation of damaged cells that produce persistent inflammation and increased fibrosis [125]. Therefore, β-receptor agonists and α1-receptor antagonists have demonstrated protective effects in models of acetaminophen-induced liver injury [126, 127].
In contrast, parasympathetic neurons exert a protective effect on hepatic inflammation by acting through α7nACh receptors expressed on macrophages. In studies involving mice treated with anti-Fas, hepatic vagotomy significantly increases mortality; however, co-treatment with an α7nACh receptor agonist attenuates this effect [128]. Vagal signaling also decreases inflammation in LPS-induced hepatitis, likely through macrophage-expressed α7nACh receptors [129]. In murine acetaminophen-induced liver injury, acetylcholinesterase inhibitors improve survival and reduce hepatocellular inflammation, damage, and apoptosis [130]. Therefore, in multiple models of chemical liver injury, vagal signaling plays a protective, anti-inflammatory role, whereas sympathetic neurons promote inflammation and liver damage (Fig. 6A).

The neuroimmune crosstalk in liver pathology. The sympathetic nervous system (SNS) and parasympathetic nervous system (PSNS) play a crucial role in the development and progression of liver diseases, including liver injury (A), liver fibrosis (B), MAFLD (C), and HCC (D). The two 3D imagines of immunolabeling in the “NAFLD” part are from ref 158. HSCs hepatic stellate cells, HCC hepatocellular carcinoma, IL-6 interleukin 6, MAOA monoamine oxidase A, MAFLD metabolic dysfunction-associated fatty liver disease, NAFLD non-alcoholic fatty liver disease, PSNS parasympathetic nervous system, SNS sympathetic nervous system, TNF-α tumor necrosis factor α, 6-OHDA 6-hydroxydopamine, α7nAChR α7 nicotinic acetylcholine receptor, α/ꞵ-R adrenergic cholinergic receptor
Liver fibrosis
Despite advances in research on the pathogenesis and treatment of liver diseases, effective therapies for liver fibrosis and cirrhosis remain elusive. Intrinsic parenchymal innervation diminishes in the pre-cirrhotic liver and is nearly absent in the regenerating nodules of established cirrhosis [131, 132]. Nevertheless, nerves persist along fibrous septae and in portal tracts within pre-cirrhotic and cirrhotic livers [131, 132]. Among these, substance P- and neuropeptide Y-positive nerve fibers tend to predominate over calcitonin gene-related peptide-containing fibers [133]. An analysis of 178 biopsy specimens from cirrhosis patients revealed concurrent increases in the densities of nerve fibers and mast cells, regardless of the underlying cause. Importantly, these density increases in inflammatory cells and nerve fibers correlated with the degree of liver fibrosis in these patients [134]. Similar findings were observed in a rat model of cirrhosis, where nerve terminals closely interacted with myofibroblasts in periseptal sinusoids [135]. In contrast to cirrhosis, which is characterized by widespread hypoinnervation, toxic hepatic injury specifically induces the proliferation of portal nerve fibers [136].
The activation of HSCs, also known as transdifferentiation, is the primary source of myofibroblasts that secrete matrix proteins, thus driving liver fibrogenesis [137]. HSC activation is induced directly or indirectly by paracrine signals from a complex immune and fibrotic microenvironment. Neural signals have long been recognized as a crucial factor in HSC activation and fibrosis progression. Patients with cirrhosis have elevated blood catecholamine levels and increased sympathetic nerve activity [138]. Studies on toxic injury models suggest that the SNS acts on nearby HSCs to exert a proinflammatory and profibrotic effect. Activated HSCs express α-adrenergic, β-adrenergic, and neuropeptide Y receptors, [139] and catecholamines induce HSC proliferation and extracellular matrix protein production in vitro [140, 141]. Mice lacking dopamine-β-hydroxylase, an enzyme essential for norepinephrine production, show reduced accumulation of hepatic α-smooth muscle actin when exposed to an antioxidant-deficient diet that induces fibrosis in control mice [142].
The role of the PSNS in liver fibrosis has not been fully investigated. However, the parasympathetic neurotransmitter Ach promotes the proliferation of myofibroblast-like HSCs and induces the expression of collagen genes in HSCs [143]. These effects are abolished by mecamylamine, an nACh receptor antagonist, suggesting that the influence of Ach on HSC proliferation is partly mediated through nACh receptors (Fig. 6B) [144]. Advanced technologies, such as single-cell and spatial multiomics, multicolor fluorescence, and optogenetics, are needed to better understand the role of neuroimmune crosstalk in the development and progression of liver fibrosis.
Metabolic dysfunction-associated steatotic liver disease
Non-alcoholic fatty liver disease (NAFLD) is a hepatic manifestation of metabolic syndromes and a major risk factor for liver injury, fibrosis, and even liver cancer [145, 146]. NAFLD is currently estimated to affect 38% of the global population [147] and is expected to become a major indication for liver transplantation [148] over the next 20 years. Recently, important conceptual advances have been made in understanding the complex pathophysiological mechanisms of this highly prevalent liver condition. In 2023, following four rounds of the Delphi survey, three large multinational liver associations proposed that metabolic dysfunction-associated steatotic liver disease (MASLD) should replace the term NAFLD [149]; this proposition garnered wide acclaim, albeit with some dissent [150]. The new nomenclature/criteria of MASLD is more suited for lean patients with NAFLD than the previously suggested metabolic dysfunction-associated fatty liver disease (MAFLD) criteria [151]. This nomenclature change better reflects the pathophysiology and cardiometabolic implications of this common and burdensome liver disease [152]. Emerging evidence suggests an excellent concordance rate between NAFLD and MASLD definitions, with over 99.5% of patients with NAFLD meeting the MASLD criteria [153]. Younossi et al. conducted research showing similar clinical profiles and mortality rates for MASLD and NAFLD, which provided evidence that data generated over the past three decades for NAFLD can be used interchangeably for MASLD [154]. Consequently, it is reasonable to consider that the findings and natural history data from older NAFLD studies remain valid under the new MASLD definition [155].
Increased sympathetic activity is predictive of the development of metabolic abnormalities, whereas reduced parasympathetic activity is linked to metabolic syndrome [156,157,158]. Autonomic dysfunction is more common in patients with NAFLD (MASLD) than those without [159, 160], suggesting that early autonomic dysfunction may exacerbate the development and progression of NAFLD (MASLD). Chemical sympathetic denervation can decrease the expression of gluconeogenic enzymes and peroxisome proliferator activated receptor alpha in the liver, reducing high-fat diet (HFD)-induced hepatic steatosis [161]. Aging and sympathetic nerve activity are believed to play significant roles in the accumulation of liver fat, with the inhibition of sympathetic activity resulting in increased fat accumulation. The cholinergic anti-inflammatory pathway, regulated by the vagus nerve, provides protection against inflammation associated with obesity and other metabolic complications [162]. KCs, which are liver macrophages, promote liver inflammation through the innate immune response via toll-like receptor (TLR) signaling. Therefore, controlling KC activity may inhibit the progression of NAFLD (MASLD).
Liu et al. used advanced 3D imaging to examine neural distributions in mice, nonhuman primates, and human livers [163]. They observed that neural innervations within the liver are predominantly sympathetic; moreover, they discovered profound and reversible loss of sympathetic innervations during metabolic challenges. This hepatic sympathetic neuropathy was induced by TNFα, produced by CD11b+ F4/80+ immune cells in response to an HFD. The researchers also found that deleting Sarm1 alleviated the hepatic sympathetic neuropathy and improved metabolic parameters in mice challenged with an HFD. The study revealed that the sympathetic neurotransmitter norepinephrine suppressed immune cell inflammation, which otherwise would cause insulin insensitivity in hepatocytes (Fig. 6C). These findings shed light on a previously unknown neuropathic event in the liver with metabolic implications [163]. Although the ANS is intricately connected to and interacts with the liver, its precise role in the development and progression of NAFLD (MASLD) remains unclear, especially in the context of the new MASLD definition. Further research is strongly encouraged to develop new neuroimmune biomarkers and drugs against MASLD to ensure patients benefit the most from the changes in disease nomenclature.
Hepatocellular carcinoma (HCC)
Liver cancer stands as the third leading cause of cancer-related mortality worldwide, [164] and HCC accounts for approximately 90% of all liver cancers [165]. The immune components within the tumor microenvironment of HCC are notably intricate, featuring diverse populations of myeloid cells and lymphocytes that play roles in inflammation, immune evasion by cancer, and responses to immunotherapy [166]. HCC is an aggressive cancer, frequently diagnosed late in the disease course when effective treatment options are limited. Fortunately, immunotherapies such as immune-checkpoint inhibitors are revolutionizing cancer management.
The ANS is being increasingly recognized as a significant contributor to tumor progression and metastasis [167, 168]. High sympathetic fiber density in HCC hepatectomy samples is associated with a poorer prognosis [169]. Monoamine oxidase A (MAOA), an enzyme involved in catecholamine neurotransmitter breakdown, is downregulated in HCC, and higher levels of MAOA are associated with improved and disease-free survival [170]. In a rat model of diethyl nitrosamine-induced HCC, sympathetic denervation using 6-OHDA reduced fibrosis and tumor formation [169]. A recent study, combining single-cell RNA sequencing and flow cytometry, identified FOXP3+ CXCR3+ mucosal-associated invariant T (MAIT) cells in patients with HCC as regulatory MAIT cells (MAITregs) with a high immunosuppressive potential [171]. Additionally, the induction and function of MAITregs were promoted by β1 adrenergic receptor signaling in pre-MAITregs and MAITregs, respectively (Fig. 6D). Together, these findings reveal an immunosuppressive subset of MAIT cells that contribute to HCC progression and suggest a control mechanism through neuroimmune crosstalk [171].
Liver pathology affecting the CNS
Hepatic encephalopathy
Hepatic encephalopathy (HE), a serious complication of both chronic liver disease and acute liver failure, is arguably the most clinically relevant entity affecting the crosstalk between liver immunity and the CNS. HE manifests in a wide spectrum of neurocognitive abnormalities, ranging from acute severe cases to more chronic subsyndromal presentations. Patients with fulminant hepatic failure exhibit rigid extremities (and neck muscles), resistance to passive movements (paratonia), worsening confusion, and agitation. These increased reflexes can diminish or be lost in patients with progressive deterioration, such as those in a coma [172, 173]. In subacute forms of HE or less severe presentations, patients may experience frequent falls, fatigue, disinterest, distraction, attention deficits, psychomotor slowing, and, sometimes, impaired driving abilities. This constellation of less-severe symptoms is termed minimal hepatic encephalopathy [172, 173]. The metabolic disturbances and pathogenic mechanisms of HE are multifaceted and complex, with key contributing factors believed to be the elevation of blood toxins such as ammonia and the accumulation of inhibitory neurotransmitters due to disturbances in nitrogenous substance metabolism in the gut [174].
Overwhelming evidence demonstrates that the gut microbiome significantly impacts the crosstalk between liver immunity and brain function in patients with HE (Fig. 7A) [175,176,177]. The liver–brain–gut axis represents a complex communication system between the gut microbiota, the liver, and the CNS. Dysregulation of this axis has been implicated in the development of several metabolic, inflammatory, neurological, and psychiatric disorders, especially HE [178]. The liver participates in bidirectional communication with the intestine via the portal circulation and bile secretion. Reduced primary bile acid entering the intestine due to liver disease leads to alterations in microbiota composition, bacterial overgrowth, and toxic metabolite production, which in turn exacerbate the course of liver disease [179]. Circulating endotoxins from the gut, together with inflammatory cytokines produced by the gut microbiome and the liver, aggravate systemic inflammation, modulating the impact of ammonia on the brain [180]. Systemic inflammation increases the permeability of the BBB and induces neuroinflammation, leading to microglia activation, astrocyte senescence, and neuronal cell death, subsequently altering neurotransmission and cognitive and motor function in HE [181,182,183]. In addition to inflammation, enteric pathogens and their metabolites affect cognitive behavior through the direct activation of vagal neurons in the enteric nervous system, altering brain-derived neurotrophic factor, gamma-aminobutyric acid, and oxytocin signaling in the brain [184].

Liver pathology affecting the CNS. The possible mechanisms of how liver pathology affects brain cognitive function in diseases of hepatic encephalopathy (A), chronic liver disease (CLD)-associated fatigue (B), depression, and anxiety (C). CLD chronic liver disease, CNS central nervous system, HE hepatic encephalopathy, HCV hepatitis c virus, IFN interferon, NO nitric oxide, PBC primary biliary cirrhosis, TLR2 toll-like receptor 2
The importance of gut microbial composition and function in the potential pathogenesis of HE is underscored by several gut-centric therapies [175]. Fecal microbiota transplantation, probiotics, prebiotics, and antibiotics can reduce serum ammonia levels and intestinal-derived neurotoxic substances by accelerating intestinal transport and changing the metabolism and abundance of intestinal bacteria [176]. A comprehensive understanding of the mechanisms and importance of the gut–brain–liver axis is imperative for developing innovative therapeutic approaches to address HE [185].
Chronic liver disease (CLD)-associated fatigue
Fatigue is typically the most common complaint and one of the most distressing symptoms reported by patients with CLD. It is closely associated with alterations in mood, such as anxiety and depression [186]. Fatigue is a multidimensional symptom experienced as tiredness in the musculoskeletal system, poor recovery from exercise, decreased motivation for usual activities, cognitive decline, fuzzy thinking, lethargy, and exhaustion [187, 188]. CLD-associated fatigue is generally classified into central (CNS dysfunction, marked by a failure to perform voluntary activities or attentional tasks) and peripheral (neuromuscular failure, characterized by muscle wasting and weakness) fatigue. [189]. Although the mechanism is poorly understood, 55–85% of patients with PBC have been reported to present with fatigue, [190, 191] with 20% experiencing significant or life-altering fatigue [192].
Defective release of corticotropin releasing hormone [193, 194] and alteration of serotonergic neurotransmission, mediated by 5-HT receptors [195, 196], have been implicated in the genesis of central fatigue occurring in an experimental model of cholestatic liver disease [197]. Clinical evidence reveals that fatigued patients with PBC exhibit stronger resting-state functional neural connectivity between the basal ganglia and motor and premotor cortices, [198] as well as a lower globus pallidus magnetization transfer ratio, [199] compared to non-fatigued patients with PBC. Communication between liver pathology and changes in endocrine function and structure in the CNS is thought to involve two potential pathways: neural and/or humoral. KCs within the liver secrete proinflammatory mediators, subsequently activating vagal afferent nerves that innervate the liver [186]. Nerve impulses are then carried through the spinal cord to the brain, thus altering neurotransmission either directly or indirectly (via microglia activation) [187]. Circulating cytokines from liver inflammation may directly interact with cerebral endothelial cells and generate secondary signals (e.g., prostaglandins and nitric oxide), creating a milieu of neuroinflammation, which can activate microglia [200]. Additionally, another immune-to-CNS communication pathway was identified, involving the entrance of immune cells into the brain. Circulating immune cells were recruited to the brain during liver inflammation to activate resident immune cells within the CNS (i.e., microglia) and amplify neuroinflammation [201]. Activation of microglia can reduce synapses between neurons in the prefrontal cortex of the brain, inhibit neural transduction, and cause behavioral alterations such as anxiety and fatigue [202]. Additionally, the imbalance of manganese homeostasis in the brain as a result of impaired biliary excretion may be an important mechanism in the genesis of fatigue in patients with pre-cirrhotic PBC (Fig. 7B) [199].
Fatigue in patients with PBC also has numerous associated peripheral features. Autonomic dysfunction, typically in the form of vasomotor abnormality, is common at all disease stages of PBC [203]. Autonomic dysfunction is strongly associated with peripheral fatigue by limiting the capacity of the muscle to respond through increased proton/lactate efflux from cells and outflow from tissues [204]. In addition, the anti-mitochondrial antibody has direct muscle-level metabolic effects, leading to over-utilization of anaerobic metabolism [204]. Other symptoms of patients with PBC can also indirectly affect the occurrence of fatigue. Pruritus, another commonly experienced symptom by patients with PBC, can lead to sleep disorders and worsen the feeling of fatigue [205]. Depression and fatigue are mutually causal and may share a common pathogenesis [206,207,208].
Fatigue in PBC has been mostly assessed with subjective and nonspecific versions of numeric ratings or questionnaires, [209, 210], but no consensus on objective or combined assessments has been reached [188]. Further work is needed to provide more specific and accurate methods to quantify the severity of fatigue and distinguish central from peripheral fatigue. Pharmacological agents to date have not shown to have a significant, reliable effect in reducing fatigue [211]. Consequently, future studies should focus on how the periphery communicates with the brain to mediate changes in central neural activity, enabling the identification of novel therapeutic targets to decrease the burden of debilitating fatigue.
CLD-associated depression and anxiety
Clinical studies have shown that some patients with CLD also experience mental health issues, such anxiety and depression, [212] including symptoms such as emotional suffering, loss of purpose, and impaired ability to function and complete daily activities. For instance, depression and anxiety have high prevalences in patients with hepatitis C [213]. Intracerebral neurobiological changes associated with hepatitis C may potentially explain these symptoms. These changes may arise from infiltration of the brain by peripheral cytokines induced by hepatitis C infection, [214] as well as direct neuropathic effects of hepatitis C viral particles penetrating the BBB. Peripherally-derived cytokines act on the endothelial cells of the BBB, stimulating the secretion of secondary messengers such as prostaglandin E2 into the brain [215]. This stimulates the release of cytokines from activated microglia cells, [216] with subsequent inhibition of neural conduction in the brain [217]. Human brain endothelial cells express specific surface functional receptors (claudin-1, tetraspanin CD81, E-2 glycoprotein, and scavenger receptor-BI) that support hepatitis C viral entry and replication [218]. Viral particles may also be introduced into the brain via infected mononuclear cells and microglia. Activated intracerebral microglia harboring hepatitis C viral infection may have inhibitory or apoptotic effects on neurons through the release of molecules such as pregnalone [219]. Furthermore, the hepatitis C virus core protein promotes neurotoxicity via the sustained activation of extracellular signal-regulated kinase through toll-like receptor 2 signaling [220]. IL-1β polymorphisms have also been confirmed to be associated with depressive symptoms in patients with hepatitis C [221]. Interferons (IFNs), as a highly beneficial therapy in hepatitis C and B, also bring about many neuropsychological disorders such as depression, anxiety, and other symptoms that affect the patient’s quality of life [222]. IFN therapy-related depression can be explained by several mechanisms, including its association with inflammation, neurotransmitter imbalance, neurodegeneration and neuroplasticity, and oxidative stress [223, 224]. Future studies should explore further mechanisms underlying the elevated incidence of depression in patients with hepatitis C, notably IFN-associated depression (Fig. 7C).
PBC, AIH, and PSC represent the three major autoimmune liver diseases [225]. Depression and anxiety have also been observed in patients with PBC and AIH [226]. Depression and anxiety in patients with PBC may have a multifactorial basis. Current supported hypotheses have implicated disrupted neurotransmitter synthesis and changes in brain structure seen on imaging. Tryptophan, as a precursor for the production of the neurotransmitter serotonin (5-hydroxytryptamine [5-HT]), is often associated with depressive mood [227,228,229]. One study indicated that lower levels of tyrosine and tryptophan were significantly associated with higher levels of fatigue and unexpected depressive condition [191]. It is speculated that treatment with steroids could influence the occurrence of depression and anxiety, since this is a well-known side effect of corticosteroid treatment (Fig. 7) [230, 231].
However, it is still unknown whether depression in turn alters the structure of liver tissue or even aggravates liver function. Further research is needed to identify biomarkers common to both biological processes, to elucidate the potential biological correlations between depression and CLD.
Clinical translations
Ongoing research is shedding light on the communication between the nervous and immune systems, with significant implications for clinical applications [91]. Clinical studies are exploring novel therapeutic approaches for inflammatory and autoimmune conditions, including pharmacological modalities and bioelectronic neuromodulation. Neurotransmitter receptor agonists are a key pharmacological modality. For instance, the anti-inflammatory and disease-ameliorating effects of GTS-21, choline, and other α7nACh receptor agonists have been demonstrated in murine models of endotoxemia, sepsis, postoperative brain inflammation, ischemia and reperfusion injury, and various other inflammatory conditions [232,233,234,235,236]. Centrally-acting acetylcholinesterase inhibitors, such as galantamine, have also been evaluated as anti-inflammatory agents in endotoxemia, inflammatory bowel disease (IBD), arthritis, and other metabolic disorders (Fig. 8) [237,238,239].

The Clinical translations and future directions of the neuroimmune crosstalk in the liver. DBS deep brain stimulation, IBD inflammatory bowel disease, TMS transcranial magnetic stimulation, tDCS transcranial direct current stimulation, TUS transcranial ultrasound stimulation, VNS vagus nerve stimulation, α7nAChR α7 nicotinic acetylcholine receptor
Bioelectronic medicine is an emerging area of neuromodulation that aims to identify molecular disease mechanisms that can be targeted therapeutically using neural signals and bioelectronic devices. This field is grounded in the significant role of the vagus nerve in the inflammatory reflex, which regulates peripheral immune function and inflammation. The therapeutic potential of electrical vagus nerve stimulation (VNS) has been demonstrated in preclinical models of rheumatoid arthritis, [240] IBD, and other inflammatory and autoimmune conditions [238, 241]. Bioelectronic devices, such as VNS, deep brain stimulation, transcranial magnetic stimulation, transcranial direct current stimulation, transcranial ultrasound stimulation [242, 243], and splenic nerve modulation [244], have received clinical approval or are under investigation for conditions like epilepsy, depression, Alzheimer’s disease, Parkinson’s disease, dystonia, and other neurodegenerative and neuropsychiatric disorders (Fig. 8). The splenic nerve plays an essential role in mediating the anti-inflammatory effects of VNS in LPS-induced endotoxemia [245, 246]. It has been reported that the anti-inflammatory efficacy of nerve stimulation might be improved by directly targeting the splenic nerve instead of the vagus nerve [247, 248]. An interesting animal study verified that splenic nerve denervation could inhibit LPS-induced depression-like behavior [244]. Collectively, research suggests a role for splenic nerve modulation in inflammation-induced abnormalities. Afferent and efferent vagus nerve signaling may contribute to communication between the brain and spleen (the brain–spleen axis), which may also indirectly participate in regulating liver functions that affect health [249]. Therefore, splenic nerve modulation as a potential therapeutic approach provides new insights into the treatment of liver diseases.
Recent progress in understanding the role of neural pathways in controlling inflammation and other physiological functions, along with advances in neuromodulatory technologies, including electrodes, physiological interfaces, and devices, have paved the way for clinical translation of this knowledge. However, several critical considerations must be addressed for clinical translation. First, evaluating intersubject variability in innervation and neuron origins across different species is essential for designing effective therapies that target specific cell populations. Additionally, understanding the influence of chronic inflammation on organ innervation is crucial, as it can lead to neural changes like sprouting, hypertrophy, and altered neurotransmitter release, affecting neural density and function. These gaps highlight the need for ongoing preclinical and clinical studies to assess inflammation's effects on organ innervation. Lastly, a deeper understanding of the neural circuitry components that regulate specific biological processes is vital for the field of neuroimmune medicine, enabling the simultaneous targeting of multiple sites. Achieving this will require collaborative efforts across various disciplines, including molecular medicine, immunology, biophysics, bioengineering, biochemistry, materials science, mechanical engineering, electrical engineering, computer science, mathematics, and artificial intelligence (Fig. 8).
Conclusions and perspectives
The nervous and immune systems hold pivotal roles in maintaining bodily regulation and defense. In this review, we have explored the experimental evidence showcasing the intricate interplay between these two systems in the context of liver diseases. As neuroscience and immunology progress, our understanding of the molecular mechanisms governing immune regulation undergoes a profound transformation. Delving into the mechanisms of neuroimmune communication has not only expanded our knowledge but has also paved the way for the evaluation of innovative therapeutic approaches. These approaches, including pharmacological modalities and bioelectronic medicine, have shown promise in preclinical and clinical settings for conditions like sepsis, IBD, and arthritis.
However, it is worth noting that neuroimmune-mediated therapies for liver diseases are still in their infancy. While local neuroimmune interactions have been extensively studied, comprehensive techniques that provide a holistic view of neuroimmune interactions between distant organs and the nervous system in vivo are currently limited. Additionally, the molecular mechanisms governing local immune regulation within the nervous system remain elusive.
The future of this field holds immense promise as cutting-edge technologies, such as multiomics, “multi-tissue” organoids, optogenetics, and single-cell and spatial transcriptomics, are shedding light on more precise anatomical connections and functional interactions within the neuroimmune network. These advancements will continue to unveil new insights into the anatomical and functional foundations of neuroimmune units at an organismal level.
In conclusion, ongoing research in the field of neuroimmune interactions promises to unravel the mysteries of how these systems collaborate and will likely yield effective therapeutic strategies for liver diseases and beyond. This interdisciplinary journey, bridging neuroscience, immunology, and advanced technologies, heralds a hopeful future where our understanding of neuroimmune communication transforms into tangible clinical benefits.
Availability of data and materials
Not applicable.
Abbreviations
- ACh:
-
Acetylcholine
- AIH:
-
Autoimmune hepatitis
- ANS:
-
Autonomic nervous system
- AVP:
-
Arginine-vasopressin
- CCl4 :
-
Carbon tetrachloride
- CLD:
-
Chronic liver disease
- CNS:
-
Central nervous system
- DCs:
-
Dendritic cells
- GI:
-
Gastrointestinal
- GLP-1:
-
Glucagon-like peptide 1
- HCC:
-
Hepatocellular carcinoma
- HE:
-
Hepatic encephalopathy
- HFD:
-
High-fat diet
- HSCs:
-
Hepatic stellate cells
- IBD:
-
Inflammatory bowel disease
- IFN:
-
Interferon
- KCs:
-
Kupffer cells
- ILC3s:
-
Group 3 innate lymphoid cell
- LPS:
-
Lipopolysaccharide
- MAFLD:
-
Metabolic dysfunction-associated fatty liver disease
- MAIT:
-
Mucosal-associated invariant T
- MAITregs:
-
Regulatory MAIT cells
- MAOA:
-
Monoamine oxidase A
- MASLD:
-
Metabolic dysfunction-associated steatotic liver disease
- NAFLD:
-
Non-alcoholic fatty liver disease
- NK:
-
Natural killer
- P2Y6:
-
UDP-specific type 6 purinergic
- PBC:
-
Primary biliary cholangitis
- PKC:
-
Protein kinase C
- PNS:
-
Peripheral nervous system
- PSC:
-
Primary sclerosing cholangitis
- PSNS:
-
Parasympathetic nervous system
- SNS:
-
Sympathetic nervous system
- TRPV4:
-
Transient receptor channel protein vanilloid 4
- TNF:
-
Tumor necrosis factor
- VIP:
-
Vasoactive intestinal peptide
- VIPR2:
-
Vasoactive intestinal peptide receptor 2
- 5-HT:
-
5-Hydroxytryptamine
- 6-OHDA:
-
6-Hydroxydopamine
References:
Schiller M, Ben-Shaanan TL, Rolls A. Neuronal regulation of immunity: why, how and where? Nat Rev Immunol. 2021;21(1):20–36.
Mizuno K, Ueno Y. Autonomic nervous system and the liver. Hepatol Res. 2017;47(2):160–5.
Kandilis AN, Papadopoulou IP, Koskinas J, Sotiropoulos G, Tiniakos DG. Liver innervation and hepatic function: new insights. J Surg Res. 2015;194(2):511–9.
Asrani SK, Devarbhavi H, Eaton J, Kamath PS. Burden of liver diseases in the world. J Hepatol. 2019;70(1):151–71.
Sharma A, Nagalli S: Chronic Liver Disease. In: StatPearls. Treasure Island (FL) ineligible companies. Disclosure: Shivaraj Nagalli declares no relevant financial relationships with ineligible companies.: StatPearls Publishing. 2024.
Santantonio T, Wiegand J, Tilman Gerlach J. Acute hepatitis C: current status and remaining challenges. J Hepatol. 2008;49(4):625–33.
Hoofnagle JH, Longo DL, Björnsson ES. Drug-induced liver injury—types and phenotypes. N Engl J Med. 2019;381(3):264–73.
Sánchez-Oro R, Jara-Díaz AM, Martínez-Sanz G. Fitz-Hugh-Curtis syndrome: a cause of right upper quadrant abdominal pain. Med Clin. 2020;154(11):447–52.
Motta RV, Saffioti F, Mavroeidis VK. Hepatolithiasis: epidemiology, presentation, classification and management of a complex disease. World J Gastroenterol. 2024;30(13):1836–50.
Bianchi G, Marchesini G, Marzocchi R, Pinna AD, Zoli M. Metabolic syndrome in liver transplantation: relation to etiology and immunosuppression. Liver Transpl. 2008;14(11):1648–54.
Laryea M, Watt KD, Molinari M, Walsh MJ, McAlister VC, Marotta PJ, et al. Metabolic syndrome in liver transplant recipients: prevalence and association with major vascular events. Liver Transpl. 2007;13(8):1109–14.
Kjaer M, Jurlander J, Keiding S, Galbo H, Kirkegaard P, Hage E. No reinnervation of hepatic sympathetic nerves after liver transplantation in human subjects. J Hepatol. 1994;20(1):97–100.
Miller BM, Oderberg IM, Goessling W. Hepatic nervous system in development, regeneration, and disease. Hepatology. 2021;74(6):3513–22.
Jensen KJ, Alpini G, Glaser S. Hepatic nervous system and neurobiology of the liver. Compr Physiol. 2013;3(2):655–65.
Robinson MW, Harmon C, O’Farrelly C. Liver immunology and its role in inflammation and homeostasis. Cell Mol Immunol. 2016;13(3):267–76.
Sousa AMM, Meyer KA, Santpere G, Gulden FO, Sestan N. Evolution of the human nervous system function, structure, and development. Cell. 2017;170(2):226–47.
Teratani T, Mikami Y, Kanai T. Neuroimmune crosstalk in the gut and liver. Int Immunol. 2022;34(9):475–84.
Wehrwein EA, Orer HS, Barman SM. Overview of the anatomy, physiology, and pharmacology of the autonomic nervous system. Compr Physiol. 2016;6(3):1239–78.
Pavlov VA, Tracey KJ. The vagus nerve and the inflammatory reflex—linking immunity and metabolism. Nat Rev Endocrinol. 2012;8(12):743–54.
Yi CX, la Fleur SE, Fliers E, Kalsbeek A. The role of the autonomic nervous liver innervation in the control of energy metabolism. Biochim Biophys Acta. 2010;1802(4):416–31.
Tsuneki KIK. Electron microscope study of vertebrate liver innervation. Arch Histol Jpn. 1981;44:1–13.
Kalra A, Yetiskul E, Wehrle CJ, Tuma F: Physiology, Liver. In: StatPearls. Treasure Island (FL) ineligible companies. Disclosure: Ekrem Yetiskul declares no relevant financial relationships with ineligible companies. Disclosure: Chase Wehrle declares no relevant financial relationships with ineligible companies. Disclosure: Faiz Tuma declares no relevant financial relationships with ineligible companies.: StatPearls Publishing Copyright © 2024, StatPearls Publishing LLC., 2024.
Trefts E, Gannon M, Wasserman DH. The liver. Curr Biol. 2017;27(21):R1147-r1151.
Grant DM. Detoxification pathways in the liver. J Inherit Metab Dis. 1991;14(4):421–30.
Kita H, Mackay IR, Van De Water J, Gershwin ME. The lymphoid liver: considerations on pathways to autoimmune injury. Gastroenterology. 2001;120(6):1485–501.
Crispe IN. The liver as a lymphoid organ. Annu Rev Immunol. 2009;27:147–63.
Shuai Z, Leung MW, He X, Zhang W, Yang G, Leung PS, et al. Adaptive immunity in the liver. Cell Mol Immunol. 2016;13(3):354–68.
Abdel-Misih SR, Bloomston M. Liver anatomy. Surg Clin North Am. 2010;90(4):643–53.
Breslin JW, Yang Y, Scallan JP, Sweat RS, Adderley SP, Murfee WL. Lymphatic vessel network structure and physiology. Compr Physiol. 2018;9(1):207–99.
Heymann F, Tacke F. Immunology in the liver—from homeostasis to disease. Nat Rev Gastroenterol Hepatol. 2016;13(2):88–110.
Wang JY, Ma JP, Nie HY, Zhang XJ, Zhang P, She ZG, et al. Hepatic regulator of G protein signaling 5 ameliorates nonalcoholic fatty liver disease by suppressing transforming growth factor beta-activated kinase 1-c-Jun-N-terminal kinase/p38 signaling. Hepatology. 2021;73(1):104–25.
Thomson AW, Knolle PA. Antigen-presenting cell function in the tolerogenic liver environment. Nat Rev Immunol. 2010;10(11):753–66.
Kubes P, Jenne C. Immune responses in the liver. Ann Rev Immunol. 2018;36:247–77.
Guo JL, Li Y, Shan YH, Shu C, Wang F, Wang X, et al. Humanized mice reveal an essential role for human hepatocytes in the development of the liver immune system. Cell Death Dis. 2018. https://doi.org/10.1038/s41419-018-0720-9.
Chung KW, Kim KM, Choi YJ, An HJ, Lee B, Kim DH, et al. The critical role played by endotoxin-induced liver autophagy in the maintenance of lipid metabolism during sepsis. Autophagy. 2017;13(7):1113–29.
Yan J, Li S, Li SL. The role of the liver in sepsis. Int Rev Immunol. 2014;33(6):498–510.
Mao KR, Baptista AP, Tamoutounour S, Zhuang LN, Bouladoux N, Martins AJ, et al. Innate and adaptive lymphocytes sequentially shape the gut microbiota and lipid metabolism. Nature. 2018;554(7691):255.
Megahed FAK, Zhou XL, Sun PN. The interactions between HBV and the innate immunity of hepatocytes. Viruses-Basel. 2020. https://doi.org/10.3390/v12030285.
Davies SP, Terry LV, Wilkinson AL, Stamataki Z. Cell-in-cell structures in the liver: a tale of four E’s. Front Immunol. 2020. https://doi.org/10.3389/fimmu.2020.00650.
Halpern KB, Shenhav R, Massalha H, Toth B, Egozi A, Massasa EE, et al. Paired-cell sequencing enables spatial gene expression mapping of liver endothelial cells. Nat Biotechnol. 2018;36(10):962.
Churchland PS, Winkielman P. Modulating social behavior with oxytocin: how does it work? What does it mean? Horm Behav. 2012;61(3):392–9.
Boone M, Deen PM. Physiology and pathophysiology of the vasopressin-regulated renal water reabsorption. Pflugers Arch. 2008;456(6):1005–24.
Jankowski M, Bissonauth V, Gao L, Gangal M, Wang D, Danalache B, et al. Anti-inflammatory effect of oxytocin in rat myocardial infarction. Basic Res Cardiol. 2010;105(2):205–18.
Oliveira-Pelegrin GR, Saia RS, Carnio EC, Rocha MJ. Oxytocin affects nitric oxide and cytokine production by sepsis-sensitized macrophages. NeuroImmunoModulation. 2013;20(2):65–71.
Poutahidis T, Kearney SM, Levkovich T, Qi P, Varian BJ, Lakritz JR, et al. Microbial symbionts accelerate wound healing via the neuropeptide hormone oxytocin. PLoS ONE. 2013;8(10):e78898.
Boyd JH, Holmes CL, Wang Y, Roberts H, Walley KR. Vasopressin decreases sepsis-induced pulmonary inflammation through the V2R. Resuscitation. 2008;79(2):325–31.
Palin K, Moreau ML, Sauvant J, Orcel H, Nadjar A, Duvoid-Guillou A, et al. Interleukin-6 activates arginine vasopressin neurons in the supraoptic nucleus during immune challenge in rats. Am J Physiol Endocrinol Metab. 2009;296(6):E1289-1299.
Tanriverdi F, Silveira L, MacColl G, Bouloux P. The hypothalamic-pituitary-gonadal axis: immune function and autoimmunity. J Endocrinol. 2003;176:293–304.
Ortona E, Pierdominici M, Rider V. Editorial: sex hormones and gender differences in immune responses. Front Immunol. 2019;10:1076.
Taneja V. Sex hormones determine immune response. Front Immunol. 2018;9:1931.
Orzołek I, Sobieraj J, Domagała-Kulawik J. Estrogens, cancer and immunity. Cancers. 2022. https://doi.org/10.3390/cancers14092265.
Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. 2016;16(10):626–38.
Markle JG, Fish EN. SeXX matters in immunity. Trends Immunol. 2014;35(3):97–104.
Butterworth M, McClellan B, Allansmith M. Influence of sex in immunoglobulin levels. Nature. 1967;214:1224–5.
Von Haam E, Rosenfeld I. The effect of estrone on antibody-production. J Immunol. 1942;43(1):109–17.
Mangalam AK, Taneja V, David CS. HLA class II molecules influence susceptibility versus protection in inflammatory diseases by determining the cytokine profile. J Immunol. 2013;190(2):513–8.
Ngo ST, Steyn FJ, McCombe PA. Gender differences in autoimmune disease. Front Neuroendocrinol. 2014;35(3):347–69.
Vancolen S, Sébire G, Robaire B. Influence of androgens on the innate immune system. Andrology. 2023;11(7):1237–44.
Sellau J, Groneberg M, Lotter H. Androgen-dependent immune modulation in parasitic infection. Semin Immunopathol. 2019;41(2):213–24.
Hreha TN, Collins CA, Cole EB, Jin RJ, Hunstad DA. Androgen exposure impairs neutrophil maturation and function within the infected kidney. mBio. 2024;15(2):e0317023.
Alwani M, Yassin A, Al-Zoubi RM, Aboumarzouk OM, Nettleship J, Kelly D, et al. Sex-based differences in severity and mortality in COVID-19. Rev Med Virol. 2021;31(6):e2223.
Duarte-Silva M, Oliveira CNS, Fuzo C, Silva-Neto PV, Toro DM, Pimentel VE, et al. Divergent androgenic modulation of SARS-CoV-2 infection cooperates with dysregulated immune response to dictate worse COVID-19 outcomes in men. Brain Behav Immun. 2023;114:275–86.
Cook MB. Epidemiology: excess cancer in men–a call for an increased research focus. Nat Rev Clin Oncol. 2013;10(4):186–8.
Terziroli Beretta-Piccoli B, Mieli-Vergani G, Vergani D. Autoimmmune hepatitis. Cell Mol Immunol. 2022;19(2):158–76.
Schwinge D, Schramm C. Sex-related factors in autoimmune liver diseases. Semin Immunopathol. 2019;41(2):165–75.
Pawlikowski M, Stepien H, Komorowski J. Hypothalamic-pituitary-thyroid axis and the immune system. Neuroimmunomodulation. 1994;1:149–52.
Barreiro Arcos ML, Gorelik G, Klecha A, Genaro AM, Cremaschi GA. Thyroid hormones increase inducible nitric oxide synthase gene expression downstream from PKC-zeta in murine tumor T lymphocytes. Am J Physiol Cell Physiol. 2006;291(2):C327-336.
Klecha AJ, Genaro AM, Gorelik G, Barreiro Arcos ML, Silberman DM, Schuman M, et al. Integrative study of hypothalamus-pituitary-thyroid-immune system interaction: thyroid hormone-mediated modulation of lymphocyte activity through the protein kinase C signaling pathway. J Endocrinol. 2006;189(1):45–55.
Tan TL, Rajeswaran H, Haddad S, Shahi A, Parvizi J. Increased risk of periprosthetic joint infections in patients with hypothyroidism undergoing total joint arthroplasty. J Arthroplasty. 2016;31(4):868–71.
Nobrega MM, Auge AP, de Toledo LG, da Silva CS, Frade AB, Salles MJ. Bacteriuria and urinary tract infection after female urodynamic studies: risk factors and microbiological analysis. Am J Infect Control. 2015;43(10):1035–9.
Shoja MM, Oyesiku NM. Clinical anatomy of the cranial nerves. Clin Anat. 2014;27(1):2–3.
Nance DM, Sanders VM. Autonomic innervation and regulation of the immune system (1987–2007). Brain Behav Immun. 2007;21:736–45.
Wang H, Yu M, Ochani M, CA A, Tanovic M, S S, et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation. Nature. 2003;421(6921):384–8.
Kawashima K, Fujii T, Moriwaki Y, Misawa H. Critical roles of acetylcholine and the muscarinic and nicotinic acetylcholine receptors in the regulation of immune function. Life Sci. 2012;91(21–22):1027–32.
Kawashima K, Fujii T, Moriwaki Y, Misawa H, Horiguchi K. Non-neuronal cholinergic system in regulation of immune function with a focus on alpha7 nAChRs. Int Immunopharmacol. 2015;29(1):127–34.
Rosas-Ballina M, Olofsson PS, Ochani M, Valdes-Ferrer SI, Levine YA, Reardon C, et al. Acetylcholine-synthesizing T cells relay neural signals in a vagus nerve circuit. Science. 2011;334(6052):98–101.
Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, et al. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015;523(7560):337–41.
Antila S, Karaman S, Nurmi H, Airavaara M, Voutilainen MH, Mathivet T, et al. Development and plasticity of meningeal lymphatic vessels. J Exp Med. 2017;214(12):3645–67.
Kipnis J. Multifaceted interactions between adaptive immunity and the central nervous system. Science. 2016;353(6301):766–71.
Da Mesquita S, Louveau A, Vaccari A, Smirnov I, Cornelison RC, Kingsmore KM, et al. Functional aspects of meningeal lymphatics in ageing and Alzheimer’s disease. Nature. 2018;560(7717):185–91.
Dantzer R. Neuroimmune interactions: from the brain to the immune system and vice versa. Physiol Rev. 2018;98(1):477–504.
Dantzer R, Konsman JP, Bluthé RM, Kelley KW. Neural and humoral pathways of communication from the immune system to the brain: parallel or convergent? Auton Neurosci. 2000;85:60–5.
Steinberg BE, Silverman HA, Robbiati S, Gunasekaran MK. Cytokine-specific neurograms in the sensory vagus nerve. Bioelectron Med. 2016;3:7–17.
von Banchet GS, Kiehl M, Schaible HG. Acute and long-term effects of IL-6 on cultured dorsal root ganglion neurones from adult rat. J Neurochem. 2005;94(1):238–48.
Jin X, Gereau RWt. Acute p38-mediated modulation of tetrodotoxin-resistant sodium channels in mouse sensory neurons by tumor necrosis factor-alpha. J Neurosci. 2006;26(1):246–55.
Shen KF, Zhu HQ, Wei XH, Wang J, Li YY, Pang RP, et al. Interleukin-10 down-regulates voltage gated sodium channels in rat dorsal root ganglion neurons. Exp Neurol. 2013;247:466–75.
Stemkowski PL, Noh MC, Chen Y, Smith PA. Increased excitability of medium-sized dorsal root ganglion neurons by prolonged interleukin-1beta exposure is K(+) channel dependent and reversible. J Physiol. 2015;593(16):3739–55.
Da Mesquita S, Fu Z, Kipnis J. The meningeal lymphatic system: a new player in neurophysiology. Neuron. 2018;100(2):375–88.
Abraira VE, Ginty DD. The sensory neurons of touch. Neuron. 2013;79(4):618–39.
Reardon C, Murray K, Lomax AE. Neuroimmune communication in health and disease. Physiol Rev. 2018;98(4):2287–316.
Chavan SS, Pavlov VA, Tracey KJ. Mechanisms and therapeutic relevance of neuro-immune communication. Immunity. 2017;46(6):927–42.
Vahl TP, Tauchi M, Durler TS, Elfers EE, Fernandes TM, Bitner RD, et al. Glucagon-like peptide-1 (GLP-1) receptors expressed on nerve terminals in the portal vein mediate the effects of endogenous GLP-1 on glucose tolerance in rats. Endocrinology. 2007;148(10):4965–73.
Balkan B, Li X. Portal GLP-1 administration in rats augments the insulin response to glucose via neuronal mechanisms. Am J Physiol Regul Integr Comp Physiol. 2000;279(4):R1449-1454.
Nakabayashi H. Neural monitoring system for circulating somatostatin in the hepatoportal area. Nutrition Nutrition. 1997;13(3):225–9.
Shiraishi T, Sasaki K, Niijima A, Oomura Y. Leptin effects on feeding-related hypothalamic and peripheral neuronal activities in normal and obese rats. Nutrition. 1999;15(7–8):576–9.
Smith GP, Jerome C, Cushin BJ, Eterno R, Simansky KJ. Abdominal vagotomy blocks the satiety effect of cholecystokinin in the rat. Science. 1981;213(4511):1036–7.
Baral P, Umans BD, Li L, Wallrapp A, Bist M, Kirschbaum T, et al. Nociceptor sensory neurons suppress neutrophil and gammadelta T cell responses in bacterial lung infections and lethal pneumonia. Nat Med. 2018;24(4):417–26.
Pinho-Ribeiro FA, Baddal B, Haarsma R, O’Seaghdha M, Yang NJ, Blake KJ, et al. Blocking neuronal signaling to immune cells treats streptococcal invasive infection. Cell. 2018;173(5):1083–97.
Kashem SW, Riedl MS, Yao C, Honda CN, Vulchanova L, Kaplan DH. Nociceptive sensory fibers drive interleukin-23 production from CD301b+ dermal dendritic cells and drive protective cutaneous immunity. Immunity. 2015;43(3):515–26.
Klein Wolterink RGJ, Wu GS, Chiu IM, Veiga-Fernandes H. Neuroimmune interactions in peripheral organs. Annu Rev Neurosci. 2022;45(1):339–60.
Kihara N, Kihara N, de la Fuente SG, Fujino K, Takahashi T, Pappas TN, et al. Vanilloid receptor-1 containing primary sensory neurones mediate dextran sulphate sodium induced colitis in rats. Gut. 2003;52:713–9.
Engel MA, Khalil M, Siklosi N, Mueller-Tribbensee SM, Neuhuber WL, Neurath MF, et al. Opposite effects of substance P and calcitonin gene-related peptide in oxazolone colitis. Dig Liver Dis. 2012;44(1):24–9.
Gomes RN, Castro-Faria-Neto HC, Bozza PT, Soares MBP, Shoemaker CB, David JR, et al. Calcitonin gene-related peptide inhibits local acute inflammation and protects mice against lethal endotoxemia. Shock. 2005;24(6):590–4.
Fernandes ES, Liang L, Smillie SJ, Kaiser F, Purcell R, Rivett DW, et al. TRPV1 deletion enhances local inflammation and accelerates the onset of systemic inflammatory response syndrome. J Immunol. 2012;188(11):5741–51.
Lechner SG, Markworth S, Poole K, Smith ES, Lapatsina L, Frahm S, et al. The molecular and cellular identity of peripheral osmoreceptors. Neuron. 2011;69(2):332–44.
Nguyen TN, Siddiqui G, Veldhuis NA, Poole DP. Diverse roles of TRPV4 in macrophages: a need for unbiased Profiling. Front Immunol. 2021;12:828115.
Stare J, Siami S, Trudel E, Prager-Khoutorsky M, Sharshar T, Bourque CW. Effects of peritoneal sepsis on rat central osmoregulatory neurons mediating thirst and vasopressin release. J Neurosci. 2015;35(35):12188–97.
Stookey JD, Constant F, Popkin BM, Gardner CD. Drinking water is associated with weight loss in overweight dieting women independent of diet and activity. Obesity. 2008;16(11):2481–8.
Kemp AH, May M, Gueler F, Barg-Hock H, Heiringhoff K-H, Engeli S, et al. Liver afferents contribute to water drinking-induced sympathetic activation in human subjects: a clinical trial. PLoS ONE. 2011. https://doi.org/10.1371/journal.pone.0025898.
Morita H, Ohyama H, Hagiike M, Horiba T, Miyake K, Yamanouchi H, et al. Effects of portal infusion of hypertonic solution on jejunal electrolyte transport in anesthetized dogs. Am J Physiol Renal Physiol. 1990;258:R1289–94.
Morita H, Abe C. Negative feedforward control of body fluid homeostasis by hepatorenal reflex. Hypertens Res. 2011;34(8):895–905.
Umpierre AD, Li B, Ayasoufi K, Simon WL, Zhao S, Xie M, et al. Microglial P2Y6 calcium signaling promotes phagocytosis and shapes neuroimmune responses in epileptogenesis. Neuron. 2024;112(12):1959–1977.e10.
Raybould HE. Sensing of glucose in the gastrointestinal tract. Auton Neurosci. 2007;133(1):86–90.
Watts AG, Donovan CM. Sweet talk in the brain: glucosensing, neural networks, and hypoglycemic counterregulation. Front Neuroendocrinol. 2010;31(1):32–43.
Yamamoto T. Brain mechanisms of sweetness and palatability of sugars. Nutr Rev. 2003;61(5 Pt 2):S5-9.
Niijima A. Afferent impulse discharges from glucoreceptors in the liver of the guinea pig. Ann N Y Acad Sci. 1969;157(2):690–700.
Caspi L, Wang PY, Lam TK. A balance of lipid-sensing mechanisms in the brain and liver. Cell Metab. 2007;6(2):99–104.
Breen DM, Yue JTY, Rasmussen BA, Kokorovic A, Cheung GWC, Lam TKT. Duodenal PKC-δ and cholecystokinin signaling axis regulates glucose production. Diabetes. 2011;60(12):3148–53.
Seillet C, Luong K, Tellier J, Jacquelot N, Shen RD, Hickey P, et al. The neuropeptide VIP confers anticipatory mucosal immunity by regulating ILC3 activity. Nat Immunol. 2020;21(2):168–77.
Wang X, Ota N, Manzanillo P, Kates L, Zavala-Solorio J, Eidenschenk C, et al. Interleukin-22 alleviates metabolic disorders and restores mucosal immunity in diabetes. Nature. 2014;514(7521):237–41.
Date Y, Shimbara T, Koda S, Toshinai K, Ida T, Murakami N, et al. Peripheral ghrelin transmits orexigenic signals through the noradrenergic pathway from the hindbrain to the hypothalamus. Cell Metab. 2006;4(4):323–31.
Lin JC, Peng YJ, Wang SY, Young TH, Salter DM, Lee HS. Role of the sympathetic nervous system in carbon tetrachloride-induced hepatotoxicity and systemic inflammation. PLoS ONE. 2015;10(3):e0121365.
Blum D, Torch S, Lambeng N, Nissou M, Benabid AL, Sadoul R, et al. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: contribution to the apoptotic theory in. Prog Neurogibol. 2001;65:135–72.
Judy WV, Watanabe AM, Murphy WR, Aprison BS, Yu PL. Sympathetic nerve activity and blood pressure in normotensive backcross rats genetically related to the spontaneously. Hypertension. 1979;1:598–604.
Hamasaki K, Nakashima M, Naito S, Akiyama Y, Ohtsuru A, Hamanaka Y, et al. The sympathetic nervous system promotes carbon tetrachloride-induced liver cirrhosis in rats by suppressing apoptosis and enhancing the growth kinetics of regenerating hepatocytes. J Gastroenterol. 2001;36:111–20.
Soeda J, Mouralidarane A, Ray S, Novelli M, Thomas S, Roskams T, et al. The beta-adrenoceptor agonist isoproterenol rescues acetaminophen-injured livers through increasing progenitor numbers by Wnt in mice. Hepatology. 2014;60(3):1023–34.
Randle LE, Sathish JG, Kitteringham NR, Macdonald I, Williams DP, Park BK. alpha(1)-Adrenoceptor antagonists prevent paracetamol-induced hepatotoxicity in mice. Br J Pharmacol. 2008;153(4):820–30.
Hiramoto T, Chida Y, Sonoda J, Yoshihara K, Sudo N, Kubo C. The hepatic vagus nerve attenuates Fas-induced apoptosis in the mouse liver via alpha7 nicotinic acetylcholine receptor. Gastroenterology. 2008;134(7):2122–31.
Li Y, Xu Z, Yu Y, Yuan H, Xu H, Zhu Q, et al. The vagus nerve attenuates fulminant hepatitis by activating the Src kinase in Kuppfer cells. Scand J Immunol. 2014;79(2):105–12.
Steinebrunner N, Mogler C, Vittas S, Hoyler B, Sandig C, Stremmel W, et al. Pharmacologic cholinesterase inhibition improves survival in acetaminophen-induced acute liver failure in the mouse. BMC Gastroenterol. 2014;14(148):1–8.
Lee JA, Ahmed Q, Hines JE, Burt AD. Disappearance of hepatic parenchymal nerves in human liver cirrhosis. Gut. 1992;33(1):87–91.
Scoazec JY, Racine L, Couvelard A, Moreau A, Flejou JF, Bernuau D, et al. Parenchymal innervation of normal and cirrhotic human liver: a light and electron microscopic study using monoclonal antibodies against the neural cell-adhesion molecule. J Histochem Cytochem. 1993;41(6):899–908.
Stoyanova II, Gulubova MV. Peptidergic nerve fibres in the human liver. Acta Histochem. 1998;100(3):245–56.
Matthews PJ, Aziz Q, Facer P, Davis JB, Thompson DG, Anand P. Increased capsaicin receptor TRPV1 nerve fibres in the inflamed human oesophagus. Eur J Gastroenterol Hepatol. 2004;16(9):897–902.
Akiyoshi H, Terada T. Mast cell, myofibroblast and nerve terminal complexes in carbon tetrachloride-induced cirrhotic rat livers. J Hepatol. 1998;29(1):112–9.
Jaskiewicz K, Robson SC, Banach L. Toxic hepatic injury is associated with proliferation of portal nerve fibers. Pathol Res Pract. 1993;189(10):1191–4.
Higashi T, Friedman SL, Hoshida Y. Hepatic stellate cells as key target in liver fibrosis. Adv Drug Deliv Rev. 2017;121:27–42.
Henriksen JH, Møller S, Ring-Larsen H, Christensen NJ. The sympathetic nervous system in liver disease. J Hepatol. 1998;29(2):328–41.
Sigala B, McKee C, Soeda J, Pazienza V, Morgan M, Lin CI, et al. Sympathetic nervous system catecholamines and neuropeptide Y neurotransmitters are upregulated in human NAFLD and modulate the fibrogenic function of hepatic stellate cells. PLoS ONE. 2013;8(9):e72928.
Oben JA, Yang S, Lin H, Ono M, Diehl AM. Norepinephrine and neuropeptide Y promote proliferation and collagen gene expression of hepatic myofibroblastic stellate cells. Biochem Biophys Res Commun. 2003;302(4):685–90.
Sancho-Bru P, Bataller R, Colmenero J, Gasull X, Moreno M, Arroyo V, et al. Norepinephrine induces calcium spikes and proinflammatory actions in human hepatic stellate cells. Am J Physiol Gastrointest Liver Physiol. 2006;291(5):G877-884.
Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Torbenson M, et al. Hepatic fibrogenesis requires sympathetic neurotransmitters. Gut. 2004;53(3):438–45.
Oben JA, Yang S, Lin H, Ono M, Diehl AM. Acetylcholine promotes the proliferation and collagen gene expression of myofibroblastic hepatic stellate cells. Biochem Biophys Res Commun. 2003;300(1):172–7.
Papke RL, Sanberg PR, Shytle RD. Analysis of mecamylamine stereoisomers on human nicotinic receptor subtypes. J Pharmacol Exp Therapeut. 2001;297(2):646–56.
Marchesini G, Brizi M, Bianchi G, Tomassetti S, Bugianesi E, Lenzi M, et al. Nonalcoholic fatty liver disease: a feature of the metabolic syndrome. Diabetes. 2001;50:1844–50.
Duseja A, Chalasani N. Epidemiology and risk factors of nonalcoholic fatty liver disease (NAFLD). Hepatol Int. 2013;7(Suppl 2):755–64.
Wong VW, Ekstedt M, Wong GL, Hagström H. Changing epidemiology, global trends and implications for outcomes of NAFLD. J Hepatol. 2023;79(3):842–52.
Benedict M, Zhang X. Non-alcoholic fatty liver disease: an expanded review. World J Hepatol. 2017;9(16):715–32.
Rinella ME, Lazarus JV, Ratziu V, Francque SM, Sanyal AJ, Kanwal F, et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. J Hepatol. 2023;79(6):1542–56.
George J. Adding to the confusion in more than just the name. Clin Mol Hepatol. 2023;29(4):973–6.
De A, Bhagat N, Mehta M, Taneja S, Duseja A. Metabolic dysfunction-associated steatotic liver disease (MASLD) definition is better than MAFLD criteria for lean patients with NAFLD. J Hepatol. 2024;80(2):e61–2.
Targher G, Byrne CD, Tilg H. MASLD: a systemic metabolic disorder with cardiovascular and malignant complications. Gut. 2024;73(4):691–702.
Hagström H, Vessby J, Ekstedt M, Shang Y. 99% of patients with NAFLD meet MASLD criteria and natural history is therefore identical. J Hepatol. 2024;80(2):e76–7.
Younossi ZM, Paik JM, Stepanova M, Ong J, Alqahtani S, Henry L. Clinical profiles and mortality rates are similar for metabolic dysfunction-associated steatotic liver disease and non-alcoholic fatty liver disease. J Hepatol. 2024;80(5):694–701.
Song SJ, Lai JC, Wong GL, Wong VW, Yip TC. Can we use old NAFLD data under the new MASLD definition? J Hepatol. 2024;80(2):e54–6.
Bai L, Mesgarzadeh S, Ramesh KS, Huey EL, Liu Y, Gray LA, et al. Genetic identification of vagal sensory neurons that control feeding. Cell. 2019;179(5):1129–43.
Licht CM, de Geus EJ, Penninx BW. Dysregulation of the autonomic nervous system predicts the development of the metabolic syndrome. J Clin Endocrinol Metab. 2013;98(6):2484–93.
Licht CM, Vreeburg SA, van Reedt Dortland AK, Giltay EJ, Hoogendijk WJ, DeRijk RH, et al. Increased sympathetic and decreased parasympathetic activity rather than changes in hypothalamic-pituitary-adrenal axis activity is associated with metabolic abnormalities. J Clin Endocrinol Metab. 2010;95(5):2458–66.
Newton JL, Pairman J, Wilton K, Jones DEJ, Day C. Fatigue and autonomic dysfunction in non-alcoholic fatty liver disease. Clin Auton Res. 2009;19(6):319–26.
Sun W, Zhang D, Sun J, Xu B, Sun K, Wang T, et al. Association between non-alcoholic fatty liver disease and autonomic dysfunction in a Chinese population. QJM. 2015;108(8):617–24.
Hurr C, Simonyan H, Morgan DA, Rahmouni K, Young CN. Liver sympathetic denervation reverses obesity-induced hepatic steatosis. J Physiol. 2019;597(17):4565–80.
Tilg H, Moschen AR. Evolution of inflammation in nonalcoholic fatty liver disease: the multiple parallel hits hypothesis. Hepatology. 2010;52(5):1836–46.
Liu K, Yang L, Wang G, Liu J, Zhao X, Wang Y, et al. Metabolic stress drives sympathetic neuropathy within the liver. Cell Metab. 2021;33(3):666–75.
Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391(10127):1301–14.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Bi G, Su M, Li N, Liang Y, Dang S, Xu J, et al. The ZAR1 resistosome is a calcium-permeable channel triggering plant immune signaling. Cell. 2021;184(13):3528–41.
Kuol N, Stojanovska L, Apostolopoulos V, Nurgali K. Role of the nervous system in cancer metastasis. J Exp Clin Cancer Res. 2018;37(1):5.
Harricharran T, Ogunwobi OO. Emergence of neural regulatory mechanisms in carcinogenesis. World J Clin Oncol. 2019;10(8):279–82.
Huan HB, Wen XD, Chen XJ, Wu L, Wu LL, Zhang L, et al. Sympathetic nervous system promotes hepatocarcinogenesis by modulating inflammation through activation of alpha1-adrenergic receptors of Kupffer cells. Brain Behav Immun. 2017;59:118–34.
Li J, Yang XM, Wang YH, Feng MX, Liu XJ, Zhang YL, et al. Monoamine oxidase A suppresses hepatocellular carcinoma metastasis by inhibiting the adrenergic system and its transactivation of EGFR signaling. J Hepatol. 2014;60(6):1225–34.
Fu S, Liu M, Zhu C, Zhang H, Zhao C, Xie Y, et al. Regulatory MAIT cells controlled by β1 adrenergic receptor signaling contribute to hepatocellular carcinoma progression. Hepatology. 2023. https://doi.org/10.1097/hep.0000000000000014.
Wijdicks EF. Hepatic encephalopathy. N Engl J Med. 2016;375(17):1660–1670.
Ohikere K, Wong RJ. Hepatic encephalopathy: clinical manifestations. Clin Liver Dis. 2024;28(2):253–63.
Lu K. Cellular pathogenesis of hepatic encephalopathy: an update. Biomolecules. 2023. https://doi.org/10.3390/biom13020396.
Bajaj JS, Hylemon PB, Ridlon JM, Heuman DM, Daita K, White MB, et al. Colonic mucosal microbiome differs from stool microbiome in cirrhosis and hepatic encephalopathy and is linked to cognition and inflammation. Am J Physiol Gastrointest Liver Physiol. 2012;303(6):G675-685.
Won SM, Oh KK, Gupta H, Ganesan R, Sharma SP, Jeong JJ, et al. The link between gut microbiota and hepatic encephalopathy. Int J Mol Sci. 2022. https://doi.org/10.3390/ijms23168999.
Bajaj JS, Ridlon JM, Hylemon PB, Thacker LR, Heuman DM, Smith S, et al. Linkage of gut microbiome with cognition in hepatic encephalopathy. Am J Physiol Gastrointest Liver Physiol. 2012;302(1):G168-175.
Albillos A, de Gottardi A, Rescigno M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol. 2020;72(3):558–77.
Zhu R, Liu L, Zhang G, Dong J, Ren Z, Li Z. The pathogenesis of gut microbiota in hepatic encephalopathy by the gut-liver-brain axis. 2023. Biosci Rep. https://doi.org/10.1042/BSR20222524.
Tranah TH, Vijay GK, Ryan JM, Shawcross DL. Systemic inflammation and ammonia in hepatic encephalopathy. Metab Brain Dis. 2013;28(1):1–5.
Görg B, Karababa A, Schütz E, Paluschinski M, Schrimpf A, Shafigullina A, et al. O-GlcNAcylation-dependent upregulation of HO1 triggers ammonia-induced oxidative stress and senescence in hepatic encephalopathy. J Hepatol. 2019;71(5):930–41.
Görg B, Karababa A, Shafigullina A, Bidmon HJ, Häussinger D. Ammonia-induced senescence in cultured rat astrocytes and in human cerebral cortex in hepatic encephalopathy. Glia. 2015;63(1):37–50.
Zemtsova I, Görg B, Keitel V, Bidmon HJ, Schrör K, Häussinger D. Microglia activation in hepatic encephalopathy in rats and humans. Hepatology. 2011;54(1):204–15.
Needham BD, Kaddurah-Daouk R, Mazmanian SK. Gut microbial molecules in behavioural and neurodegenerative conditions. Nat Rev Neurosci. 2020;21(12):717–31.
Teratani T, Mikami Y, Nakamoto N, Suzuki T, Harada Y, Okabayashi K, et al. The liver–brain–gut neural arc maintains the Treg cell niche in the gut. Nature. 2020;585(7826):591–6.
Swain MG. Fatigue in liver disease: pathophysiology and clinical management. Can J Gastroenterol. 2006;20(3):181–8.
Swain MG, Jones DEJ. Fatigue in chronic liver disease: new insights and therapeutic approaches. Liver Int. 2019;39(1):6–19.
Gerber LH, Weinstein AA, Mehta R, Younossi ZM. Importance of fatigue and its measurement in chronic liver disease. World J Gastroenterol. 2019;25(28):3669–83.
Weinstein AA, Diao G, Baghi H, Escheik C, Gerber LH, Younossi ZM. Demonstration of two types of fatigue in subjects with chronic liver disease using factor analysis. Qual Life Res. 2017;26(7):1777–84.
Cauch-Dudek K, Abbey S, Stewart DE, Heathcote EJ. Fatigue in primary biliary cirrhosis. Gut. 1998;43(5):705–10.
Huet PM, Deslauriers J, Tran A, Faucher C, Charbonneau J. Impact of fatigue on the quality of life of patients with primary biliary cirrhosis. Am J Gastroenterol. 2000;95(3):760–7.
Jopson L, Jones DE. Fatigue in primary biliary cirrhosis: prevalence, pathogenesis and management. Dig Dis. 2015;33(Suppl 2):109–14.
Swain MG, Maric M. Defective corticotropin-releasing hormone mediated neuroendocrine and behavioral responses in cholestatic rats: implications for cholestatic liver disease-related sickness behaviors. Hepatology. 1995;22(5):1560–4.
Burak KW, Le T, Swain MG. Increased sensitivity to the locomotor-activating effects of corticotropin-releasing hormone in cholestatic rats. Gastroenterology. 2002;122(3):681–8.
Celik T, Gören MZ, Cinar K, Gürdal H, Onder FO, Tan A, et al. Fatigue of cholestasis and the serotoninergic neurotransmitter system in the rat. Hepatology. 2005;41(4):731–7.
Swain MG, Maric M. Improvement in cholestasis-associated fatigue with a serotonin receptor agonist using a novel rat model of fatigue assessment. Hepatology. 1997;25(2):291–4.
Bhandari K, Kapoor D. Fatigue in cirrhosis. J Clin Exp Hepatol. 2022;12(2):617–24.
Mosher VAL, Swain MG, Pang JXQ, Kaplan GG, Sharkey KA, MacQueen GM, et al. Primary biliary cholangitis alters functional connections of the brain’s deep gray matter. Clin Transl Gastroenterol. 2017;8(7):e107.
Forton DM, Patel N, Prince M, Oatridge A, Hamilton G, Goldblatt J, et al. Fatigue and primary biliary cirrhosis: association of globus pallidus magnetisation transfer ratio measurements with fatigue severity and blood manganese levels. Gut. 2004;53(4):587–92.
Kobayashi Y. The regulatory role of nitric oxide in proinflammatory cytokine expression during the induction and resolution of inflammation. J Leukoc Biol. 2010;88(6):1157–62.
D’Mello C, Le T, Swain MG. Cerebral microglia recruit monocytes into the brain in response to tumor necrosis factoralpha signaling during peripheral organ inflammation. J Neurosci. 2009;29(7):2089–102.
Socodato R, Henriques JF, Portugal CC, Almeida TO, Tedim-Moreira J, Alves RL, et al. Daily alcohol intake triggers aberrant synaptic pruning leading to synapse loss and anxiety-like behavior. Sci Signal. 2020. https://doi.org/10.1126/scisignal.aba5754.
Newton JL, Davidson A, Kerr S, Bhala N, Pairman J, Burt J, et al. Autonomic dysfunction in primary biliary cirrhosis correlates with fatigue severity. Eur J Gastroenterol Hepatol. 2007;19(2):125–32.
Griffiths L, Jones DE. Pathogenesis of primary biliary cirrhosis and its fatigue. Dig Dis. 2014;32(5):615–25.
Newton JL, Gibson GJ, Tomlinson M, Wilton K, Jones D. Fatigue in primary biliary cirrhosis is associated with excessive daytime somnolence. Hepatology. 2006;44(1):91–8.
Abbas G, Jorgensen RA, Lindor KD. Fatigue in primary biliary cirrhosis. Nat Rev Gastroenterol Hepatol. 2010;7(6):313–9.
Goldblatt J, James OFW, Jones DEJ. Grip strength and subjective fatigue in patients with primary biliary cirrhosis. JAMA. 2001;285(17):2196–7.
Blackburn P, Freeston M, Baker CR, Jones DE, Newton JL. The role of psychological factors in the fatigue of primary biliary cirrhosis. Liver Int. 2007;27(5):654–61.
Kim HP, Lieber SR, Rogers ME, Moon AM, Loiselle M, Walker J, et al. A systematic review of patient-reported outcomes in primary biliary cholangitis and primary sclerosing cholangitis. Hepatol Commun. 2020;4(10):1502–15.
Machado MO, Kang NC, Tai F, Sambhi RDS, Berk M, Carvalho AF, et al. Measuring fatigue: a meta-review. Int J Dermatol. 2021;60(9):1053–69.
Lynch EN, Campani C, Innocenti T, Dragoni G, Biagini MR, Forte P, et al. Understanding fatigue in primary biliary cholangitis: from pathophysiology to treatment perspectives. World J Hepatol. 2022;14(6):1111–9.
Mullish BH, Kabir MS, Thursz MR, Dhar A. Review article: depression and the use of antidepressants in patients with chronic liver disease or liver transplantation. Aliment Pharmacol Ther. 2014;40(8):880–92.
Golden J, O’Dwyer AM, Conroy RM. Depression and anxiety in patients with hepatitis C: prevalence, detection rates and risk factors. Gen Hosp Psychiatry. 2005;27(6):431–8.
Huckans M, Fuller BE, Olavarria H, Sasaki AW, Chang M, Flora KD, et al. Multi-analyte profile analysis of plasma immune proteins: altered expression of peripheral immune factors is associated with neuropsychiatric symptom severity in adults with and without chronic hepatitis C virus infection. Brain Behav. 2014;4(2):123–42.
Nadeau S, Rivest S. Effects of circulating tumor necrosis factor on the neuronal activity and expression of the genes encoding the tumor necrosis factor receptors (p55 and p75) in the rat brain: a view from the blood-brain barrier. Neuroscience. 1999;93(4):1449–64.
Wilkinson J, Radkowski M, Eschbacher JM, Laskus T. Activation of brain macrophages/microglia cells in hepatitis C infection. Gut. 2010;59(10):1394–400.
Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9(1):46–56.
Fletcher NF, Wilson GK, Murray J, Hu K, Lewis A, Reynolds GM, et al. Hepatitis C virus infects the endothelial cells of the blood-brain barrier. Gastroenterology. 2012;142(3):634-643.e636.
Grover VP, Pavese N, Koh SB, Wylezinska M, Saxby BK, Gerhard A, et al. Cerebral microglial activation in patients with hepatitis C: in vivo evidence of neuroinflammation. J Viral Hepat. 2012;19(2):e89-96.
Paulino AD, Ubhi K, Rockenstein E, Adame A, Crews L, Letendre S, et al. Neurotoxic effects of the HCV core protein are mediated by sustained activation of ERK via TLR2 signaling. J Neurovirol. 2011;17(4):327–40.
Huang HY, Yu RL, Tsai WF, Chuang WL, Huang JF, Dai CY, et al. Impact of interleukin-1β single nucleotide polymorphisms and depressive symptoms in individuals with chronic viral hepatitis. Kaohsiung J Med Sci. 2024;40(1):94–104.
Lai JY, Ho JX, Kow ASF, Liang G, Tham CL, Ho YC, et al. Interferon therapy and its association with depressive disorders—a review. Front Immunol. 2023;14:1048592.
Chiu WC, Su YP, Su KP, Chen PC. Recurrence of depressive disorders after interferon-induced depression. Transl Psychiatry. 2017;7(2):e1026.
Hoyo-Becerra C, Schlaak JF, Hermann DM. Insights from interferon-α-related depression for the pathogenesis of depression associated with inflammation. Brain Behav Immun. 2014;42:222–31.
Carbone M, Neuberger JM. Autoimmune liver disease, autoimmunity and liver transplantation. J Hepatol. 2014;60(1):210–23.
Schramm C, Wahl I, Weiler-Normann C, Voigt K, Wiegard C, Glaubke C, et al. Health-related quality of life, depression, and anxiety in patients with autoimmune hepatitis. J Hepatol. 2014;60(3):618–24.
Strasser B, Gostner JM, Fuchs D. Mood, food, and cognition: role of tryptophan and serotonin. Curr Opin Clin Nutr Metab Care. 2016;19(1):55–61.
Kraus C, Castrén E, Kasper S, Lanzenberger R. Serotonin and neuroplasticity—links between molecular, functional and structural pathophysiology in depression. Neurosci Biobehav Rev. 2017;77:317–26.
Grace AA. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat Rev Neurosci. 2016;17(8):524–32.
Patten SB. Exogenous corticosteroids and major depression in the general population. J Psychosom Res. 2000;49(6):447–9.
Murphy BE. Steroids and depression. J Steroid Biochem Mol Biol. 1991;38(5):537–59.
Parrish WR, Rosas-Ballina M, Gallowitsch-Puerta M, Ochani M, Ochani K, Yang LH, et al. Modulation of TNF release by choline requires alpha7 subunit nicotinic acetylcholine receptor-mediated signaling. Mol Med. 2008;14(9–10):567–74.
Pavlov VA, Ochani M, Yang LH, Gallowitsch-Puerta M, Ochani K, Lin X, et al. Selective alpha7-nicotinic acetylcholine receptor agonist GTS-21 improves survival in murine endotoxemia and severe sepsis. Crit Care Med. 2007;35(4):1139–44.
Norman GJ, Morris JS, Karelina K, Weil ZM, Zhang N, Al-Abed Y, et al. Cardiopulmonary arrest and resuscitation disrupts cholinergic anti-inflammatory processes: a role for cholinergic alpha7 nicotinic receptors. J Neurosci. 2011;31(9):3446–52.
Terrando N, Yang T, Ryu JK, Newton PT, Monaco C, Feldmann M, et al. Stimulation of the α7 nicotinic acetylcholine receptor protects against neuroinflammation after tibia fracture and endotoxemia in mice. Mol Med. 2014;20(1):667–75.
Hoover DB. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol Ther. 2017;179:1–16.
Koopman FA, Chavan SS, Miljko S, Grazio S, Sokolovic S, Schuurman PR, et al. Vagus nerve stimulation inhibits cytokine production and attenuates disease severity in rheumatoid arthritis. Proc Natl Acad Sci USA. 2016;113(29):8284–9.
Sinniger V, Pellissier S, Fauvelle F, Trocme C, Hoffmann D, Vercueil L, et al. A 12-month pilot study outcomes of vagus nerve stimulation in Crohn’s disease. Neurogastroenterol Motil. 2020;32(10):e13911.
Genovese M, Gaylis N, Sikes D, Kivitz A, Horowitz D, Peterfy C, et al. Safety and efficacy of neurostimulation with a miniaturised vagus nerve stimulation device in patients with multidrugrefractory rheumatoid arthritis: a two-stage multicentre, randomised pilot study. Lancet Rheumatol. 2020;2:e527–38.
Andersson U, Tracey KJ. A new approach to rheumatoid arthritis: treating inflammation with computerized nerve stimulation. Cerebrum. 2012;2012:3.
Hoover DB. Cholinergic modulation of the immune system presents new approaches for treating inflammation. Pharmacol Ther. 2017;179:1–16.
Peng S, Dhawan V, Eidelberg D, Ma Y. Neuroimaging evaluation of deep brain stimulation in the treatment of representative neurodegenerative and neuropsychiatric disorders. Bioelectron Med. 2021;7(1):4.
Pavlov VA, Tracey KJ. Bioelectronic medicine: preclinical insights and clinical advances. Neuron. 2022;110(21):3627–44.
Ma L, Zhang J, Fujita Y, Shinno-Hashimoto H, Shan J, Wan X, et al. Effects of spleen nerve denervation on depression-like phenotype, systemic inflammation, and abnormal composition of gut microbiota in mice after administration of lipopolysaccharide: a role of brain-spleen axis. J Affect Disord. 2022;317:156–65.
Rosas-Ballina M, Ochani M, Parrish WR, Ochani K, Harris YT, Huston JM, et al. Splenic nerve is required for cholinergic antiinflammatory pathway control of TNF in endotoxemia. Proc Natl Acad Sci USA. 2008;105(31):11008–13.
Huston JM, Ochani M, Rosas-Ballina M, Liao H, Ochani K, Pavlov VA, et al. Splenectomy inactivates the cholinergic antiinflammatory pathway during lethal endotoxemia and polymicrobial sepsis. J Exp Med. 2006;203(7):1623–8.
Guyot M, Simon T, Panzolini C, Ceppo F, Daoudlarian D, Murris E, et al. Apical splenic nerve electrical stimulation discloses an anti-inflammatory pathway relying on adrenergic and nicotinic receptors in myeloid cells. Brain Behav Immun. 2019;80:238–46.
Zachs DP, Offutt SJ, Graham RS, Kim Y, Mueller J, Auger JL, et al. Noninvasive ultrasound stimulation of the spleen to treat inflammatory arthritis. Nat Commun. 2019;10(1):951.
Yang B, Ren Q, Zhang JC, Chen QX, Hashimoto K. Altered expression of BDNF, BDNF pro-peptide and their precursor proBDNF in brain and liver tissues from psychiatric disorders: rethinking the brain-liver axis. Transl Psychiatry. 2017;7(5):e1128.
Acknowledgements
We would like to thank Editage for English language editing.
Funding
This manuscript was supported by grants from the National Natural Sciences Foundation of China (82070613 and 82370638 to R.C.), the Science and Technology Innovation Program of Hunan Province (2022RC1212 to R.C.), and the Natural Science Foundation of Hunan Province China (2023JJ10095 to R.C.).
Author information
Authors and Affiliations
Contributions
Ruochan Chen: conceptualization, investigation, and writing (original draft, review & editing). Ju Zou: writing and graphics (original draft, review and editing). Jie Li, Xiaoxu Wang, and Daoling Tang: writing (original draft, review and editing).
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Zou, J., Li, J., Wang, X. et al. Neuroimmune modulation in liver pathophysiology. J Neuroinflammation 21, 188 (2024). https://doi.org/10.1186/s12974-024-03181-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12974-024-03181-w