
Abstract
The Bacillus Calmette-Guérin (BCG) vaccine was discovered a century ago and has since been clinically applicable. BCG can not only be used for the prevention of tuberculosis, but also has a non-specific protective effect on the human body called trained immunity that is mediated by innate immune cells such as monocytes, macrophages, and natural killer cells. Mechanisms of trained immunity include epigenetic reprogramming, metabolic reprogramming, and long-term protection mediated by hematopoietic stem cells. Trained immunity has so far shown beneficial effects on cancer, viral-infections, autoimmune diseases, and a variety of other diseases, especially bladder cancer, respiratory viruses, and type 1 diabetes. The modulation of the immune response by BCG has led to the development of a variety of recombinant vaccines. Although the specific mechanism of BCG prevention on diseases has not been fully clarified, the potential role of BCG deserves further exploration, which is of great significance for prevention and treatment of diseases.
Similar content being viewed by others
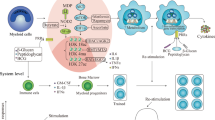
Bacillus Calmette-Guérin (BCG) vaccine is an attenuated strain of Mycobacterium bovis obtained by serial passage. Mycobacterium bovis was firstly isolated in 1908 by Albert Calmette and Camille Guéri from a glycerol bile potato medium at the Pasteur Institute in Lille [1]. From 1908 to 1921, they serially passaged the strain and obtained a low-virulence strain and finally found that the strain protects the body from attack by the virulent Mycobacterium tuberculosis, and named it as BCG [1]. Trained immunity is the long-term functional reprogramming of innate immune cells, which is evoked by exogenous or endogenous insults, and leads to an increased effector function upon secondary stimulation after returning to an inactive state [2]. Compared to classical immunological memory, trained immunity has a number of characteristics. First, cells (myeloid cells, natural killer cells) and germline encoded recognition and effector molecules (e.g., pattern recognition receptors, cytokines) that differ from classical immunological memory are involved. Second, the increased responsiveness to secondary stimuli during trained immunity is not specific for a particular pathogen. Finally, trained immunity relies on changes in the functional state of innate immune cells that persists for weeks to months, rather than years, after the elimination of the initial stimulus [3]. In this review, we summarized the history, mechanisms, and potential applications of BCG-induced trained immunity.
History of BCG-induced trained immunity
BCG was first administered as a vaccine on 18 July 1921 at the Charité Hospital in Paris to an infant boy whose mother had died of tuberculosis [4]. Mass production of BCG began in 1924, and with the widespread vaccination of BCG through intradermal injection, its role was found to be more than just the prevention of tuberculosis. In 1928, Pearl found in an autopsy study that the incidence of cancer in tuberculosis patients was low [5], and subsequent epidemiological studies also showed that BCG could prevent child mortality independent of its effect on tuberculosis [6,7,8]. This sparked great interest and presented a new approach to studying the role of BCG in other diseases (Fig. 1).

BCG and the history of trained immunity
The study by MacKaness GB et al. in 1964 showed that infection with certain bacterial pathogens would confer a high degree of resistance in the host against other unrelated pathogens, the host had nonspecific protection ("cross-protection"). It has been shown that BCG can induce host resistance to other infections [9, 10]. As further evidence for BCG induced non-specific protection, studies in the following years have also shown that BCG-inoculated mice can prevent infections such as Plasmodium [11], Schistosoma manson [12]. In 1988, Bistoni et al. elicited substantial protection against infection by Candida albicans vaccination in athymic mice, showing that cytotoxic T cells and B lymphocytes do not play a key role in the protection against C. albicans infections [13]. This protective effect, is independent of T/B cells, suggesting that BCG may exert a non-specific protective effect through a mechanism independent of adaptive immunity. In 2003, Garly et al. showed that BCG vaccination in West African children could reduce the morbidity caused by infections other than tuberculosis, thereby reducing the overall mortality, which profits from the non-specific protection of BCG vaccination [14]. There is a strong argument for BCG to induce non-specific protection against other infections, but the mechanism still requires further investigation.
Until 2011, studies had found that these nonspecific protective effects were mediated by innate immune cells such as monocytes (Mo), macrophages (Mφ), natural killer cells (NK), dendritic cells (DC), and neutrophils. Netea MG et al. showed that innate immunity conferred immune memory to innate host defenses. The feature is called “trained immunity” [3, 15]. A 2012 study, which combined in vivo and in vitro experiments, demonstrated that a NOD2-mediated epigenetic change at the level of histone methylation (H3K4me3) is the mechanism through which BCG enhances innate immune responses [16]. Saeed et al. in 2014 demonstrated the importance of epigenetic regulation in monocyte-to-macrophage differentiation and trained immune pathways [17]. At the same time, Cheng et al. proved that the shift of metabolic pathways to oxygen glycolysis is critical for the maintenance of trained immunity [18]. Regarding the relationship between epigenetics and metabolism-induced trained immunity, the study in 2016 showed that cellular metabolic reprogramming is a central process of BCG-induced trained immunity; metabolism and epigenetic modification are interwoven; and positive feedback loops may enhance trained immunophenotypes [19]. In view of the problem that mature innate immune cells (e.g., monocytes) have a short lifetime in circulation compared to the duration of trained immunity [20], a study in 2018 showed that the introduction of BCG into bone marrow (BM) alters the transcriptional patterns of hematopoietic stem cells (HSCs) and multifunctional progenitor cells (MPPs), and the monocytes/macrophages trained by BCG-induced HSC reprogramming are sustainable in vivo and provides better protection [21], which further extends the trained immune mechanism to the level of hematopoietic progenitors.
Mechanisms of trained immunity
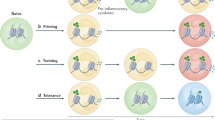
Epigenetic reprograming is one of the molecular mechanisms that induces the development of trained immunity [22]. The different types of epigenetic modifications include DNA modifications, noncoding RNAs, histone modifications, and chromatin remodeling [23]. In addition to epigenetic reprogramming, different cellular metabolic pathways are also involved in the regulation and development of monocytes, macrophages, and NK cell-trained immunity. Epigenetic modification is a method of controlling gene expression that requires the coordination of cellular metabolism. Furthermore, epigenetic reprogramming is regulated by changes in immune cell metabolic flux. Trained immunity can play a long-term protective effect against infection, which is the result of interaction with hematopoietic stem cells [21] (Fig. 2).

BCG induces trained immunity by binding to NOD2 receptors. The Akt/mTOR pathway is then activated for the metabolic switch to glycolysis. BCG induces epigenetic and metabolic reprogramming of innate immune cells to train innate immunity. Epigenetic reprogramming and metabolic reprogramming influence each other. Through the interaction with hematopoietic stem cells, it plays a long-term protective effect against infection. When the body was stimulated by BCG again, the cells after the training immunization produced more pro-inflammatory factors
Epigenetic reprogramming
BCG induces trained immunity by binding to NOD2 receptors, and epigenetic recombination is the key to regulating gene expression to induce trained immunity [16]. Epigenetic mechanisms mainly include DNA methylation, post-translational modifications of histones, and noncoding RNA regulation. DNA methylation normally hinders transcription, whereas histone modifications can have more complex effects [24].
Epigenetic reprogramming of monocytes, characterized by deposition of chromatin marks and altered DNA methylation status, promotes the expression of pro-inflammatory genes, and metabolic reorganization underlies the long-term alterations in immune responses [19]. BCG vaccination induces histone modifications and epigenetic reprogramming in human monocytes at the promoter sites of genes encoding inflammatory cytokines such as TNF-α and IL-6 [16]. Trained monocytes and macrophages exhibited functional and epigenetic reprogramming, resulting in increased production of proinflammatory cytokines IL-6, IL-1, and TNF-α and chemokines, and enhanced phagocytosis and mortality [25]. At the same time, BCG also promotes NK cells to produce pro-inflammatory cytokines such as IL-1β and IL-6 [26]. Pro-inflammatory cytokines produced by cells, such as TNF-α, IL-1β and IL-6 coordinate local and systemic inflammatory responses. TNF-α and IL-1β sequentially activate the local endothelium, induce vasodilation, increase vascular permeability, and enable the recruitment of serum proteins and leukocytes to the site of infection. In addition, IL-1β, together with IL-6, activates hepatocytes to produce acute phase proteins. These proteins activate the complement and cause phagocytosis of pathogens by macrophages and neutrophils [27]. In addition, increased release of tumor necrosis TNF-α and IL-6 may prevent tuberculosis and viral infections [21, 28]. The reduction in viremia was highly correlated with the upregulation of IL-1β [28]. Enhanced neutrophil function persists for at least 3 months and is associated with genome-wide epigenetic modification of histone 3 trimethylation of lysine 4 (H3K4me3) [29]. Macrophages are trained to increase the expressions of various pattern recognition receptors (TLR4, CD206, and CD14); chemokine receptors (CCR2 and CXCR4); and costimulatory and/or signaling molecules (CD43, CD14, CD40) that correlated with chromatin remodeling marker H3K4me3 [30]. These receptors facilitate the stimulation of T cells, angiogenesis, and wound healing. As major players in the innate immune-triggered inflammatory response, inflammatory cytokines and chemokines play a key role in host defense against microbial infection. Inflammatory cytokines and chemokines play a crucial role in host defense against microbial infection [27, 31].
In addition to innate immune cells in local tissues, trained immunity has also been reported to be induced in myeloid progenitors of the BM, resulting in monocytes with higher immune potential and longer duration [21]. A recent study further demonstrated that BCG vaccination induces trained immunity through transcriptomic, epigenomic, and functional reprogramming of hematopoietic stem cells, progenitors and monocytes [32]. BCG may alter the BM microenvironment, leading to the production of cytokines via pathogen-associated molecular patterns (PAMPs), which may indirectly affect the function of hematopoietic stem and progenitor cells [21]. BCG can train innate immune cells to produce more cytokines, of which IL-1β has a strong effect on myelopoiesis [33]. IL-1β may be an endogenous mediator which links peripheral activation of monocytes and macrophages by BCG to long-term functional reprogramming at the level of myeloid progenitors.
Metabolic reprogramming
Glycolysis, oxidative phosphorylation, and glutamine catabolism pathways were up-regulated in peripheral blood mononuclear cells after BCG inoculation [19]. Increased glycolytic metabolism in monocytes leads to a shift in the cellular metabolic program from oxidative phosphorylation to aerobic glycolysis (Warburg effect) [18]. Arts et al. [19] showed that BCG-induced trained immunity in monocytes was accompanied by a metabolic shift towards glycolysis through activation of the Akt/mTOR pathway. Metabolites not only provide substrates for biosynthesis but also regulate immune responses through signaling pathways and gene expression [34]. Most metabolites are important substrates and cofactors for epigenetic enzymatic activity.
Inhibited metabolism can reverse epigenetic changes in an in vitro trained immune model, resulting in reduced cytokine responses upon re-stimulation [35]. Healthy individuals taking metformin, an inhibitor of the mammalian target of rapamycin-dependent glycolysis, have reduced immune responses after training, through decreased histone H3 lysine 9 trimethyl marks (H3K9me3) [19].
Potential applications
Prevention and treatment of tuberculosis
The BCG has been used for prevention and treatment of tuberculosis for 100 years, but the effect of BCG varies from person to person. For children, BCG is mainly used to prevent disseminated tuberculosis [36]. The test conducted in Uganda from 1962 to 1970 showed that BCG had a major and sustained protective effect on the early childhood tuberculosis leprosy in this area [37]. In recent years, it has been found that after BCG is given to infant or school-age children after strict tuberculin testing, the lung tuberculosis can be prevented, and the protective effect of meningeal tuberculosis and millet tuberculosis seems to be greater than the protection of lung tuberculosis [38]. Therefore, early vaccination with BCG is beneficial to reducing the incidence of these diseases.
Cancers therapy
BCG can also play a role against cancers. In 1959, Old LJ. et al. proved that the BCG had anti-tumor effect [39], and slowed the progress of the tumor by inducing immune stimulation effects [40]. In 1976, Morales A. et al. firstly used BCG to treat superficial bladder cancer [41]. Until now, BCG therapy is still the standard therapy for non-muscle invasive bladder cancer [42]. Although the study found that BCG treatment on the bladder cancer caused the poor sleep quality of patients with non-muscle invasive bladder cancer, effectiveness of BCG on bladder cancer is positive [43]. It is found that trained immunity level and antitumor effect may be improved by modifying BCG to express a higher level of key PAMP molecules [44]. This further proves the efficacy of BCG in the treatment of bladder cancer. In addition, BCG can also reduce the risk of melanoma [45], but also had used to the treatment of patients with melanoma in III stage [46]. The recent study showed that BCG can induce gastric cancer cell apoptosis and autophagy, activate lymphocytes, enhance anti-tumor activity of immune cells [47]. Moreover, the early BCG vaccination of children is associated with the risk of reducing lung cancer [48] and leukemia [49], and the earlier BCG vaccination is likely to reduce tumor mortality [50]. Another study showed that BCG may have an efficacy against kidney cancer and prostate cancer [51]. It can be seen that exploring the mechanism of the role of BCG is significant to the treatment of cancer.
Antiviral effects
BCG can play an antiviral effect on a variety of viruses. For example, BCG-induced genome-wide epigenetic reprogramming in monocytes are proved to protect humans from experimental infections from yellow-fever attenuated vaccine strains, which may be critical for monocyte-produced IL-1β function to achieve a protective effect [28]. Since the body immunity of the elderly has different degrees of decline as the young, the elderly is easy to be infected by viruses, then the inoculation of BCG may be an effective prevention choice. The study has found that inoculating the elderly with BCG for 3 consecutive months, once a month, can significantly prevent their acute respiratory infections [52]. A mouse experimental study showed that BCG can prevent infections of various DNA and RNA viruses, including herpes and influenza viruses [53]. Moreover, in a randomized, placebo-controlled study, it is found that BCG vaccination before influenza vaccination can significantly improve the reaction amplitude and accelerated induction of the antibody against 2009 epidemic influenza A(H1N1), and the inoculation of BCG can also regulate the influence of the influenza vaccine on the production ability of cytokines [54]. The appearance of this phenomenon may be because BCG affects the humoral and cellular response to influenza vaccine. When influenza vaccine is administered, BCG in the body plays a certain strengthening role, which acts as an adjuvant and enhances the immune response of the body to influenza. Furthermore, BCG can also enhance the heterologous reactions against tetanus toxoid and poliovirus vaccines [55]. In a recent study, the recombinant BCG vaccine was constructed by inserting BZLF1 and LMP2 genes of EB viruses, rBCG vaccine expressing two genes exhibits an obvious effective immunosuppression for tumors of positive EB virus [56], which provides a good idea for continuing development of more effective antiviral vaccines.
For children’s common and flat warts, the local immunotherapy of BCG may be a new, effective, safe choice [57]. Nevertheless, different types of warts have different responses to BCG. A double-blind, randomized control study showed that intradermal injection of BCG is more effective in treating viral warts to a certain extent [58]. In addition, early BCG vaccination may be beneficial to relieve clinical syndrome and its long-term process of warts [59]. In summary, the time, the way and the dose of BCG vaccination have different effects on different diseases. Therefore, it is necessary to clarify the specific mechanism of BCG to prevent diseases.
In recent years, BCG has been found to have a huge potential against coronavirus Disease 2019 (COVID-19). Trained immunity induced by BCG vaccination during birth may have resistance to COVID-19 [60, 61]. First, when people are exposed to pathogens or pathogen components, they can strengthen BCG-induced trained immunity at birth, which is similar to re-vaccination, thus playing a role in resistance. Second, its resistance may be achieved through the inhibition of viral replication by BCG, resulting in a reduction in viral load, followed by a reduction in inflammation and symptoms. However, the protective effect of BCG was not found in another analysis [62]. In other words, although the people were inoculated with BCG vaccine, the analysis results may be different due to different characteristics of the population. Then BCG is insufficient to prevent COVID-19. Hence, the role of BCG on COVID-19 has no clear evidence, and further researches are needed to conduct.
Treating autoimmune diseases
BCG may also have huge potential against autoimmune diseases. Studies have found that BCG inoculation is related to the reduction of hyperglycemia in patients with advanced type 1 diabetes [63]. The decrease of blood sugar is the joint action of the decrease in glucose production and the increase in glucose consumption. BCG vaccination can stabilize or even reduce the level of glycosylated hemoglobin, and induced TNF-α accelerate the death of islet self-reactive T-cells. In addition, the systematic transformation of glucose metabolism from oxidative phosphorylation to aerobic glycolysis showed a high utilization rate of glucose. Then the combined effect of the above mechanisms can achieve the goal of lowering blood sugar levels. The observations of autoimmune diabetic animal models are consistent with that since BCG has an immunomodulatory effect, perhaps it is connected with the decrease of positive autoantibodes of GAD65 and IA-2 in patients of South Indian diabetes [64]. In addition, in a mouse experiment, BCG infection can inhibit the development of experimental autoimmune encephalomyelitis [65]. BCG also can reduce the advancement of multiple sclerosis and retard the progress of brain lesions [66]. The above-mentioned studies have shown that BCG plays a major role in the treatment of autoimmune brain diseases. Interestingly, a cohort study showed that BCG did not affect the prevalence of asthma, eczema or pollen heat during children's period. However, neonatal vaccination BCG is associated with a low incidence of asthma [67]. This appears to be achieved by suppressing the immune response of helper Th2-type cells specific to asthma. At the same time, since BCG may change the microenvironment of the bone marrow, BCG may regulate the maturation of immune cells and the production of cytokines in the early stage or even in the whole process, thereby achieving long-term protective effects. A systematic review concluded that BCG vaccination might not represent an effective primary preventative strategy against the development of allergic sensitization and diseases [68]. Even so, there is still a study suggesting that BCG vaccination in the early days of life may prevent asthma by adjusting the immune maturation process [69]. Although there is no clear evidence for the effective effect of BCG on asthma, it is worth further study to clarify the value BCG on other autoimmune diseases.
Other diseases and potential adverse effects
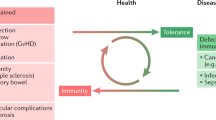
BCG may have certain effects on many other diseases. For instance, BCG's immunotherapy on bladder cancer is in connection with the significantly reduced risk of Alzheimer's disease and Parkinson's disease [70], and BCG reduces human malaria infection in a part of volunteers [71]. Surprisingly, there is insufficient evidence to prevent malaria by BCG vaccination [72, 73]. And BCG can promote the progression of chronic inflammatory diseases [74]. This shows trained immunity has beneficial effects on treating cancer, viruses, and autoimmune diseases, but trained immunity can also have harmful effects.
To sum up, although there is literature supporting the prote ctive effect of BCG induced trained immunity in some cases, the evidence in other infections and diseases is not always so strong.
Conclusions
There is still much to learn about trained immunity. First, the mechanisms of trained immunity should be studied more deeply, including the molecular mechanisms and signaling pathways on the different cells involved, and further elucidation of metabolic reprogramming and epigenetic processes are needed. Second, to facilitate its application to more clinical diseases, it is necessary to clarify the effect of trained immunization on different diseases. Finally, the nonspecific protection of trained immunization can be used to develop a new generation of vaccines to realize the cross-protection potential of different vaccines.
References
Petroff SA, Branch A. Bacillus calmette-guérin (BCG): animal experimentation and prophylactic immunization of children. Am J Public Health Nations Health. 1928;18(7):843–64.
Netea MG, Domínguez-Andrés J, Barreiro LB, Chavakis T, Divangahi M, Fuchs E, Joosten LAB, van der Meer JWM, Mhlanga MM, Mulder WJM, et al. Defining trained immunity and its role in health and disease. Nat Rev Immunol. 2020;20(6):375–88.
Netea MG, Joosten LA, Latz E, Mills KH, Natoli G, Stunnenberg HG, O’Neill LA, Xavier RJ. Trained immunity: a program of innate immune memory in health and disease. Science. 2016;352(6284):1098.
Ravenel M. La vaccination préventive contre la tuberculose par le “BCG.” Am Publ Health Assoc. 1928;18(8):1075.
Pearl R. On the pathological relations between cancer and tuberculosis. Proc Soc Exp Biol Med. 1928;26(1):73–5.
Levine MI, Sackett MF. Results of BCG immunization in New York City. Am Rev Tuberc. 1946;53:517–32.
Ferguson RG, Simes AB. BCG vaccination of Indian infants in Saskatchewan. Tubercle. 1949;30(1):5–11.
Rosenthal SR, Loewinsohn E, Graham ML, Liveright D, Thorne MG, Johnson V. BCG vaccination in tuberculous households. Am Rev Respir Dis. 1961;84:690–704.
Mackaness GB. The immunological basis of acquired cellular resistance. J Exp Med. 1964;120(1):105–20.
Mackaness GB. The influence of immunologically committed lymphoid cells on macrophage activity in vivo. J Exp Med. 1969;129(5):973–92.
Clark IA, Allison AC, Cox FE. Protection of mice against babesia and plasmodium with BCG. Nature. 1976;259(5541):309–11.
Civil RH, Warren KS, Mahmoud AA. Conditions for bacille calmette-guérin-induced resistance to infection with schistosoma mansoni in mice. J Infect Dis. 1978;137(5):550–5.
Bistoni F, Verducci G, Perito S, Vecchiarelli A, Puccetti P, Marconi P, Cassone A. Immunomodulation by a low-virulence, agerminative variant of candida albicans further evidence for macrophage activation as one of the effector mechanisms of nonspecific anti-infectious protection. J Med Vet Mycol. 1988;26(5):285–99.
Garly ML, Martins CL, Balé C, Baldé MA, Hedegaard KL, Gustafson P, Lisse IM, Whittle HC, Aaby P. BCG scar and positive tuberculin reaction associated with reduced child mortality in West Africa a non-specific beneficial effect of BCG. Vaccine. 2003;21:2782–90.
Netea MG, Quintin J, van der Meer JW. Trained immunity: a memory for innate host defense. Cell Host Microbe. 2011;9(5):355–61.
Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Ifrim DC, Saeed S, Jacobs C, van Loenhout J, de Jong D, Stunnenberg HG, et al. Bacille calmette-guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc Natl Acad Sci USA. 2012;109(43):17537–42.
Saeed S, Quintin J, Kerstens HH, Rao NA, Aghajanirefah A, Matarese F, Cheng SC, Ratter J, Berentsen K, van der Ent MA, et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science. 2014;345(6204):1251086.
Cheng S-c, Quintin J, Cramer R, Shepardson K, Saeed S, Kumar V, Giamarellos-Bourboulis E, Martens J, Rao N, Aghajanirefah A, et al. MTOR- and HIF-1α-mediated aerobic glycolysis as metabolic basis for trained immunity. Science. 2014. https://doi.org/10.1126/science.1250684.
Arts RJW, Carvalho A, La Rocca C, Palma C, Rodrigues F, Silvestre R, Kleinnijenhuis J, Lachmandas E, Goncalves LG, Belinha A, et al. Immunometabolic pathways in BCG-induced trained immunity. Cell Rep. 2016;17(10):2562–71.
Mitroulis I, Ruppova K, Wang B, Chen LS, Grzybek M, Grinenko T, Eugster A, Troullinaki M, Palladini A, Kourtzelis I, et al. Modulation of myelopoiesis progenitors is an integral component of trained immunity. Cell. 2018;172(1–2):147-161.e12.
Kaufmann E, Sanz J, Dunn JL, Khan N, Mendonca LE, Pacis A, Tzelepis F, Pernet E, Dumaine A, Grenier JC, et al. BCG educates hematopoietic stem cells to generate protective innate immunity against tuberculosis. Cell. 2018;172(1–2):176–90.
Kleinnijenhuis J, van Crevel R, Netea MG. Trained immunity: consequences for the heterologous effects of BCG vaccination. Trans R Soc Trop Med Hyg. 2015;109(1):29–35.
Zhang Q, Cao X. Epigenetic regulation of the innate immune response to infection. Nat Rev Immunol. 2019;19(7):417–32.
Donohoe DR, Bultman SJ. Metaboloepigenetics: interrelationships between energy metabolism and epigenetic control of gene expression. J Cell Physiol. 2012;227(9):3169–77.
Quintin J, Saeed S, Martens JHA, Giamarellos-Bourboulis EJ, Ifrim DC, Logie C, Jacobs L, Jansen T, Kullberg BJ, Wijmenga C, et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe. 2012;12(2):223–32.
Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Jacobs C, Xavier RJ, van der Meer JW, van Crevel R, Netea MG. BCG-induced trained immunity in NK cells: role for non-specific protection to infection. Clin Immunol. 2014;155(2):213–9.
Medzhitov R. Recognition of microorganisms and activation of the immune response. Nature. 2007;449(7164):819–26.
Arts RJW, Moorlag S, Novakovic B, Li Y, Wang SY, Oosting M, Kumar V, Xavier RJ, Wijmenga C, Joosten LAB, et al. BCG vaccination protects against experimental viral infection in humans through the induction of cytokines associated with trained immunity. Cell Host Microbe. 2018;23(1):89-100.e5.
Moorlag S, Rodriguez-Rosales YA, Gillard J, Fanucchi S, Theunissen K, Novakovic B, de Bont CM, Negishi Y, Fok ET, Kalafati L, et al. BCG vaccination induces long-term functional reprogramming of human neutrophils. Cell Rep. 2020;33(7): 108387.
Jeljeli M, Riccio LGC, Doridot L, Chêne C, Nicco C, Chouzenoux S, Deletang Q, Allanore Y, Kavian N, Batteux F. Trained immunity modulates inflammation-induced fibrosis. Nat Commun. 2019;10(1):5670.
Ishii KJ, Koyama S, Nakagawa A, Coban C, Akira S. Host innate immune receptors and beyond: making sense of microbial infections. Cell Host Microbe. 2008;3(6):352–63.
Cirovic B, de Bree LCJ, Groh L, Blok BA, Chan J, van der Velden W, Bremmers MEJ, van Crevel R, Handler K, Picelli S, et al. BCG vaccination in humans elicits trained immunity via the hematopoietic progenitor compartment. Cell Host Microbe. 2020;28(2):322–34.
Dinarello CA. The IL-1 family and inflammatory diseases. Clin Exp Rheumatol. 2002;20(5):S1-13.
Jha AK, Huang SC, Sergushichev A, Lampropoulou V, Ivanova Y, Loginicheva E, Chmielewski K, Stewart KM, Ashall J, Everts B, et al. Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity. 2015;42(3):419–30.
Bekkering S, Blok BA, Joosten LA, Riksen NP, van Crevel R, Netea MG. In vitro experimental model of trained innate immunity in human primary monocytes. Clin Vaccine Immunol. 2016;23(12):926–33.
Trunz BB, Fine P, Dye C. Effect of BCG vaccination on childhood tuberculous meningitis and miliary tuberculosis worldwide: a meta-analysis and assessment of cost-effectiveness. Lancet. 2006;367(9517):1173–80.
Stanley SJ, Howland C, Stone MM, Sutherland I. BCG vaccination of children against leprosy in uganda: final results. J Hyg (Lond). 1981;87(2):233–48.
Mangtani P, Abubakar I, Ariti C, Beynon R, Pimpin L, Fine PE, Rodrigues LC, Smith PG, Lipman M, Whiting PF, et al. Protection by BCG vaccine against tuberculosis: a systematic review of randomized controlled trials. Clin Infect Dis. 2014;58(4):470–80.
Old LJ, Clarke DA, Benacerraf B. Effect of bacillus calmette-guerin infection on transplanted tumours in the mouse. Nature. 1959;184(5):291–2.
Han RF, Pan JG. Can intravesical bacillus calmette-guérin reduce recurrence in patients with superficial bladder cancer? A meta-analysis of randomized trials. Urology. 2006;67(6):1216–23.
Morales A, Eidinger D, Bruce A. Intracavitary bacillus calmette-guerin in the treatment of superficial bladder tumors. J Urol. 1976;116(2):180–3.
Larsen ES, Joensen UN, Poulsen AM, Goletti D, Johansen IS. Bacillus calmette-guérin immunotherapy for bladder cancer: a review of immunological aspects, clinical effects and BCG infections. APMIS. 2020;128(2):92–103.
Miyake M, Nishimura N, Oda Y, Owari T, Hori S, Morizawa Y, Gotoh D, Nakai Y, Anai S, Torimoto K, et al. Intravesical bacillus calmette-guerin treatment-induced sleep quality deterioration in patients with non-muscle invasive bladder cancer: functional outcome assessment based on a questionnaire survey and actigraphy. Support Care Cancer. 2022;30(1):887–95.
Singh AK, Praharaj M, Lombardo KA, Yoshida T, Matoso A, Baras AS, Zhao L, Srikrishna G, Huang J, Prasad P, et al. Re-engineered BCG overexpressing cyclic di-AMP augments trained immunity and exhibits improved efficacy against bladder cancer. Nat Commun. 2022;13(1):878.
Pfahlberg A, Kölmel KF, Grange JM, Mastrangelo G, Krone B, Botev IN, Niin M, Seebacher C, Lambert D, Shafir R, et al. Inverse association between melanoma and previous vaccinations against tuberculosis and smallpox: results of the FEBIM study. J Invest Dermatol. 2002;119(3):570–5.
Sfakianos JP, Salome B, Daza J, Farkas A, Bhardwaj N, Horowitz A. Bacillus calmette-guerin (BCG): Its fight against pathogens and cancer. Urol Oncol. 2021;39(2):121–9.
Yao K, Wang W, Li H, Lin J, Tan W, Chen Y, Guo L, Lin D, Chen T, Zhou J, et al. Bacillus calmette guérin (BCG) activates lymphocyte to promote autophagy and apoptosis of gastric cancer MGC-803 cell. Cell Mol Biol. 2018;64(6):11–6.
Usher NT, Chang S, Howard RS, Martinez A, Harrison LH, Santosham M, Aronson NE. Association of BCG vaccination in childhood with subsequent cancer diagnoses: a 60-year follow-up of a clinical trial. JAMA Netw Open. 2019;2(9): e1912014.
Morra M, Kien N, Elmaraezy A, Abdelaziz O, Elsayed A, Halhouli O, Montasr A, Vu T, Ho C, Foly A, et al. Early vaccination protects against childhood leukemia: a systematic review and meta-analysis. Sci Rep. 2017;7(1):15986.
Biering-Sørensen S, Aaby P, Napirna BM, Roth A, Ravn H, Rodrigues A, Whittle H, Benn CS. Small randomized trial among low-birth-weight children receiving bacillus calmette-guérin vaccination at first health center contact. Pediatr Infect Dis J. 2012;31(3):306–8.
Gandhi NM, Morales A, Lamm DL. Bacillus calmette-guérin immunotherapy for genitourinary cancer. BJU Int. 2013;112(3):288–97.
Wardhana DE, Sultana A, Mandang V, Jim E. The efficacy of bacillus calmette-guerin vaccinations for the prevention of acute upper respiratory tract infection in the elderly. Acta Medica Indonesiana. 2011;43(3):185–90.
Moorlag S, Arts RJW, van Crevel R, Netea MG. Non-specific effects of BCG vaccine on viral infections. Clin Microbiol Infect. 2019;25(12):1473–8.
Leentjens J, Kox M, Stokman R, Gerretsen J, Diavatopoulos DA, van Crevel R, Rimmelzwaan GF, Pickkers P, Netea MG. BCG vaccination enhances the immunogenicity of subsequent influenza vaccination in healthy volunteers: a randomized, placebo-controlled pilot study. J Infect Dis. 2015;212(12):1930–8.
Libraty DH, Zhang L, Woda M, Acosta LP, Obcena A, Brion JD, Capeding RZ. Neonatal BCG vaccination is associated with enhanced T-helper 1 immune responses to heterologous infant vaccines. Trials Vaccinol. 2014;1:31–5.
Xue QJ, Yu HX, Liu A, Wang H, Li YQ, Chen T, Wang QL. The inhibitory effect of rBCG on EB virus-positive tumours using an EB virus fusion gene. Appl Microbiol Biotechnol. 2022;106(1):185–95.
Salem A, Nofal A, Hosny D. Treatment of common and plane warts in children with topical viable bacillus calmette-guerin. Pediatr Dermatol. 2013;30(1):60–3.
Podder I, Bhattacharya S, Mishra V, Sarkar TK, Chandra S, Sil A, Pal S, Kumar D, Saha A, Shome K, et al. Immunotherapy in viral warts with intradermal bacillus calmette-guerin vaccine versus intradermal tuberculin purified protein derivative: a double-blind, randomized controlled trial comparing effectiveness and safety in a tertiary care center in Eastern India. Indian J Dermatol Venereol Leprol. 2017;83(3):411.
Ristori G, Romano S, Cannoni S, Visconti A, Tinelli E, Mendozzi L, Cecconi P, Lanzillo R, Quarantelli M, Buttinelli C, et al. Effects of bacille calmette-guerin after the first demyelinating event in the CNS. Neurology. 2014;82(1):41–8.
Covian C, Retamal-Diaz A, Bueno SM, Kalergis AM. Could BCG vaccination induce protective trained immunity for SARS-CoV-2? Front Immunol. 2020. https://doi.org/10.3389/fimmu.2020.00970.
Berg MK, Yu Q, Salvador CE, Melani I, Kitayama S. Mandated bacillus calmette-guérin (BCG) vaccination predicts flattened curves for the spread of COVID-19. Sci Adv. 2020;6(32):1463.
Lindestam Arlehamn CS, Sette A, Peters B. Lack of evidence for BCG vaccine protection from severe COVID-19. Proc Natl Acad Sci USA. 2020;117(41):25203–4.
Kühtreiber WM, Tran L, Kim T, Dybala M, Nguyen B, Plager S, Huang D, Janes S, Defusco A, Baum D, et al. Long-term reduction in hyperglycemia in advanced type 1 diabetes: the value of induced aerobic glycolysis with BCG vaccinations. NPJ Vaccines. 2018. https://doi.org/10.1038/s41541-018-0062-8.
Sanjeevi CB, Das AK, Shtauvere-Brameus A. BCG vaccination and GAD65 and IA-2 autoantibodies in autoimmune diabetes in southern India. Ann N Y Acad Sci. 2002;958:293–6.
Lee J, Reinke EK, Zozulya AL, Sandor M, Fabry Z. Mycobacterium bovis bacille calmette-guérin infection in the CNS suppresses experimental autoimmune encephalomyelitis and Th17 responses in an IFN-gamma-independent manner. J Immunol. 2008;181(9):6201–12.
Faustman DL, Wang L, Okubo Y, Burger D, Ban L, Man G, Zheng H, Schoenfeld D, Pompei R, Avruch J, et al. Proof-of-concept, randomized, controlled clinical trial of bacillus-calmette-guerin for treatment of long-term type 1 diabetes. PLoS ONE. 2012;7(8): e41756.
Marks GB, Ng K, Zhou J, Toelle BG, Xuan W, Belousova EG, Britton WJ. The effect of neonatal BCG vaccination on atopy and asthma at age 7 to 14 years: an historical cohort study in a community with a very low prevalence of tuberculosis infection and a high prevalence of atopic disease. J Allergy Clin Immunol. 2003;111(3):541–9.
Arnoldussen DL, Linehan M, Sheikh A. BCG vaccination and allergy: a systematic review and meta-analysis. J Allergy Clin Immunol. 2011;127(1):246–53.
El-Zein M, Parent ME, Benedetti A, Rousseau MC. Does BCG vaccination protect against the development of childhood asthma? A systematic review and meta-analysis of epidemiological studies. Int J Epidemiol. 2010;39(2):469–86.
Klinger D, Hill BL, Barda N, Halperin E, Gofrit ON, Greenblatt CL, Rappoport N, Linial M, Bercovier H. Bladder cancer immunotherapy by BCG is associated with a significantly reduced risk of Alzheimer’s disease and Parkinson’s disease. Vaccines. 2021;9(5):491.
Walk J, de Bree LCJ, Graumans W, Stoter R, van Gemert GJ, van de Vegte-Bolmer M, Teelen K, Hermsen CC, Arts RJW, Behet MC, et al. Outcomes of controlled human malaria infection after BCG vaccination. Nat Commun. 2019;10(1):874.
Witschkowski J, Behrends J, Frank R, Eggers L, von Borstel L, Hertz D, Mueller AK, Schneider BE. BCG provides short-term protection from experimental cerebral malaria in mice. Vaccines. 2020;8(4):745.
Rodrigues A, Schellenberg JA, Roth A, Benn CS, Aaby P, Greenwood B. Revaccination with bacillus calmette-guerin (BCG) vaccine does not reduce morbidity from malaria in African children. Trop Med Int Health. 2007;12(2):224–9.
Li X, Wang H, Yu X, Saha G, Kalafati L, Ioannidis C, Mitroulis I, Netea MG, Chavakis T, Hajishengallis G. Maladaptive innate immune training of myelopoiesis links inflammatory comorbidities. Cell. 2022;185:1709–27.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China and Yunnan Province Department of Science and Technology-Kunming Medical University Joint Fund Projects. The funding institutions had no involvement in the design of the study or review of the manuscript.
Funding
National Natural Science foundation of China (No. 82160304, 32060180, 81860644, and 81560596) and Joint foundation of Yunnan Province Department of Science, Technology-Kunming Medical University [No. 2019FE001 (-002) and 2017FE467 (-001)].
Author information
Authors and Affiliations
Contributions
BFK, LAH, CJJ, GL, and WXY conceived and designed the study. FYX and LBX conducted the database search and screening. SJQ and FYX evaluated the data. LMX, PL, and CJJ conducted the quality assessment. CJJ, GL, and WXY drafted the manuscript. BFK and LAH revised and approved the manuscript. Everyone agreed to publish it. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Competing interests
We declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chen, J., Gao, L., Wu, X. et al. BCG-induced trained immunity: history, mechanisms and potential applications. J Transl Med 21, 106 (2023). https://doi.org/10.1186/s12967-023-03944-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-023-03944-8