
Abstract
Acute myocardial infarction has long been the leading cause of death in coronary heart disease, which is characterized by irreversible cardiomyocyte death and restricted blood supply. Conventional reperfusion therapy can further aggravate myocardial injury. Stem cell therapy, especially with mesenchymal stem cells (MSCs), has emerged as a promising approach to promote cardiac repair and improve cardiac function. MSCs may induce these effects by secreting exosomes containing therapeutically active RNA, proteins and lipids. Notably, normal cardiac function depends on intracardiac paracrine signaling via exosomes, and exosomes secreted by cardiac cells can partially reflect changes in the heart during disease, so analyzing these vesicles may provide valuable insights into the pathology of myocardial infarction as well as guide the development of new treatments. The present review examines how exosomes produced by MSCs and cardiac cells may influence injury after myocardial infarction and serve as therapies against such injury.
Graphical Abstract

Video Abstract
Similar content being viewed by others

Introduction
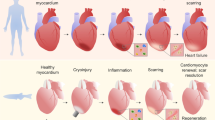
Despite considerable improvements in healthcare worldwide, acute myocardial infarction (AMI), defined as the death of myocardial cells due to prolonged ischemia, is the most serious manifestation of coronary artery disease and the primary cause of associated death [1, 2]. Cells of adult heart cannot regenerate after loss due to ischemia, so myocardial infarction (MI) can cause the irreversible loss of too many cardiomyocytes, leading to left ventricular remodeling and even progressive heart failure [3].
The current standard treatment for AMI is reperfusion, but this can aggravate cardiomyocyte dysfunction and tissue injury, a process known as ischemia/reperfusion (I/R) injury [4]. Such injury can lead initially to oxidative stress and inflammation, and can rapidly develop into apoptosis and necrosis [5]. This highlights the need to develop new treatments to reduce cardiomyocyte death and promote cardiac repair after MI.
A fruitful approach may be to exploit exosomes, extracellular vesicles containing regulatory RNAs, lipids and proteins that are normally secreted by many types of cells, including cardiomyocytes, endothelial cells (ECs), immune cells, and stem cells. Exosomes serve as important paracrine signaling vehicles, helping to promote angiogenesis, inhibit harmful ventricular remodeling, improve heart function, inhibit local inflammation and regulate immune responses [6].
This review provides an overview of recent developments in our understanding of exosomes secreted by mesenchymal stem cells (MSCs) and cardiac cells over the past five years. Certain exosomes exhibit great potential in the treatment of MI, while others impact the progression and prognosis of MI [6, 7]. Although no studies have compared or independently described exosomes of animal and human origin, multiple studies have demonstrated that exosomes derived from both human and rat MSCs attenuate cardiomyocyte apoptosis in rats with MI [8, 9]. Consequently, we postulate that exosomes of homologous cellular origin from different races may share similar biological properties. Furthermore, this review examines the prospects and challenges of utilizing exosomes to treat patients after MI, with the aim of providing valuable information for future research that compares the specific biological functions of exosomes of animal and human origin.
Characteristics of exosomes
Exosomes are a type of extracellular vesicles (EVs) with diameters between 40 and 160 nm [10]. They are secreted by dendritic cells, mast cells, platelets, and MSCs as well as other cell types, and they are found in most body fluids of humans and animals, including plasma, serum, saliva, amniotic fluid, breast milk, and urine [11]. They were discovered by Pan and Johnstone during their studies of how sheep reticulocytes mature into erythrocytes [12]. Since its discovery, exosomes have been extensively studied for their biological characteristics, function, and potential clinical application.
Exosomes are generated from multivesicular endosomes that fuse with the cytoplasmic membrane and are secreted, instead of being degraded by lysosomes or autophagosomes [13]. Exosomes can enter recipient cells by employing different mechanisms such as endocytosis, direct fusion with membranes, or binding to receptors on the cell surface, and thus achieve their complex biological effects (Table 1) [14,15,16,17,18]. Exosomal cargo comprises cell surface and intracellular proteins, RNA, DNA, lipids, and metabolites (e.g. amino acids, ATP, amides), and its type and content are influenced by donor cells, microenvironment, or physiological conditions. By transferring these biomolecules containing important information to different cells, exosomes are able to influence gene transcription and cell proliferation [10]. Investigation of exosomal cargo and its functions may provide insight into many cell–cell communication processes in health and disease.
Biomarkers of exosomes are also a key research focus. The complexity of exosomes is further amplified by their specific biomarkers, which possess characteristic compositions of donor cells. For example, tumor-secreted exosomes are usually highly expressed in heat shock proteins, whereas B cell-secreted exosomes contain large amounts of tetraspanins [19, 20]. Nevertheless, the biomarkers of exosomes also exhibit homogeneity. Typical biomarkers of exosomes include proteins such as CD9, CD81, CD63, ceramide, tumor susceptibility gene 101 (TSG101), as well as apoptosis-linked gene 2-interacting protein X (ALIX), all of which are involved in the origin and biogenesis of exosomes (Table 2) [14, 21,22,23]. The properties of the biomarkers suggest an important role in the identification and purification of exosomes, and also give exosomes great potential for clinical applications such as disease diagnosis and targeted therapies.
Exosomes possess a wide and unique advantage in drug delivery and treatment due to their endogeneity and heterogeneity in applied research. Their endogeneity provides superior biocompatibility and low toxicity compared to common nanocarriers, making them excellent gene drug candidate vectors [24]. On the other hand, heterogeneity is one of the most important features of exosomes, which largely determines their function. The heterogeneity of exosomes is reflected by various factors, such as multiple mechanisms of exosome biogenesis, diverse cellular sources, content, and functional heterogeneity in their impact on recipient cells [25]. A deeper understanding of exosome characteristics will significantly advance related research in this field.
Exosomes derived from MSCs
If the ability to divide and differentiate into multiple cell types makes stem cells particularly interesting for medicine [26], then exosomes that can promote the repair and regeneration of injured tissue are also particularly attractive. Among the various types of stem cells that have shown therapeutic potential [27]. MSCs stand out because they have the potential of multi-directional differentiation and can function in a variety of adult tissues, including bone marrow, fat, umbilical cord blood and placenta [28,29,30,31]. Their ability to promote angiogenesis and recovery of ischemic tissue make them attractive for treating heart disease [32]. They secrete exosomes containing a broad array of regulatory RNAs with potential to ameliorate tissue injury after MI [33]. Exosomes from different MSCs mostly have common therapeutic effects, but their specific mechanisms are slightly different, which is closely related to their contents [34,35,36,37]. Multiple studies have shown that functionally diverse non-coding RNAs (ncRNAs) in exosomal cargoes can account for the vast majority of their therapeutic effects, and the mechanisms responsible for their effects have been explored in depth. Figure 1 depicts the biogenesis of exosome, and the potentially therapeutic functions of ncRNAs in exosomes derived from MSCs. We aim to elucidate the molecular mechanisms underlying the action of MSC-derived exosomes in order to facilitate their clinical translation.

Biogenesis of mesenchymal stem cell-derived exosomes and potentially therapeutic functions of non-coding RNAs they carry. The process of exosome biogenesis involves a double invagination of the plasma membrane and the formation of intracellular multivesicular bodies (MVBs). The plasma membrane invaginates for the first time and undergoes endocytosis, generating early sorting endosomes in the cytoplasm. Early sorting endosomes can mature into late sorting endosomes, which further generate MVBs. MVBs are formed by the second invagination of the plasma membrane which can either fuse with lysosomes or autophagosomes to be degraded or fuse with the plasma membrane to release intraluminal vesicles (i.e., exosomes). CircRNA, circular RNA; LncRNA, long non-coding RNA; miRNA, microRNA; ncRNA, non-coding RNA; ROS, reactive oxygen species, TSG101, Tumor susceptibility gene 101
Exosomes derived from bone marrow MSCs
Bone marrow MSCs (BMSCs) show weak immunogenicity, multidirectional differentiation and high transplantability, making them well-suited for treating cardiovascular diseases. BMSCs have been shown to be safe in clinical studies [38], and exosomes derived from BMSCs can influence various pathways in cardiomyocytes and macrophages to reduce myocardial injury and improve cardiac function. On the whole, exosomes derived from BMSCs can achieve good therapeutic effects through the joint action of various mechanisms.
Inhibition of cardiomyocyte death
Reperfusion of ischemic myocardium can exacerbate apoptosis, necrosis and cell death [39]. MicroRNA (miRNA) is a small (~ 21 nucleotide) ncRNA, and miRNAs in BMSC-derived exosomes can coordinately inhibit genes that drive cardiomyocyte apoptosis and heart injury. For example, miR-150-5p in BMSC-derived exosomes downregulates the pro-apoptotic B-cell lymphoma-associated X (Bax) protein to reduce cardiomyocyte apoptosis and improve cardiac function in a mouse model of MI [40]. BMSC-derived exosomes also carry miR-183-5p, which inhibits expression of forkhead box O1 (FOXO1) to reduce apoptosis and oxidative stress in cardiomyocytes [41]; miR-21a-5p, which downregulates the pro-apoptotic genes programmed cell death 4 (Pdcd4) and factor related apoptosis ligand (Fasl), protecting infarcted myocardium [42]; miR-25-3p, which downregulates the pro-apoptotic genes Fasl and phosphatase and tensin homolog (Pten) [43]; and miR-125b, which downregulates the pro-apoptotic genes P53 and B-cell lymphoma-2 antagonist/killer 1 (Bak1) [44]. This reflects the complexity of the signaling network in target cells regulated by exosomes. It involves not only multiple miRNAs acting on the same gene or pathway, but also the generation of synergistic effects through a single miRNA targeting multiple gene products.
During myocardial I/R injury, the activation of NOD-like receptor thermal protein domain associated protein 3 (NLRP3) inflammasomes can lead to the secretion of inflammatory cytokines and pyroptosis of cardiomyocytes [45]. BMSC-derived exosomes containing miR-182-5p can inhibit the expression of pro-pyroptosis protein gasdermin D (GSDMD) and toll-like receptor 4 (TLR4), as well as the TLR4/nuclear factor-κB (NF-κB) signaling pathway, thereby exerting powerful anti-inflammatory and anti-pyroptosis effects [46, 47].
Interestingly, BMSC-derived exosomes contain miRNAs that regulate several therapeutically useful signaling pathways (Fig. 2). For instance, regulatory RNAs in BMSC-derived exosomes can influence the phosphatidylinositol3-kinase/ protein kinase B (PI3K/AKT) pathway to attenuate cardiac fibrosis after MI and myocardial injury after hypoxia [48, 49]. Additionally, several miRNAs can regulate the C-Jun N-terminal kinase (JNK) pathway, which is a key regulatory pathway of apoptosis. For example, miR-338 acts through the mitogen-activated protein kinase 2/JNK (MAP3K2/JNK) signaling pathway to attenuate cardiomyocyte apoptosis, while miR-455-3p can inhibits mitogen-activated protein kinase 1/mitogen-activated protein kinase 4/JNK (MEKK1-MKK4-JNK) signaling pathway [50, 51]. Moreover, miR-143-3p regulates autophagy by inhibiting the checkpoint kinase 2 (CHK2)/Beclin2 pathway to effectively inhibit cardiomyocyte apoptosis [52].

Selected signaling pathways through which exosomes from MSCs exert therapeutic effects. ADMSC-exo, exosomes derived from adipose mesenchymal stem cells; AIFM3, apoptosis-inducing factor, mitochondrion-associated 3; AKT, protein kinase B; BMSC-exo, exosomes derived from bone marrow mesenchymal stem cells; CHK2, checkpoint kinase 2; DMBT1, deleted in malignant brain tumors 1; EZH2, enhancer of zeste 2 polycomb repressive complex 2; FOXO3, forkhead box O3; GSK3β, glycogen synthase kinase 3β; HMGA2, high mobility group AT-hook 2; IGF-1, insulin-like growth factor-1; JNK, C-Jun N-terminal kinase; MAP3K2, mitogen-activated protein kinase kinase kinase 2; MEKK1, mitogen-activated protein kinase kinase kinase 1; MKK4, mitogen-activated protein kinase kinase 4; NF-κB, nuclear factor-κB; p-AKT, phosphorylated protein kinase B; PI3K, phosphatidylinositol3-kinase; PP2A, protein phospholipase 2A; S1P, sphingosine 1-phosphate; S1PR1, sphingosine-1-phosphate receptor 1; SK1, sphingosine kinase 1; TLR4, toll-like receptor 4; UMSC-exo, exosomes derived from umbilical cord mesenchymal stem cells
BMSC-derived exosomes also carry long non-coding RNAs (lncRNAs), which usually regulate gene expression by interacting with miRNAs. For example, lncRNA KLF3-AS1 binds to miR-138-5p to upregulate sirtuin 1 (Sirt1), thereby inhibiting pyroptosis and slowing progression of injury following MI [53]. Another study is lncA2M-AS1, which sponges miR-556-5p to upregulate the anti-apoptotic protein X-linked inhibitor of apoptosis protein (XIAP), reducing myocardial apoptosis and oxidative stress caused by I/R injury [54]. By targeting miR-497/IGF1 axis, the lncRNA HCP5P in BMSC-derived exosomes also inhibits the PI3K/AKT signaling pathway, thereby increasing the viability of cardiomyocytes following hypoxia/reperfusion and decreasing apoptosis [55].
MI kills cardiomyocytes not only by promoting autophagy and apoptosis but also ferroptosis, in which the intracellular glutathione-dependent antioxidant defense system is defective, resulting in the accumulation of toxic reactive oxygen species (ROS) and iron [56]. BMSC-derived exosomes contain lncRNA Mir9-3hg that upregulates peroxoredoxin 6 (PRDX6) to decrease levels of iron and ROS in cardiomyocytes after I/R [57]. These endogenous lncRNAs contained within exosomes serve as natural therapeutic agents and offer a highly valuable gene therapy strategy for the treatment of cardiovascular diseases.
Promotion of angiogenesis
MI injures the vascular endothelium and restricts myocardial blood supply, so rescuing angiogenesis is critical [58]. BMSC-derived exosomes upregulate platelet-derived growth factor receptor-β (PDGFR-β) to enhance microvascular regeneration, inhibit fibrosis, and maintain long-term cardiac function after I/R injury [59]. This study also found that BMSC-exo had even better therapeutic effects than BMSCs themselves, suggesting that exosomes have great potential for future practical applications. These effects appear to depend on miR-221-3p, since lower levels of this miRNA in exosomes lead to weaker therapeutic effects [60].
While naturally occurring exosomes show therapeutic potential, genetic engineering can be used to create more potent exosomes in the laboratory. For example, infecting BMSCs with recombinant lentivirus allowed researchers to create exosomes containing abundant hypoxia-inducible factor 1 (HIF-1α), which rescued angiogenic, migratory and proliferative functions of hypoxia-injured human umbilical vein endothelial cells (HUVECs), and led to neovessel formation and inhibition of fibrosis in a rat model of MI [58]. In another approach, exposing BMSCs to ischemic heart extract rich in interleukin (IL)-22 led to exosomes carrying high levels of deleted in malignant brain tumors 1 (DMBT1), which can activate PI3K-AKT/ glycogen synthase kinase 3β(GSK3β)/β-catenin/ vascular endothlial growth factor (VEGF) signaling to enhance migration and proliferation of HUVECs [34].
Inhibition of inflammation
Death of cardiomyocytes during myocardial I/R injury activates an inflammatory response that has a significant impact on the final extent of myocardial injury. Activation of inflammasomes can induce pyroptosis, leading to the release of intracellular contents into the extracellular environment. This, in turn, activates the inflammatory response and further exacerbates myocardial injury [53, 61]. Excessive or prolonged inflammation degrades the extracellular matrix, leading to ventricular remodeling [62]. Therefore, treating MI requires timely suppression of the inflammatory response.
Macrophages play a critical role in the progression and regression of inflammation [63]. Macrophages can be broadly divided into two types: M1 or “classically activated” macrophages secrete large amounts of pro-inflammatory factors, while M2 or “alternatively activated” macrophages produce various anti-inflammatory factors and growth factors. Soon after MI, M1 macrophages are recruited into the infarcted myocardium and exhibit strong phagocytic activity and pro-inflammatory activity, after which M2 macrophages come to dominate and resolve inflammation to allow myocardial repair [64]. BMSC-derived exosomes have been shown to activate AKT1/AKT2 signaling and the nuclear factor erythroid 2-related factor 2/heme oxygenase 1 (NRF2/HO-1) axis, while inhibiting NF-κB signaling. This shift in signaling pathways can change macrophages from an M1 to an M2 phenotype, which helps to reduce inflammation after MI [65, 66]. Treating BMSCs with fibronectin type III domain-containing protein 5 (FNDC5) can further enhance the ability of exosomes to promote this shift in macrophage phenotype [66].
BMSC-derived exosomes can also reduce the activity of the NLRP3 inflammasome in a mouse model of left coronary artery ligation [67]. As well, they carry miR-181a-5p that downregulates activating transcription factor 2 (ATF2) to protect the myocardium [68]. Exosomes carrying higher levels of this miRNA can be prepared by first treating the BMSCs with lipopolysaccharide [68]. Future research should continue to explore how to improve the therapeutic effects of exosomes by pretreating the parental cells. For example, studies should explore pretreating MSCs with inflammatory factors that mimic the inflammatory environment after MI, or with stromal factors or other cytokines that promote MSC growth.
Other effects
BMSC-derived exosomes can inhibit harmful ventricular remodeling. They contain miR-185, which downregulates suppressor of cytokine signaling 2 (SOCS2), an inhibitor of growth hormone/IGF signaling [69]; and miR-29b-3p, which downregulates A disintegrin and metalloproteinase with thrombospondin 16 (ADAMTS16), blocking myocardial fibrosis and promoting angiogenesis in a rat model of MI [70].
BMSCs have been shown to promote myocardial recovery after infarction by influencing the renin-angiotensin system (RAS). Following MI, the activation of the neuro-humoral regulation mechanism, particularly the RAS, can accelerate the process of ventricular remodeling [71], which is associated with poor prognosis in MI patients [72]. Angiotensin-converting-enzyme inhibitors and angiotensin-receptor blockers, which can inhibit the RAS, occupy an important part in the pharmacotherapy of MI [73]. BMSC-derived exosomes accelerate the conversion of angiotensin II to angiotensin-(1-7), reducing the adverse effects of angiotensin II on cardiomyocytes and improving cardiac remodeling, as well as protecting cardiomyocytes from oxidative stress and associated apoptosis [72, 74]. This humoral therapeutic mechanism warrants further exploration in future research.
Work to explore the therapeutic potential of BMSC-derived exosomes should also explore their combination with BSMCs themselves. One study found that first injecting the exosomes followed by the cells effectively reduced scar size and restored cardiac function after acute MI [75]. Those authors proposed that the exosomes dampened inflammation in the heart and thereby created an environment to ensure survival of the subsequently injected BMSCs. This study not only demonstrates the effectiveness of exosome therapy, but also provides additional possibilities for future methods of clinical application of exosomes.
Exosomes derived from adipose MSCs
Adipose MSCs (ADMSCs) have similar biological properties and multidirectional differentiation potential as BMSCs [76]. The source tissue is abundant and exosomes can be extracted from it efficiently and painlessly [77]. For these reasons, ADMSCs may replace BMSCs as a new stem cell source for regenerative medicine.
Inhibition of cardiomyocyte apoptosis
Exosomes derived from ADMSCs contain miR-671, which downregulates transforming growth factor β receptor 2 (TGFBR2) and decreases mothers against decapentaplegic homolog (Smad2) phosphorylation, thereby reducing cardiomyocyte apoptosis, fibrosis and inflammation [78]. They also attenuate apoptosis after I/R and protect ischemic myocardium by activating the Wnt/β-catenin signaling pathway [35]. It is important to note that the Wnt/β-catenin signaling pathway influences macrophage polarization, angiogenesis, and other aspects [79, 80]. However, this study only examines its effects from one perspective, and a comprehensive and in-depth investigation could provide a fuller understanding of its role.
Inhibition of inflammation
Similar to BMSC exosomes, exosomes derived from ADMSCs can shift macrophages toward an M2 phenotype and inhibit fibrosis and inflammation after MI, but the specific mechanisms may be different. Deng et al. found that this effect is achieved by activating the sphingosine 1-phosphate/sphingosine kinase 1/sphingosine-1-phosphate receptor 1 (S1P/SK1/S1PR1) axis [81].
Promotion of angiogenesis
In hypoxic/ischemic environment, HIF-1 can drive the transactivation of dozens of genes involved in angiogenesis and play a critical role in mediating cardioprotection [82, 83]. Exosomes derived from ADMSCs contain miR-31, which downregulates an asparagine hydroxylase that blocks the binding of HIF-1α to coactivator p300, thereby “rescuing” expression of angiogenic genes [83].
Exosomes derived from umbilical cord MSCs
Inhibition of myocardial cell death
Exosomes derived from UMSCs exhibit a potential to promote cardiomyocyte survival after MI by inhibiting various cell death pathways such as apoptosis, autophagy, pyroptosis, and ferroptosis. They contain both miR-19a and circular RNA circ-0001273 that attenuate myocardial injury and inhibit cardiomyocyte apoptosis, which act by downregulating the transcription factor SRY (sex determining region Y)-box 6 (SOX6) and inhibiting miR-199b-3p, respectively [86, 87]. Moreover, UMSC-derived exosomes can promote the expression of Smad7 by inhibiting miR-125b-5p in the infarcted myocardium, providing additional myocardial protection [9]. They also carry lncRNA UCA1, which inhibits both autophagy and apoptosis [88]. Interestingly, Sun et al. showed that lncRNA UCA1 in exosomes secreted by hypoxic MSCs also achieved myocardial protection by sponging miR-873-5p [89]. Although this study does not specify the origin of MSCs, it suggests the importance of exploring the functional diversity of ncRNAs in exosomes.
Furthermore, UMSC-derived exosomes contain miR-100-5p that downregulates forkhead box O3 (FOXO3), which suppresses inflammation by suppressing NLRP3 inflammasome activation and protects cardiomyocytes from pyroptosis and injury [90]. By delivering miR-23a-3p, the exosomes can also inhibit ferroptosis by targeting the divalent metal transporter recombinant divalent metal transporter 1 (DMT1), which promotes iron accumulation in cardiomyocytes [91].
Inhibition of inflammation
In contrast to regulation of macrophage polarization, exosomes derived from UMSCs exert anti-inflammatory effects through cardiac fibroblasts (CFs). Activated CFs after MI are characterized by fibroblast-myofibroblast transdifferentiation, accelerated proliferation, and extracellular matrix accumulation, which are essential for the dynamic homeostasis and remodeling of cardiac tissue, cardiomyocyte proliferation and angiogenesis [92, 93]. Analogous to macrophages, these fibroblasts adopt a pro-inflammatory phenotype soon after MI, after which they differentiate into myofibroblasts, which secrete anti-inflammatory factors and extracellular matrix proteins to repair and stabilize cardiac tissue [94]. Exosomes from human UMSCs promote differentiation of CFs toward myofibroblasts, thus attenuating the inflammatory response, reducing cardiomyocyte apoptosis, and promoting cardiac repair [95].
These exosomes may also inhibit inflammation and promote repair by activating regulatory T cells through a mechanism involving activation of protein phospholipase 2A (PP2A)/p-AKT/FOXO3 signaling [96]. That study demonstrated not only the potential for exosome-mediated immunomodulation but also the effectiveness of intrapericardial injection of exosomes.
Other effects
Exosomes derived from UMSCs contain tissue inhibitor of metalloproteinase 2 (TIMP2), which inhibits matrix metalloproteinases and thereby inhibits harmful remodeling of the extracellular matrix and deposition of collagen, and the exosomes promote ECs proliferation and migration in damaged myocardium, leading to angiogenesis, while also inhibiting apoptosis [97, 98]. That study showed that levels of TIMP2 in exosomes can be increased by overexpressing TIMP2 in the parental UMSCs.
Notably, exosomes derived from UMSCs can also repair myocardium by affecting the function of other MSCs. Exosomes from UMSCs can enhance the ability of BMSCs to repair myocardium by delivering miR-136, which downregulates apoptotic protease activating factor- 1 (Apaf1) [99].
Exosomes derived from cardiac MSCs
Cardiac MSCs play an important role in various aspects of ventricular remodeling, angiogenesis, and myocardial repair after MI [100,101,102]. Cardiac MSCs maintain cardiac homeostasis and promote cardiac repair through paracrine signaling [103]. Exosomes from cardiac MSCs can inhibit apoptosis, enhance myocardial angiogenesis and increase proliferation of cardiomyocytes in ischemic myocardium [104]. However, the related studies of cardiac MSC-derived exosomes are still limited, and further exploration is needed.
Exosomes derived from cardiac cells
The ischemic-hypoxic microenvironment during MI induces apoptosis of most cell types within the heart, including cardiomyocytes, ECs, and macrophages [105]. Apoptosis of cardiomyocytes induces CFs to proliferate, leading to myocardial remodeling and impairing cardiac function. Exosomes produced by cardiac cells, especially cardiomyocytes, are an important source of exosomes in the heart and have a regulatory effect on cardiac function. For example, cardiomyocytes and ECs in the heart produce abundant exosomes under normal conditions and after MI [106]; and exosomes produced by CFs regulate cardiac hypertrophy [107]. Understanding the range of exosomes produced by cardiac cells and their molecular contents is important for exploiting them as potential therapies following MI (Table 3).
Exosomes derived from cardiomyocytes
Although cardiomyocytes are not typical secretory cells, they secrete exosomes that mediate communication between healthy and damaged cells under healthy and ischemic conditions [131]. These exosomes can also regulate cell proliferation, migration, differentiation, survival, and angiogenesis.
Promotion of cell death
Exosomes from cardiomyocytes can exert both harmful and beneficial effects. For example, their content of miR-328-3p increases after infarction, and this miRNA activates intracellular caspase signaling to promote apoptosis [108]. In fact, infarcted cardiomyocytes can send exosomes to nearby cardiomyocytes, promoting apoptosis. Similarly, exosomes from infarcted cardiomyocytes contain abundant miR-19a-3p, which downregulates HIF-1α to inhibit proliferation of ECs and angiogenesis [109].
The same hypoxia that triggers cardiomyocyte apoptosis after MI upregulates the lncRNA AK139128 in cardiomyocytes and increases its levels in exosomes secreted by these cells. This lncRNA promotes apoptosis and inhibits proliferation of CFs, exacerbating myocardial injury after infarction [110]. However, hypoxia also activates transforming growth factor β1 (TGF-β1) to inhibit the apoptosis of CFs [132]. Interestingly, TGF-β signaling can interact with the Wnt signaling pathway and plays a key role in the differentiation of CFs [133]. This highlights the complex interplay within the signaling pathway network, which should be thoroughly explored in future studies.
Regulation of inflammation
AMI temporarily increases the production of exosomes and microvesicles, mainly by cardiomyocytes and ECs. These vesicles accumulate in the ischemic myocardium, where they are rapidly taken up by infiltrating monocytes, which release chemokines and pro-inflammatory cytokines [134]. Ferroptosis lowers the levels of miR-106b-3p in exosomes from cardiomyocytes, leading to activation of Wnt signaling and a shift of cardiac macrophages to the M1 phenotype, exacerbating myocardial inflammation [80].
On the other hand, exosomes from hypoxic cardiomyocytes promote macrophage polarization toward the M2 phenotype, attenuating hypoxia-induced cardiomyocyte injury [111]. Further research is needed to clarify the how these exosomes influence macrophage polarization.
Other effects
Excessive activation of CFs leads to harmful cardiac remodeling that can culminate in cardiac fibrosis. Hypoxic cardiomyocytes secrete exosomes containing miR-208a, which are taken up by CFs, in which the miRNA promotes fibroblast proliferation and myofibroblast differentiation, exacerbating cardiac fibrosis and the associated deterioration of cardiac function [112].
On the other hand, exosomes from ischemic cardiomyocytes can protect myocardium from oxidation-induced injury; promote ECs proliferation, sprouting, and tube formation through circHIPK3; and stimulate neovascularization through miR-222 and miR-143 [113, 114]. These studies elucidate the intricacy of intercellular interactions among cardiac cells resulting from the overlapping stages of inflammation, fibrosis, and angiogenesis in damaged myocardium. They emphasize the significance of adopting a comprehensive approach to comprehend the multifaceted factors and mechanisms involved in MI.
Exosomes from cardiomyocytes can regulate cardiac cells but also MSCs. In co-cultures of ischemic cardiomyocytes with MSCs, the cardiomyocytes secreted exosomes containing the lncRNA KLF3-AS1, which upregulated signal transducer and activator of transcription 5B (STAT5B) in MSCs, leading them to secrete IGF-1 to promote repair of myocardial tissue damaged by I/R [115]. To explore therapeutic possibilities, future research should explore the full range of cells that exosomes from ischemic/hypoxic cardiomyocytes can target, as well as the resulting biological effects.
Exosomes derived from cardiosphere-derived cells
Cardiosphere-derived cells (CDCs) are a type of cardiac stromal/progenitor cells that exert beneficial immunomodulatory, anti-fibrotic and pro-regenerative properties in diseased cardiac and skeletal muscle [135]. Studies in various animal models of MI and phase I trials with patients have shown that CDCs can reduce scarring, prevent unfavorable remodeling and increase survival [136, 137]. As a result, phase II trials are underway to assess the ability of CDCs to reduce scarring after MI. It appears that exosomes secreted by CDCs explain most if not all the cardioprotective, anti-apoptotic and regenerative effects [138, 139].
Inhibit inflammation
After cardiac I/R, exosomes from CDCs localize to ischemic tissues, reduce infarct size, and polarize macrophages into a highly phagocytic state similar to M2. These macrophages express abundant anti-inflammatory genes, efficiently phagocytose necrotic cellular debris, and attenuate excessive inflammatory stress within the infarcted heart. These effects appear to be mediated at least in part by miR-181b within the exosomes; this miRNA downregulates protein kinase δ (PKCδ) in macrophages and polarizes macrophages toward a cardioprotective phenotype [119]. Besides, Exosome-enriched EVs derived from CDCs contain abundant non-coding Y RNA called EV-YF1, which upregulates the pro-inflammatory factor IL-10 in macrophages, helping protect cardiomyocytes from oxidative stress [118]. These results suggest the importance of exploring ncRNA cargo in exosomes more extensively.
Other effects
The potent pro-angiogenic and anti-fibrotic properties of CDC-secreted exosomes can produce multiple beneficial effects, such as reducing scarring and improving contractile function when injected intramyocardially in an ischemic cardiomyopathy model [136]. This can be attributed to the fact that the new capillaries not only supply nutrients to the infarction border zone, but also provide energy for the differentiation of CFs, thereby compensating for the maintenance of cardiac structure and functional integrity [82, 140]. Additionally, through the reduction of myocardial fibrosis, they can enhance conduction and make the post-ischemic heart less susceptible to arrhythmias [141].
Exosomes derived from ECs and endothelial progenitor cells
ECs regulate vascular tone, angiogenesis, homeostasis, and inflammation in vivo and are major players in cardiovascular physiology and pathophysiology. Given the ability of exosomes from ECs to protect the brain and kidney from I/R injury, several studies have explored their potential against myocardial I/R injury [142]. Endothelial progenitor cells (EPCs), which serve as precursors to ECs, also secrete exosomes that may have therapeutic benefits in treating injury following MI [143].
Promotion of angiogenesis
ECs rely on exosomes, in part, to induce angiogenesis [144]. For example, the exosomal level of profilin 2 (PFN2) increased in the serum of post-MI patients and animal models during angiogenesis. When exosomes derived from endothelial cells overexpressing PNF2 were injected into MI mice, they delivered PNF2 to endothelial cells, promoting angiogenesis and protecting the cells from inflammatory damage through the PI3K-PFN2-extracellular regulated protein kinase (ERK) axis [120].
Although the functional substrates and potential molecular mechanisms of EPCs therapy for MI have not been fully elucidated, studies have shown that the angiogenesis effects of exosomes from EPCs are closely related to ECs. Exosomes from these progenitor cells contain miR-1246 and miR-1290, which upregulate E74-like ETS transcription factor 5 (ELF5) and specificity protein 1 (SP1), promoting the conversion of CFs to ECs, leading to angiogenesis [124]. The exosomes also contain miR-218-5p and miR-363-3p that upregulate p53 signaling and downregulate JMY to promote the mesenchymal-endothelial transition and angiogenesis [125].
Inhibit cardiomyocyte death
Exosomes from ECs can inhibit apoptosis of cardiomyocytes, such as by stimulating PI3K/AKT signaling [121]. For example, exosomes from vascular ECs also contain the lncRNA LINC00174, which downregulates P53 in cardiomyocytes, thereby inhibiting the AKT/ adenosine 5 ‘-monophosphate (AMP)-activated protein kinase (AMPK) pathway and attenuating cellular autophagy and apoptosis [122].
Other effects
In the post-MI ischemic microenvironment, aggregation and hyperactivation of neutrophils will further exacerbate tissue damage. The number of circulating neutrophils is strongly correlated with the infarct size [145]. MI rapidly mobilizes neutrophils from the spleen into the peripheral blood, and one of the drivers of this process is vascular cell adhesion molecule-1 in exosomes secreted by injured ECs [123]. This study reinforces the idea that exosomes can strongly influence immune responses to MI.
Exosomes derived from other cardiac cells
Other types of cardiac cells also secrete exosomes potentially useful for treating MI. For example, exosomes from cardiac resident progenitor cells contain pregnancy-associated plasma protein-A (PPAP-A) that promotes the release of IGF-1 and phosphorylation of AKT and ERK1/2, while reducing caspase activation, preventing cardiomyocyte apoptosis and enhancing ventricular function after I/R injury [126]. In fact, such exosomes can activate multiple signaling pathways to promote cardiomyocyte migration, survival and proliferation, as well as stimulate angiogenesis directly and indirectly through monocytes, thereby promoting tissue repair and regeneration in multiple ways [127].
Cardiac telocytes are cardiac interstitial cells in the cardiac mesenchyme that help guide myocardial development and that can promote myocardial regeneration after infarction. These cells secrete exosomes that can mitigate injury after MI, such as by carrying miRNA-21-5p, which downregulates cell death inducing protein 1 (Cdip1) gene to inhibit apoptosis of cardiac microvascular ECs and promote cardiac angiogenesis [128, 146].
Exosomes from cardiomyocytes can influence the proliferation and differentiation of CFs [112], and the converse is true: CFs can affect cardiomyocytes after I/R injury via a paracrine pathway [147], such as by delivering miR-133a, which downregulates ELAV-like RNA-binding protein 1 (ELAVL1), thereby inhibiting pyroptosis [129].
Although epicardial cells have rarely been studied as a source of exosomes, probably because these cells are largely quiescent in adults [148], exosomes from these cells can promote cardiomyocyte proliferation after myocardial injury [130]. These findings highlight the danger of neglecting “unremarkable” intracardial cell populations that may take on important therapeutic functions after MI. Future studies should explore the potential therapeutic usefulness of these exosomes.
Application forms of exosomes in MI therapy
Data from several preclinical studies suggest that exosomes have advantages over cell-based therapies in terms of safety, low immunogenicity, and ease of storage, but there are still some inadequacies to be addressed [37]. Short half-life and low retention rate are among the drawbacks of direct exosome injection. Therefore, advanced engineering techniques are required to functionalize exosomes, enhancing their efficiency and potency as therapeutics for MI. Currently, the main forms of preclinical research applications of exosomes include using biocompatible and biodegradable delivery systems to help deliver exosomes or engineering exosomes to enhance their therapeutic capability.
Exosome co-delivery system for application
The application of delivery systems can overcome the issue of exosomes' low tissue residence time and provide a controlled release system to maintain their biological activity, thereby improving therapeutic effects on the infarcted myocardium. The mechanical properties, biocompatibility, biostability, and unique maneuverability of biomaterials make them the optimal choice for delivery systems [149]. Delivery methods typified by hydrogels have demonstrated significant therapeutic potential in laboratory settings and preclinical studies [150,151,152,153,154]. Researchers have utilized biomaterials as a foundation to better address the complex physiological environment of the heart following MI by chemically modifying, connecting self-assembling peptides, adding therapeutic molecules, and other approaches. These strategies enhance the accumulation of exosome-based delivery systems at disease sites, ultimately improving therapeutic efficacy. We summarize the current delivery systems for MI therapy, focusing on MSC and cardiac cell-derived exosomes (Table 4). Notably, both Zhu and Cheng et al. demonstrated the safety, efficacy, and clinical feasibility of minimally invasive intrapericardial injections in a clinically relevant porcine model and further explored the potential for treatment in humans [155, 156].
By combining with other therapeutic modalities, exosomes can be developed for a wider range of applications. Given their ability to regulate cellular function directly and effectively, exosomes-based delivery systems are currently commonly used in both drug carrier and therapeutic agents. As mentioned earlier, sequential administration of exosomes and BMSCs has shown improved recruitment and survival of BMSCs, providing a promising strategy for the co-delivery of exosomes and living cells in synergistic therapy for heart-related disease [75]. Similarly, Chachques et al. utilized the therapeutic properties of exosomes by seeding MSC-derived exosomes along with MSCs and macrophages into an elastomeric cardiowrap scaffold [162]. The results demonstrated that exosomes significantly enhanced the wound healing properties of MSCs and shifted macrophages towards an M2 phenotype. This suggests that exosomes enable the implementation of multi-mechanism therapeutic strategies by influencing multiple cellular functions, representing another promising direction for exosome-based delivery systems.
In future exosome research, there is a need for further exploration of the development of exosome-based delivery systems for disease therapy, the determination of optimal release periods for exosomes, and ways to improve loading efficiency, among other issues.
Engineering exosomes for drug delivery
Exosome engineering technology not only enables exosomes to act as natural nanocarriers for delivering molecular drugs but also enhances the target specificity of exosomes through surface modifications, making them more clinically applicable.
Drug loading strategies can be divided into two categories: endogenous (collecting drug-laden exosomes after pretreatment of parental cells) and exogenous (directly loading drugs into isolated exosomes) [163]. The former allows exosomes to be loaded with drugs at the cellular level, resulting in stronger therapeutic effects [164,165,166,167]. However, exosomes obtained through pretreatment usually exhibit relatively low drug loading efficiency and may have uneven drug levels [163]. In contrast, exogenous strategies provide better control over drug loading and have also been proven effective in animal models [168, 169]. Various drug loading methods have been developed and are described in detail in the review by Xi et al. [170]. The effectiveness of exosomes as drug carriers in preclinical studies has been initially established, and further validation of long-term safety should be conducted in the future. To achieve maximum drug loading efficiency, it’s suggested to coat exosome to metal organic framework or inorganic nanoparticles, both of which possess high drug loading efficiency [171].
Surface modification can enhance the specific targeting property and residence time of exosomes in cardiac tissues, thus improving the issue of exosomes diffusing out of the infarcted heart following direct injection and avoiding off-target effects. For example, Lee et al. used iron oxide nanoparticles to pretreat BMSCs, followed by serial cell extrusion, resulting in exosome-mimetic nanovesicles with enhanced therapeutic molecules and higher targeting efficiency [172]. Magnetic guidance further increased their retention in the infarcted hearts of MI rats. Similarly, exosomes can be modified either through genetic engineering or direct modification to bind cardiac homing peptides on their surface, enabling them to specifically target and treat ischemic myocardium when administered via tail vein injection [173, 174]. Intravenous delivery is expected to avoid the trauma caused by myocardial injection and provides a new direction for exosome delivery. Additionally, surface modifications can ameliorate the issue of circulating clearance of exosomes by immune cells due to intravenous injection. Exosomes incorporating CD47 protein on the surface can be protected from phagocytosis of macrophages and monocytes and have a longer half-life in circulation [175]. These studies suggest that exosomes can be delivered in a more convenient, safe and feasible way, offering broader prospects for clinical applications of exosomes. From the current perspective, the development of engineering exosomes is still in its infancy, with high costs for translational research and clinical applications.
Conclusions and outlook
The causes affecting the prognosis of MI are diverse, but a fundamental problem is the irreversible loss of cardiomyocytes and the formation of scar tissue. Also, dysregulation of immune pathways, impaired suppression of postinfarction inflammation, disturbed spatial inhibition of the inflammatory response, and overactive fibrosis can contribute to poor remodeling of the heart, which may eventually lead to serious consequences such as heart failure [62]. Stem cell therapy has opened up new perspectives for treating MI, but cells transplanted into the ischemic heart often show poor recruitment and survival [75]. A more effective alternative may be delivering the therapeutic components of exosomes secreted by these cells. This review highlights the diverse range of exosome sources and contents that can mitigate injury, promote tissue repair, and regenerate after MI.
Preclinical evidence of exosome efficacy in animal models of MI is abundant [104, 176], with MSC-derived exosomes showing strong potential in many studies [177]. Although most exosome research is preclinical, there have been clinical trials that have initially verified the safety of exosomes, proving that exosomes purified by current methods may already be safe to use in patients. For example, injecting exosomes from MSCs into the brain parenchyma of five patients with ischemic stroke did not lead to serious adverse effects within 3-month follow-up [178]. The only clinical trial using MSC-derived exosomes for MI treatment is still under recruitment (NCT05669144), and further longitudinal clinical studies are necessary to confirm the safety and efficacy of exosome therapy.
In the case of exosomes secreted by cardiac cells, their cargo can depend on the pathological state of the cells. This characteristic could potentially make them useful as biomarkers for timely diagnosis, staging, and monitoring of disease during treatment. For instance, multiple studies have shown that miR-208a in exosomes released from injured cardiomyocytes after MI has the advantages of strong myocardial specificity, high sensitivity and strong stability, and is positively correlated with the indexes of myocardial function before and after therapy [112, 179,180,181]. The functional diversity of exosomes secreted by cardiac cells also suggests that these various cell types should be further explored as potential sources of therapeutic exosomes.
Intensive research on the molecular content and biological function of exosomes has facilitated the development of various engineering approaches. These methods have enhanced the targeting or therapeutic benefits of exosomes, but most engineering methods are only suitable for small-scale laboratory use. To make exosomes suitable for clinical therapeutics, several challenges still need to be addressed. Firstly, current techniques to isolate exosomes are not compatible with large-scale production for the clinic. Appropriate production methods and quality standards need to be established for exosome purification and characterization to ensure efficacy, safety and cost-effectiveness. Secondly, the low yield of exosomes makes it challenging to achieve industrialization, which also puts forward higher requirements for their cellular source. The cellular sources of exosomes should undergo differentiation and division as little as possible, so genetic engineering should be explored to modify existing cell types, such as the recent immortalization of MSCs [182]. Another potential source of therapeutic exosomes is plants, as illustrated in the ability of exosome-like nanoparticles from garlic to reduce obesity in a mouse model [183]. Genetic engineering should also be explored for creating exosomes that target specific tissues within the target organ(s), reducing their accumulation in off-target tissues [174].
Exosomes derived from MSCs and cardiac cells appear poised to play an important role in treating MI and potentially other cardiovascular diseases in the future. The functional diversity of cardiac cells and the ability of their exosomes to either mitigate or exacerbate myocardial injury highlight the need for future research to investigate the specific effects of exosome cargo on particular signaling pathways. Additionally, future studies should explore methods for pretreating or genetically modifying parental cells in order to optimize the therapeutic effects of their exosomes.
Availability of data and materials
Not applicable.
Abbreviations
- ADAMTS16:
-
A disintegrin and metalloproteinase with thrombospondin 16
- ADMSCs:
-
Adipose mesenchymal stem cells
- AIFM3:
-
Apoptosis-inducing factor, mitochondrion-associated 3
- AKT:
-
Protein kinase B
- ALIX:
-
Apoptosis-linked gene 2-interacting protein X
- AMI:
-
Acute myocardial infarction
- AMPK:
-
Adenosine 5 ‘-monophosphate (AMP)-activated protein kinase
- Apaf1:
-
Apoptotic protease activating factor- 1
- ATF2:
-
Activating transcription factor 2
- Bak1:
-
B-cell lymphoma-2 antagonist/killer 1
- Bax:
-
B-cell lymphoma-associated X
- BMSCs:
-
Bone marrow mesenchymal stem cells
- CDCs:
-
Cardiosphere-derived cells
- Cdip1:
-
Cell death inducing protein
- CFs:
-
Cardiac fibroblasts
- CHK2:
-
Checkpoint kinase 2
- DMBT1:
-
Deleted in malignant brain tumors 1
- DMT1:
-
Recombinant divalent metal transporter 1
- Dyrk2:
-
Dual specificity tyrosine (Y) phosphorylation regulated kinase 2
- ECs:
-
Endothelial cells
- ELAVL1:
-
ELAV-like RNA-binding protein 1
- ELF5:
-
E74-like factor 5
- EPCs:
-
Endothelial progenitor cells
- ERK:
-
Extracellular regulated protein kinase
- EVs:
-
Extracellular vesicles
- EZH2:
-
Enhancer of zeste 2 polycomb repressive complex 2
- Fasl:
-
Factor related apoptosis ligand
- FNDC5:
-
Fibronectin type III domain-containing protein 5
- FOXO1:
-
Forkhead box O1
- FOXO3:
-
Forkhead box O3
- GSDMD:
-
Gasdermin D
- GSK3 β:
-
Glycogen synthase kinase 3β
- HIF-1 α:
-
Hypoxia-inducible factor 1
- HMGA2:
-
High mobility group AT-hook 2
- HO-1:
-
Heme oxygenase 1
- I/R:
-
Ischemia/Reperfusion
- IGF-1:
-
Insulin-like growth factor- 1
- IL:
-
Interleukin
- JMY:
-
Regulatory protein
- JNK:
-
C-Jun N-terminal kinase
- MAP3K2:
-
Mitogen-activated protein kinase kinase kinase 2
- MAPK:
-
Mitogen-activated protein kinase
- MEKK1:
-
Mitogen-activated protein kinase kinase kinase 1
- MI:
-
Myocardial infarction
- miRNA:
-
Micro RNA
- MKK4:
-
Mitogen-activated protein kinase kinase 4
- MSCs:
-
Mesenchymal stem cells
- MVB:
-
Multivesicular body
- ncRNA:
-
Non-coding RNA
- NF-κB:
-
Nuclear factor-κB
- NLRP3:
-
NOD-like receptor thermal protein domain associated protein 3
- Nrf2:
-
Nuclear factor erythroid 2-related factor 2
- p-AKT:
-
Phosphorylated protein kinase B
- PAPP-A:
-
Pregnancy-associated plasma protein-A
- Pdcd4:
-
Programmed cell death 4
- PDGFR- β:
-
Platelet-derived growth factor receptor-β
- PFN2:
-
Profilin 2
- PI3K:
-
Phosphatidylinositol3-kinase
- PKC δ:
-
Protein Kinase C δ
- PP2A:
-
Protein phospholipase 2A
- Pten:
-
Phosphatase and tensin homolog
- RAS:
-
Renin-angiotensin system
- ROS:
-
Reactive oxygen species
- S1P:
-
Sphingosine 1-phosphate
- S1PR1:
-
Sphingosine-1-phosphate receptor 1
- Sirt1:
-
Sirtuin 1
- SK1:
-
Sphingosine kinase 1
- Smad:
-
Mothers against decapentaplegic homolog
- SOCS2:
-
Suppressor of cytokine signaling 2
- SOX6:
-
SRY (sex determining region Y)-box 6
- SP1:
-
Specificity protein 1
- SRSF1:
-
Serine and arginine rich splicing factor 1
- STAT5B:
-
Signal transducer and activator of transcription 5B
- TGFBR2:
-
Transforming growth factor-beta receptor type 2
- TGF-β1:
-
Transforming growth factor-beta 1
- TIMP2:
-
Tissue inhibitor of metalloproteinase 2
- TLR4:
-
Toll-like receptor 4
- TNF:
-
Tumor necrosis factor
- TSG101:
-
Tumor susceptibility gene 101
- UMSCs:
-
Umbilical cord mesenchymal stem cells
- VCAM-1:
-
Vascular cell adhesion molecule 1
- VEGF:
-
Vascular endothlial growth factor
- VEGFA:
-
Vascular endothlial growth factor A
- XIAP:
-
X-linked inhibitor of apoptosis protein
References
Wang W, Zheng H. Myocardial Infarction: the protective role of mirnas in myocardium pathology. Front Cardiovasc Med. 2021;8: 631817.
Mozaffarian D, Benjamin EJ, Go AS, Arnett DK, Blaha MJ, Cushman M, et al. Heart disease and stroke statistics-2016 update: a report from the american heart association. Circulation. 2016;133(4):e38-360.
Wu X, Iroegbu CD, Peng J, Guo J, Yang J, Fan C. Cell death and exosomes regulation after myocardial infarction and ischemia-reperfusion. Front Cell Dev Biol. 2021;9: 673677.
Neri M, Riezzo I, Pascale N, Pomara C, Turillazzi E. Ischemia/Reperfusion injury following acute myocardial infarction: a critical issue for clinicians and forensic pathologists. Mediators Inflamm. 2017;2017:7018393.
Braunwald E, Kloner RA. Myocardial reperfusion: a double-edged sword? J Clin Invest. 1985;76(5):1713–9.
Cui M, Han Y, Yang J, Li G, Yang C. A narrative review of the research status of exosomes in cardiovascular disease. Ann Palliat Med. 2022;11(1):363–77.
Shafei AE, Ali MA, Ghanem HG, Shehata AI, Abdelgawad AA, Handal HR, et al. Mesenchymal stem cell therapy: A promising cell-based therapy for treatment of myocardial infarction. J Gene Med. 2017;19(12).
Zou L, Ma X, Lin S, Wu B, Chen Y, Peng C. Bone marrow mesenchymal stem cell-derived exosomes protect against myocardial infarction by promoting autophagy. Exp Ther Med. 2019;18(4):2574–82.
Wang XL, Zhao YY, Sun L, Shi Y, Li ZQ, Zhao XD, et al. Exosomes derived from human umbilical cord mesenchymal stem cells improve myocardial repair via upregulation of Smad7. Int J Mol Med. 2018;41(5):3063–72.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478).
Lin J, Li J, Huang B, Liu J, Chen X, Chen XM, et al. Exosomes: novel biomarkers for clinical diagnosis. ScientificWorldJournal. 2015;2015: 657086.
Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–78.
Colombo M, Raposo G, Théry C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol. 2014;30:255–89.
Hade MD, Suire CN, Suo Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells. 2021;10(8).
Gonda A, Kabagwira J, Senthil GN, Wall NR. Internalization of Exosomes through Receptor-Mediated Endocytosis. Mol Cancer Res. 2019;17(2):337–47.
McKelvey KJ, Powell KL, Ashton AW, Morris JM, McCracken SA. Exosomes: mechanisms of uptake. J Circ Biomark. 2015;4:7.
Mulcahy LA, Pink RC, Carter DR. Routes and mechanisms of extracellular vesicle uptake. J Extracell Vesicles. 2014;3.
Ginini L, Billan S, Fridman E, Gil Z. Insight into Extracellular Vesicle-Cell Communication: From Cell Recognition to Intracellular Fate. Cells. 2022;11(9).
Chanteloup G, Cordonnier M, Isambert N, Bertaut A, Hervieu A, Hennequin A, et al. Monitoring HSP70 exosomes in cancer patients’ follow up: a clinical prospective pilot study. J Extracell Vesicles. 2020;9(1):1766192.
Wubbolts R, Leckie RS, Veenhuizen PT, Schwarzmann G, Möbius W, Hoernschemeyer J, et al. Proteomic and biochemical analyses of human B cell-derived exosomes. Potential implications for their function and multivesicular body formation. J Biol Chem. 2003;278(13):10963–72.
Roucourt B, Meeussen S, Bao J, Zimmermann P, David G. Heparanase activates the syndecan-syntenin-ALIX exosome pathway. Cell Res. 2015;25(4):412–28.
Simons M, Raposo G. Exosomes–vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21(4):575–81.
He C, Zheng S, Luo Y, Wang B. Exosome theranostics: biology and translational medicine. Theranostics. 2018;8(1):237–55.
Duan L, Xu L, Xu X, Qin Z, Zhou X, Xiao Y, et al. Exosome-mediated delivery of gene vectors for gene therapy. Nanoscale. 2021;13(3):1387–97.
Han QF, Li WJ, Hu KS, Gao J, Zhai WL, Yang JH, et al. Exosome biogenesis: machinery, regulation, and therapeutic implications in cancer. Mol Cancer. 2022;21(1):207.
Weissman IL. Stem cells: units of development, units of regeneration, and units in evolution. Cell. 2000;100(1):157–68.
Segers VF, Lee RT. Stem-cell therapy for cardiac disease. Nature. 2008;451(7181):937–42.
Fridenshteĭn A, Piatetskiĭ S II, Petrakova KV. Osteogenesis in transplants of bone marrow cells. Arkh Anat Gistol Embriol. 1969;56(3):3–11.
Bieback K, Kern S, Klüter H, Eichler H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells. 2004;22(4):625–34.
Hass R, Kasper C, Böhm S, Jacobs R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun Signal. 2011;9:12.
Pittenger MF, Martin BJ. Mesenchymal stem cells and their potential as cardiac therapeutics. Circ Res. 2004;95(1):9–20.
Shafei AE, Ali MA, Ghanem HG, Shehata AI, Abdelgawad AA, Handal HR, et al. Mechanistic effects of mesenchymal and hematopoietic stem cells: New therapeutic targets in myocardial infarction. J Cell Biochem. 2018;119(7):5274–86.
Moghaddam AS, Afshari JT, Esmaeili SA, Saburi E, Joneidi Z, Momtazi-Borojeni AA. Cardioprotective microRNAs: lessons from stem cell-derived exosomal microRNAs to treat cardiovascular disease. Atherosclerosis. 2019;285:1–9.
Xiao Y, Zhang Y, Li Y, Peng N, Liu Q, Qiu D, et al. Exosomes derived from mesenchymal stem cells pretreated with ischemic rat heart extracts promote angiogenesis via the delivery of DMBT1. Cell Transplant. 2022;31:9636897221102898.
Cui X, He Z, Liang Z, Chen Z, Wang H, Zhang J. Exosomes from adipose-derived mesenchymal stem cells protect the myocardium against ischemia/reperfusion injury through Wnt/β-catenin signaling pathway. J Cardiovasc Pharmacol. 2017;70(4):225–31.
Xu H, Wang Z, Liu L, Zhang B, Li B. Exosomes derived from adipose tissue, bone marrow, and umbilical cord blood for cardioprotection after myocardial infarction. J Cell Biochem. 2020;121(3):2089–102.
Sun SJ, Wei R, Li F, Liao SY, Tse HF. Mesenchymal stromal cell-derived exosomes in cardiac regeneration and repair. Stem Cell Reports. 2021;16(7):1662–73.
Miao C, Lei M, Hu W, Han S, Wang Q. A brief review: the therapeutic potential of bone marrow mesenchymal stem cells in myocardial infarction. Stem Cell Res Ther. 2017;8(1):242.
Amani H, Habibey R, Hajmiresmail SJ, Latifi S, Pazoki-Toroudi H, Akhavan O. Antioxidant nanomaterials in advanced diagnoses and treatments of ischemia reperfusion injuries. J Mater Chem B. 2017;5(48):9452–76.
Wu Z, Cheng S, Wang S, Li W, Liu J. BMSCs-derived exosomal microRNA-150-5p attenuates myocardial infarction in mice. Int Immunopharmacol. 2021;93: 107389.
Mao S, Zhao J, Zhang ZJ, Zhao Q. MiR-183-5p overexpression in bone mesenchymal stem cell-derived exosomes protects against myocardial ischemia/reperfusion injury by targeting FOXO1. Immunobiology. 2022;227(3): 152204.
Luther KM, Haar L, McGuinness M, Wang Y, Lynch Iv TL, Phan A, et al. Exosomal miR-21a-5p mediates cardioprotection by mesenchymal stem cells. J Mol Cell Cardiol. 2018;119:125–37.
Peng Y, Zhao JL, Peng ZY, Xu WF, Yu GL. Exosomal miR-25-3p from mesenchymal stem cells alleviates myocardial infarction by targeting pro-apoptotic proteins and EZH2. Cell Death Dis. 2020;11(5):317.
Zhu LP, Tian T, Wang JY, He JN, Chen T, Pan M, et al. Hypoxia-elicited mesenchymal stem cell-derived exosomes facilitates cardiac repair through miR-125b-mediated prevention of cell death in myocardial infarction. Theranostics. 2018;8(22):6163–77.
Toldo S, Mauro AG, Cutter Z, Abbate A. Inflammasome, pyroptosis, and cytokines in myocardial ischemia-reperfusion injury. Am J Physiol Heart Circ Physiol. 2018;315(6):H1553–68.
Yue R, Lu S, Luo Y, Zeng J, Liang H, Qin D, et al. Mesenchymal stem cell-derived exosomal microRNA-182-5p alleviates myocardial ischemia/reperfusion injury by targeting GSDMD in mice. Cell Death Discov. 2022;8(1):202.
Sun C, Li W, Li Y, Chen J, An H, Zeng G, et al. MiR-182-5p mediated by exosomes derived from bone marrow mesenchymal stem cell attenuates inflammatory responses by targeting TLR4 in a Mouse Model of Myocardial Infraction. Immune Netw. 2022;22(6): e49.
Jiao W, Hao J, Xie Y, Meng M, Gao W. EZH2 mitigates the cardioprotective effects of mesenchymal stem cell-secreted exosomes against infarction via HMGA2-mediated PI3K/AKT signaling. BMC Cardiovasc Disord. 2022;22(1):95.
Cheng H, Chang S, Xu R, Chen L, Song X, Wu J, et al. Hypoxia-challenged MSC-derived exosomes deliver miR-210 to attenuate post-infarction cardiac apoptosis. Stem Cell Res Ther. 2020;11(1):224.
Fu DL, Jiang H, Li CY, Gao T, Liu MR, Li HW. MicroRNA-338 in MSCs-derived exosomes inhibits cardiomyocyte apoptosis in myocardial infarction. Eur Rev Med Pharmacol Sci. 2020;24(19):10107–17.
Wang Y, Shen Y. Exosomal miR-455-3p from BMMSCs prevents cardiac ischemia-reperfusion injury. Hum Exp Toxicol. 2022;41:9603271221102508.
Chen G, Wang M, Ruan Z, Zhu L, Tang C. Mesenchymal stem cell-derived exosomal miR-143-3p suppresses myocardial ischemia-reperfusion injury by regulating autophagy. Life Sci. 2021;280: 119742.
Mao Q, Liang XL, Zhang CL, Pang YH, Lu YX. LncRNA KLF3-AS1 in human mesenchymal stem cell-derived exosomes ameliorates pyroptosis of cardiomyocytes and myocardial infarction through miR-138-5p/Sirt1 axis. Stem Cell Res Ther. 2019;10(1):393.
Yu H, Pan Y, Dai M, Wang X, Chen H. Mesenchymal Stem Cell-Originated Exosomal Lnc A2M-AS1 Alleviates Hypoxia/Reperfusion-Induced Apoptosis and Oxidative Stress in Cardiomyocytes. Cardiovasc Drugs Ther. 2022.
Li KS, Bai Y, Li J, Li SL, Pan J, Cheng YQ, et al. LncRNA HCP5 in hBMSC-derived exosomes alleviates myocardial ischemia reperfusion injury by sponging miR-497 to activate IGF1/PI3K/AKT pathway. Int J Cardiol. 2021;342:72–81.
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta R, Zaitsev EM, Gleason CE, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–72.
Zhang JK, Zhang Z, Guo ZA, Fu Y, Chen XJ, Chen WJ, et al. The BMSC-derived exosomal lncRNA Mir9-3hg suppresses cardiomyocyte ferroptosis in ischemia-reperfusion mice via the Pum2/PRDX6 axis. Nutr Metab Cardiovasc Dis. 2022;32(2):515–27.
Sun J, Shen H, Shao L, Teng X, Chen Y, Liu X, et al. HIF-1α overexpression in mesenchymal stem cell-derived exosomes mediates cardioprotection in myocardial infarction by enhanced angiogenesis. Stem Cell Res Ther. 2020;11(1):373.
Wang X, Bai L, Liu X, Shen W, Tian H, Liu W, et al. Cardiac microvascular functions improved by MSC-derived exosomes attenuate cardiac fibrosis after ischemia-reperfusion via PDGFR-β modulation. Int J Cardiol. 2021;344:13–24.
Sun L, Zhu W, Zhao P, Zhang J, Lu Y, Zhu Y, et al. Down-regulated exosomal microRNA-221 - 3p derived from senescent mesenchymal stem cells impairs heart repair. Front Cell Dev Biol. 2020;8:263.
Kesavardhana S, Malireddi RKS, Kanneganti TD. Caspases in cell death, inflammation, and pyroptosis. Annu Rev Immunol. 2020;38:567–95.
Xiong YY, Gong ZT, Tang RJ, Yang YJ. The pivotal roles of exosomes derived from endogenous immune cells and exogenous stem cells in myocardial repair after acute myocardial infarction. Theranostics. 2021;11(3):1046–58.
de Couto G, Liu W, Tseliou E, Sun B, Makkar N, Kanazawa H, et al. Macrophages mediate cardioprotective cellular postconditioning in acute myocardial infarction. J Clin Invest. 2015;125(8):3147–62.
Ben-Mordechai T, Palevski D, Glucksam-Galnoy Y, Elron-Gross I, Margalit R, Leor J. Targeting macrophage subsets for infarct repair. J Cardiovasc Pharmacol Ther. 2015;20(1):36–51.
Xu R, Zhang F, Chai R, Zhou W, Hu M, Liu B, et al. Exosomes derived from pro-inflammatory bone marrow-derived mesenchymal stem cells reduce inflammation and myocardial injury via mediating macrophage polarization. J Cell Mol Med. 2019;23(11):7617–31.
Ning H, Chen H, Deng J, Xiao C, Xu M, Shan L, et al. Exosomes secreted by FNDC5-BMMSCs protect myocardial infarction by anti-inflammation and macrophage polarization via NF-κB signaling pathway and Nrf2/HO-1 axis. Stem Cell Res Ther. 2021;12(1):519.
Kore RA, Wang X, Ding Z, Griffin RJ, Tackett AJ, Mehta JL. MSC exosome-mediated cardioprotection in ischemic mouse heart comparative proteomics of infarct and peri-infarct areas. Mol Cell Biochem. 2021;476(4):1691–704.
Liu HY, Yu LF, Zhou TG, Wang YD, Sun DH, Chen HR, et al. Lipopolysaccharide-stimulated bone marrow mesenchymal stem cells-derived exosomes inhibit H2O2-induced cardiomyocyte inflammation and oxidative stress via regulating miR-181a-5p/ATF2 axis. Eur Rev Med Pharmacol Sci. 2020;24(19):10069–77.
Li Y, Zhou J, Zhang O, Wu X, Guan X, Xue Y, et al. Bone marrow mesenchymal stem cells-derived exosomal microRNA-185 represses ventricular remolding of mice with myocardial infarction by inhibiting SOCS2. Int Immunopharmacol. 2020;80: 106156.
Zheng J, Zhang X, Cai W, Yang Y, Guo T, Li J, et al. Bone marrow mesenchymal stem cell-derived exosomal microRNA-29b-3p promotes angiogenesis and ventricular remodeling in rats with myocardial infarction by targeting ADAMTS16. Cardiovasc Toxicol. 2022;22(8):689–700.
Sun Y. Intracardiac renin-angiotensin system and myocardial repair/remodeling following infarction. J Mol Cell Cardiol. 2010;48(3):483–9.
Xiao M, Zeng W, Wang J, Yao F, Peng Z, Liu G, et al. Exosomes protect against acute myocardial infarction in rats by regulating the renin-angiotensin system. Stem Cells Dev. 2021;30(12):622–31.
Anderson JL, Morrow DA. Acute myocardial infarction. N Engl J Med. 2017;376(21):2053–64.
Burchill LJ, Velkoska E, Dean RG, Griggs K, Patel SK, Burrell LM. Combination renin-angiotensin system blockade and angiotensin-converting enzyme 2 in experimental myocardial infarction: implications for future therapeutic directions. Clin Sci (Lond). 2012;123(11):649–58.
Huang P, Wang L, Li Q, Xu J, Xu J, Xiong Y, et al. Combinatorial treatment of acute myocardial infarction using stem cells and their derived exosomes resulted in improved heart performance. Stem Cell Res Ther. 2019;10(1):300.
Woo DH, Hwang HS, Shim JH. Comparison of adult stem cells derived from multiple stem cell niches. Biotechnol Lett. 2016;38(5):751–9.
Frese L, Dijkman PE, Hoerstrup SP. Adipose tissue-derived stem cells in regenerative medicine. Transfus Med Hemother. 2016;43(4):268–74.
Wang X, Zhu Y, Wu C, Liu W, He Y, Yang Q. Adipose-derived mesenchymal stem cells-derived exosomes carry microRNA-671 to alleviate myocardial infarction through inactivating the TGFBR2/Smad2 Axis. Inflammation. 2021;44(5):1815–30.
Zhang Q, Wang L, Wang S, Cheng H, Xu L, Pei G, et al. Signaling pathways and targeted therapy for myocardial infarction. Signal Transduct Target Ther. 2022;7(1):78.
Sun S, Wu Y, Maimaitijiang A, Huang Q, Chen Q. Ferroptotic cardiomyocyte-derived exosomes promote cardiac macrophage M1 polarization during myocardial infarction. PeerJ. 2022;10: e13717.
Deng S, Zhou X, Ge Z, Song Y, Wang H, Liu X, et al. Exosomes from adipose-derived mesenchymal stem cells ameliorate cardiac damage after myocardial infarction by activating S1P/SK1/S1PR1 signaling and promoting macrophage M2 polarization. Int J Biochem Cell Biol. 2019;114: 105564.
Wu X, Reboll MR, Korf-Klingebiel M, Wollert KC. Angiogenesis after acute myocardial infarction. Cardiovasc Res. 2021;117(5):1257–73.
Zhu D, Wang Y, Thomas M, McLaughlin K, Oguljahan B, Henderson J, et al. Exosomes from adipose-derived stem cells alleviate myocardial infarction via microRNA-31/FIH1/HIF-1α pathway. J Mol Cell Cardiol. 2022;162:10–9.
Akyurekli C, Le Y, Richardson RB, Fergusson D, Tay J, Allan DS. A systematic review of preclinical studies on the therapeutic potential of mesenchymal stromal cell-derived microvesicles. Stem Cell Rev Rep. 2015;11(1):150–60.
Heo JS, Choi Y, Kim HS, Kim HO. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int J Mol Med. 2016;37(1):115–25.
Huang L, Yang L, Ding Y, Jiang X, Xia Z, You Z. Human umbilical cord mesenchymal stem cells-derived exosomes transfers microRNA-19a to protect cardiomyocytes from acute myocardial infarction by targeting SOX6. Cell Cycle. 2020;19(3):339–53.
Li CX, Song J, Li X, Zhang T, Li ZM. Circular RNA 0001273 in exosomes derived from human umbilical cord mesenchymal stem cells (UMSCs) in myocardial infarction. Eur Rev Med Pharmacol Sci. 2020;24(19):10086–95.
Diao L, Zhang Q. Transfer of lncRNA UCA1 by hUCMSCs-derived exosomes protects against hypoxia/reoxygenation injury through impairing miR-143-targeted degradation of Bcl-2. Aging (Albany NY). 2021;13(4):5967–85.
Sun L, Zhu W, Zhao P, Wang Q, Fan B, Zhu Y, et al. Long noncoding RNA UCA1 from hypoxia-conditioned hMSC-derived exosomes: a novel molecular target for cardioprotection through miR-873-5p/XIAP axis. Cell Death Dis. 2020;11(8):696.
Liang C, Liu Y, Xu H, Huang J, Shen Y, Chen F, et al. Exosomes of human umbilical cord mscs protect against hypoxia/reoxygenation-induced pyroptosis of cardiomyocytes via the miRNA-100-5p/FOXO3/NLRP3 pathway. Front Bioeng Biotechnol. 2020;8: 615850.
Song Y, Wang B, Zhu X, Hu J, Sun J, Xuan J, et al. Human umbilical cord blood-derived MSCs exosome attenuate myocardial injury by inhibiting ferroptosis in acute myocardial infarction mice. Cell Biol Toxicol. 2021;37(1):51–64.
Furtado MB, Costa MW, Rosenthal NA. The cardiac fibroblast: origin, identity and role in homeostasis and disease. Differentiation. 2016;92(3):93–101.
Xu Y, Wu A, Chen J, Song X, Chen M, Liu Q. Limb-Bud and Heart (LBH) upregulation in cardiomyocytes under hypoxia promotes the activation of cardiac fibroblasts via exosome secretion. Mediators Inflamm. 2022;2022:8939449.
Shinde AV, Frangogiannis NG. Fibroblasts in myocardial infarction: a role in inflammation and repair. J Mol Cell Cardiol. 2014;70:74–82.
Shi Y, Yang Y, Guo Q, Gao Q, Ding Y, Wang H, et al. Exosomes derived from human umbilical cord mesenchymal stem cells promote fibroblast-to-myofibroblast differentiation in inflammatory environments and benefit cardioprotective effects. Stem Cells Dev. 2019;28(12):799–811.
Zhu D, Liu S, Huang K, Wang Z, Hu S, Li J, et al. Intrapericardial exosome therapy dampens cardiac injury via activating Foxo3. Circ Res. 2022;131(10):e135–50.
Vanhoutte D, Heymans S. TIMPs and cardiac remodeling: “Embracing the MMP-independent-side of the family.” J Mol Cell Cardiol. 2010;48(3):445–53.
Ni J, Liu X, Yin Y, Zhang P, Xu YW, Liu Z. Exosomes derived from TIMP2-modified human umbilical cord mesenchymal stem cells enhance the repair effect in rat model with myocardial infarction possibly by the Akt/Sfrp2 pathway. Oxid Med Cell Longev. 2019;2019:1958941.
Zhang N, Zhu J, Ma Q, Zhao Y, Wang Y, Hu X, et al. Exosomes derived from human umbilical cord MSCs rejuvenate aged MSCs and enhance their functions for myocardial repair. Stem Cell Res Ther. 2020;11(1):273.
Hamid T, Xu Y, Ismahil MA, Rokosh G, Jinno M, Zhou G, et al. Cardiac mesenchymal stem cells promote fibrosis and remodeling in heart failure: role of PDGF signaling. JACC Basic Transl Sci. 2022;7(5):465–83.
Klopsch C, Skorska A, Ludwig M, Gaebel R, Lemcke H, Kleiner G, et al. Cardiac mesenchymal stem cells proliferate early in the ischemic heart. Eur Surg Res. 2017;58(5–6):341–53.
Klopsch C, Skorska A, Ludwig M, Lemcke H, Maass G, Gaebel R, et al. Intramyocardial angiogenetic stem cells and epicardial erythropoietin save the acute ischemic heart. Dis Model Mech. 2018;11(6).
Ruan XF, Li YJ, Ju CW, Shen Y, Lei W, Chen C, et al. Exosomes from Suxiao Jiuxin pill-treated cardiac mesenchymal stem cells decrease H3K27 demethylase UTX expression in mouse cardiomyocytes in vitro. Acta Pharmacol Sin. 2018;39(4):579–86.
Ju C, Shen Y, Ma G, Liu Y, Cai J, Kim IM, et al. Transplantation of cardiac mesenchymal stem cell-derived exosomes promotes repair in ischemic myocardium. J Cardiovasc Transl Res. 2018;11(5):420–8.
Park M, Shen YT, Gaussin V, Heyndrickx GR, Bartunek J, Resuello RR, et al. Apoptosis predominates in nonmyocytes in heart failure. Am J Physiol Heart Circ Physiol. 2009;297(2):H785–91.
Davidson SM, Riquelme JA, Zheng Y, Vicencio JM, Lavandero S, Yellon DM. Endothelial cells release cardioprotective exosomes that may contribute to ischaemic preconditioning. Sci Rep. 2018;8(1):15885.
Bang C, Batkai S, Dangwal S, Gupta SK, Foinquinos A, Holzmann A, et al. Cardiac fibroblast-derived microRNA passenger strand-enriched exosomes mediate cardiomyocyte hypertrophy. J Clin Invest. 2014;124(5):2136–46.
Huang J, Wang F, Sun X, Chu X, Jiang R, Wang Y, et al. Myocardial infarction cardiomyocytes-derived exosomal miR-328-3p promote apoptosis via Caspase signaling. Am J Transl Res. 2021;13(4):2365–78.
Gou L, Xue C, Tang X, Fang Z. Inhibition of Exo-miR-19a-3p derived from cardiomyocytes promotes angiogenesis and improves heart function in mice with myocardial infarction via targeting HIF-1α. Aging (Albany NY). 2020;12(23):23609–18.
Wang L, Zhang J. Exosomal lncRNA AK139128 derived from hypoxic cardiomyocytes promotes apoptosis and inhibits cell proliferation in cardiac fibroblasts. Int J Nanomedicine. 2020;15:3363–76.
Zhang Z, Xu Y, Cao C, Wang B, Guo J, Qin Z, et al. Exosomes as a messager to regulate the crosstalk between macrophages and cardiomyocytes under hypoxia conditions. J Cell Mol Med. 2022;26(5):1486–500.
Yang J, Yu X, Xue F, Li Y, Liu W, Zhang S. Exosomes derived from cardiomyocytes promote cardiac fibrosis via myocyte-fibroblast cross-talk. Am J Transl Res. 2018;10(12):4350–66.
Ribeiro-Rodrigues TM, Laundos TL, Pereira-Carvalho R, Batista-Almeida D, Pereira R, Coelho-Santos V, et al. Exosomes secreted by cardiomyocytes subjected to ischaemia promote cardiac angiogenesis. Cardiovasc Res. 2017;113(11):1338–50.
Wang Y, Zhao R, Shen C, Liu W, Yuan J, Li C, et al. Exosomal CircHIPK3 released from hypoxia-induced cardiomyocytes regulates cardiac angiogenesis after myocardial infarction. Oxid Med Cell Longev. 2020;2020:8418407.
Chen G, Yue A, Wang M, Ruan Z, Zhu L. The Exosomal lncRNA KLF3-AS1 from ischemic cardiomyocytes mediates IGF-1 secretion by MSCs to rescue myocardial ischemia-reperfusion injury. Front Cardiovasc Med. 2021;8: 671610.
Namazi H, Namazi I, Ghiasi P, Ansari H, Rajabi S, Hajizadeh-Saffar E, et al. Exosomes secreted by normoxic and hypoxic cardiosphere-derived cells have anti-apoptotic effect. Iran J Pharm Res. 2018;17(1):377–85.
Namazi H, Mohit E, Namazi I, Rajabi S, Samadian A, Hajizadeh-Saffar E, et al. Exosomes secreted by hypoxic cardiosphere-derived cells enhance tube formation and increase pro-angiogenic miRNA. J Cell Biochem. 2018;119(5):4150–60.
Cambier L, de Couto G, Ibrahim A, Echavez AK, Valle J, Liu W, et al. Y RNA fragment in extracellular vesicles confers cardioprotection via modulation of IL-10 expression and secretion. EMBO Mol Med. 2017;9(3):337–52.
de Couto G, Gallet R, Cambier L, Jaghatspanyan E, Makkar N, Dawkins JF, et al. Exosomal microRNA transfer into macrophages mediates cellular postconditioning. Circulation. 2017;136(2):200–14.
Li Z, Huo X, Chen K, Yang F, Tan W, Zhang Q, et al. Profilin 2 and endothelial exosomal profilin 2 promote angiogenesis and myocardial infarction repair in mice. Front Cardiovasc Med. 2022;9: 781753.
Liu W, Feng Y, Wang X, Ding J, Li H, Guan H, et al. Human umbilical vein endothelial cells-derived exosomes enhance cardiac function after acute myocardial infarction by activating the PI3K/AKT signaling pathway. Bioengineered. 2022;13(4):8850–65.
Su Q, Lv XW, Xu YL, Cai RP, Dai RX, Yang XH, et al. Exosomal LINC00174 derived from vascular endothelial cells attenuates myocardial I/R injury via p53-mediated autophagy and apoptosis. Mol Ther Nucleic Acids. 2021;23:1304–22.
Akbar N, Braithwaite AT, Corr EM, Koelwyn GJ, van Solingen C, Cochain C, et al. Rapid neutrophil mobilisation by VCAM-1+ endothelial extracellular vesicles. Cardiovasc Res. 2022.
Huang Y, Chen L, Feng Z, Chen W, Yan S, Yang R, et al. EPC-Derived Exosomal miR-1246 and miR-1290 regulate phenotypic changes of fibroblasts to endothelial cells to exert protective effects on myocardial infarction by targeting ELF5 and SP1. Front Cell Dev Biol. 2021;9: 647763.
Ke X, Yang R, Wu F, Wang X, Liang J, Hu X, et al. Exosomal miR-218-5p/miR-363-3p from Endothelial Progenitor Cells Ameliorate Myocardial Infarction by Targeting the p53/JMY Signaling Pathway. Oxid Med Cell Longev. 2021;2021:5529430.
Barile L, Cervio E, Lionetti V, Milano G, Ciullo A, Biemmi V, et al. Cardioprotection by cardiac progenitor cell-secreted exosomes: role of pregnancy-associated plasma protein-A. Cardiovasc Res. 2018;114(7):992–1005.
Hocine HR, Brunel S, Chen Q, Giustiniani J, San Roman MJ, Ferrat YJ, et al. extracellular vesicles released by allogeneic human cardiac stem/progenitor cells as part of their therapeutic benefit. Stem Cells Transl Med. 2019;8(9):911–24.
Liao Z, Chen Y, Duan C, Zhu K, Huang R, Zhao H, et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics. 2021;11(1):268–91.
Liu N, Xie L, Xiao P, Chen X, Kong W, Lou Q, et al. Cardiac fibroblasts secrete exosome microRNA to suppress cardiomyocyte pyroptosis in myocardial ischemia/reperfusion injury. Mol Cell Biochem. 2022;477(4):1249–60.
Del Campo CV, Liaw NY, Gunadasa-Rohling M, Matthaei M, Braga L, Kennedy T, et al. Regenerative potential of epicardium-derived extracellular vesicles mediated by conserved miRNA transfer. Cardiovasc Res. 2022;118(2):597–611.
Gennebäck N, Hellman U, Malm L, Larsson G, Ronquist G, Waldenström A, et al. Growth factor stimulation of cardiomyocytes induces changes in the transcriptional contents of secreted exosomes. J Extracell Vesicles. 2013;2.
Chu W, Li X, Li C, Wan L, Shi H, Song X, et al. TGFBR3, a potential negative regulator of TGF-β signaling, protects cardiac fibroblasts from hypoxia-induced apoptosis. J Cell Physiol. 2011;226(10):2586–94.
Hermans KC, Daskalopoulos EP, Blankesteijn WM. The Janus face of myofibroblasts in the remodeling heart. J Mol Cell Cardiol. 2016;91:35–41.
Loyer X, Zlatanova I, Devue C, Yin M, Howangyin KY, Klaihmon P, et al. Intra-cardiac release of extracellular vesicles shapes inflammation following myocardial infarction. Circ Res. 2018;123(1):100–6.
Rogers RG, Fournier M, Sanchez L, Ibrahim AG, Aminzadeh MA, Lewis MI, et al. Disease-modifying bioactivity of intravenous cardiosphere-derived cells and exosomes in mdx mice. JCI Insight. 2019;4(7).
Gallet R, Dawkins J, Valle J, Simsolo E, de Couto G, Middleton R, et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur Heart J. 2017;38(3):201–11.
Malliaras K, Makkar RR, Smith RR, Cheng K, Wu E, Bonow RO, et al. Intracoronary cardiosphere-derived cells after myocardial infarction: evidence of therapeutic regeneration in the final 1-year results of the CADUCEUS trial (CArdiosphere-Derived aUtologous stem CElls to reverse ventricUlar dySfunction). J Am Coll Cardiol. 2014;63(2):110–22.
Ibrahim AG, Cheng K, Marbán E. Exosomes as critical agents of cardiac regeneration triggered by cell therapy. Stem Cell Reports. 2014;2(5):606–19.
Nguyen CT, Dawkins J, Bi X, Marbán E, Li D. Diffusion tensor cardiac magnetic resonance reveals exosomes from cardiosphere-derived cells preserve myocardial fiber architecture after myocardial infarction. JACC Basic Transl Sci. 2018;3(1):97–109.
Fraccarollo D, Galuppo P, Bauersachs J. Novel therapeutic approaches to post-infarction remodelling. Cardiovasc Res. 2012;94(2):293–303.
Dawkins JF, Ehdaie A, Rogers R, Soetkamp D, Valle J, Holm K, et al. Biological substrate modification suppresses ventricular arrhythmias in a porcine model of chronic ischaemic cardiomyopathy. Eur Heart J. 2022;43(22):2139–56.
Shyu KG, Wang BW, Lin CM. Reply to the letter to the Editor “ECs-derived exosomes: A novel therapeutic target for myocardial ischemia-reperfusion injury.” Int J Cardiol. 2021;332:39.
Zhou W, Zheng X, Cheng C, Guo G, Zhong Y, Liu W, et al. Rab27a deletion impairs the therapeutic potential of endothelial progenitor cells for myocardial infarction. Mol Cell Biochem. 2021;476(2):797–807.
Wang J, Sun X, Zhao J, Yang Y, Cai X, Xu J, et al. Exosomes: a novel strategy for treatment and prevention of diseases. Front Pharmacol. 2017;8:300.
Sreejit G, Abdel-Latif A, Athmanathan B, Annabathula R, Dhyani A, Noothi SK, et al. Neutrophil-Derived S100A8/A9 Amplify Granulopoiesis After Myocardial Infarction. Circulation. 2020;141(13):1080–94.
Yang J, Li Y, Xue F, Liu W, Zhang S. Exosomes derived from cardiac telocytes exert positive effects on endothelial cells. Am J Transl Res. 2017;9(12):5375–87.
Abrial M, Da Silva CC, Pillot B, Augeul L, Ivanes F, Teixeira G, et al. Cardiac fibroblasts protect cardiomyocytes against lethal ischemia-reperfusion injury. J Mol Cell Cardiol. 2014;68:56–65.
Streef TJ, Smits AM. Epicardial contribution to the developing and injured heart: exploring the cellular composition of the epicardium. Front Cardiovasc Med. 2021;8: 750243.
Li P, Hu J, Wang J, Zhang J, Wang L, Zhang C. The Role of Hydrogel in Cardiac Repair and Regeneration for Myocardial Infarction: Recent Advances and Future Perspectives. Bioengineering (Basel). 2023;10(2).
Liu S, Chen X, Bao L, Liu T, Yuan P, Yang X, et al. Treatment of infarcted heart tissue via the capture and local delivery of circulating exosomes through antibody-conjugated magnetic nanoparticles. Nat Biomed Eng. 2020;4(11):1063–75.
Han C, Zhou J, Liang C, Liu B, Pan X, Zhang Y, et al. Human umbilical cord mesenchymal stem cell derived exosomes encapsulated in functional peptide hydrogels promote cardiac repair. Biomater Sci. 2019;7(7):2920–33.
Zhang Y, Cai Z, Shen Y, Lu Q, Gao W, Zhong X, et al. Hydrogel-load exosomes derived from dendritic cells improve cardiac function via Treg cells and the polarization of macrophages following myocardial infarction. J Nanobiotechnology. 2021;19(1):271.
Zou Y, Li L, Li Y, Chen S, Xie X, Jin X, et al. Restoring cardiac functions after myocardial infarction-ischemia/reperfusion via an exosome anchoring conductive hydrogel. ACS Appl Mater Interfaces. 2021;13(48):56892–908.
Yao J, Huang K, Zhu D, Chen T, Jiang Y, Zhang J, et al. A Minimally invasive exosome spray repairs heart after myocardial infarction. ACS Nano. 2021;15(7):11099–111.
Zhu D, Li Z, Huang K, Caranasos TG, Rossi JS, Cheng K. Minimally invasive delivery of therapeutic agents by hydrogel injection into the pericardial cavity for cardiac repair. Nat Commun. 2021;12(1):1412.
Cheng G, Zhu D, Huang K, Caranasos TG. Minimally invasive delivery of a hydrogel-based exosome patch to prevent heart failure. J Mol Cell Cardiol. 2022;169:113–21.
Hu X, Ning X, Zhao Q, Zhang Z, Zhang C, Xie M, et al. Islet-1 mesenchymal stem cells-derived exosome-incorporated angiogenin-1 hydrogel for enhanced acute myocardial infarction therapy. ACS Appl Mater Interfaces. 2022;14(32):36289–303.
Shafei S, Khanmohammadi M, Ghanbari H, Nooshabadi VT, Tafti SHA, Rabbani S, et al. Effectiveness of exosome mediated miR-126 and miR-146a delivery on cardiac tissue regeneration. Cell Tissue Res. 2022;390(1):71–92.
Chung JJ, Kim ST, Zaman S, Helmers MR, Arisi MF, Li EC, et al. Therapeutic efficacy of cryopreserved, allogeneic extracellular vesicles for treatment of acute myocardial infarction. Int Heart J. 2021;62(2):381–9.
Monguió-Tortajada M, Prat-Vidal C, Moron-Font M, Clos-Sansalvador M, Calle A, Gastelurrutia P, et al. Local administration of porcine immunomodulatory, chemotactic and angiogenic extracellular vesicles using engineered cardiac scaffolds for myocardial infarction. Bioact Mater. 2021;6(10):3314–27.
Yuan J, Yang H, Liu C, Shao L, Zhang H, Lu K, et al. Microneedle patch loaded with exosomes containing microRNA-29b prevents cardiac fibrosis after myocardial infarction. Adv Healthc Mater. 2023;12(13): e2202959.
Chachques JC, Gardin C, Lila N, Ferroni L, Migonney V, Falentin-Daudre C, et al. Elastomeric Cardiowrap Scaffolds Functionalized with Mesenchymal Stem Cells-Derived Exosomes Induce a Positive Modulation in the Inflammatory and Wound Healing Response of Mesenchymal Stem Cell and Macrophage. Biomedicines. 2021;9(7).
Yin T, Liu Y, Ji W, Zhuang J, Chen X, Gong B, et al. Engineered mesenchymal stem cell-derived extracellular vesicles: a state-of-the-art multifunctional weapon against Alzheimer’s disease. Theranostics. 2023;13(4):1264–85.
Gong XH, Liu H, Wang SJ, Liang SW, Wang GG. Exosomes derived from SDF1-overexpressing mesenchymal stem cells inhibit ischemic myocardial cell apoptosis and promote cardiac endothelial microvascular regeneration in mice with myocardial infarction. J Cell Physiol. 2019;234(8):13878–93.
Xu L, Fan Y, Wu L, Zhang C, Chu M, Wang Y, et al. Exosomes from bone marrow mesenchymal stem cells with overexpressed Nrf2 inhibit cardiac fibrosis in rats with atrial fibrillation. Cardiovasc Ther. 2022;2022:2687807.
Liu X, Li X, Zhu W, Zhang Y, Hong Y, Liang X, et al. Exosomes from mesenchymal stem cells overexpressing MIF enhance myocardial repair. J Cell Physiol. 2020;235(11):8010–22.
Lv LH, Wan YL, Lin Y, Zhang W, Yang M, Li GL, et al. Anticancer drugs cause release of exosomes with heat shock proteins from human hepatocellular carcinoma cells that elicit effective natural killer cell antitumor responses in vitro. J Biol Chem. 2012;287(19):15874–85.
Gupta D, Zickler AM, El Andaloussi S. Dosing extracellular vesicles. Adv Drug Deliv Rev. 2021;178: 113961.
Okamura A, Yoshioka Y, Saito Y, Ochiya T. Can Extracellular Vesicles as Drug Delivery Systems Be a Game Changer in Cardiac Disease? Pharm Res. 2023;40(4):889–908.
Xi XM, Xia SJ, Lu R. Drug loading techniques for exosome-based drug delivery systems. Pharmazie. 2021;76(2):61–7.
Cheng G, Li W, Ha L, Han X, Hao S, Wan Y, et al. Self-assembly of extracellular vesicle-like metal-organic framework nanoparticles for protection and intracellular delivery of biofunctional proteins. J Am Chem Soc. 2018;140(23):7282–91.
Lee JR, Park BW, Kim J, Choo YW, Kim HY, Yoon JK, et al. Nanovesicles derived from iron oxide nanoparticles-incorporated mesenchymal stem cells for cardiac repair. Sci Adv. 2020;6(18):eaaz0952.
Vandergriff A, Huang K, Shen D, Hu S, Hensley MT, Caranasos TG, et al. Targeting regenerative exosomes to myocardial infarction using cardiac homing peptide. Theranostics. 2018;8(7):1869–78.
Wang X, Chen Y, Zhao Z, Meng Q, Yu Y, Sun J, et al. Engineered exosomes with ischemic myocardium-targeting peptide for targeted therapy in myocardial infarction. J Am Heart Assoc. 2018;7(15): e008737.
Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546(7659):498–503.
Zheng YL, Wang WD, Cai PY, Zheng F, Zhou YF, Li MM, et al. Stem cell-derived exosomes in the treatment of acute myocardial infarction in preclinical animal models: a meta-analysis of randomized controlled trials. Stem Cell Res Ther. 2022;13(1):151.
Meng H, Cheng W, Wang L, Chen S, Teng Y, Lu Z, et al. mesenchymal stem cell exosomes in the treatment of myocardial infarction: a systematic review of preclinical in vivo studies. J Cardiovasc Transl Res. 2022;15(2):317–39.
Dehghani L, Khojasteh A, Soleimani M, Oraee-Yazdani S, Keshel SH, Saadatnia M, et al. Safety of intraparenchymal injection of allogenic placenta mesenchymal stem cells derived exosome in patients undergoing decompressive craniectomy following malignant middle cerebral artery infarct, a pilot randomized clinical trial. Int J Prev Med. 2022;13:7.
Wang L, Jin Y. Noncoding RNAs as biomarkers for acute coronary syndrome. Biomed Res Int. 2020;2020:3298696.
Chen S, Hong X, Wu Y, Chen Z. Diagnostic and prognostic significance of microRNA-208a in acute myocardial infarction. Dis Markers. 2022;2022:7030722.
Salama AM, Khalil WA, Al-Zaky M, Abdallah SH, Kandil NT, Abdelsabour A, et al. MicroRNA-208a: a good diagnostic marker and a predictor of no-reflow in STEMI patients undergoing primary percutaneuos coronary intervention. J Cardiovasc Transl Res. 2020;13(6):988–95.
Zhang Y, Liu J, Mo Y, Chen Z, Chen T, Li Y, et al. Immortalized mesenchymal stem cells: a safe cell source for cellular or cell membrane-based treatment of glioma. Stem Cells Int. 2022;2022:6430565.
Sundaram K, Mu J, Kumar A, Behera J, Lei C, Sriwastva MK, et al. Garlic exosome-like nanoparticles reverse high-fat diet induced obesity via the gut/brain axis. Theranostics. 2022;12(3):1220–46.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (82030007, 2020), Science and Technology Project of Sichuan Province [23NSFSC1345, 2022YFS0614, 2022YFS0578].
Author information
Authors and Affiliations
Contributions
Yuchang Liu, Minrui Wang and Yang Yu contributed equally in preparing the original draft of the manuscript. Chunhong Li and Chunxiang Zhang guided the entire study. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
We give our consent for the manuscript to be published in Cell Communication and Signaling.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, Y., Wang, M., Yu, Y. et al. Advances in the study of exosomes derived from mesenchymal stem cells and cardiac cells for the treatment of myocardial infarction. Cell Commun Signal 21, 202 (2023). https://doi.org/10.1186/s12964-023-01227-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-023-01227-9