
Abstract
Background
Testis is an immune privileged organ, which prevents the immune response against sperm antigens and inflammation. Testicular cells responsible for immune tolerance are mainly Sertoli cells, which form the blood-testis barrier and produce immunosuppressive factors. Sertoli cells prevent inflammation in the testis and maintain immune tolerance by inhibiting proliferation and inducing lymphocyte apoptosis. It has been shown that 9-cis-retinoic acid (9cRA) blocks ex vivo apoptosis of peripheral blood lymphocytes and promotes the differentiation of Treg cells in the gut. However, the role of retinoid signaling in regulating the immune privilege of the testes remains unknown.
Objective
The aim of this study was to determine whether 9cRA, acting via the retinoic acid receptors (RAR) and the retinoic X receptors (RXR), controls the immunomodulatory functions of Sertoli cells by influencing the secretion of anti-inflammatory/pro-inflammatory factors, lymphocyte physiology and Treg cell differentiation.
Methods
Experiments were performed using in vitro model of co-cultures of murine Sertoli cells and T lymphocytes. Agonists and antagonists of retinoic acid receptors were used to inhibit/stimulate retinoid signaling in Sertoli cells.
Results
Our results have demonstrated that 9cRA inhibits the expression of immunosuppressive genes and enhances the expression of pro-inflammatory factors in Sertoli cells and lymphocytes, increases lymphocyte viability and decreases apoptosis rate. Moreover, we have found that 9cRA blocks lymphocyte apoptosis acting through both RAR and RXR and inhibiting FasL/Fas/Caspase 8 and Bax/Bcl-2/Caspase 9 pathways. Finally, we have shown that 9cRA signaling in Sertoli cells inhibits Treg differentiation.
Conclusion
Collectively, our results indicate that retinoid signaling negatively regulates immunologically privileged functions of Sertoli cells, crucial for ensuring male fertility. 9cRA inhibits lymphocyte apoptosis, which can be related to the development of autoimmunity, inflammation, and, in consequence, infertility.
Similar content being viewed by others


Introduction
Immune privilege implies to a specific immunological status in several places in the mammalian body where cells expressing autoantigens or foreign antigens are tolerated without provoking harmful inflammatory immune responses [1]. The mammalian testis is considered an immune privileged organ because it is able to tolerate autoantigens expressed by haploid germ cells (predominantly spermatids). These cells are produced during spermatogenesis as a result of meiotic division completed after puberty when immunocompetence is already established [2]. The immunologically privileged status of the testis has been confirmed by numerous studies on allogeneic and xenogeneic transplantation of tissues that survive for different times after implantation into the testis of small and large animals [3]. However, the testicular environment does not entirely prevent pathological conditions, such as infection, trauma, and inflammation, which might harm both steroidogenesis and spermatogenesis. Previous studies have shown that infection and inflammation of the testis is a cause of approximately 6–15% of all cases of fertility disturbances [4]. Chronic autoimmune orchitis, a frequent factor of male infertility, is thought to impair the function of the testes and epididymis through the inhibition of steroidogenesis and induction of germ cell apoptosis [5]. Therefore, it is particularly important to understand the molecular mechanisms involved in the interactions between immune system cells and testicular cells.
Research conducted in the past 25 years indicate that of all the non-immune testicular cells, Sertoli cells are main cell population responsible for immune tolerance in the testis. It was confirmed by observation that grafts transplanted into testis have been protected from rejection after most of the germ cells and testosterone producing Leydig cells were removed [6]. Furthermore, primary Sertoli cells isolated from mouse testes protected co-transplanted allogeneic and xenogeneic cells from rejection. Transplantation of Sertoli cells prolonged the survival of co-transplanted islets when they were grafted across immunological barriers, which proves that Sertoli cells are capable of retaining immunoprotective properties outside of the testis by creating an immune-privileged ectopic site [7, 8].
Sertoli cells provides structural and functional support for germ cells development. Adjacent Sertoli cells form tight junctions creating a physical blood–testis barrier (BTB) that protects the auto-immunogenic germ cells from the host’s immune system by limiting the access of germ cell antigens to interstitial immune cells and the passage of antibodies from interstitium to adluminal compartment. Blood–testis barrier was believed for many years to be critical for testicular immune privilege [9]. However, the interstitial compartment and early-stage germ cells that localize outside the BTB (spermatogonia and preleptotene spermatocytes) are also immune privilege. These observations suggested that other mechanisms are also involved in the maintenance of testicular immune privilege. Indeed, only some meiotic germ cell antigens are sequestered behind the Sertoli cell barrier while others egress from normal tubules in residual bodies during spermiation and reach the interstitium [10]. It was reported that Sertoli cells have the capacity to modulate the immune response by production and local secretion of immunomodulatory factors and cytokines, such as transforming growth factor β (TGFβ), tumor necrosis factor-α (TNFα) and its receptor TNFR1, interferons (IFN α, β, γ), interleukins (IL-1, IL-6, IL-10), galectin-1 (GAL-1), activin A, and growth factors (e.g. insulin growth factor-1, granulocyte monocyte-colony stimulating factor) [11]. These factors inhibit proliferation, regulate activation and survival of T lymphocytes present under physiological conditions in the interstitial area of the rodent testis, preventing inflammation in the testicle. In infertile patients with sperm autoimmunity, the number of lymphocytes is significantly high suggesting their involvement in testicular pathology under inflammatory conditions [12].
Several different subsets of T cells are detected in the testes, including regulatory T (Treg) cells, helper T (Th) cells, cytotoxic T (Tc) cells, and natural killer (NK) cells. It is well known that Treg cells play a relevant role in the induction of testicular immunosuppression and prevention of autoimmune diseases by suppressing other reactive T cells [5]. The Treg cell family consists of the natural CD4 + CD25 + Treg cells differentiated in thymus and the induced Treg cells generated from CD4 + CD25- precursors in periphery organs. Tregs are characterized based on the expression of the transcription factor, forkhead box P3 (FOXP3), currently the most reliable marker to identify this T cell subset [13]. Latest study demonstrated that Sertoli cells are able to induce Foxp3 expression and subsequently a regulatory phenotype in CD4 + CD25- T cells to maintain testis immunosuppression by producing TGFβ and indoleamine-2, 3-dioxygenase (IDO) [14].
Sertoli cells may also induce apoptosis of T cells, which is thought to be critical mechanism in maintaining testicular immune privilege. In the extrinsic apoptotic pathway FAS ligand (FASL) expressing Sertoli cells interacts with FAS-bearing lymphocytes, which leads to caspase activation and lymphocyte apoptosis [15]. The importance of this FAS - FASL pathway was confirmed by the study indicating that allograft rejection can be controlled by grafting of FASL-positive Sertoli cells [16, 17]. The other mechanism that mediates lymphocyte apoptosis important for the maintenance of peripheral tolerance is Bcl-2-associated X protein/B-cell lymphoma 2 (BAX/BCL-2) pathway (intrinsic mitochondrial pathway). It has been reported that the increase in BAX/BCL-2 ratio causes a caspase cascade and finally leads to the generation of an apoptotic phenotype. Notably, defects in Bcl2 expression correlate with autoimmunity in mice and men [18].
Retinoic acids (RA), the active metabolites of vitamin A, are essential for postnatal testicular development and male fertility. In rodents, deficiency of vitamin A during spermatogenesis can cause reversible infertility within a few months. Retinoic acid signaling promotes Sertoli cell maturation at puberty by suppressing their proliferation, initiates spermatogonial differentiation and the process of meiosis [19]. Retinoids regulate cellular functions by binding to two types of intracellular receptors, retinoic acid receptors (RARs: α, β, and γ) and retinoic X receptors (RXRs: α, β, and γ), which function as ligand-dependent transcription factors. After ligand binding, homo- or heterodimers of the receptors recruit coactivators and initiate gene transcription by binding to retinoic acid response elements located within the promoters of target genes [20]. 9-cis-retinoic acid (9cRA), endogenous bioactive metabolite formed by spontaneous isomerization of all-trans RA, binds with high affinity to both RARs and RXRs in vitro [21]. It was found, that rodent Sertoli cells express both RARα and RXRβ [22]. Early investigations exhibited disruption of Sertoli cell maturation and function and spermiation failure in mice with a Sertoli cell-specific knockout of the RXRβ [23, 24]. Similarly, mice with a Sertoli cell-specific ablation of RARα showed a delay in the final phase of Sertoli cell differentiation, defects in the integrity of the Sertoli-cell barrier, progressive spermatogenic degeneration, and infertility [25]. The study showed that the overexpression of a dominant-negative form of RARα in Sertoli cells disrupts the BTB at stages VII-XII and causes the large-scale apoptosis of differentiating germ cells, which indicates that retinoids contribute to immune privilege by regulating blood-testis barrier integrity [26]. However to date, the role of retinoid signaling in regulating other Sertoli cell-dependent mechanisms involved in testicular immune privilege remains unknown.
Importantly, in the immune system retinoids blocks the ex vivo apoptosis of peripheral blood lymphocytes by regulating FASL expression and suppresses inflammatory responses [27]. Recent reports have demonstrated the role of all-trans RA in inducing Treg cells in the gut, which may be one of the mechanisms by which retinoids regulate autoimmune reactions in vivo [28]. Nevertheless, it is unclear whether retinoids may modulate testicular lymphocytes activity.
The aim of this study was to determine whether 9cRA, acting via RARs and RXRs controls the immunomodulatory functions of Sertoli cells by influencing the secretion of anti-inflammatory/pro-inflammatory factors, lymphocyte physiology, and the differentiation of Treg cells.
Materials and methods
Murine primary Sertoli cell culture and treatment
Sertoli cells were isolated from testes of ~ 16-day-old CD1 mice according to previously described protocol, which enables to obtain differentiated non-proliferating Sertoli cell of high purity [29]. Testes denuded of tunica albuginea were incubated with the mixture of collagenase Type I (0.5 mg/mL) (Sigma-Aldrich, St. Louis, MO, USA) and DNase I (200 µg/mL) (Sigma-Aldrich, St. Louis, MO, USA) for 15 min at 35 °C to disperse the seminiferous tubules. Next, tubules were washed twice in Enriched DMEM: F12 medium (VWR International, Radnor, PA, USA), layered over 5% Percoll (Sigma-Aldrich, St. Louis, MO, USA)/95% 1× Hank’s balanced salt solution (HBSS; Invitrogen, Carlsbad, CA, USA), and allowed to settle for 20 min in room temperature. The bottom part of Percoll solution was transferred and incubated for 20 min at 35 °C in the mixture of trypsin (1 mg/mL) (Thermo Fisher Scientific, Waltham, MA, USA) and DNase I (200 µg/mL). After incubation, to halt the digestion 5 mL charcoal-stripped FBS (Thermo Fisher Scientific, Waltham, MA, USA) was immediately added. The fragmented tubules were washed twice in Enriched DMEM: F12 medium (800 g, 5 min) and further digested for 30 min with the mixture of collagenase (1 mg/mL), hyaluronidase (2 mg/mL) (Sigma-Aldrich, St. Louis, MO, USA), and DNase I (50 µg/mL) to generate loose cells. The residual tubules in the suspension were filtered away from the single cell suspension with use of 100 μm cell strainer. Sertoli cells and testicular germ cells were then washed 3 times by centrifugation (800 g, 5 min) in isolation medium. Pelleted cells were plated onto 6-well plates at 1 × 106 cells/cm2 in Enriched DMEM: F12 supplemented with serum and an antibiotic in a humidified atmosphere of 95% air and 5% CO2 (vol/vol) at 35oC. After 24 h, cultures were treated with 10% HBSS to lyse contaminating germ cells and obtain primary Sertoli cell cultures with > 90% purity. In all experiments the monolayers composed of pure Sertoli cells were used. Next, primary Sertoli cells were treated with 10− 7 M 9cRA (Sigma-Aldrich, St. Louis, MO, USA), 10− 6 M CD2665 (Tocris Bioscience, Bristol, UK), 2 × 10− 6 M Adapalene (Tocris Bioscience, Bristol, UK), 10− 6 M HX531 (Tocris Bioscience, Bristol, UK), or 10− 5 M Bexarotene (Tocris Bioscience, Bristol, UK) for 24 h. Control cells were incubated in the presence of the vehicle (0.01% dimethyl sulfoxide).
Transplantation and inhibin A (INHA) expression analysis of 9cRA-treated Sertoli cells underneath kidney capsule
Twelve male CD1 mice were obtained from the Animal House, Faculty of Pharmacy, Jagiellonian University in Krakow. Animals were maintained on a 12-h light and 12-h dark cycle and received ad libitum access to standard LSM food (Agropol, Motycz, Poland) and water. Allograft transplantation was proceeded as previously described [30, 31]. Briefly, two million control or 9cRA-treated Sertoli cells (as described in 2.1) were transplanted underneath both left and right kidney capsules of healthy anesthetized mice. Negative controls constituted mice treated only with vehicle (PBS). The present study was performed according to Polish ethical and legal requirements and in compliance with the Directive 2010/63/EU on the Protection of Animals Used for Scientific Purposes. The use of the animals for the experiments was approved by 2nd Local Institutional Animal Care and Use Committee in Krakow, Poland (217/2022). The kidneys were removed from mice after 21 days and the presence of Sertoli cells was assessed by immunohistochemical analysis of inhibin A (INHA). Since INHA is not expressed in kidney cells, it was used as a marker of Sertoli cells [32]. Immunohistochemical staining was performed on paraffin sections fixed in 4% paraformaldehyde according to previously published protocol [33]. To achieve antigen retrieval, slides were immersed in 10 mM citrate buffer (pH 6.0) and heated for 5 min in a microwave oven (750 W). Sections were incubated with INHA primary antibody (PA5-13681; Thermo Fisher Scientific, Waltham, MA, USA), at 4 °C overnight followed by biotinylated secondary antibody (goat anti-rabbit IgG; 1:400; Vector, Burlingame CA, USA) for 1 h, and avidin-biotinylated horseradish peroxidase complex (ABC/HRP; 1:100; Vectastain Elite ABC Reagent, Vector Lab. Burlingame CA, USA) for 30 min. Bound antibody was visualized with 0.05% 3,3′-diaminobenzidine tetrachloride (DAB; Sigma-Aldrich, St. Louis, MO, USA). In negative control sections incubation with a primary antibody was omitted. Thereafter, sections were washed and counterstained with Mayer’s hematoxylin, dehydrated, and mounted using DPX mounting media (Sigma-Aldrich, St. Louis, MO, USA). Analyses were repeated three times. The presence of SCs and immune cell infiltration were subsequently observed with Nikon Eclipse Ni microscope at ×400 magnification (Nikon Instech Co., Ltd., Tokyo, Japan). The presence of immune cell infiltration was assessed by visual evaluation of hematoxylin-stained samples based on the morphological characteristic of immune cells (small cells with dense, dark blue nuclei; [34]).
Isolation of murine spleen lymphocytes; 9-cis-retinoid acid-treated Sertoli cell – spleen lymphocytes co-culture
Spleen lymphocytes were isolated from healthy ~ 60-day-old mice according to previously described protocol [35]. Spleens were isolated into ice cold sterile RPMI/10% FBS medium (VWR International, Radnor, PA, USA), and gently cut into small pieces. To release the cells, spleen tissue pieces were gently pressed through the 70 μm strainer using syringe plunger. Isolated cells were centrifuged (900 g, 10 min), cell pellet was immediately disaggregated, re-suspended in 5 ml ACK (Ammonium-Chloride-Potassium) lysis buffer (Thermo Fisher Scientific, Waltham, MA, USA) and incubated on ice for 3 min to lyse red blood cells. Following the incubation, cells were washed in ice cold PBS and filtered through 70 μm cell strainer. Spleen lymphocytes (1 × 106 cells/well) were co-cultured with 9cRA-pretreated Sertoli cells (1 × 106 cells/well) for 48 h (1:1 ratio). Subsequently, Sertoli cells and lymphocytes were separately collected for further analyses.
Isolation of murine CD4 + CD25- lymphocytes; 9-cis-retinoid acid-treated Sertoli cell – CD4 + CD25-lymphocytes co-culture
Single-cell suspension of CD4 + CD25- T cells was isolated and purified using EasySep™ CD4 + CD25 + T cell isolation kit (StemCell, Auburn, CA, USA) according to the manufacturer’s instructions (purity > 95%). Shortly, cells were isolated by negative selection. First, CD4 + T cells were pre-enriched using EasySep™ Mouse CD4 + T Cell Isolation Cocktail with antbodies recognizing specific cell surface markers. Then, CD25- were negatively selected using EasySep™ Mouse CD25 Regulatory T Cell Selection Cocktail, which contains antibodies recognizing CD25. The EasySep™ cocktails labeled cells with antibodies and the cells were separated using an EasySep™ magnet. Spleen CD4 + CD25- lymphocytes were co-cultured with 9cRA-treated Sertoli cells for 4 days in 1:1 ratio (1 × 106 cells/well) and separately collected for further analyses.
Lymphocyte viability and apoptosis assays
After 48 h of Sertoli cell-lymphocyte co-culture, the lymphocyte suspension was transferred to another culture plate to detect the cell viability and apoptosis by 3-(4,5- dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay and TUNEL (TdT-mediated dUTP nick end labeling) assay, respectively.
Briefly, MTT stock solution (5 mg/ml) (Thermo Fisher Scientific, Waltham, MA, USA) was added to each well and the cells were incubated at 35 °C for 6 h. A negative control was included by adding 10 µL of the MTT stock solution to medium alone. Next, 100 µL of the sodium dodecyl sulfate (SDS) – HCl solution was added to each well to dissolve the formazan. After 4-hour incubation absorbance was measured at 570 nm with microtiter plate reader (LT-4500, Labtech International Ltd., Heathfield, UK).
For TUNEL, the lymphocyte apoptosis rate was evaluated using In Situ Cell Death Detection Kit, Fluorescein (Roche, Basel, Switzerland) following the instructions of the manufacturer. Cells were counted and the percentages of TUNEL-positive lymphocytes were calculated with Leica DM IL LED microscope (Leica Microsystems Inc., Wetzlar, Germany).
RNA isolation, reverse transcription, and quantitative RT-PCR (RT-qPCR)
Total RNA was extracted with TRIzol reagent (Life Technologies, Gaithersburg, MD, USA). Contaminating DNA and DNase was removed using TURBO DNA-free Kit (Ambion, Austin, TX, USA). The purity and quality of the RNA were examined by measuring the A260:A280 ratio (NanoDrop ND2000 Spectrophotometer, Thermo Scientific, Rocheford, IL, USA) and electrophoresis. To generate 20 µL of cDNA, High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA) was used. To estimate genomic DNA contamination of RNA samples, reactions in the absence of reverse transcriptase were carried out. RT-qPCR analyses were performed with the 10 ng cDNA templates, 0.5 µM primers (Institute of Biochemistry and Biophysics, Polish Academy of Sciences; Table 1) and SYBR Green mastermix (Applied Biosystems, Carlsbad, CA, USA) using the StepOne Real-time PCR system (Applied Biosystems, Carlsbad, CA, USA) in conditions: 55 °C for 2 min, 94 °C for 10 min, denaturation temperature 95 °C for 15 s, following by annealing temperature for 60 s to determine the cycle threshold (Ct) for quantitative measurement. Amplification efficiency was measured according to Svec et al. [36] and was between 90 and 112%. Melting curve analysis and agarose gel electrophoresis were performed to confirm amplification specificity. Negative control reactions were carried out. NormFinder was used to analyze the expression stability of commonly used reference genes and to select the most stable genes. Based on these analyses, housekeeping genes for normalizing RNA expression were selected: Rn18s, B2m, Rpl13a, and Hprt1. mRNA expressions of target genes were normalized to the geometric mean expression of the reference genes (relative quantification, RQ = 1) using the 2−∆∆Ct method.
Western blot analysis
Lysates were obtained by cell homogenization and sonification with RIPA buffer (Thermo Fischer Scientific, Rocheford, IL, USA) containing protease inhibitor coctail (Thermo Fisher Scientific, Rocheford, IL, USA). To determined protein concentration DC protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA) was used. 30 µg of proteins were resolved by SDS-PAGE under reducing conditions followed by transfer to polyvinylidene difluoride membranes (Sigma-Aldrich, St. Louis, MO, USA). Next, 5% (wt/vol) non-fat dry milk, 0.1% (vol/vol) Tween 20 was used to block non-specific binding sites. Successively membranes were incubated with the respective primary antibody for 24 h at 4 °C (Table 2) and then with a horseradish peroxidase-conjugated secondary antibody (1:3000; Thermo Fischer Scientific, Rocheford, IL, USA) for 1 h at room temperature to detect studied proteins. Proteins were imaged with Clarity Western ECL Blotting Substrate (Bio-Rad Laboratories, Hercules, CA, USA) and chemiluminescence detection system (ChemiDoc TM XRS + System, Bio–Rad Laboratories, Hercules, CA, USA). The molecular weights of targeted proteins were assessed by reference to standard proteins (PageRulerPrestained Protein Ladder, Thermo Fisher Scientific, Rocheford, IL, USA). Finally, all membranes were stripped and reprobed with an antibody against protein loading control – β-actin (Table 2). Relative intensities of protein bands were quantified using the Image Lab software (Bio-Rad Laboratories, Hercules, CA, USA).
Immunofluorescence
Immunofluorescence was performed on Sertoli cells seeded on coverslips, according to a previously published protocol [37]. Briefly, cells were washed with phosphate-buffered saline (PBS) and fixed with cold methanol–acetone. Non-specific binding was blocked with normal goat serum (NGS), then primary antibodies (listed in Table 2) followed by Cy3-conjugated goat anti-rabbit IgG secondary antibody (1:200; Thermo Fischer Scientific, Rocheford, IL, USA) were used. Images were captured with epifluorescence microscope Nikon Eclipse Ni (Nikon Instech Co., Tokyo, Japan). To confirm the specifity of primary antibodies negative controls with the omission of the primary antibody were performed (not shown).
Statistical analysis
Each data point was a mean ± SD of the results from three independent experiments. Normality and homogeneity of variance were tested with Shapiro-Wilk W-test and Levene’s test. Statistical differences in expression levels were assessed using one-way ANOVA, followed by Dunnett’s or Tukey’s post hoc comparison test or Kruskal-Wallis test. Analyses were done on raw data with Statistica 10 software (StatSoft Inc.). Data were considered statistically significant at *p < 0.05, **p < 0.01, ***p < 0.001.
Results
The role of 9-cis-retinoic acid in Sertoli cells and lymphocytes in the regulation of pro-inflammatory and immunosupressive genes involved in immunoregulation in the testis
First, we tested the effect of 9cRA on the expression of pro-inflammatory proteins IFN-γ, TNFR1, IL-1α, and IL-6 (Fig. 1A), and anti-inflammatory proteins TGFβ, IL-10, GAL-1, and IDO in primary murine Sertoli cells (Fig. 1B). Following the exposure of Sertoli cells to 9cRA the expression of IFN-γ, TNFR1, IL-1α, and IL-6 mRNA (Fig. 1A) and protein (Fig. 1A’) was upregulated (p < 0.05; p < 0.01; p < 0.001). In the cytoplasm, enhanced immunofluorescence signal of IFN-γ, TNFR1, and IL-6 was observed, whereas IL-1α signal increased in perinuclear area when compared to the control (Fig. 1A”). In contrast, 9cRA administration to Sertoli cells caused decrease of the expression of TGFβ, IL-10, GAL-1, and IDO transcripts and proteins (p < 0.05; p < 0.01; p < 0.001) (Fig. 1B; B’). Also immunofluorescence signal was clearly reduced in cytoplasm of 9cRA-treated Sertoli cells (Fig. 1B”).

Effect of 9cRA on the expression of pro-inflammatory (A-A”) and anti-inflammatory (B-B”) genes and proteins in murine Sertoli cells. Cells were treated with a vehicle (control, C), or 10− 7M 9cRA for 24 h. (A, B) Relative expression of Ifng, Tnfr1, Il1a, Il6, Tgfb1, Il10, Gal1, and Ido mRNAs was determined using quantitative real-time RT–PCR analysis. The histograms are the quantitative representation of data of three independent experiments, each in triplicate. The expression of the individual genes was normalized to the mean expression of the reference genes (Rn18s, B2m, Rpl13a, and Hprt1) as an internal control (relative quantification, RQ). (A’, B’) Western blot detection of TGFβ, IL-10, GAL-1, IDO, IFN-γ, TNFR1, IL-1α, and IL-6 proteins. The histograms are the quantitative representation after densitometry of data (mean ± SD) of three independent experiments, each in triplicate. The relative level of studied protein was normalized to β-actin. The protein levels within the control group were arbitrarily set as 1. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001 (A”,B”) Subcellular localization of TGFβ, IL-10, GAL-1, IDO, IFN-γ, TNFR1, IL-1α, and IL-6 proteins in murine Sertoli cells was visualized by immunofluorescence. Scale bar = 100 μm
Further, we tested whether retinoid signaling activation in Sertoli cells affects lymphocyte function. For this purpose, we co-cultured 9cRA-pretreated Sertoli cells with lymphocytes and analyzed the expression of lymphocyte pro-inflammatory proteins: signal transducer and activator of transcription 3 (STAT3), signal transducer and activator of transcription 5 (STAT5), interleukin-2 (IL-2) and c-Jun N-terminal kinases 1/2/3 (JNK) (Fig. 2). Following the exposure of murine lymphocytes to 9cRA-pretreated Sertoli cells the expression of STAT3, STAT5, IL-2, and JNK mRNA (Fig. 2A) and proteins (Fig. 2B) was upregulated (p < 0.05; p < 0.01; p < 0.001), indicating that retinoid signaling in Sertoli cells controls the function of testicular lymphocytes.

Effect of 9cRA on the expression of pro-inflammatory genes and proteins in murine lymphocytes. Cells were treated with a vehicle (control, C), or 10− 7M 9cRA for 24 h. (A) Relative expression of Stat3, Stat5, Il2, and Jnk mRNAs was determined using quantitative real-time RT–PCR analysis. The histograms are the quantitative representation of data of three independent experiments, each in triplicate. The expression of the individual genes was normalized to the mean expression of the reference genes (Rn18s, B2m, Rpl13a, and Hprt1) as an internal control (relative quantification, RQ). (B) Western blot detection of STAT3, STAT5, IL-2, and JNK proteins. The histograms are the quantitative representation after densitometry of data (mean ± SD) of three independent experiments, each in triplicate. The relative level of studied protein was normalized to β-actin. The protein levels within the control group were arbitrarily set as 1. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001
In order to evaluate the immunologically privileged capacity of 9cRA-treated Sertoli cells, Sertoli cells were transplanted into kidney capsule as allografts (Fig. 3). Clusters of INHA-positive Sertoli cells occurred both in the control and 9cRA-treated group. No positive signal was found in the intact kidneys of vehicle-treated mice. The presence of clear immune cell infiltration was visible only after transplantation of 9cRA-treated Sertoli cells. This confirms the results of in vitro studies and indicates the proinflammatory role of 9cRA signaling in Sertoli cells.

The effect of 9cRA-treated Sertoli cells transplantation underneath the kidney capsule. Immunohistochemical staining for INHA in control Sertoli cell graft (C),9cRA-treated Sertoli cell grafts (9cRA), and in the intact kidney (IK). Rectangles indicate the location of the higher magnification view (bottom images). No positive staining is observed when the primary antibody is omitted (see, right bottom section). Sections were counterstained with hematoxylin. K - kidney; arrows - Sertoli cells; open arrows - immune cells. Scale bars represent 20 μm
The role of 9-cis-retinoic acid signaling in Sertoli cells in the control of viability and apoptosis of lymphocytes
The co-culture model of murine Sertoli cells and lymphocytes allowed to analyze the impact of 9cRA-pretreated Sertoli cells on the lymphocyte viability and apoptosis. Based on MTT analysis we found that 9cRA significantly enhanced lymphocyte viability compared with the control group (p < 0.01) (Fig. 4A). RXR agonist (Bexarotene) increased lymphocyte viability to the level comparable to 9cRA-treated cells (p < 0.01), while RXR antagonist (HX531) completely abolished the effect of 9cRA on cell viability (p < 0.05). In contrast, neither RAR agonist (Adapalene) nor RAR antagonist (CD2665) had an effect on basal and 9cRA-stimulated lymphocyte viability level. This indicates that RXR-dependent signaling is involved in the regulation of lymphocytes viability.
The results of TUNEL assay revealed that 9cRA as well as RAR and RXR agonists significantly reduced the apoptosis rate of lymphocytes (p < 0.001) (Fig. 4B). The effect of 9cRA was partly suppressed by both RAR and RXR antagonists (p < 0.05), indicating that 9cRA acting through both RAR and RXR decreases the apoptosis rate in lymphocytes co-cultured with murine Sertoli cells (Fig. 4B).

Effect of retinoid signaling on murine T lymphocytes viability (MTT assay; A) and apoptosis (TUNEL assay; B). Sertoli cells were pre-treated with a vehicle (control, C), 10− 7 M 9cRA, 10− 6 M CD2665, 9cRA + CD2665, 2 × 10− 6 M Adapalene, 10− 6 M HX531, 9cRA + HX531, or 10− 5 M Bexarotene for 24 h. Spleen lymphocytes were co-cultured with 9cRA-pretreated Sertoli cells for 48 h. Bars represent mean ± SEM. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001
Effects of 9-cis-retinoic acid signaling in Sertoli cells on the FASL/FAS/Caspase 8 and BAX/BCL-2/Caspase 9 pathways
To explain the mechanism of the induction of lymphocyte apoptosis by 9cRA-pretreated Sertoli cells, the expression of proteins involved in both intrinsic and extrinsic apoptotic pathways was analyzed in 9cRA-pretreated Sertoli cells co-cultured with lymphocytes.
The expression of FASL, FAS, and Caspase 8 was reduced after incubation with 9cRA and this effect was abrogated only by RXR antagonist (p < 0.05; p < 0.01; p < 0.001). Moreover, only RXR agonist suppressed proteins expression (p < 0.05; p < 0.01). RAR agonist and antagonist had no effect on the expression of analyzed proteins. This indicates that primarily RXR is involved in the regulation of FASL/FAS/Caspase 8 pathway in Sertoli cells and lymphocytes (Fig. 5A-C’).

The expression of FASL (A, A’) in Sertoli cells, FAS (B, B’) and Caspase 8 (C, C’) in lymphocytes co-cultured with Sertoli cells pre-treated with 9cRA or RAR/RXR agonists or antagonists. Sertoli cells were pre-treated with a vehicle (control, C), 10− 7 M 9cRA, 10− 6 M CD2665, 9cRA + CD2665, 2 × 10− 6 M Adapalene, 10− 6 M HX531, 9cRA + HX531, or 10− 5 M Bexarotene for 24 h. Spleen lymphocytes were co-cultured with 9cRA-treated Sertoli cells for 48 h. (A, B, C) Relative expression of Fasl, Fas, and Caspase8 mRNAs was determined using quantitative real-time RT–PCR analysis. The histograms are the quantitative representation of data of three independent experiments, each in triplicate. The expression of the individual genes was normalized to the mean expression of the reference genes (Rn18s, B2m, Rpl13a, and Hprt1) as an internal control (relative quantification, RQ). (A’, B’, C’) Western blot detection of FASL, FAS, and Caspase8 proteins. The histograms are the quantitative representation after densitometry of data (mean ± SD) of three independent experiments, each in triplicate. The relative level of studied protein was normalized to β-actin. The protein levels within the control group were arbitrarily set as 1. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001
Upregulation of BCL-2 expression and downregulation of BAX/BCL-2 ratio and Caspase 9 expression were found in murine lymphocytes after 9cRA treatment when compared to the control (p < 0.01; p < 0.001) (Fig. 6B-D’). The effect of 9cRA on BAX, BCL-2, and Caspase 9 expression was completely abolished only by RAR antagonist (p < 0.05; p < 0.01; p < 0.001) (Fig. 6A-D’). What is more, only exposure to the RAR agonist altered the expression of studied proteins (p < 0.01; p < 0.001). Thus, RAR seems to play a dominant role in the control of BAX/BCL-2/Caspase 9 pathway in lymphocytes.

The expression of BAX (A, A’), BCL-2 (B, B’), and Caspase 9 (D, D’) in lymphocytes co-cultured with Sertoli cells pre-treated with 9cRA or RAR/RXR agonists or antagonists. Sertoli cells were pre-treated with a vehicle (control, C), 10− 7 M 9cRA, 10− 6 M CD2665, 9cRA + CD2665, 2 × 10− 6 M Adapalene, 10− 6 M HX531, 9cRA + HX531, or 10− 5 M Bexarotene for 24 h. Spleen lymphocytes were co-cultured with 9cRA-treated Sertoli cells for 48 h. (A, B, D) Relative expression of Bax, Bcl2, and Caspase9 mRNAs was determined using quantitative real-time RT–PCR analysis. The histograms are the quantitative representation of data of three independent experiments, each in triplicate. The expression of the individual genes was normalized to the mean expression of the reference genes (Rn18s, B2m, Rpl13a, and Hprt1) as an internal control (relative quantification, RQ). (A’, B’, D’) Western blot detection of BAX, BCL-2, and Caspase 9 proteins. The histograms are the quantitative representation after densitometry of data (mean ± SD) of three independent experiments, each in triplicate. The relative level of studied protein was normalized to β-actin. The calculated ratio of BAX/BCL-2 mRNA (C) or protein (C’) expression. The protein levels within the control group were arbitrarily set as 1. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001
Effects of 9-cis-retinoic acid signaling in Sertoli cells on the Treg cell differentiation
Latest study demonstrated that TGFβ is able to induce Foxp3 expression and subsequently a regulatory phenotype in CD4 + CD25- murine T cells [38]. Herein, we demonstrated that treatment with 9cRA decreased the expression of TGFβ in Sertoli cells (see subsection 3.1). Therefore, indirect effect of 9cRA signaling in Sertoli cells on Treg differentiation may be proposed. To test such a possibility, we isolated CD4 + CD25- T cells from spleen, co-cultured them with 9cRA-treated Sertoli cells, and analyzed FOXP3 expression.
Both RT-qPCR and western blot revealed that 9cRA as well as RAR and RXR agonists downregulate the FOXP3 expression (p < 0.05; p < 0.01; p < 0.001). In T cells, RAR and RXR antagonists partly abrogated the effect of 9cRA on FOXP3 expression, which suggests that 9cRA signaling through both receptors affects Treg cell differentiation (p < 0.05; p < 0.01) (Fig. 7A-B).

The expression of FOXP3 in CD4 + CD25- lymphocytes co-cultured with Sertoli cells pre-treated with 9cRA or RAR/RXR agonists or antagonists. Sertoli cells were pre-treated with a vehicle (control, C), 10− 7 M 9cRA, 10− 6 M CD2665, 9cRA + CD2665, 2 × 10− 6 M Adapalene, 10− 6 M HX531, 9cRA + HX531, or 10− 5 M Bexarotene for 24 h. CD4 + CD25- lymphocytes were co-cultured with 9cRA-treated Sertoli cells for 4 days. (A) Relative expression of Foxp3 mRNAs was determined using quantitative real-time RT–PCR analysis. The histograms are the quantitative representation of data of three independent experiments, each in triplicate. The expression of the individual genes was normalized to the mean expression of the reference genes (Rn18s, B2m, Rpl13a, and Hprt1) as an internal control (relative quantification, RQ). (B) Western blot detection of FOXP3 protein. The histograms are the quantitative representation after densitometry of data (mean ± SD) of three independent experiments, each in triplicate. The relative level of studied protein was normalized to β-actin. The protein levels within the control group were arbitrarily set as 1. Significant differences from control values are denoted as *p < 0.05, **p < 0.01, and ***p < 0.001
Discussion
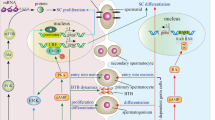
We have presented here novel data demonstrating a link between retinoid signaling in Sertoli cells and testis immune privilege. Our results provided evidence that 9cRA signaling in Sertoli cells increases the expression of pro-inflammatory genes, while reducing the expression of anti-inflammatory genes, affects lymphocyte viability, blocks their apoptotic death and inhibits Treg differentiation, which can be related to the development of autoimmunity and inflammation (Fig. 8).

Schematic illustration summarizing main results obtained in the present study
Previous studies demonstrated that 9cRA signaling induces production of pro-inflammatory cytokines by dendritic cells, promotes the differentiation of effector T cells and the formation of lymphoid structures [39, 40]. In parallel to the activation of the innate immune response, RA can also promote human dendritic cells to induce production of T cells expressing anti-inflammatory Il10 gene, controlling inflammation and tissue homeostasis [41]. Therefore, the outcome of retinoid signaling depends on the cell type or its physiological state.
Herein, we revealed for the first time that exposure of murine primary Sertoli cells to 9cRA leads to upregulation of Ifng, Tnfr1, Il1a, and Il6 genes. It is well established that elevated expression of these genes is closely related with the induction and progression of the inflammation in the testis. Elhija and coworkers [42] observed over-expression of IL-6 in testicular homogenates of mature and immature mice after systemic inflammation. Moreover, intratesticular injection of IL-6 induced inflammation and damage of seminiferous tubules. Interestingly, rat Sertoli cells cultured in the presence of IL-6 exhibited a redistribution of tight junction proteins [43]. In rat in vivo model of experimental autoimmune orchitis (EAO), an increase in the number of TNFR1 and IL-6 positive cells has been linked to progression of the disease [44, 45]. In EAO rat testis involvement of IFN-γ in inflammatory infiltration was also detected [46]. Equally important, an increase in IL-1 expression during testis inflammation and its role in neutrophil recruitment from the testicular vasculature have also been demonstrated [47]. Moreover, a few studies proved that IL-1, IL6, and TNFα stimulate each other’s production in an amplification loop, progressing T lymphocyte activation in many tissues [48, 49]. Notably, TNFα was able to increase IL-1α and IL-6 production in rat Sertoli cells [50].
On account of these data, increase in IFN-γ, TNFR1, IL-1α, and IL-6 expression in Sertoli cells following 9cRA allows us to consider 9cRA as a factor promoting pro-inflammmatory phenotype of murine Sertoli cells. This is confirmed by the observation that in murine Sertoli cells 9cRA downregulates the expression of genes involved in maintaining immune tolerance in the testis: Tgfb, Il10, Gal1, and Ido. It was previously found that TGFβ protected islet β-cell grafts after co-transplantation with Sertoli cells [51]. Moreover, overexpression of IL-10 in the testis significantly reduced inflammation in EAO model [52], as well as administration of exogenous GAL-1 suppressed the clinical signs of EAO and testicular inflammation, and promoted the differentiation of tolerogenic dendritic cells and Tregs [53, 54]. However, a pro-inflammatory role of GAL-1 in Sertoli cells was also reported [55]. Finally, IDO expressed by rat Sertoli exerts immunosuppressive effect, modulating inflammatory response to germ cell antigens [56].
To further confirm that 9cRA signaling in Sertoli cells may disrupt testicular immune privilege, we examined the expression of pro-inflammatory proteins in lymphocytes co-cultured with 9cRA-stimulated Sertoli cells and found upregulation of IL-2, STAT3, STAT5, and JNK expression. It was previously reported that T lymphocytes responsible for the induction of EAO secrete IL-2 and inhibition of IL-2/STAT5 signaling is essential for the development of FOXP3 + Tregs [44, 57, 58]. Activation of STAT3 is associated with the progression of autoimmunological aggression [59]. Notably, IL-6 strongly regulates STAT3 activation [60], which is in agreement with our results, showing an increase in the expression of both proteins in Sertoli cells. JNK regulates the proliferation, maturation, and activity of T cells and synthesis of pro-inflammatory cytokines IL-2, IL-6, and TNF-α. Several studies demonstrated the importance of JNK pathway in inflammatory disorders such as rheumatoid arthritis and atherosclerosis [61, 62]. In the light of the above-mentioned data, enhanced expression of IL-2, STAT3, STAT5, and JNK in response to 9cRA detected in our study may contribute to impaired T lymphocyte physiology and disruption of testis immune privilege. An in vivo model was used to confirm the role of 9cRA in the control of immune privilege. In the present study, lymphocyte infiltrations were observed underneath the kidney capsule 20 days after transplantation of 9cRA-treated Sertoli cells, which indicates that 9cRA weakened the ability of Sertoli cells to prevent the aggression of lymphocytes.
Testis as immune-privileged site can prolong the survival of allografts and xenografts [30, 63]. The capability of Sertoli cells to inhibit T lymphocyte proliferation, confirmed in cell culture experiments, could be highly related to the graft survival [64]. Therefore, to explain the effects of retinoid-dependent changes in Sertoli cell secretory activity on lymphocyte fate, we examined whether 9cRA signaling in Sertoli cells regulates T lymphocyte proliferation and apoptosis.
We observed increase in viability of lymphocytes co-cultured with 9cRA-pretreated Sertoli cells, which indicates that retinoid signaling activation limits the ability of Sertoli cells to prevent the proliferation of lymphocytes. Our results showed that prolonged lymphocyte survival is dependent on 9cRA action through RXRs in Sertoli cells. Previously, only direct relationship between RXRα expression in T lymphocytes and cell cycle progression and cell proliferation was demonstrated [65]. Although (based on our results) RARs activation in Sertoli cells does not affect lymphocyte viability, activation of RARs signaling directly in T cells or neural stem cells was found to affect proliferation of these cells [66, 67].
We further demonstrated that 9cRA, acting in Sertoli cells via both RARs and RXRs, inhibited lymphocyte apoptotic cell death, for the first time providing evidence for Sertoli cell-mediated effect of retinoids on lymphocytes apoptosis. So far, only the direct effect of retinoids on cell apoptosis has been detected in T cell hybridomas, thymocytes and peripheral blood lymphocytes [68,69,70]. This effect was described as mediated through both RARs and RXRs and involved in blockade of FASL expression [70, 71].
In our co-culture experiments, mRNA and protein expressions of FASL in RA-treated Sertoli cells as well as FAS and Caspase 8 in lymphocytes were significantly suppressed, which indicates that Sertoli cell - lymphocyte FAS/FASL/Caspase8 system was disturbed by 9cRA. Of note, earlier studies revealed inhibitory effect of TNFα on FASL expression in Sertoli cells and demonstrated a link between Sertoli cell expression of TGFβ1 and FASL and lymphocyte apoptosis [72, 73]. Therefore, it is likely that TNFα and TGFβ1, which have been upregulated in our study after exposure of Sertoli cells to 9cRA, contribute to indirect inhibition of lymphocyte apoptosis by retinoids. It is worth to mentioning, however, that in fibroblasts 9cRA exerts stimulatory effect on the expression of FASL, resulting in the induction of Jurkat T cells death [74].
Based on the results presented herein, the regulation of FASL/FAS/Caspase8 pathway in murine Sertoli cell - lymphocytes co-culture by 9cRA is dependent on RXRs. In contrast, RARs activation in Sertoli cells downregulates intrinsic apoptotic pathway (reducing BAX/ BCL-2 ratio and Caspase 9) in this model. It was previously reported that Sertoli cells prevent germ cell apoptosis via contact dependent or paracrine cell-to-cell interactions by upregulating BCL-2 [75]. Notably, direct inhibitory effects of RARγ activation on this pathway has been reported to limit cell apoptosis in adenocarcinoma cells [76].
Collectively, these results indicate the inhibitory effect of 9cRA on lymphocyte apoptosis mediated through activation of both RARs and RXRs in Sertoli cells, leads to suppresion of the FASL/FAS/Caspase8 and BAX/BCL-2/Caspase 9 pathways, which may reduce testis immune privilege.
Lastly, we provided evidence that retinoid signaling in Sertoli cells can affect Treg differentiation. The ability of Sertoli cells to induce the generation of Tregs was reported previously in diabetic mouse model [77]. Although, the exact mechanism has not been fully understood, earlier in vitro studies showed that mouse Sertoli cells induce Tregs from naive T cells after exposure to IFN-γ [78]. It has been proposed that Sertoli cells can promote Tregs differentiation (within or outside the testis) either directly by secreting regulatory factors such as soluble JAGGED1, TFG-β, IL-10, and IDO, or indirectly through activation of other immunotolerant cells [79, 80]. In view of these data, inhibitory effect of 9cRA/RAR/RXR signaling on the expression of Treg marker FOXP3 detected in our study may result from the downregulation of TFG- β, IL-10, and IDO found in Sertoli cells after 9cRA treatment.
Finally, it is important to note that RXRs control gene expression by forming heterodimers not only with RARs, but also with other nuclear receptors (such as peroxisome proliferator-activated receptors (PPAR), pregnane X receptor (PXR), liver X receptor (LXR), farnesoid X receptor (FXR)). Thus, the observed effect of RXRs in the regulation of immune privilege in the testis may be additionally related to other signaling pathways, which requires further investigation.
Conclusion
It is widely known that vitamin A is necessary for the proper course of spermatogenesis and maintaining male fertility. However, our work has shown for the first time that the activation of retinoid signaling in Sertoli cells by 9cRA may limit their immunosuppressive functions. It is likely, therefore, that excessive activation of retinoid receptors negatively affects Sertoli cells immunomodulatory properties, which can potentially lead to inflammation and impaired fertility. Thus, our findings point to the importance of maintaining the appropriate level of retinoic acid signaling in the testis.
Despite many years of research knowledge regarding the interaction of the immune system cells and the male reproductive system in the process of creating and maintaining an immunologically privileged environment in the male gonad of mammals is still incomplete. Nevertheless, our present results shed light on the mechanisms involved in the indirect effect of Sertoli cells on the function of lymphocytes and the induction of Treg differentiation, which is particularly important to understand the molecular basis of the interactions between immune and testicular cells.
Data availability
The data that support the findings of this study are available on request from the corresponding author.
Abbreviations
- 9cRA:
-
9-cis-Retinoic Acid
- ACK:
-
Ammonium-Chloride-Potassium
- Bax:
-
Bcl-2-associated x protein
- Bcl-2:
-
B-cell lymphoma 2
- BTB:
-
Blood-Testis Barrier
- DMEM:
-
Dulbecco’s Modified Eagle Medium
- FasL:
-
Fas Ligand
- FBS:
-
Fetal Bovine Serum
- FoxP3:
-
Forkhead box P3
- FXR:
-
Farnesoid X Receptor
- GAL-1:
-
Galectin-1
- HBSS:
-
Hank’s Balanced Salt Solution
- IDO:
-
Iindoleamine-2, 3-Dioxygenase
- IFN α,β:
-
Interferonαβ
- IL-1:
-
Interleukin 1
- IL-2:
-
Interleukin 2
- IL-6:
-
Interleukin 6
- IL-10:
-
Interleukin 10
- INHA:
-
Inhibin A
- JNK:
-
c-Jun N-terminal Kinases 1/2/3
- LXR:
-
Liver X Receptor
- NHS:
-
Normal Horse Serum
- NK:
-
Natural Killer T cells
- PPAR:
-
Peroxisome Proliferator-Activated Receptors
- PXR:
-
Pregnane X Receptor
- RAR:
-
Retinoid Acid Receptors
- RXR:
-
Retinoid X Receptors
- STAT3:
-
Signal Transducer and Activator of Transcription 3
- STAT5:
-
Signal Transducer and Activator of Transcription 5
- Tc:
-
Cytotoxic T cells
- TGFβ:
-
Transforming Growth Factor β
- Th:
-
Helper T cells
- TNFα:
-
Tumor Necrosis Factor-α
- TNFR1:
-
Tumor Necrosis Factor-α Receptor
- Treg:
-
Regulatory T cells
References
Kaur G, Mital P, Dufour JM. Testis immune privilege - assumptions versus facts. Anim Reprod. 2013;1:3–15.
Qu N, Ogawa Y, Kuramasu M, Nagahori K, Sakabe K, Itoh M. Immunological microenvironment in the testis. Reprod Med Biol. 2019;1:24–31.
Washburn RL, Hibler T, Kaur G, Dufour JM. Sertoli cell immune regulation: a double-edged sword. Front Immunol. 2022;13:913502.
Punab M, Poolamets O, Paju P, Vihljajev V, Pomm K, Ladva R, Korrovits P, Laan M. Causes of male infertility: a 9-year prospective monocentre study on 1737 patients with reduced total sperm counts. Hum Reprod. 2017;1:18–31.
Gong J, Zeng Q, Yu D, Duan YG. T lymphocytes and testicular immunity: a new insight into immune regulation in testes. Int J Mol Sci. 2020;22:57.
Cameron DF, Whittington K, Schultz RE, Selawry HP. Successful islet/abdominal testis transplantation does not require leydig cells. Transplantation. 1990;50:649–53.
Korbutt GS, Elliott JF, Rajotte RV. Cotransplantation of allogeneic islets with allogeneic testicular cell aggregates allows long-term graft survival without systemic immunosuppression. Diabetes. 1997;46:317–22.
Dufour JM, Rajotte RV, Kin T, Korbutt GS. Immunoprotection of rat islet xenografts by cotransplantation with Sertoli cells and a single injection of antilymphocyte serum. Transplantation. 2003;7:1594–6.
Mital P, Hinton BT, Dufour JM. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol Reprod. 2011;84:851–8.
Tung KS, Harakal J, Qiao H, Rival C, Li JC, Paul AG, Wheeler K, Pramoonjago P, Grafer CM, Sun W, Sampson RD, Wong EW, Reddi PP, Deshmukh US, Hardy DM, Tang H, Cheng CY, Goldberg E. Egress of sperm autoantigen from seminiferous tubules maintains systemic tolerance. J Clin Invest. 2017;127:1046–60.
Pérez CV, Theas MS, Jacobo PV, Jarazo-Dietrich S, Guazzone VA, Lustig L. Dual role of immune cells in the testis: protective or pathogenic for germ cells? Spermatogenesis. 2013;3:e23870.
Havrylyuk A, Chopyak V, Boyko Y, Kril I, Kurpisz M. Cytokines in the blood and semen of infertile patients. Cent Eur J Immunol. 2015;40:337–44.
Sakaguchi S, Yamaguchi T, Nomura T, Ono M. Regulatory T cells and immune tolerance. Cell. 2008;133:775–87.
Song X, Si Q, Qi R, Liu W, Li M, Guo M, Wei L, Yao Z. Indoleamine 2,3-dioxygenase 1: a promising therapeutic target in malignant tumor. Front Immunol. 2021;12:800630.
Ferguson TA, Griffith TS. A vision of cell death: Fas ligand and immune privilege 10 years later. Immunol Rev. 2006;213:228–38.
Bellgrau D, Gold D, Selawry H, Moore J, Franzusoff A, Duke RC. A role for CD95 ligand in preventing graft rejection. Nature. 1995;377:630–2.
Griffith TS, Brunner T, Fletcher SM, Green DR, Ferguson TA. Fas ligand-induced apoptosis as a mechanism of immune privilege. Science. 1995;270:1189–92.
Droin NM, Green DR. Role of Bcl-2 family members in immunity and disease. Biochim Biophys Acta. 2004;1644:179–88.
Nicholls PK, Harrison CA, Rainczuk KE, Wayne Vogl A, Stanton PG. Retinoic acid promotes Sertoli cell differentiation and antagonises activin-induced proliferation. Mol Cell Endocrinol. 2013;377:33–43.
Ghyselinck NB, Vernet N, Dennefeld C, Giese N, Nau H, Chambon P, Viville S, Mark M. Retinoids and spermatogenesis: lessons from mutant mice lacking the plasma retinol binding protein. Dev Dyn. 2006;235:1608–22.
Yoo HS, Moss KO, Cockrum MA, Woo W, Napoli JL. Energy status regulates levels of the RAR/RXR ligand 9-cis-retinoic acid in mammalian tissues: glucose reduces its synthesis in β-cells. J Biol Chem. 2023;299:105255.
Vernet N, Dennefeld C, Rochette-Egly C, Oulad-Abdelghani M, Chambon P, Ghyselinck NB, Mark M. Retinoic acid metabolism and signaling pathways in the adult and developing mouse testis. Endocrinology. 2006;147:96–110.
Vernet N, Dennefeld C, Klopfenstein M, Ruiz A, Bok D, Ghyselinck NB, Mark M. Retinoid X receptor beta (RXRB) expression in Sertoli cells controls cholesterol homeostasis and spermiation. Reproduction. 2008;136:619–26.
Lucas TF, Nascimento AR, Pisolato R, Pimenta MT, Lazari MF, Porto CS. Receptors and signaling pathways involved in proliferation and differentiation of Sertoli cells. Spermatogenesis. 2014;4:e28138.
Chung SS, Choi C, Wang X, Hallock L, Wolgemuth DJ. Aberrant distribution of junctional complex components in retinoic acid receptor alpha-deficient mice. Microsc Res Tech. 2010;73:583–96.
Hasegawa K, Saga Y. Retinoic acid signaling in Sertoli cells regulates organization of the blood-testis barrier through cyclical changes in gene expression. Development. 2012;139:4347–55.
Lee MO, Kang HJ, Kim YM, Oum JH, Park J. Repression of FasL expression by retinoic acid involves a novel mechanism of inhibition of transactivation function of the nuclear factors of activated T-cells. Eur J Biochem. 2002;269:1162–70.
Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, Belkaid Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J Exp Med. 2007;204:1775–85.
Chang YF, Lee-Chang JS, Panneerdoss S, MacLean JA, Rao MK. Isolation of Sertoli, Leydig, and spermatogenic cells from the mouse testis. Biotechniques. 2011;51:341–4.
Doyle TJ, Kaur G, Putrevu SM, Dyson EL, Dyson M, McCunniff WT, Pasham MR, Kim KH, Dufour JM. Immunoprotective properties of primary Sertoli cells in mice: potential functional pathways that confer immune privilege. Biol Reprod. 2012;86:1–14.
Sun Z, Nie Q, Zhang L, Niu R, Wang J, Wang S. Fluoride reduced the immune privileged function of mouse Sertoli cells via the regulation of Fas/FasL system. Chemosphere. 2017;168:318–25.
Gravesen E, Nordholm A, Mace M, Morevati M, Høgdall E, Nielsen C, Kjær A, Olgaard K, Lewin E. Effect of inhibition of CBP-coactivated β-catenin-mediated wnt signalling in uremic rats with vascular calcifications. PLoS ONE. 2018;13: e0201936.
Kamińska A, Pardyak L, Marek S, Górowska-Wójtowicz E, Kotula-Balak M, Bilińska B, Hejmej A. Bisphenol A and Dibutyl Phthalate affect the expression of juxtacrine signaling factors in rat testis. Chemosphere. 2018;199:182–90.
da Costa Souza P, Parra ER, Atanazio MJ, da Silva OB, Noleto GS, Ab’saber AM, de Morais Fernezlian S, Takagaki T, Capelozzi VL. Different morphology, stage and treatment affect immune cell infiltration and long-term outcome in patients with non-small-cell lung carcinoma. Histopathology. 2012;61:587–96.
Lim JF, Berger H, Su IH. Isolation and activation of murine lymphocytes. J Vis Exp. 2016;116:54596.
Svec D, Tichopad A, Novosadova V, Pfaffl MW, Kubista M. How good is a PCR efficiency estimate: recommendations for precise and robust qPCR efficiency assessments. Biomol Detect Quantif. 2015;3:9–16.
Kamińska A, Pardyak L, Marek S, Wróbel K, Kotula-Balak M, Bilińska B, Hejmej A. Notch signaling regulates nuclear androgen receptor AR and membrane androgen receptor ZIP9 in mouse Sertoli cells. Andrology. 2020;8:457–72.
Barrachina F, Ottino K, Elizagaray ML, Gervasi MG, Tu LJ, Markoulaki S, Spallanzani RG, Capen D, Brown D, Battistone MA. Regulatory T cells play a crucial role in maintaining sperm tolerance and male fertility. Proc Natl Acad Sci U S A. 2023;120. Article e2306797120.
Hall JA, Cannons JL, Grainger JR. Essential role for retinoic acid in the promotion of CD4(+) T cell effector responses via retinoic acid receptor alpha. Immunity. 2011;34:435–47.
Hall JA, Grainger JR, Spencer SP, Belkaid Y. The role of retinoic acid in tolerance and immunity. Immunity. 2011;35:13–22.
Bakdash G, Vogelpoel LT, van Capel TM, Kapsenberg ML, de Jong EC. Retinoic acid primes human dendritic cells to induce gut-homing, IL-10-producing regulatory T cells. Mucosal Immunol. 2015;8:265–78.
Elhija MA, Potashnik H, Lunenfeld E, Potashnik G, Schlatt S, Nieschlag E, Huleihel M. Testicular interleukin-6 response to systemic inflammation. Eur Cytokine Netw. 2005;16:167–72.
Pérez CV, Sobarzo CM, Jacobo PV, Pellizzari EH, Cigorraga SB, Denduchis B, Lustig L. Loss of occludin expression and impairment of blood-testis barrier permeability in rats with autoimmune orchitis: effect of interleukin 6 on Sertoli cell tight junctions. Biol Reprod. 2012;122:1–12.
Yule TD, Tung KSK. Experimental autoimmune orchitis induced by testis and sperm antigen-specific T cell clones: an important pathogenic cytokine is tumor necrosis factor. Endocrinology. 1993;133:1098–107.
Rival C, Theas MS, Guazzone VA, Lustig L. Interleukin-6 and IL-6 receptor cell expression in testis of rats with autoimmune orchitis. J Reprod Immunol. 2006;70:43–58.
Jacobo P, Pérez CV, Theas MS, Guazzone VA, Lustig L. CD4 + and CD8 + T cells producing Th1 and Th17 cytokines are involved in the pathogenesis of autoimmune orchitis. Reproduction. 2011;141:249–58.
Lysiak JJ, Nguyen QAT, Kirby JL, Turner TT. Ischemia-reperfusion of the murine testis stimulates the expression of proinflammatory cytokines and activation of c-jun N-terminal kinase in a pathway to E-selectin expression. Biol Reprod. 2003;69:202–10.
Stylianou E, Saklatvala J. Interleukin-1. Int J Biochem Cell Biol. 2000;30:1075–9.
De Cesaris P, Starace D, Riccioli A, Padula F, Filippini A, Ziparo E. Tumor necrosis factor-alpha induces interleukin-6 production and integrin ligand expression by distinct transduction pathways. J Biol Chem. 1998;273:7566–71.
Stephan JP, Syed V, Jegou B. Regulation of Sertoli cell IL-1 and IL-6 production in vitro. Mol Cell Endocrinol. 1997;134:109–18.
Avallet O, Vigier M, Leduque P, Dubois PM, Saez JM. Expression and regulation of transforming growth factor-beta 1 messenger ribonucleic acid and protein in cultured porcine Leydig and Sertoli cells. Endocrinology. 1994;134:2079–87.
Watanabe M, Kashiwakura Y, Kusumi N, Tamayose K, Nasu Y, Nagai A. Adeno-associated virus-mediated human IL-10 gene transfer suppresses the development of experimental autoimmune orchitis. Gene Ther. 2005;12:1126–32.
Pérez CV, Gómez LG, Gualdoni GS, Lustig L, Rabinovich GA, Guazzone VA. Dual roles of endogenous and exogenous galectin-1 in the control of testicular immunopathology. Sci Rep. 2015;5:12259.
Gao J, Wang X, Wang Y, Han F, Cai W, Zhao B, Li Y, Han S, Wu X, Hu D. Murine Sertoli cells promote the development of tolerogenic dendritic cells: a pivotal role of galectin-1. Immunology. 2016;148:253–65.
Lei T, Moos S, Klug J, Aslani F, Bhushan S, Wahle E, Frohlich S, Meinhardt A, Fijak M. Galectin-1 enhances TNFalpha-induced inflammatory responses in Sertoli cells through activation of MAPK signalling. Sci Rep. 2018;8:3741.
Gualdoni GS, Jacobo PV, Sobarzo CM, Pérez CV, Matzkin ME, Höcht C, Frungieri MB, Hill M, Anegon I, Lustig L, Guazzone VA. Role of indoleamine 2,3-dioxygenase in testicular immune-privilege. Sci Rep. 2019;9:15919.
Burchill MA, Yang J, Vogtenhuber C, Blazar BR, Farrar MA. IL-2 receptor beta-dependent STAT5 activation is required for the development of Foxp3 + regulatory T cells. J Immunol. 2007;178:280–90.
Laurence A, Tato CM, Davidson TS, Kanno Y, Chen Z, Yao Z, Blank RB, Meylan F, Siegel R, Hennighausen L, Shevach EM. O’shea JJ. Interleukin-2 signaling via STAT5 constrains T helper 17 cell generation. Immunity. 2007;26:371–81.
Nowell MI, Williams AS, Carty SA, Scheller J, Hayes AJ, Jones GW. Therapeutic targeting of IL-6 trans signaling counteracts STAT3 control of experimental inflammatory arthritis. J Immunol. 2009;182:613–22.
Leu CM, Wong FH, Chang C, Huang SF, Hu CP. Interleukin-6 acts as an antiapoptotic factor in human esophageal carcinoma cells through the activation of both STAT3 and mitogen-activated protein kinase pathways. Oncogene. 2003;22:7809–18.
Han Z, Boyle DL, Chang L, Bennett B, Karin M, Yang L, Manning AM, Firestein GS. c-Jun N-terminal kinase is required for metalloproteinase expression and joint destruction in inflammatory arthritis. J Clin Invest. 2001;108:73–81.
Adhikari N, Charles N, Lehmann U, Hall JL. Transcription factor and kinase-mediated signaling in atherosclerosis and vascular injury. Curr Atheroscler Rep. 2006;8:252–60.
Mital P, Kaur G, Dufour JM. Immunoprotective Sertoli cells: making allogeneic and xenogeneic transplantation feasible. Reproduction. 2010;139:495–504.
De Cesaris P, Filippini A, Cervelli C, Riccioli A, Muci S, Starace G, Stefanini M, Ziparo E. Immunosuppressive molecules produced by Sertoli cells cultured in vitro: biological effects on lymphocytes. Biochem Biophys Res Commun. 1992;186:1639–46.
Ishaq M, Zhang YM, Natarajan V. Activation-induced down-regulation of retinoid receptor RXRalpha expression in human T lymphocytes. Role of cell cycle regulation. J Biol Chem. 1998;273:21210–6.
Friedman A, Halevy O, Schrift M, Arazi Y. Retinoic acid promotes proliferation and induces expression of retinoic acid receptor-gene in murine T lymphocytes. Cell Immunol. 1993;152:240–8.
Zhao M, Chen S, Yang ML, Li SY, Jiang W, Xiao N. Vitamin a regulates neural stem cell proliferation in rats after hypoxic-ischemic brain damage via RARɑ-mediated modulation of the β-catenin pathway. Neurosci Lett. 2020;727:134922.
Yang Y, Merćep M, Ware CF, Ashwell JD. Fas and activation-induced Fas ligand mediate apoptosis of T cell hybridomas: inhibition of Fas ligand expression by retinoic acid and glucocorticoids. J Exp Med. 1995;181:1673–82.
Szondy Z, Reichert U, Fésüs L. Retinoic acids regulate apoptosis of T lymphocytes through an interplay between RAR and RXR receptors. Cell Death Differ. 1998;5:4–10.
Tóth K, Sarang Z, Scholtz B, Brázda P, Ghyselinck N, Chambon P, Fésüs L, Szondy Z. Retinoids enhance glucocorticoid-induced apoptosis of T cells by facilitating glucocorticoid receptor-mediated transcription. Cell Death Differ. 2010;18:783–92.
Bissonnette RP, Brunner T, Lazarchik SB, Yoo NJ, Boehm MF, Green DR, Heyman RA. 9-cis retinoic acid inhibition of activation-induced apoptosis is mediated via regulation of fas ligand and requires retinoic acid receptor and retinoid X receptor activation. Mol Cell Biol. 1995;15:5576–85.
Pentikäinen V, Erkkilä K, Suomalainen L, Otala M, Pentikäinen MO, Parvinen M, Dunkel L. TNFalpha down-regulates the Fas ligand and inhibits germ cell apoptosis in the human testis. J Clin Endocrinol Metab. 2001;86:4480–8.
Yin ZZ, Xie L, Zeng MH, Li R, Huang YB, Zhu M, Chen G, Chen S. Sertoli cells induce xenolymphocyte apoptosis in vitro. Transpl Proc. 2006;38:3309–11.
Nguyen T, Russell J. The regulation of FasL expression during activation-induced cell death (AICD). Immunology. 2001;103:426–34.
Russell LD, Chiarini-Garcia H, Korsmeyer SJ, Knudson CM. Bax-dependent spermatogonia apoptosis is required for testicular development and spermatogenesis. Biol Reprod. 2002;66:950–8.
Pettersson F, Dalgleish AG, Bissonnette RP, Colston KW. Retinoids cause apoptosis in pancreatic cancer cells via activation of RAR-gamma and altered expression of Bcl-2/Bax. Br J Cancer. 2002;87:555–61.
Fallarino F, Luca G, Calvitti M, Mancuso F, Nastruzzi C, Fioretti MC, Grohmann U, Becchetti E, Burgevin A, Kratzer R, van Endert P, Boon L, Puccetti P, Calafiore R. Therapy of experimental type 1 diabetes by isolated Sertoli cell xenografts alone. J Exp Med. 2009;206:2511–26.
Dal Secco V, Riccioli A, Padula F, Ziparo E, Filippini A. Mouse Sertoli cells display phenotypical and functional traits of antigen-presenting cells in response to interferon gamma. Biol Reprod. 2008;78:234–42.
Campese AF, Grazioli P, de Cesaris P, Riccioli A, Bellavia D, Pelullo M, Padula F, Noce C, Verkhovskaia S, Filippini A, Latella G, Screpanti I, Ziparo E, Starace D. Mouse Sertoli cells sustain de novo generation of regulatory T cells by triggering the notch pathway through soluble JAGGED1. Biol Reprod. 2014;90:53.
Porubska B, Vasek D, Somova V, Hajkova M, Hlaviznova M, Tlapakova T, Holan V, Krulova M. Sertoli cells possess immunomodulatory properties and the ability of mitochondrial transfer similar to mesenchymal stromal cells. Stem Cell Rev Rep. 2021;17:1905–16.
Acknowledgements
Not applicable.
Funding
This research was funded by the Priority Research Area BioS under the program Excellence Initiative – Research University at the Jagiellonian University in Krakow (grant no. U1U/P03/NO/03.15 to AK). The open-access publication of this article was funded by the programme “Excellence Initiative – Research University” at the Faculty of Biology of the Jagiellonian University in Kraków, Poland.
Author information
Authors and Affiliations
Contributions
The work presented here was conducted in collaboration with all authors. A.K., Z.A., and A.H. designed the research study; A.K., S.L., L.P., A.PF., and K.G. performed the research; A.K. and S.L. analyzed the data; A.K. prepared figures, and A.K. and A.H. wrote the paper. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kamińska, A., Pardyak, L., Lustofin, S. et al. 9-cis-retinoic acid signaling in Sertoli cells regulates their immunomodulatory function to control lymphocyte physiology and Treg differentiation. Reprod Biol Endocrinol 22, 75 (2024). https://doi.org/10.1186/s12958-024-01246-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12958-024-01246-2