
Abstract
Background
Obesity is strongly associated to insulin resistance, inflammation, and elevated plasma free fatty acids, but the mechanisms behind this association are not fully comprehended. Evidences suggest that endoplasmic reticulum (ER) stress may play a role in this complex pathophysiology. The aim of the present study was to investigate the involvement of inflammation and ER stress in the modulation of glucose transporter GLUT4, encoded by Slc2a4 gene, in L6 skeletal muscle cells.
Methods
L6 cells were acutely (2 h) and chronically (6 and 12 h) exposed to palmitate, and the expression of several proteins involved in insulin resistance, ER stress and inflammation were analyzed.
Results
Chronic and acute palmitate exposure significantly reduced GLUT4 protein (~ 39%, P < 0.01) and its mRNA (18%, P < 0.01) expression. Only acute palmitate treatment increased GRP78 (28%, P < 0.05), PERK (98%, P < 0.01), eIF-2A (35%, P < 0.01), IRE1a (60%, P < 0.05) and TRAF2 (23%, P < 0.05) protein content, and PERK phosphorylation (106%, P < 0.001), but did not elicit eIF-2A, IKK phosphorylation or increased XBP1 nuclear content. Additionally, acute and chronic palmitate increased NFKB p65 nuclear content (~ 30%, P < 0.05) and NFKB binding activity to Slc2a4 gene promoter (~ 45%, P < 0.05).
Conclusion
Different pathways are activated in acute and chronic palmitate induced-repression of Slc2a4/GLUT4 expression. This regulation involves activation of initial component of ER stress, such as the formation of a IRE1a-TRAF2-IKK complex, and converges to NFKB-induced repression of Slc2a4/GLUT4. These results link ER stress, inflammation and insulin resistance in L6 cells.
Similar content being viewed by others

Background
In the postprandial state, glucose uptake occurs mainly in skeletal muscle [1] through the insulin responsive glucose transporter GLUT4. Many reports have demonstrated that reduced GLUT4 expression in both skeletal muscle [2] and adipose tissue [3,4,5] is related to insulin resistance. Indeed, while the knockout of the Slc2a4 gene, which encodes GLUT4 protein, results in insulin resistance [6], its overexpression ameliorates diabetes [7].
Insulin resistance states, such as diabetes and obesity, have been associated to low-grade systemic inflammation [3, 8, 9]. Recently, we have reported the participation of inflammation in insulin resistance by showing that the nuclear factor kappa B (NFKB) downregulates specifically the Slc2a4 gene [10,11,12,13].
In addition to insulin resistance and inflammation, obesity is strongly related to elevated plasma free fatty acids (FFA), but the mechanisms behind this association are not fully comprehended. Recent evidences suggest that the endoplasmic reticulum (ER) stress may play a role in this complex pathophysiology.
Disturbed ER homeostasis results in unfolded and misfolded proteins, which can lead to cell dysfunction and death. In consequence of ER stress, an adaptive process named unfolded protein response (UPR) is triggered. UPR signaling is mainly mediated by ER transmembrane signal transducers, including the following: protein kinase R-like endoplasmic reticulum kinase (PERK), serine/threonine-protein kinase/endoribonuclease IRE1 (IRE1) and cyclic AMP-dependent transcription factor ATF-6 (ATF6). In non-stressed cells, 78 kDa glucose-regulated protein (GRP78), a central regulator of ER stress, binds and inactivates PERK, IRE1 and ATF6. In stressed cells, the accumulated unfolded proteins bind to GRP78, allowing the ER transmembrane proteins to be released from it and to become activated [14].
Palmitic acid (C16:0) is one of the most abundant saturated FFA in both human and rodent plasma [15]. It is able to induce peripheral insulin resistance, characterized by impaired glucose uptake by skeletal muscle [16]. As a matter of fact, it has been shown that palmitate is able to downregulate GLUT4 expression in skeletal muscle cells [17, 18] but little is known about the involved mechanisms.
The objective of the present study was to investigate the involvement of inflammation and ER stress in the modulation of Slc2a4/GLUT4 expression by acute and chronic palmitate exposure in L6 skeletal muscle cells.
Methods
Reagents
Rat L6 myoblasts (CRL 1458) were purchased from ATCC (APABCAM, Rio de Janeiro Brazil). Dulbecco’s-modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were from Vitrocell Embriolife (Campinas, SP, Brazil). Antibiotic (penicillin/streptomycin), bovine serum albumin (BSA), palmitate (P5585) and 3-(4,5-dimethylthiazol-2-yI)-2,5-diphenyltetrazolium bromide (MTT) were obtained from Sigma (St. Louis, MO, EUA). Trizol, Platinum SYBR Green qPCR SuperMix UDG and Taqman primers and probes were obtained from Life Technologies (Carlsbad, CA, USA), ImProm-II Reverse Transcriptase from Promega (Madison, WI, USA). Anti-GLUT4 (#07–1404) antibody was obtained from Millipore (Billerica, MA, USA), and anti-phospho-IKK-A/B (#2681) from Cell Signaling (Beverly, MA, USA). Anti phospho-PERK (Thr 981) (sc-32577), anti-PERK (H-300) (sc-13073), anti- eIF-2A (FL-315) (sc-11386), anti- IRE1a (H-190) (sc-20790), anti-PR15A (C-19) (sc-825), anti-DDIT-3 (F-168) (sc-575), anti-TRAF2 (C-20) (sc-876), and anti-NFKB p50 subunit (C-19) (sc-1190) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Anti-phospho-eIF-2A (Ser 51) (ab 32157), anti-XBP1 (ab 37152), anti-NFKB p65 subunit (ab 7970), and anti-GRP78 (ab 108615) antibodies were from Abcam (Cambridge, MA, USA), and horseradish peroxidase-linked immunoglobulin was from Amersham Biosciences (Buckinghamshire, UK).
Cell culture and treatment
L6 cells were propagated and differentiated as previously reported [19]. Briefly, cells were propagated in growth medium [DMEM (25 mM glucose), 10% FBS and 1% (v/v) antibiotic] until reach 70% of confluence. Cells were then differentiated for 6 days in differentiation medium [DMEM (25 mM glucose), 2% FBS and 1% (v/v) antibiotic]. At seventh day, cells were incubated with FBS-free restriction/treatment medium [DMEM (5.5 mM glucose), 1% BSA and 1% (v/v) antibiotic] for 12 h before treatment. Palmitate was diluted in ethanol (vehicle) and added to a final concentration of 0.75 mM palmitate and 0.5% ethanol for 2, 6 and 12 h. In parallel, control plates were treated with a non-toxic final concentration of 0.5% ethanol [15, 20].
Cell viability assay
L6 cells were plated in 12-well plate and treated with 0.5% ethanol alone for 12 h or with 0.75 mM palmitate/0.5% ethanol for 2, 6 or 12 h. Cell viability was assessed using MTT reagent according to the manufacturer’s recommendations with some modifications. Briefly, after the incubation with the tested compounds, the medium was changed to a medium with 10% MTT (5 mg/ml) and incubated in dark for 4 h at 37 °C. The medium was removed and replaced with a solvent solution (0.04 N HCl in absolute isopropanol), and the plates were placed on a plate shaker for 15 min to solubilize the formazan crystals derived from MTT reduction. The absorbance of the converted dye was measured at 570 nm with a background subtraction at 690 nm using a microplate reader (BioTek Microquant, BioTek Instruments, Winooski, VT, USA).
Western blotting
Total membrane protein fraction for GLUT4 protein quantification was performed as previously reported [19]. For RE proteins (cytosolic proteins) extraction, L6 cells were removed from plates with PBS and centrifuged at 1500 g for 10 min at 4 °C. The resulting pellet was resuspended with sonication in ice-cold extraction buffer [1% SDS, 100 mM Tris (pH 7.4), 100 mM sodium pyrophosphate, 100 mM sodium fluoride, 10 mM EDTA, 10 mM sodium vanadate, 2 mM PMSF, and 0.1 mg/ml aprotinin], incubated at 96 °C for 10 min. and submitted to centrifugation at 15,000 g for 10 min at 4 °C. The resulting supernatants were frozen at − 20 °C or at − 80 °C. Nuclear proteins for Western blotting analysis were extracted from L6 cells as earlier described [11, 12]. Thirty micrograms of total protein were resolved by dodecyl sulfate polyacrylamide gel electrophoresis (10%T and 2.7%C for GLUT4, NFKB p50, NFKB p65, TRAF2, and XBP1; 12%T and 2.7%C for DDIT-3, phosho-eIF-2A, eIF-2A, IRE1a, phosho-PERK, and PERK; and, 8%T and 2.7%C for GRP78, phosho-IKK-A/B, PR15A), transferred to a nitrocellulose membrane, and immunoblotted for 12 h at 4 °C with specific primary antibody in the following concentrations: anti- DDIT-3 (F-168) (1:500, 1× TBS), anti-eIF-2A (FL-315) (1:500, 1× TBS), anti-phospho-eIF-2A (Ser 51) (1:2000, 1× TBS/3% BSA), anti-GLUT4 (1:3000 in 1× PBS/8% BSA), anti-GRP78 (1:500, 1× TBS/3% BSA), anti-phospho-IKK-A/B (1:500, 1× TBS/5% BSA), anti-IRE1a (H-190) (1:1000, 1× TBS/3% BSA), and anti-NFKB p50 (C-19) (1:500, 1× TBS/1% BSA), anti-NFKB p65 (1:500, 1× TBS/1% BSA), anti-PERK (H-300) (1:1000, 1× TBS/3% BSA), anti-phospho-PERK (Thr 981) (1:1000, 1× TBS/3% BSA), anti-PR15A (C-19) (1:1000, 1× TBS/3% BSA), anti-TRAF2 (C-20) (1:1000, 1× TBS/3% BSA), anti-XBP1 (1:1000, 1× TBS/3% BSA and antibodies. The membrane was then incubated with horseradish peroxidase-linked secondary antibody, and signal was detected by chemiluminescence. Blots were quantified by optical densitometry (ImageScanner III, GE Healthcare, Uppsala, Sweden). Because protocols of protein extraction of membrane (GLUT4), nuclear and cytosolic fractions are different, different approaches for protein-loaded normalization were undertaken. GLUT4 and nuclear protein content were normalized by analyzing post-transferring Coomassie Blue-stained gel [5, 21], and cytosolic protein content was normalized by ACTB content.
Real time RT-PCR
One microgram of total RNA was extracted with Trizol. cDNA, obtained by reverse transcription, was then amplified with Taqman system. Commercially available Taqman primers for rat Gapdh (#Rn99999916_s1) was used for normalization. The primer sequences for rat Slc2a4 were custom designed: 5 ′ GGCTGTGCCATCTTGATGAC-3′ (fw), 5′-CACGATGGACACATAACTCATGGA-3′ (rv) and FAM AACCCGCTCCAGCAGC MGB, as previously described [19].
Eletrophoretic mobility shift assay (EMSA)
Nuclear proteins were extracted from L6 cells and subjected to EMSA as earlier described [12]. The sequence of the oligonucleotide specific for NFKB binding site in Slc2a4 promoter (Slc2a4-NFKB) was 5`-GGGTTGGGGGCGTGGCCTTTTGG-3` [12, 19].
Statistical analysis
Data are expressed as means ± S.E.M. One-way analysis of variance (ANOVA), with Student–Newman–Keuls as a post hoc test was used for analysis of comparison, after comparison of the variances by the Bartelett’s test.
Results
Evaluation of cell viability
The viability of L6 muscle cells incubated with 0.5% ethanol (vehicle) alone or with 0.75 mM palmitate was assessed with MTT assay. As shown in Fig. 1, treatment with palmitate as long as 12 h resulted in 92% of viable cells.

Evaluation of cell viability after exposure to ethanol (vehicle) and palmitate. L6 cells were treated with 0.5% ethanol alone for 12 h or 0.75 mM palmitate in 0.5% ethanol for 2, 6 or 12 h. Cell viability of L6 cells was evaluated by MTT assay. Values are means ± SEM of 12 to 20 samples; one-way ANOVA, Student–Newman–Keuls as post hoc test
Palmitate effects on Slc2a4 gene expression in L6 myotubes
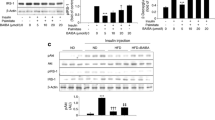
The effect of palmitate treatment on Slc2a4 gene expression was investigated over a time course of 2, 6 and 12 h in L6 myotubes. We have previously demonstrated that 0.75 mM palmitate for 20 h reduces GLUT4 protein content and glucose uptake in L6 cells [18]. Here we showed that 0.75 mM palmitate reduced the Slc2a4 gene transcript content (14, 22 and 16% after 2 h, 6 and 12 h, respectively, P < 0.05 and P < 0.01 versus C; Fig. 2a), and its encoded protein, GLUT4 (33% after 2 h and 6 h and 42% after 12 h, P < 0.01 versus C; Fig. 2b). As it can be observed, the repression effect of palmitate was greater in GLUT4 protein expression than in expression of its respective mRNA, suggesting the presence of a transcriptional and posttranscriptional regulation, such as UPR signaling. To test this hypothesis, the ER stress-mediated pathways were investigated.

Palmitate reduces the expression of the Slc2a4 gene in L6 myotubes. L6 myotubes were treated with 0.5% ethanol (vehicle) for 12 h (control sample, C) or 0.75 mM palmitate for 2, 6 or 12 h. a Relative Slc2a4 mRNA normalized by Gapdh mRNA (RT-qPCR). b Relative GLUT4 protein normalized by total protein analysis with Coomassie Blue-stained gel (Western blotting). Values are means ± SEM of 6 to 10 samples; *P < 0.05 and **P < 0.01 versus C; one-way ANOVA, Student–Newman–Keuls as post hoc test
Palmitate effects on ER stress signaling pathways in L6 cells
L6 muscle cells treated with palmitate for 2 h showed an increase in GRP78 protein content (28%, P < 0.05 versus C, Fig. 3a), which was subsequently dissipated. Thus, two branches of ER stress pathway were investigated including PERK and IRE1.

Palmitate effects on ER stress signaling in L6 cells. L6 myotubes were treated with 0.5% ethanol (vehicle) for 12 h (control sample, C) or 0.75 mM palmitate for 2, 6 or 12 h. Relative cytosolic (a-f, h, j, and k) or nuclear (g and i) protein content. Protein content was normalized by ACTB (a-f, h, j, and k), or by total protein analysis with Coomassie Blue-stained gel (G and I). Values are means ± SEM of 7 to 8 (a), 6 to 7 (b), 8 to 12 (c), 10 to 11 (d), 7 to 8 (e), 4 to 6 (f), 8 to 12 (g), 6 to 9 (h), 9 to 10 (i), 5 to 7 (j), and 8 to 9 (k) samples; *P < 0.05 and **P < 0.01 versus C; #P < 0.05 versus 2 h, §P < 0.05 versus 6 h; one-way ANOVA, Student–Newman–Keuls as post hoc test
PERK activation is the most immediate response to ER stress. In fact, as a consequence of increased GRP78 protein content (Fig. 3a), L6 cells showed enhanced PERK total content (98%, P < 0.01 versus C, Fig. 3b) and phosphorylation (106%, P < 0.001 versus C, Fig. 3c) after 2 h-palmitate treatment. Further, PERK activation is usually followed by phosphorylation of eukaryotic translation initiation factor 2 subunit 1 (eIF-2A); interestingly, while palmitate increased eIF-2A protein content (35, 25 and 39% after 2 h, 6 and 12 h, respectively; P < 0.05 and P < 0.01, versus C; Fig. 3d), it decreased eIF-2A phosphorylation (41, 40 and, 33% after 2, 6 and 12 h, respectively; P < 0.01 versus C; Fig. 3e).
In order to further investigate the discrepancy between the eIF-2A content and phosphorylation, a phosphatase that dephosphorylates eIF-2A, PR15A (protein phosphatase 1 regulatory subunit 15A), also known as growth arrest and DNA damage-inducible protein 34 (GADD34) was analyzed. Surprisingly, palmitate did not exert any effect in PR15A content in L6 cells (Fig. 3f).
The activation of PERK regulates numerous genes involved in apoptosis such as DDIT-3 (DNA damage-inducible transcript 3 protein), also known as growth arrest and DNA damage-inducible protein (GADD153) [22]; thus, the effect of palmitate on the expression of this protein was investigated. Interestingly, palmitate reduced the nuclear content of DDIT-3 3 (66, 60 and 40%, after 2, 6 and 12 h, respectively; P < 0.01 versus C; Fig. 3g) indicating that PERK signaling was not activated downstream.
Next, another pathway of UPR signaling was investigated. In parallel to the increased GRP78 protein content after 2 h-palmitate incubation, the content of protein kinase IRE1a also increased (60%, P < 0.05 versus C, Fig. 3h). The activation of IRE1 results in spliced isoform of XBP1, which positively regulates genes involved in UPR signaling [23]. The present work used an anti-XBP1 antibody that recognizes both spliced and unspliced isoform of XBP1. Here, acute palmitate slightly, but not significantly, increased XBP1 protein content (Fig. 3i).
Alternatively, ER stress can also trigger IRE1a to interact with the kinase adapter tumor necrosis factor receptor-associated factor 2 (TRAF2) and form a complex with inhibitor of nuclear factor kappa-B kinase (IKK) promoting activation of NFKB, independently of IKK activation [24]. Here, it was found that TRAF2 increased (23%, P < 0.05 versus C, Fig. 3j) in L6 myotubes after 2 h-palmitate incubation, albeit IKK phosphorylation was not modified (Fig. 3k).
Participation of NFKB transcription factor in the Slc2a4 gene downregulation by palmitate in L6 muscle cells
Finally, the participation of NFKB in the modulation of Slc2a4 gene by palmitate was investigated.
Recently, we have reported that NFKB downregulates the Slc2a4 gene; in L6 muscle cells, the binding of NFKB dimers to a double-stranded oligonucleotide containing a NFKB site sequence inside the Slc2a4 promoter results in two complexes named a and b [12]. According to our previous report [12], we confirmed here the oligonucleotide binding specificity by competition assay with unlabeled probe (Fig. 4c). Additionally, we have previously confirmed the specific binding of NFKB p50 and p65 subunits in the protein/DNA binding complex in L6 cells. As shown in Fig. 4a and b, treatment with palmitate increased NFKB binding activity after exposure of 2 h (16% for complex a, P < 0.05 versus C; and, 68% for complex b; P < 0.01 versus C), and after 12 h (42% for complex a; P < 0.01 versus C; and, 53% for complex b; P < 0.05 versus C). Additionally, palmitate increased the nuclear content of NFKB p65 in L6 cells (30, 23 and 32% after 2, 6 and 12 h, respectively; P < 0.05 versus C, Fig. 4d).

NFKB participates in Slc2a4 gene downregulation by palmitate in L6 cells. L6 myotubes were treated with 0.5% ethanol (vehicle) for 12 h (control sample, C) or 0.75 mM palmitate for 2, 6 or 12 h. a and b Relative nuclear protein content of complexes a and b (Slc2a4-NFKB); c EMSA competition assay (left panel) and representative image of EMSA assay showing complexes a and b (right panel); d Relative NFKB p65 nuclear protein content normalized by total protein analysis with Coomassie Blue-stained gel. Values are means ± SEM of 10 (a), 9 to 10 (b), and 5 to 10 (d) samples; *P < 0.05 and **P < 0.01 versus C; #P < 0.05 versus 2 h, §P < 0.05 versus 6 h; &P < 0.05 versus 12 h, one-way ANOVA, Student–Newman–Keuls as post hoc test
Discussion
Palmitate is known to induce insulin resistance by imparing glucose uptake [16] and reducing GLUT4 expression [17, 18] in skeletal muscle; nonetheless, little is known about the involved mechanisms.
Here we reported that palmitate reduced GLUT4 expression in L6 muscle cells; still, the correlation of mRNA and protein content was not consistent. Since post-transcriptional modulation has been reported to cause discrepancies between GLUT4 protein and its transcript abundancies [25], it was hypothesized that ER stress could participate in the modulation of the Slc2a4 gene. Actually, it has been reported that activation of UPR signaling represses the Slc2a4 gene in 3 T3-L1 adipocytes [26]. Also, it has been reported that palmitate activates UPR system in hepatocytes [27], pancreatic beta-cells [28], and C2C12 mouse muscle cell [29]. Up to date, only one report has shown that palmitate (1 mM, 24-h treatment) induces ER stress in L6 cells [30]. The present findings showed that lower concentration of palmitate (0.75 mM) acutely (2 h) and transiently induced the expression and activation of proteins related to the initial steps of UPR signaling in L6 myotubes. In fact, palmitate has a weaker effect on UPR compared to tunicamycin, a chemical ER stress inducer in C2C12 myotube [31]. Here, we showed that acute palmitate was able to increase the expression of GRP78 protein, a chaperone positively regulated in stress conditions. When unfolded proteins bind to GRP78, transmembrane proteins from ER such as IRE, PERK and ATF6 are released from GRP78 binding and become activated [14]. Here we investigated two branches of UPR pathway, PERK and eIF-2A.
Acute palmitate exposure resulted in increased expression of PERK and eIF-2A; and surprisingly, while PERK activation was increased, eIF-2A phosphorylation was reduced. Some phosphatases such as PR15A can dephosphorylate eIF-2A, attenuating response to stress and promoting recovery of protein synthesis [32]. In our study, since no alteration on PR15A was observed by palmitate treatment, it is suggested that other factors may be involved in the observed intense reduction of eIF-2A phosphorylation. Besides, the activation of PERK regulates numerous genes involved in apoptosis such as DDIT-3 [22]. Here, we found a decrease in DDIT-3 nuclear content pointing out that acute palmitate, at the present concentration, does not result in apoptosis in L6 myotubes, as confirmed by the unchanged cellular viability observed up to 12 h of culture.
We also demonstrated that acute palmitate even though was able to increase IRE1a expression, it did not result in its activation since phosphorylation of IRE1a and XBP1 nuclear content was not altered. However, our results strongly suggest that IRE1a can contribute to NFKB activation after acute palmitate exposure. It has been reported that basal IKK activity is sufficient to activate NFKB by IRE1a, and this occurs by physical interaction among IRE1a, TRAF2 and IKK [24]. The formation of this complex is transient and can occur in basal state, even if there is no activation of UPR signaling [24]. Albeit increased IKK phosphorylation was not observed after acute palmitate treatment, increased IRE1a and TRAF2 protein expression, besides NFKB p65 activation were observed suggesting that the formation of the complex IRE1a-TRAF2-IKK may activate NFKB in L6 cells after 2 h-palmitate exposure. Besides this non-classical NFKB activation, other IKK phosphorylation-independent pathways have been described. One example of this could be the direct phosphorylation of NFKB p65 by protein kinases such as PKC and PKA, independently of IKK [33,34,35]. Another example could be the activation of NFKB in nuclear factor-kappa-B inhibitor (IKB)-depleted cells [36].
Recently, it has been demonstrated the rat/mouse Slc2a4 gene promoter has a functional kB site to which NFKB binds and downregulates its transcription [12]. Additionally, many studies have reported that reduced GLUT4 expression occurs in insulin resistant states [2,3,4,5]. Thus, NFKB signaling is a prominent molecular signaling that links inflammation to insulin resistance. The present data point out that 2 h- and 12 h-treatment with palmitate increases the nuclear content of NFKB p65 subunit and NFKB binding specifically to the Slc2a4 promoter, and consequently reduces Slc2a/GLUT4 expression. Consistent with these results, we have previously demonstrated that the same concentration of palmitate for 20 h reduces GLUT4 expression and glucose uptake; moreover, it induces tumor necrosis factor gene expression and NFKB accumulation in the nucleus in L6 cells [18]. Our data suggest that while acute palmitate treatment induces NFKB by the formation of IRE1a-TRAF2-IKK complex, chronic treatment effect should occur by some other non-classical pathway. In addition, treatment with palmitate for 6 h also reduced the expression of Slc2a4/GLUT4 in L6 cells, but did not increase NFKB binding in Slc2a4 gene promoter. Of note, a different pathway should be involved.
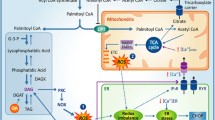
In summary, the data reveal that short-term exposition of muscle cells to palmitate can repress Slc2a4/GLUT4 expression, in a ER- and inflammatory-stress mediated way; thus, contributing to induce insulin resistance (Fig. 5). Considering that, we highlight that the increased consumption of fats, as observed in Western diet and processed foods, may contribute to acute and intermittent increases in circulant FFA levels, favoring the establishment or worsening of muscle metabolic disorders related to inflammation, insulin resistance and diabetes onset.

Participation of ER stress in palmitate-induced inflammation and insulin resistance in skeletal muscle cells. Acute palmitate treatment induces the physical interaction among IREa, TRAF2 and IKK, resulting in the formation of IREa-TRAF2-IKK complex. This complex activates NFKB nuclear factor, which translocates to the nucleus and binds to the Slc2a4 gene promoter, downregulating it and leading to a decreased expression of GLUT4 protein, and, consequently to insulin resistance. In vivo, this mechanism could be triggered by high-fat meals, favoring the establishment or the worsening of muscle metabolic disorders related to insulin resistance
Conclusion
Palmitate reduces Slc2a4/GLUT4 expression in L6 muscle cells. That is related to formation of a IRE1a-TRAF2-IKK complex, activation of NFKB and repression of Slc2a4 gene transcription. These findings indicate that ER stress may connect palmitate-induced inflammation and insulin resistance in L6 muscle cells.
Abbreviations
- BSA:
-
Bovine serum albumin
- DMEM:
-
Dulbecco’s-modified Eagle’s medium
- EMSA:
-
Electrophoretic mobility shift assay
- ER:
-
Endoplasmic reticulum
- FBS:
-
Fetal bovine serum
- FFA:
-
Free fatty acids
- MTT:
-
3-(4,5-dimethylthiazol-2-yI)-2,5-diphenyltetrazolium bromide)
- UPR:
-
Unfolded protein response
- ᅟ:
-
Acronyms used for genes and proteins were based on the Rat Genome Database (RGD) and the Protein Knowledgebase (UniProtKB) respectively
- Actb :
-
Actin beta
- Gapdh :
-
Glyceraldehyde-3-phosphate dehydrogenase
- Slc2a4 :
-
Solute carrier family 2 member 4
- ACTB:
-
Actin cytoplasmic 1 (former beta-actin)
- ATF6:
-
Cyclic AMP-dependent transcription factor (indiscriminate subunit)
- DDIT-3:
-
DNA damage-inducible transcript 3 protein (former growth arrest and DNA damage-inducible protein 153, GADD153)
- eIF-2A:
-
Eukaryotic translation initiation factor 2 subunit 1
- GLUT4:
-
Glucose transporter 4; GRP78, 78 kDa glucose-regulated protein
- IKB:
-
Nuclear factor-kappa-B inhibitor (former IκB)
- IKK-A/B:
-
Inhibitor of nuclear factor kappa-B kinase subunits alpha/beta (former IKKα/β)
- NFKB:
-
Nuclear factor NF-kappa-B (indiscriminate isoform)
- PR15A:
-
Protein phosphatase 1 regulatory subunit 15A (former growth arrest and DNA damage-inducible protein 34, GADD34)
- IRE1:
-
Serine/threonine-protein kinase/endoribonuclease IRE1
- PERK:
-
Protein kinase R-like endoplasmic reticulum kinase
- PKA:
-
cAMP-dependent protein kinase catalytic subunit
- PKC:
-
Protein kinase C
- TRAF2:
-
Tumor necrosis factor receptor-associated factor 2
- XBP1:
-
X Box Protein-1
References
DeFronzo RA, Tripathy D. Skeletal Muscle Insulin Resistance Is the Primary Defect in Type 2 Diabetes. Diabetes Care. 2009;32(Suppl 2):157–63.
Papa PC, Seraphim PM, Machado UF. Loss of weight restores GLUT 4 content in insulin-sensitive tissues of monosodium glutamate-treated obese mice. Int J Obes Relat Metab Disord. 1997;21:1065–70.
Furuya DT, Poletto AC, Favaro RR, Martins JO, Zorn TM, Machado UF. Anti-inflammatory effect of atorvastatin ameliorates insulin resistance in monosodium glutamate-treated obese mice. Metabolism. 2010;59:395–9.
Poletto AC, Anhê GF, Eichler P, Takahashi HK, Furuya DT, Okamoto MM, Curi R, Machado UF. Soybean and sunflower oil-induced insulin resistance correlates with impaired GLUT4 protein expression and translocation specifically in white adipose tissue. Cell Biochem Funct. 2010;28:114–21.
Poletto AC, David-Silva A, Yamamoto AP, Machado UF, Furuya DT. Reduced Slc2a4/GLUT4 expression in subcutaneous adipose tissue of monosodium glutamate obese mice is recovered after atorvastatin treatment. Diabetol Metab Syndr. 2015;7:18.
Stenbit AE, Tsao TS, Li J, Burcelin R, Geenen DL, Factor SM, Houseknecht K, Katz EB, Charron MJ. GLUT4 heterozygous knockout mice develop muscle insulin resistance and diabetes. Nat Med. 1997;3:1096–101.
Brozinick JT Jr, McCoid SC, Reynolds TH, Nardone NA, Hargrove DM, Stevenson RW, Cushman SW, Gibbs EM. GLUT4 overexpression in db/db mice dose-dependently ameliorates diabetes but is not a lifelong cure. Diabetes. 2001;50:593–600.
Schmidt MI, Duncan BB. Diabesity: an inflammatory metabolic condition. Clin Chem Lab Med. 2003;41:1120–30.
Shoelson SE, Lee J, Goldfine AB. Inflammation and insulin resistance. J Clin Invest. 2006;116:1793–801.
Silva JL, Giannocco G, Furuya DT, Lima GA, Moraes PA, Nachef S, Bordin S, Britto LR, Nunes MT, Machado UF. NF-kappaB, MEF2A, MEF2D and HIF1-a involvement on insulin- and contraction-induced regulation of GLUT4 gene expression in soleus muscle. Mol Cell Endocrinol. 2005;240:82–93.
Furuya DT, Poletto AC, Freitas HS, Machado UF. Inhibition of cannabinoid CB1 receptor upregulates Slc2a4 expression via nuclear factor-κB and sterol regulatory element-binding protein-1 in adipocytes. J Mol Endocrinol. 2012;49(2):97–106.
Furuya DT, Neri EA, Poletto AC, Anhê GF, Freitas HS, Campello RS, Rebouças NA, Machado UF. Identification of nuclear factor-κB sites in the Slc2a4 gene promoter. Mol Cell Endocrinol. 2013;370:87–95.
Campello RS, Alves-Wagner AB, Lucas TF, Mori RC, Furuya DT, Porto CS, Machado UF. Estrogen receptor 1 agonist PPT stimulates Slc2a4 gene expression and improves insulin-induced glucose uptake in adipocytes. Curr Top Med Chem. 2012;12:2059–69.
Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP and ER stress transducers in the unfolded-protein response. Nat Cell Biol. 2000;2:326–32.
Lambertucci RH, Hirabara SM, Silveira LR, levada-Pires AC, Curi R, Pithon-Curi TC. Palmitate increases superoxide production through mitochondrial electron transport chain and NADPH oxidase activity in skeletal muscle cells. J Cell Physiol. 2008;216:796–804.
Tsuchiya Y, Hatakeyama H, Emoto N, Wagatsuma F, Matsushita S, Kanzaki M. Palmitate-induced down-regulation of sortilin and impaired GLUT4 trafficking in C2C12 myotubes. J Biol Chem. 2010;285:34371–81.
Jové M, Planavila A, Sánchez RM, Merlos M, Laguna JC, Vázquez-Carrera M. Palmitate induces tumor necrosis factor-alpha expression in C2C12 skeletal muscle cells by a mechanism involving protein kinase C and nuclear factor-kappaB activation. Endocrinology. 2006;147:552–61.
Anhê GF, Okamoto MM, Kinote A, Sollon C, Lellis-Santos C, Anhê FF, Lima GA, Hirabara SM, Velloso LA, Bordin S, Machado UF. Quercetin decreases inflammatory response and increases insulin action in skeletal muscle of ob/ob mice and in L6 myotubes. Eur J Pharmacol. 2012;689:285–93.
Poletto AC, Furuya DT, David-Silva A, Ebersbach-Silva P, Santos CL, Corrêa-Giannella ML, Passarelli M, Machado UF. Oleic and linoleic fatty acids downregulate Slc2a4/GLUT4 expression via NFKB and SREBP1 in skeletal muscle cells. Mol Cell Endocrinol. 2015;401:65–72.
Haber EP, Hirabara SM, Gomes AD, Curi R, Carpinelli AR, Carvalho CR. Palmitate modulates the early steps of insulin signalling pathway in pancreatic islets. FEBS Lett. 2003;544:185–8.
Ferguson RE, Carroll HP, Harris A, Maher ER, Selby PJ, Banks RE. Housekeeping proteins: a preliminary study illustrating some limitations as useful references in protein expression studies. Proteomics. 2005;5:566–71.
Malhotra JD, Kaufman RJ. The endoplasmic reticulum and the unfolded protein response. Semin Cell Dev Biol. 2007;18:716–31.
Hetz C, Glimcher LH. Fine tuning of the Unfolded Protein Response: Assembling the IRE1α interactome. Mol Cell. 2009;35:551–61.
Tam AB, Mercado EL, Hoffmann A, Niwa M. ER Stress Activates NF-κB by Integrating Functions of Basal IKK Activity, IRE1 and PERK. PLoS One. 2012;7:e45078.
Seraphim PM, Nunes MT, Giannocco G, Machado UF. Age related obesity-induced shortening of GLUT4 mRNA poly(A) tail length in rat gastrocnemius skeletal muscle. Mol Cell Endocrinol. 2007;276:80–7.
Miller RS, Diaczok D, Cooke DW. Repression of GLUT4 expression by the endoplasmic reticulum stress response in 3T3-L1 adipocytes. Biochem Biophys Res Commun. 2007;362:188–92.
Achard CS, Laybutt DR. Lipid-induced endoplasmic reticulum stress in liver cells results in two distinct outcomes: adaptation with enhanced insulin signaling or insulin resistance. Endocrinology. 2012;153:2164–77.
Green CD, Olson LK. Modulation of palmitate-induced endoplasmic reticulum stress and apoptosis in pancreatic β-cells by stearoyl-CoA desaturase and Elovl6. Am J Physiol Endocrinol Metab. 2011;300:640–9.
Peng G, Li L, Liu Y, Pu J, Zhang S, Yu J, Zhao J, Liu P. Oleate blocks palmitate-induced abnormal lipid distribution, endoplasmic reticulum expansion and stress, and insulin resistance in skeletal muscle. Endocrinology. 2011;152:2206–18.
Yuzefovych LV, LeDoux SP, Wilson GL, Rachek LT. Mitochondrial DNA damage via augmented oxidative stress regulates endoplasmic reticulum stress and authophagy: crosstalk, links and signaling. PLoS One. 2013;8:83349.
Hage Hassan R, Hainault I, Vilquin JT, Samama C, Lasnier F, Ferré P, Foufelle F, Hajduch E. Endoplasmic reticulum stress does not mediate palmitate-induced insulin resistance in mouse and human muscle cells. Diabetologia. 2012;55:204–14.
Brush MH, Weiser DC, Shenolikar S. Growth arrest and DNA damage-inducible protein GADD34 targets protein phosphatase 1 alpha to the endoplasmic reticulum and promotes dephosphorylation of the alpha subunit of eukaryotic translation initiation factor 2. Mol Cell Biol. 2003;23:1292–303.
Zhong H, SuYang H, Erdjument-Bromage H, Tempst P, Ghosh S. The transcriptional activity of NF-kappaB is regulated by the IkappaB-associated PKAc subunit through a cyclic AMP-independent mechanism. Cell. 1997;89:413–24.
Ghosh S, Karin M. Missing pieces in the NF-kappaB puzzle. Cell. 2002;109(Suppl):81–96.
Mut M, Amos S, Hussaini IM. PKC alpha phosphorylates cytosolic NF-kappaB/p65 and PKC delta delays nuclear translocation of NF-kappaB/p65 in U1242 glioblastoma cells. Turk Neurosurg. 2010;20:277–85.
Tergaonkar V, Correa RG, Ikawa M, Verma IM. Distinct roles of IkappaB proteins in regulating constitutive NF-kappaB activity. Nat Cell Biol. 2005;7(9):921–3.
Acknowledgements
This research was supported by grants from FAPESP (São Paulo State Foundation for Research): 2010/09984-5, 2013/18841-1 and 2016/15603-1.
Availability of data and materials
All data generated during the current study are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
PES, UFM, GFA and PMS designed the study. PES, ACP and ADS conducted the experiments. PES, UFM and DTF analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Ebersbach-Silva, P., Poletto, A.C., David-Silva, A. et al. Palmitate-induced Slc2a4/GLUT4 downregulation in L6 muscle cells: evidence of inflammatory and endoplasmic reticulum stress involvement. Lipids Health Dis 17, 64 (2018). https://doi.org/10.1186/s12944-018-0714-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12944-018-0714-8