
Abstract
Background
Apolipoprotein E (APOE) plays a major role in lipid metabolism and inflammation. However, the association between APOE gene polymorphisms and serum triglyceride levels remains controversial. We tested the effects of APOE variants on triglyceride levels and their interactions with the inflammatory marker C-reactive protein (CRP) in a Taiwanese population.
Methods
Two APOE single nucleotide polymorphisms (SNPs) rs429358 and rs7412 were genotyped by TaqMan Assay using real time PCR in 595 healthy subjects attending the clinic for routine visits.
Results
After adjustment for clinical covariates, subjects carrying the rs429358-TT genotype and non-ε4 alleles were found to have higher CRP levels, whereas those with rs7412-CC genotype and non-ε2 alleles had significantly higher total and low-density lipoprotein cholesterol levels (all P < 0.01). Using subgroup and interaction analyses, we observed significantly lower triglyceride levels in subjects carrying the rs429358-TT genotype and non-ε4 alleles in the low CRP group (P = 2.71× 10−4 and P = 4.32 × 10−4, respectively), but not in those in the high CRP group (interaction P = 0.013 and 0.045, respectively). In addition, multivariate stepwise linear regression analysis showed that subjects carrying the rs429358-TT genotype and non-ε4 alleles with low CRP levels had significantly lower triglyceride levels (P < 0.001 and P < 0.001, respectively). In addition, when combined with the risk alleles of GCKR, APOA5 and LPL gene variants, we observed that triglyceride levels increased significantly with the number of risk alleles (P = 2.9 × 10−12).
Conclusions
The combination of SNPs and ε alleles at the APOE locus is involved in managing lipid and CRP levels in the Taiwanese population. APOE polymorphisms interact with CRP to regulate triglyceride levels, thus triglyceride concentration is influenced by both the genetic background of the APOE locus and the inflammatory status of a subject.
Similar content being viewed by others

Background
Human apolipoprotein E (APOE) is a 299-amino acid polypeptide responsible for lipid metabolism and cholesterol and triglyceride transport in the blood [1]. APOE is synthesized and secreted by the hepatocytes and is a surface component of primarily triglyceride-rich lipoproteins, such as very low-density lipoproteins (VLDLs), their remnants, chylomicron remnants, and high density lipoproteins (HDLs) [1]. APOE exerts effects by binding to its own receptor and the low-density lipoprotein (LDL) receptor and is the main ligand for clearance of VLDLs and chylomicron remnants, and as such affects circulating concentrations of lipoproteins and plasma levels of cholesterol and triglycerides [2, 3]. Moreover, it was shown that apoE has proinflammatory properties and mediates the presentation of lipid antigens to the immune system and in this way lead to chronic inflammation [4, 5]. In addition, APOE transcripts are found in the brain, kidneys, and spleen, suggestive of a crucial function of APOE in these organs [6–8]. APOE knockout mice developed severe hypercholesterolemia and accelerated the development of atherosclerotic lesions [9]. In humans, the APOE variants are known for their roles in atherosclerosis, cardiovascular disease, Alzheimer disease, aging, and longevity [10–12]. Three APOE isoforms are encoded by three codominant alleles, ε2, ε3, and ε4, which are the direct results of two amino acid substitutions at positions 112 and 158. Therefore, depending on an individual’s genotype, three homozygous (APOE2/2, APOE3/3, and APOE4/4) and three heterozygous (APOE2/3, APOE2/4, and APOE3/4) isoform combinations are present in the human population [13].
Many clinical investigations have shown that serum triglyceride level is an excellent indicator of inflammation, insulin resistance, metabolic syndrome, atherosclerosis, and coronary heart disease (CHD) [14–17]. Previous genome-wide association studies (GWAS) and meta-analyses have mapped the genetic determinants of lipid levels, including that of triglycerides, to several loci enriched with single nucleotide polymorphisms (SNPs) [18]. However, GWAS and fine-mapping studies on the sets of APOE SNPs responsible for the lipid traits have yielded conflicting results. Some APOE SNPs have been discovered by GWAS; however, fine-mapping studies have suggested a different set of SNPs, which vary substantially in frequency and effect size [18, 19]. Many studies have discovered factors that interact with APOE gene polymorphisms in regulating physiological functions. One such factor is C-reactive protein (CRP), an indispensable biomarker for chronic systemic inflammation. Serum CRP levels considerably influenced the relationship between APOE SNPs and disease states [20]. Previous studies have found elevated CRP levels associated with hyperlipidemias [21, 22]. CRP gene variants have been independently associated with serum lipid levels [23] and individuals carrying the risk allele for CRP also encounter higher lipid levels and risk of type 2 diabetes [24]. These results suggested CRP may play a role in lipid metabolism.
Since APOE is a multifunctional protein involving lipid metabolism and inflammatory process, this study aimed to investigate the role of APOE variants on CRP and lipid levels as well as the interactive effect of CRP levels. Apolipoprotein A-V (APOA5), Lipoprotein lipase (LPL) and glucokinase regulator (GCKR) gene region variants have been consistently associated with serum triglyceride levels [25–27]. The independent effect of APOE SNP on triglyceride levels and the genetic risk allele analysis were also investigated with the LPL, GCKR and ApoA5 loci variants.
Methods
Subjects
In total, 595 patients (312 men, mean age: 44.9 ± 10.4 years; 283 women, mean age: 46.6 ± 10.0 years) were recruited between October 2003 and September 2005at Chang Gung Memorial Hospital. During routine cardiovascular health examinations, the participants answered a questionnaire on medical history and lifestyle characteristics. They underwent physical examinations in which height, weight, waist and hip circumferences, and blood pressure in the sitting position after 15 min of rest were measured. Fasting blood samples, including serum and plasma, were obtained from each patient via the antecubital vein, and were then centrifuged at 3000 × g for 15 min at 4 °C. All patients provided informed consent. Exclusion criteria included a history of myocardial infarction, stroke or transient ischemic attack, cancer, current renal or liver disease. Furthermore, to avoid enrolling patients with acute inflammatory disease, those with CRP levels of >10 mg/L [28] and use of lipid-lowering medication relate to variation in CRP and lipid levels were excluded. The clinical and biometrical features of the study population are summarized in Table 1. Current smokers were those who smoked regularly during the survey period. This study was approved by the Ethics Committees of Chang Gung Memorial Hospital (92–315, 6 May 2003).
Genomic DNA extraction and genotyping
Genomic DNA was extracted as previously reported [29]. Two APOE SNPs, rs429358 (TGC → CGC, Cys112Arg) and rs7412 (CGC → TGC, Arg158Cys), were selected according to the NCBI SNP database (http://www.ncbi.nlm.nih.gov/SNP) for genotyping APOE isoforms and were performed using TaqMan SNP Genotyping Assays from Applied Biosystems (ABI; Foster City, CA, USA). The two APOE SNPs were further stratified into three common alleles, ɛ2, ɛ3 and ɛ4: ɛ2, T-T (Cys-Cys); ɛ3, T-C (Cys-Arg); and ɛ4, C-C (Arg-Arg) [13]. Genotyping for GCKR rs1260326 and LPL rs13702 polymorphisms were also performed using TaqMan SNP Genotyping Assays from Applied Biosystems. Genotyping for APOA5 rs662799 was performed with polymerase chain reaction and restriction enzymes digestion. Genotyping data are shown in Additional file 1: Table S1. For quality control purposes, approximately 10 % of the samples were re-genotyped in a blinded fashion and the same results were obtained.
Assays
The levels of most markers, including serum CRP, serum amyloid A (SAA), soluble intercellular adhesive molecule (sICAM1), soluble vascular cell adhesive molecule (sVCAM1), soluble E-selectin (sE-selectin), and matrix metalloproteinase 9 (MMP-9), were measured using a sandwich enzyme-linked immunosorbent assay (ELISA) developed in-house. All in-house kits showed excellent correlation when compared with commercially available ELISA kits [30–32]. Circulating serum matrix metalloproteinase 2 (MMP-2) levels were measured using a commercially available ELISA kit (R&D; Minneapolis, MN, USA). Plasma glucose levels were measured in a central laboratory as previously reported [33]. Total cholesterol and triglyceride levels were measured through automatic enzymatic colorimetry. High-density lipoprotein cholesterol (HDL-C) levels were measured enzymatically after phosphotungsten and magnesium precipitation. Low-density lipoprotein cholesterol (LDL-C) levels were measured using the Friedewald formula for patients with triglyceride levels of <400 mg/dL. When triglyceride levels were >400 mg/dL, serum LDL-C levels were measured with commercial reagents following standard protocol. Plasma fibrinogen levels were measured using the Clauss method adapted for a Sysmex CA1-1500 instrument in the clinical hematology laboratory.
Statistical analyses
The chi-square test was used to examine differences in the distribution of categorical data including smoking status, hypertension, hypercholesterolemia, and diabetes mellitus. Clinical characteristics that were continuous variables were expressed as means ± SD and tested using the two-sample t test or analysis of variance (ANOVA). A generalized linear model was used to analyze lipid and inflammatory marker levels regarding investigated genotypes and confounders. Dominant models were used for numeric association tests after recoding our SNPs from categorical variables to continuous variables, such as 0 and 1 of a particular allele. Triglyceride, HDL-C, LDL-C, total cholesterol, CRP, SAA, sICAM1, sVCAM1, sE-selectin, MMP-2, and MMP-9 level values were logarithmically transformed before statistical analyses to follow a normality assumption. P < 0.05, obtained using two-sided tests, was considered statistically significant. Interactions between each SNP, the levels of lipid traits or CRP, and the CRP status were tested using two-way ANOVA. When interaction terms were significant, stratified analyses of the APOE genotypes (i.e., genotypes affected by the CRP status) and lipid or CRP levels were performed to further study interactive effects while controlling for other variables including age, smoking status, BMI, and antihypertensive and antidiabetic medication use. In addition, stepwise linear regression analysis was performed to determine independent predictors of triglyceride levels. Deviation from the Hardy–Weinberg equilibrium (HWE), the linkage disequilibrium between polymorphisms, the association of genotypes with lipid or CRP levels, and genotype–CRP status interactions were investigated using Golden Helix SVS Win32 7.3.1 software (Golden Helix). The genetic risk alleles were created to assume each SNP to be independently associated with triglyceride levels (i.e., no interaction between SNPs). The count model assumed each SNP in the panel contributed equally to triglyceride levels, and the genetic risk allele was determined by a simple summation of the number of risk alleles from the GCKR, APOA5, LPL SNPs and APOE − CRP subgroups.
Results
Characteristics of clinical biochemical factors and biomarkers
Demographic data, clinical biochemical data, and lipid and inflammatory biomarker profiles of the subjects, stratified by sex, are summarized in Table 1. A significantly higher percentage of the men were current smokers (P < 0.001) and hypertensive (P = 0.006). In addition, the men had significantly higher body mass index (BMI) (P < 0.001), diastolic blood pressure (P < 0.001), fasting plasma glucose (P = 0.01), triglyceride (P < 0.001), sE-selectin (P < 0.001), and MMP-9 (P = 0.011) levels than the women did. By contrast, HDL-C levels (P < 0.001) were lower in the men than in the women. No significant deviation from the HWE was observed for the studied polymorphisms (Additional file 1: Table S1, P = 0.812 and 0.935 for rs429358 and rs7412, respectively).
Relationship between APOE SNPs and lipid biomarkers
To determine the effects of APOE SNPs on lipid levels, we created an additive model using four lipid traits as variables of interest. After adjustment for age, sex, smoking status, BMI, and antihypertensive and antidiabetic treatments, subjects carrying rs429358-TT genotype had lower LDL-C levels and higher HDL-C levels than those carrying rs429358-CT and –CC genotypes did (Table 2, P = 0.025 and 0.040, respectively). The subjects carrying rs429358-CT genotypes had the highest triglyceride levels compared to that of the other two carriers (P = 0.034). Among rs7412 carriers, subjects carrying rs7412-CC genotype had the highest total cholesterol and LDL-C levels compared to those carrying the rs7412-CT and TT genotypes (Table 2, P = 3.01× 10−5 and 4.55× 10−9, respectively). Using a recessive model, we discovered that subjects carrying rs429358-CT and –CC genotypes had higher LDL-C and triglyceride levels than those carrying the rs429358-TT genotype did (P = 0.043 and 0.027, respectively), whereas subjects carrying rs7412-CT and TT genotypes had lower total cholesterol and LDL-C levels compared to those carrying rs7412-CC genotype (P = 3.2 × 10−5 and 3.1 × 10−8, respectively), which largely confirmed most of the findings from the additive model.
Association between APOE polymorphisms and inflammatory biomarkers
To determine whether the APOE genotypes influenced any inflammatory marker, we analyzed the levels of eight inflammatory markers, CRP, fibrinogen, SAA, sICAM1, sVCAM1, sE-selectin, MMP-2, and MMP-9. Except for CRP levels, no significant differences were observed in inflammatory marker levels among the APOE genotypes (Additional file 1: Table S2). Table 2 shows variation in serum CRP levels across the APOE genotypes. After an additive model was adjusted for age, sex, smoking status, BMI, and antihypertensive and antidiabetic medication use, only rs429358 genotypes was significantly associated with CRP levels (P = 0.005). In addition, a dominant model adjusted for age, sex, smoking status, BMI, and antihypertensive and antidiabetic medication use revealed that subjects carrying rs429358-CC and -CT genotypes were associated with lower CRP levels than those carrying TT genotype did (P = 3.8 × 10−4) (Table 2). In contrast, none of the rs7412 genotypes were associated with CRP levels.
APOE allele combinations, lipids, and CRP
We assigned APOE allele combinations into three groups, ε2 (ε2ε2 and ε2ε3), ε3 (ε3ε3), and ε4 (ε4ε4 and ε4ε3), and used multiple linear regressions to analyze their effects on lipid and CRP levels. After adjustment for age, sex, smoking status, BMI, and antihypertensive and antidiabetic treatments using an additive model, different allele groups were shown as exerting significant differences in the levels of total cholesterol, LDL-C, HDL-C, and CRP (Table 3, P = 4.24× 10−6, 1.94× 10−7, 0.008, and 0.018, respectively). The ε2 carriers had significantly lower total cholesterol and LDL-C levels than the non-ε2 carriers did (P = 6.65 × 10−6 and 5.12 × 10−9, respectively). The ε4 carriers had significantly higher LDL-C and triglyceride levels (P = 0.031 and 0.018, respectively) and lower HDL-C and CRP levels than the non-ε4 carriers did (P = 0.020 and 2.12× 10−4, respectively).
APOE and CRP interaction
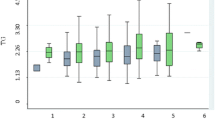
Because we were interested in the effects of CRP–APOE interaction on lipid levels, we divided the subjects according to their CRP levels and APOE genotypes, and analyzed the triglyceride levels among different APOE genotypes and allele carriers in the low and high CRP groups (Fig. 1). After adjustment for clinical covariates, ε4 carriers exhibited significantly higher triglyceride levels exclusively in the low CRP group (P = 4.32× 10−4), and interaction was observed between CRP levels and different APOE allele carrier groups (interaction P = 0.045). Similarly, subjects carrying the rs429358-CC and CT genotypes had higher triglyceride levels than those carrying rs429358-TT genotypes did in the low CRP group (P = 2.71× 10−4). None of these observations were seen in the high CRP group.

The triglyceride levels according to rs429358 genotypes a, b and APOE isoforms c, d in Taiwanese subjects in high and low C-reaction protein (CRP) subgroups. a After adjusting for clinical covariates, subjects carrying the APOE rs429358-TT genotype had lower triglyceride levels compared to those carrying non-TT genotypes in the low CRP subgroup but not in the high CRP subgroup. b Significantly lower triglyceride levels were also noted when subjects with the APOE rs429358-TT genotype and low CRP levels were compared to the other subjects of the study. c, d Similar results were also noted when triglyceride levels from carriers of the APOE ε4 and non-ε4 isoforms were compared. Low CRP level was defined as CRP levels lower or equal to 0.62 mg/L, whereas high CRP level was defined as CRP levels higher than 0.62 mg/L
Stepwise regression on triglyceride levels
Because we observed an interaction between APOE genotypes and CRP, we next studied the effects of APOE − CRP subgroups on triglyceride by using stepwise regression to adjust for smoking status, BMI, age, and the three LPL, GCKR and ApoA5 SNPs that were independent variables associated with triglyceride levels in previous studies [25–27]. Our results also showed that the studies LPL, GCKR and ApoA5 SNPs were significantly associated triglyceride levels (Additional file 1: Table S3). The effect of the non-ε4 − low CRP subgroup on triglyceride levels was significant (β: 0.087, P < 0.001) (Table 4). Similar results were observed for the rs429358-TT − low CRP subgroup (β: 0.081, P < 0.001) (Table 4). When CRP levels were included in the independent variables, the effect of the non-ε4 − low CRP or rs429358-TT − low CRP subgroup on triglyceride levels was still significant (data not shown). We next assayed the combined effects of risk alleles of GCKR, APOA5, LPL, and APOE − CRP subgroups on triglyceride levels (Fig. 2). We observed that triglyceride levels increased significantly with the number of risk alleles (P = 2.9 × 10−12).

The triglyceride levels according to number of risk alleles calculated from number of SNPs of the GCKR, APOA5, and LPL genes, and the APOE − CRP subgroups. The triglyceride levels were as follows: no risk allele (2.3 % of the population): 87.7 ± 34.2 mg/dL; one risk allele (40.8 % of the population): 113.3 ± 109.3 mg/dL; two risk alleles (42.0 % of the population): 154.9 ± 105.3 mg/dL; three risk alleles (14.4 % of the population): 197.6 ± 167.5 mg/dL; three or four risk alleles (0.5 % of the population): 267.3 ± 84.7 mg/dL
Discussion
This investigation analyzed the association and interaction among APOE genotypes, various lipids including triglycerides, and the inflammatory marker CRP in a Taiwanese population. First, regarding the relationship between APOE SNPs and lipid levels, our data revealed significant associations between various genotypes or allelic combinations of the APOE SNP rs7412 and total cholesterol and LDL-C levels. Borderline significance (i.e., 0.01 < P < 0.05) was noted between various rs429358 genotypes and triglyceride, LDL-C, and HDL-C levels. Regarding the relationship between various APOE ε alleles and lipid levels, we observed significant differences in total cholesterol, LDL-C, and HDL-C levels among the carriers of the three APOE ε allele groups. In addition, when ε2 and ε4 were analyzed, we found significant differences in total cholesterol and LDL-C levels between the ε2 and non-ε2 carriers. Second, regarding the relationship between APOE and CRP, our data revealed significant associations between various alleles or allelic combinations of the APOE SNP rs429358 and CRP levels and a significant difference in CRP levels between the ε4 and non-ε4 carriers. Finally, an interaction was discovered between CRP and APOE, and differences in triglyceride levels were clearly evident between the carriers of different APOE genotypes when stratified by low CRP status. In univariate and multivariate analyses, significant differences were found in triglyceride levels in carriers of the rs429358-TT genotype or non-ε4 alleles from the low CRP group compared with the triglyceride levels in the other carriers. The association between APOE-CRP subgroups and triglyceride levels was independent of SNPs of APOA5, GCKR, and LPL. However, these risk alleles of APOA5, GCKR, and LPL interacted with APOE-CRP subgroups synergistically to increase triglyceride levels. These results will certainly increase our understanding of the involvement of APOE in cardiovascular diseases.
APOE plays a major role in lipid metabolism and inflammatory response. Our results regarding lipid metabolism echo those of past studies that have shown that ε4 carriers have high total cholesterol and LDL-C levels [34–36]. Regarding inflammation, our results also support previous results showing that ε4 carriers have low CRP levels [37–40]. In addition to these genotype-dependent effects on lipids and CRP levels, ε4 is associated with infectious diseases [41] and a high risk of age-related cardiovascular and Alzheimer disease [42, 43]. The differential regulation of lipids and CRP, in particular by APOE ε4, could be explained by a structural change caused by a cysteine-to-arginine substitution at residue 112 [37], down regulation of the mevalonate/cholesterol synthetic pathway in ε4 carriers [39], or different patterns of linkage disequilibrium between ε4 and other SNPs that are yet to be characterized [44].
The association between APOE polymorphisms and triglyceride levels is particularly complicated because many studies, such as our initial comparison among the ε2, ε3, ε4 carriers, yield negative results. Nevertheless, some studies have reported positive findings, particularly for the ε4 variant. Dong et al. (2013) showed that ε4 was associated with higher triglyceride levels in patients with renal disease [35]. Carvarlho-Wells et al. (2010) reported that ε4 carriers have higher postprandial triglyceride levels only in adults aged >50 years [45]. The results of both studies are consistent with our finding that ε4 carriers have slightly higher triglyceride levels than non-ε4 carriers do. Maxwell et al. (2013) reported that though ε4 negatively affects the hazard ratio of CHD divided by triglycerides, APOE genotypes, overall, do interact with triglycerides to increase the risk of CHD in a linear model of a pooled European-American population, but not an African-American population [46]. According to these studies, each APOE isoform seems to interact with triglyceride metabolism differently, and under various physiological conditions (i.e., age, ethnicity, disease states, and subcategories), these interactions affect triglyceride levels and contribute to the differential odds of the same disease in a multifactorial manner. This may explain why observed odds for coronary disease among ε4 carriers decreases when subjects with different backgrounds were pooled into a substantial sample for meta-analysis [47]. Although we did not measure odd ratios of any disease in this study, an increased difference was observed in triglyceride levels between the ε4 and non-ε4 carriers in the low CRP group, thus providing an acceptable example of the inconsistent regulation of the triglyceride trait. Furthermore, previous studies suggested that the genetic profiles such as the genotype score may be useful for the early detection and treatment of dyslipidemias and triglyceride levels [48, 49]. We also found a cumulative effect in common risk alleles for triglyceride levels. There was a significant association between triglyceride levels and an increasing genotype number of risk alleles. These findings indicated that multiple variants might act together to influence the development of various phenotypes and disease.
Studies have investigated the gene − environment interaction of gene variants on various traits. Qi et al. (2009) reported that the variants of the interleukin-6 receptor SNP rs8192284 modified diabetes risk in women with different CRP levels, and the highest risk was observed in those with the subjects carrying rs8192284-AA genotype in the highest CRP quartile [50]. Our findings provide another example of complex gene–environment interaction, with the subjects carrying APOE rs429358-TT genotype and non-ε4 alleles substantially reducing the levels of triglyceride in subjects with lower CRP concentration. The regulation of triglyceride levels is associated with inflammation and insulin resistance; therefore, increased triglyceride levels for both the ε4 and non-ε4 carriers in the high CRP group are acceptable. Unexpectedly, triglyceride levels remained elevated for the ε4 carriers with a low CRP level. Although an association was observed between ε4 and decreased CRP levels [40, 51], ε4 exhibited proinflammatory activities [52]. Our finding thus suggests that ε4 may, possibly through an alternative pro-inflammatory pathway, regulate triglyceride independently of CRP. By contrast, CRP levels might be upregulated by some proinflammatory factors, directly or indirectly masking the influence of ε4. For example, hs-CRP levels were not considerably different between APOE genotype carriers in Moroccan patients with end-stage renal disease (ESRD) [53], and, in our study, some ε4 carriers exhibited high CRP levels. Although the cause of high CRP levels regardless of APOE genotypes in our sample population is unknown, we suspect that certain health conditions (e.g., ESRD) are responsible for this phenomenon. Further investigation is required to address this possible association.
We propose that the CRP − APOE − triglyceride triad might be a common motif of information processing that bridges clinical manifestations of genetic variants to outputs of signaling units in the body (e.g., systemic inflammation) and generates pleiotropic phenotypes. Its relationship with CHD states requires further investigation, and discovering more loci, covariates, or traits that interact with APOE in a relationship quantitative trait loci pattern [46, 54, 55] might be insightful.
Conclusion
In conclusion, our data revealed a CRP-dependent association between APOE polymorphisms and triglyceride levels. APOE ε4 alleles along with low CRP levels were an independent determinant of triglyceride levels in our multivariate regression analysis. CRP is considered an inflammatory marker, thus we propose that the effects of APOE genotypes on triglyceride levels in the blood might be associated with the inflammatory status of an individual.
Limitations
One limitation of this study is the relatively low number of subjects genotyped; replication of the current results in a second cohort would support the strength of the study. Independent association studies with larger sample size and functional data are required to confirm our results before any definitive conclusions can be drawn. Another limitation is the study’s cross-sectional design, which means that the results may be used to draw only limited inference regarding the relationship between exposure and outcome.
Abbreviations
- AC:
-
antecubital
- APOA5:
-
apoliporotein A-V
- APOE:
-
Apolipoprotein E
- BMI:
-
body mass index
- BP:
-
blood pressure
- CHD:
-
coronary heart disease
- CRP:
-
C-reactive protein
- ESRD:
-
end-stage renal disease
- GCKR:
-
glucokinase regulator
- GWAS:
-
genome-wide association studies
- HDL-C:
-
high-density lipoprotein-cholesterol
- LDL-C:
-
low-density lipoprotein-cholesterol
- LPL:
-
lipoprotein lipase
- MMP2:
-
matrix metalloproteinase 2
- MMP9:
-
matrix metalloproteinase 9
- SAA:
-
serum amyloid A
- sICAM1:
-
soluble intercellular adhesive molecule 1
- SNP:
-
single nucleotide polymorphism
- sVCAM1:
-
soluble vascular cell adhesive molecule 1
References
Weisgraber KH, Mahley RW. Human apolipoprotein E: the Alzheimer’s disease connection. FASEB J. 1996;10:1485–94.
Mahley RW, Ji ZS. Remnant lipoprotein metabolism: Key pathways involving cell-surface heparan sulfate proteoglycans and apolipoprotein E. J Lipid Res. 1999;40:1–16.
Holtzman DM, Herz J, Bu G. Apolipoprotein E and apolipoprotein E receptors: normal biology and roles in Alzheimer disease. Cold Spring Harb Perspect Med. 2012;2:a006312.
van den Elzen P, Garg S, Leon L, Brigl M, Leadbetter EA, et al. Apolipoprotein-mediated pathways of lipid antigen presentation. Nature. 2005;437:906–10.
Mooijaart SP, Berbée JF, van Heemst D, Havekes LM, de Craen AJ, Slagboom PE, et al. ApoE plasma levels and risk of cardiovascular mortality in old age. PLoS Med. 2006;3:e176.
Williams DL, Dawson PA, Newman TC, Rudel LL. Synthesis of apolipoprotein E by peripheral tissues. Potential functions in reverse cholesterol transport and cellular cholesterol metabolism. Ann N Y Acad Sci. 1985;454:222–9.
Elshourbagy NA, Liao WS, Mahley RW, Taylor JM. Apolipoprotein E mRNA is abundant in the brain and adrenals, as well as in the liver, and is present in other peripheral tissues of rats and marmosets. Proc Natl Acad Sci U S A. 1985;82:203–7.
Zhang H, Wu LM, Wu J. Cross-talk between apolipoprotein E and cytokines. Mediators Inflamm. 2011;2011:949072.
Plump AS, Smith JD, Hayek T, Aalto-Setala K, Walsh A, Verstuyft JG, et al. Severe hypercholesterolemia and atherosclerosis in apolipoprotein E-deficient mice created by homologous recombination in ES cells. Cell. 1992;71:343–53.
Huang Y. Mechanisms linking apolipoprotein E isoforms with cardiovascular and neurological diseases. Curr Opin Lipidol. 2010;21:337–45.
Seripa D, D’Onofrio G, Panza F, Cascavilla L, Masullo C, Pilotto A. The genetics of the human APOE polymorphism. Rejuvenation Res. 2011;14:491–500.
Phillips MC. Apolipoprotein E, isoforms and lipoprotein metabolism. IUBMB Life. 2014;66:616–23.
Takei N, Miyashita A, Tsukie T, Arai H, Asada T, Imagawa M, et al. Genetic association study on in and around the APOE in late-onset Alzheimer disease in Japanese. Genomics. 2009;93:441–8.
Shimodaira M, Niwa T, Nakajima K, Kobayashi M, Hanyu N, Nakayama T. Impact of serum triglyceride and high density lipoprotein cholesterol levels on early-phase insulin secretion in normoglycemic and prediabetic subjects. Diabetes Metab J. 2014;38:294–301.
Hadaegh F, Khalili D, Ghasemi A, Tohidi M, Sheikholeslami F, Azizi F. Triglyceride/HDL-cholesterol ratio is an independent predictor for coronary heart disease in a population of Iranian men. Nutr Metab Cardiovasc Dis. 2009;19:401–8.
Patel S, Puranik R, Nakhla S, Lundman P, Stocker R, Wang XS, et al. Acute hypertriglyceridaemia in humans increases the triglyceride content and decreases the anti-inflammatory capacity of high density lipoproteins. Atherosclerosis. 2009;204:424–8.
Labreuche J, Deplanque D, Touboul PJ, Bruckert E, Amarenco P. Association between change in plasma triglyceride levels and risk of stroke and carotid atherosclerosis: systematic review and meta-regression analysis. Atherosclerosis. 2010;212:9–15.
Global Lipids Genetics C, Willer CJ, Schmidt EM, Sengupta S, Peloso GM, Gustafsson S, et al. Discovery and refinement of loci associated with lipid levels. Nat Genet. 2013;45:1274–83.
Sanna S, Li B, Mulas A, Sidore C, Kang HM, Jackson AU, et al. Fine mapping of five loci associated with low-density lipoprotein cholesterol detects variants that double the explained heritability. PLoS Genet. 2011;7.
Aiello AE, Nguyen HO, Haan MN. C-reactive protein mediates the effect of apolipoprotein E on cytomegalovirus infection. J Infect Dis. 2008;197:34–41.
Yin G, Hu G, Cang X, Yu G, Hu Y, Xing M, et al. C-reactive protein: rethinking its role in evaluating the severity of hyperlipidemic acute pancreatitis. Pancreas. 2014; 43:1323-8.
Johnsson H, Panarelli M, Cameron A, Sattar N. Analysis and modelling of cholesterol and high-density lipoprotein cholesterol changes across the range of C-reactive protein levels in clinical practice as an aid to better understanding of inflammation-lipid interactions. Ann Rheum Dis. 2014;73:1495–9.
Wei W, Yang S, Qiu Y, Wang H, Zhao X, Zhao Y, et al. CRP gene polymorphism contributes genetic susceptibility to dyslipidemia in Han Chinese population. Mol Biol Rep. 2014;41:2335–43.
Ligthart S, de Vries PS, Uitterlinden AG, Hofman A, CHARGE Inflammation working group, Franco OH, et al. Pleiotropy among common genetic loci identified for cardiometabolic disorders and C-reactive protein. PLoS One. 2015;10:e0118859.
Varbo A, Benn M, Tybjaerg-Hansen A, Grande P, Nordestgaard BG. TRIB1 and GCKR polymorphisms, lipid levels, and risk of ischemic heart disease in the general population. Arterioscler Thromb Vasc Biol. 2011;31:451–7.
Hsu LA, Ko YL, Chang CJ, Teng MS, Wu S, Hu CF. Apolipoprotein A5 gene -1131 T/C polymorphism is associated with the risk of metabolic syndrome in ethnic Chinese in Taiwan. Clin Chem Lab Med. 2008;46:1714–9.
Tang W, Apostol G, Schreiner PJ, Jacobs Jr DR, Boerwinkle E, Fornage M. Associations of lipoprotein lipase gene polymorphisms with longitudinal plasma lipid trends in young adults: The Coronary Artery Risk Development in Young Adults (CARDIA) study. Circ Cardiovasc Genet. 2010;3:179–86.
Ridker PM. Clinical application of C-reactive protein for cardiovascular disease detection and prevention. Circulation. 2003;107:363–9.
Ko YL, Hsu LA, Hsu KH, Ko YH, Lee YS. The interactive effects of hepatic lipase gene promoter polymorphisms with sex and obesity on high-density-lipoprotein cholesterol levels in Taiwanese-Chinese. Atherosclerosis. 2004;172:135–42.
Chan EC, Chang PY, Wu TL, Wu JT. Enzymatic assay of homocysteine on microtiter plates or a TECAN analyzer using crude lysate containing recombinant methionine gamma-lyase. Ann Clin Lab Sci. 2005;35:155–60.
Wu TL, Tsai IC, Chang PY, Tsao KC, Sun CF, Wu LL, et al. Establishment of an in-house ELISA and the reference range for serum amyloid A (SAA): complementarity between SAA and C-reactive protein as markers of inflammation. Clin Chim Acta. 2007;376:72–6.
Wu TL, Tsao KC, Chang CP, Li CN, Sun CF, Wu JT. Development of ELISA on microplate for serum C-reactive protein and establishment of age-dependent normal reference range. Clin Chim Acta. 2002;322:163–8.
Teng MS, Hsu LA, Wu S, Chou HH, Chang CJ, Sun YZ, et al. Mediation analysis reveals a sex-dependent association between ABO gene variants and TG/HDL-C ratio that is suppressed by sE-selectin level. Atherosclerosis. 2013;228:406–12.
Corella D, Guillen M, Saiz C, Portoles O, Sabater A, Cortina S, et al. Environmental factors modulate the effect of the APOE genetic polymorphism on plasma lipid concentrations: ecogenetic studies in a Mediterranean Spanish population. Metabolism. 2001;50:936–44.
Dong CQ, Luo YG, Dong K, Chen C, Liu Q, Yang TQ. Relationship between apolipoprotein E gene polymorphism with triglyceride level in patients with renal diseases. Ren Fail. 2013;35:1449–54.
Kolovou GD, Anagnostopoulou KK. Apolipoprotein E polymorphism, age and coronary heart disease. Ageing Res Rev. 2007;6:94–108.
Chasman DI, Kozlowski P, Zee RY, Kwiatkowski DJ, Ridker PM. Qualitative and quantitative effects of APOE genetic variation on plasma C-reactive protein, LDL-cholesterol, and apoE protein. Genes Immun. 2006;7:211–9.
Hubacek JA, Peasey A, Pikhart H, Stavek P, Kubinova R, Marmot M, et al. APOE polymorphism and its effect on plasma C-reactive protein levels in a large general population sample. Hum Immunol. 2010;71:304–8.
Marz W, Scharnagl H, Hoffmann MM, Boehm BO, Winkelmann BR. The apolipoprotein E polymorphism is associated with circulating C-reactive protein (the Ludwigshafen risk and cardiovascular health study). Eur Heart J. 2004;25:2109–19.
Vasunilashorn S, Glei DA, Lan CY, Brookmeyer R, Weinstein M, Goldman N. Apolipoprotein E is associated with blood lipids and inflammation in Taiwanese older adults. Atherosclerosis. 2011;219:349–54.
Kuhlmann I, Minihane AM, Huebbe P, Nebel A, Rimbach G. Apolipoprotein E genotype and hepatitis C, HIV and herpes simplex disease risk: a literature review. Lipids Health Dis. 2010;9:8.
Farrer LA, Cupples LA, Haines JL, Hyman B, Kukull WA, Mayeux R, et al. Effects of age, sex, and ethnicity on the association between apolipoprotein E genotype and Alzheimer disease. A meta-analysis. APOE and Alzheimer Disease Meta Analysis Consortium. JAMA. 1997;278:1349–56.
Gustavsson J, Mehlig K, Leander K, Strandhagen E, Bjorck L, Thelle DS, et al. Interaction of apolipoprotein E genotype with smoking and physical inactivity on coronary heart disease risk in men and women. Atherosclerosis. 2012;220:486–92.
Khan TA, Shah T, Prieto D, Zhang W, Price J, Fowkes GR, et al. Apolipoprotein E genotype, cardiovascular biomarkers and risk of stroke: systematic review and meta-analysis of 14,015 stroke cases and pooled analysis of primary biomarker data from up to 60,883 individuals. Int J Epidemiol. 2013;42:475–92.
Carvalho-Wells AL, Jackson KG, Gill R, Olano-Martin E, Lovegrove JA, Williams CM, et al. Interactions between age and apoE genotype on fasting and postprandial triglycerides levels. Atherosclerosis. 2010;212:481–7.
Maxwell TJ, Ballantyne CM, Cheverud JM, Guild CS, Ndumele CE, Boerwinkle E. APOE modulates the correlation between triglycerides, cholesterol, and CHD through pleiotropy, and gene-by-gene interactions. Genetics. 2013;195:1397–405.
Bennet AM, Di Angelantonio E, Ye Z, Wensley F, Dahlin A, Ahlbom A, et al. Association of apolipoprotein E genotypes with lipid levels and coronary risk. JAMA. 2007;298:1300–11.
Tikkanen E, Tuovinen T, Widén E, Lehtimäki T, Viikari J, Kähönen M, Peltonen L, Raitakari OT, Ripatti S. Association of known loci with lipid levels among children and prediction of dyslipidemia in adults. Circ Cardiovasc Genet. 2011;4(6):673–80.
Spracklen CN, Saftlas AF, Triche EW, Bjonnes A, Keating B, Saxena R, Breheny PJ, Dewan AT, Robinson JG, Hoh J, Ryckman KK. Genetic Predisposition to Dyslipidemia and Risk of Preeclampsia. Am J Hypertens. 2015;28(7):915–23.
Qi L, Rifai N, Hu FB. Interleukin-6 receptor gene, plasma C-reactive protein, and diabetes risk in women. Diabetes. 2009;58:275–8.
Toms TE, Smith JP, Panoulas VF, Blackmore H, Douglas KM, Kitas GD. Apolipoprotein E gene polymorphisms are strong predictors of inflammation and dyslipidemia in rheumatoid arthritis. J Rheumatol. 2012;39:218–25.
Guo L, LaDu MJ, Van Eldik LJ. A dual role for apolipoprotein e in neuroinflammation: anti- and pro-inflammatory activity. J Mol Neurosci. 2004;23:205–12.
Lahrach H, Essiarab F, Timinouni M, Hatim B, El Khayat S, Er-Rachdi L, et al. Association of apolipoprotein E gene polymorphism with end-stage renal disease and hyperlipidemia in patients on long-term hemodialysis. Ren Fail. 2014;36:1504–9.
Pavlicev M, Wagner GP. A model of developmental evolution: selection, pleiotropy and compensation. Trends Ecol Evol. 2012;27:316–22.
Pavlicev M, Cheverud JM, Wagner GP. Evolution of adaptive phenotypic variation patterns by direct selection for evolvability. Proc Biol Sci. 2011;278:1903–12.
Acknowledgements
This study was supported by a grant from the Taipei Tzu Chi Hospital, Buddhist Tzu Chi Medical Foundation (TCRD-TPE-103-RT-2, TCRD-I103-01-01, TCRD-TPE-NCS-102-01, TCRD-TPE-MOST-103-01, TCRD-TPE-MOST-104-09), a grant from the Tzu Chi University, Hualien, Taiwan (TCIRP102001-02Y1, TCMMP104-06-03), and a grant from the National Science Council (NSC 101-2314-B-303 -023 -MY3, MOST 104-2314-B-303-013-MY3) to Y.-L. Ko. We greatly appreciate technical support from the Core Laboratory of the Taipei Tzu Chi Hospital, Buddhist Tzu Chi Medical Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
SW and Y-LK wrote the paper and conceived and designed the experiments; L-AH analyzed the data; J-FL and H-HC collected and provided the sample for this study; M-ST and Y-MW performed the genotype data; M-CL and C-WS performed the ELISA. All authors read and approved the final manuscript.
Additional file
Additional file 1: Table S1.
Primer sequences and restriction enzymes (RE) used in all gene polymorphisms. Table S2. Analysis of inflammation markers with APOE genotypes. Table S3. Associations between different SNPs and triglyceride levels (mg/dL). (DOCX 21 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wu, S., Hsu, LA., Teng, MS. et al. Interactive effects of C-reactive protein levels on the association between APOE variants and triglyceride levels in a Taiwanese population. Lipids Health Dis 15, 94 (2016). https://doi.org/10.1186/s12944-016-0262-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12944-016-0262-z