
Abstract
Background
Pathogenic Escherichia coli are an important cause of bacterial infections in both humans and pigs and many of antimicrobials are used for the treatment of E. coli infection. The objective of this study was to investigate the characteristics and relationship between humans and pigs regarding third-generation cephalosporin resistance and CMY-2-producing E. coli in Korea.
Results
All 103 third-generation cephalosporin-resistant E. coli isolates showed multidrug resistance. Also, except for β-lactam/β-lactamase inhibitor combinations, all antimicrobials resistant rates were higher in pigs than in humans. A total of 36 isolates (humans: five isolates; pigs: 31 isolates) were positive for the CMY-2-encoding genes and thirty-two (88.9%) isolates detected class 1 integrons with 10 different gene cassette arrangements, and only 1 isolate detected a class 2 integron. The most common virulence genes in pigs were LT (71.0%), F18 (51.6%), and STb (51.6%), while stx2 (80.0%) was the most frequently detected gene in humans. Stx2 gene was also detected in pigs (6.5%). Interestingly, 36 CMY-2-producing E. coli isolates showed a high diversity of sequence types (ST), and ST88 was present in E. coli from both pigs (11 isolates) and humans (one isolate).
Conclusion
Our findings suggest that a critical need for comprehensive surveillance of third-generation cephalosporin resistance is necessary to preserve the usefulness of third-generation cephalosporins in both humans and pigs.
Similar content being viewed by others


Introduction
Escherichia coli is member of a large bacterial family, Enterobacteriaceae, which consists of facultative anaerobic Gram-negative rods that live in the intestinal microflora of humans and animals. Although many E. coli are harmless commensals, pathogenic E. coli are an important cause of bacterial infections like colibacillosis. In humans, these strains are the foremost cause of diarrhea and hemorrhagic colitis as well as hemolytic uremic syndrome [1]. In pigs, these strains cause diarrhea with edema disease and economic loss on pig farming due to decreasing weight gain, and costs for feed supplements, vaccinations and treatments [2]. Trimethoprim-sulfamethoxazole, fluoroquinolones, and third-generation cephalosporins are the important antimicrobials for treating infections caused by pathogenic E. coli. Third-generation cephalosporins are widely used for the treatment of bacterial infections in both human and veterinary medicines [3]. Misuse of third-generation cephalosporins increases the emergence of extended-spectrum β-lactamase (ESBL) and AmpC β-lactamases producing E. coli isolates in humans and animals [4,5,6]. These isolates are resistant to β-lactam antibiotics because the ESBL and AmpC β-lactamases allow bacteria to hydrolyze β-lactam antibiotics. AmpC beta-lactamase genes can be located and encoded on chromosomes or plasmids [7].
The genes that encode on plasmid called plasmid-mediated AmpC beta-lactamases (pAmpC) and are often overexpressed [5, 6, 8]. Eight families of pAmpC have been described based on differences in the amino acid sequence: ACC (Ambler class C), ACT (AmpC type), CMY (cephamycin), DHA (Dhahran hospital in Saudi Arabia), FOX (cefoxitin), LAT (latamoxef), MIR (Miriam hospital in Providence), and MOX (moxalactam) [9, 10]. Of these groups, CMY-2 is the most widely and prevalent existed in Enterobacteriaceae including E. coli [7].
In pathogenic E. coli, antimicrobial resistant genes and virulence genes are frequently detected in isolates from both humans and pigs. Pigs are considered the primary reservoirs of pathogenic E. coli which can lead the contamination of food products such as pork, and human infection [11,12,13]. Although some studies about third-generation cephalosporins resistance of pathogenic E. coli have been reported in either human or pigs [4, 5, 8], a relative paucity of information exists showing a relationship between humans and pigs. Therefore, this study aimed to investigate the characteristics and relationship between pathogenic isolates from humans and pigs with third-generation cephalosporin resistance of CMY-2-producing E. coli.
Materials and methods
Escherichia coli isolates
Between 2008 and 2020, 392 pathogenic E. coli isolates were collected from 401 pigs with colibacillosis clinical case in 120 different farms, respectively. The farms consisted of different pig herds (50 to 100 sows per each herd). Samples were not collected repeatedly from the same farm. The aseptically collected intestinal contents and feces were inoculated on MacConkey agar (BD Biosciences, Sparks, MD) and Eosin methylene blue agar (BD Biosciences) and incubated at 37℃ for 20 to 24 h. Only one typical colony was selected from each sample and transferred to blood agar. Suspected colonies were identified as E. coli using the VITEK II system (bioMéreiux, Craponne, France). Hemolysis was also determined in blood agar (Asan Pharmaceutical, Seoul, Korea). Also, 197 strains from 197 diarrheic patients from 1981–2019 were provided by the National Culture Collection for Pathogens (NCCP; Korea, 51 strains), Gyeongsang National University Hospital Branch of the NCCP (GNUH-NCCP; Korea, 138 strains), and Kyungpook National University Hospital Branch of the NCCP (KNUH-NCCP; Korea, eight strains).
Third-generation cephalosporin resistant E. coli identification
Two-hundred microliters of standardized inoculum (0.5 McFarland) of each E. coli isolate (collected on MacConkey agar without antimicrobial) was plated on Mueller–Hinton agar plates supplemented with 2 μg /mL cefotaxime (Sigma-Aldrich, St.Louis, MO) and incubated at 37 ℃ for 24 h to select third-generation cephalosporin-resistant E. coli [14]. Ultimately, a total of 103 third-generation cephalosporin-resistant E. coli strains (8 strains isolated from KNUH-NCCP, 15 strains isolated from GNUH-NCCP, 8 strains isolated from NCCP, and 72 strains isolated from 19 different pig farms in this study) were tested in this study (Additional file 1).
Antimicrobial susceptibility test
All third-generation cephalosporin-resistant E. coli isolates were investigated for their antimicrobial resistance using the disc diffusion test with the following 19 discs (BD Biosciences): amikacin (30 μg), amoxicillin/clavulanate (20/10 μg), ampicillin (10 μg), cefazolin (30 μg), cefepime (30 μg), cefoxitin (30 μg), cephalothin (30 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), colistin (10 μg), doxycycline (30 μg), gentamicin (10 μg), kanamycin (30 μg), nalidixic acid (30 μg), neomycin (30 μg), norfloxacin (10 μg), streptomycin (10 μg), tetracycline (30 μg), and trimethoprim/sulfamethoxazole (1.25/23.75 μg). Results were interpreted according to the Clinical and Laboratory Standards Institute guidelines [15, 16]. The minimum inhibitory concentrations (MICs) for cefazolin, cephalothin, cefoxitin, cefotaxime, cefpodoxime, ceftazidime, ceftriaxone, and cefepime were determined by standard broth microdilution methods with Mueller–Hinton broth (BD Biosciences) according to the recommendations of the CLSI [15, 16]. Escherichia coli ATCC 25,922 strain was used the control organisms in the antimicrobial susceptibility tests. Multi-drug resistance (MDR) was defined as acquired non-susceptibility to at least 1 agent in 3 or more antimicrobial categories [17].
Detection of β-lactamase-encoding genes
PCR amplification was conducted with primers (Table 1) for CTX-M, TEM, SHV, OXA, and pAmpC β-lactamase genes in the 103 third-generation cephalosporin-resistant E. coli. PCR products were sequenced using an automatic sequencer (Cosmogenetech, Seoul, Korea). The sequences were confirmed with those in the GenBank nucleotide database using the Basic Local Alignment Search Tool (BLAST) program available through the National Center for Biotechnology Information website (http://www.ncbi.nlm.nih.gov/BLAST). Therefore, 36 CMY-2-producing E. coli isolates were identified from 103 third-generation cephalosporin-resistant E. coli isolates.
Molecular analysis
For detection of antimicrobial resistance genes and virulence genes, PCR amplification was performed using DNA extracted from 36 CMY-2-producing E. coli isolates (Table 1). DNA was extracted using QIAamp DNA Mini kit (QIAGEN, Germany) according to the protocol for bacteria provided by the manufacturer. All CMY-2-producing E. coli isolates were tested for resistance genes related to aminoglycosides (aac (3)-II, aac(6’)-Ib, and ant(2’’)-I), chloramphenicols (cmlA and catA1), quinolone (qnrA, qnrB, qnrC, qnrD, qnrS, and qepA), sulfonamides (sul1 and sul2), and tetracyclines (tetA, tetB, tetC, tetD, tetE and tetG). Virulence factor genes associated with the toxins (LT, STa, STb, Stx2e, and EAST-1), fimbriae (F4, F5, F6, F18, and F41), and non-fimbrial adhesins (AIDA-1, paa, eae) were also confirmed by PCR as previously described [31].
Plasmid replicon typing and detection of integrons and gene cassettes
For plasmid replicon typing and detection of integrons and gene cassettes, PCR amplification was performed using DNA extracted from CMY-2-producing E. coli isolates. The DNA was extracted as described above. The primers used in this study targeted 18 different replicons [32] and class 1 and 2 integrons (Table 1). Gene cassettes were tested for integron-positive isolates. The PCR products of the gene cassettes were sequenced as described above (Additional file 2).
Multi-locus sequence typing (MLST)
All processes, including genomic DNA extraction, PCR amplification, Sanger sequencing, and assembly were performed by Macrogen (Macrogen, Seoul, South Korea). Genomic DNA were extracted using a QIAamp DNA Mini kit (QIAGEN, Germany). MLST was performed using partial sequences of seven house-keeping genes (adk, fumC, gyrB, icd, mdh, purA and recA). PCR was performed with 20 ng of genomic DNA as the template in a 30 μl reaction mixture, using Dr. MAX DNA Polymerase (Doctor Protein INC, South Korea) as follows: activation of Taq polymerase at 95 °C for 5 min; 35 cycles at 95 °C for 30 s, 52 °C for 30 s, and 72 °C for 1 min; and a final 10 min step at 72 °C. The products obtained after amplification were purified using a multiscreen filter plate (Millipore Corp, USA). Sequencing was performed using a PRISM BigDye Terminator v3.1 Cycle Sequencing Kit. The mixture was incubated at 95 °C for 5 min followed by 5 min on ice and then analyzed in an ABI PRISM 3730XL DNA analyzer (Applied Biosystems, USA). Sequence types (ST) were assigned online (http://pubmlst.org/biqsdb?db=pubmlst_ecoli_achtman_seqdef).
Statistical and data analysis
The statistical package SPSS 23 was used for the description of antimicrobial patterns. A comparison of antimicrobial resistance rate between third-generation cephalosporin-resistant E. coli isolated from human and third-generation cephalosporin-resistant E. coli isolated from pig was conducted with the two-sample t-test. Differences were considered significant at P < 0.05.
Results
Antimicrobial resistance
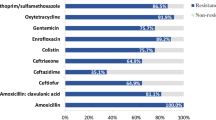
The MDR patterns of third-generation cephalosporin-resistant E. coli isolated from humans and pigs are shown in Table 2. All 103 third-generation cephalosporin-resistant E. coli isolates showed MDR against three to nine classes of antimicrobial agents. Isolates from pigs showed resistance to 9 classes (48.6%), while no isolates from humans showed resistance to 9 classes. Although, in pigs, resistance rates of penicillins (100%), aminoglycosides (98.6%), β-lactam/β-lactamase inhibitor combinations (87.5%), folate pathway inhibitors (80.6%), phenicols (91.7%), quinolone (86.1%), and tetracycline (88.9%) were higher than 80%, resistance rates of penicillins (100%), β-lactam/β-lactamase inhibitor combinations (100%), and aminoglycosides (98.6%), were higher than 80% in humans. Also, fluoroquinolones, folate pathway inhibitors, phenicols, quionolone, and tetracycline resistant rates were significantly higher in pigs than in humans (P < 0.05).
Characteristics of CMY-2-producing E. coli
The phenotypic and genotypic characteristics of the 36 CMY-2-producing E. coli isolates (humans: five isolates; pigs: 31 isolates) among the 103 third-generation cephalosporin-resistant E. coli isolates are shown in Table 3. All CMY-2-producing E. coli isolates had high MICs for most cephalosporins. Among the 36 CMY-2-producing E. coli isolates, TEM-1 and OXA-1 genes were detected in 27 (75.0%) and 4 (11.1%) isolates, respectively. Also, 5 human isolates and 22 isolates recovered from pigs harbored both CMY-2 and TEM-1 genes, respectively. Tetracycline-resistance genes were detected in all CMY-2-producing E. coli isolates from both humans and pigs. In pigs, tetA (100.0%) was the most prevalent resistance gene, but in humans, tetB (100.0%) was predominant. Two types of aminoglycoside-modifying enzyme genes (aac(6’)-Ib and aac(3)-II) were examined, but there was no ant(2’’)-I gene in CMY-2-producing E. coli isolate from either humans or pigs. Sul1 and sul2 sulfonamide-resistance genes were detected in 41.7% (humans: 100.0%; pigs: 32.3%;) and 75.0% (humans: 40.0%; pigs: 80.6%) of isolates, respectively. In pigs, both cmlA (58.1%) and catA1 (3.2%) chloramphenicol-resistance genes were identified; there were no chloramphenicol-resistance genes found in strains isolated from humans. The qnrS quinolone-resistance gene was the only quinolone-resistance gene detected in both pigs (32.3%) and humans (40.0%).
Among the 36 CMY-2-producing E. coli isolates, 88.9% (humans: 4 isolates; pigs: 28 isolates) were found to have class 1 integrons and only 1 isolate harbored class 2 integrons (Table 3). Class 1 integrons harbored ten types of gene cassette arrangements, aadA2-dfrA12 (five isolates), aadA5- dfrA17 (four isolates), aadA1 (two isolates), aadA1-dfrA1 (two isolates), aadA2-linF (two isolates), aadA1-aadA2-aadB-cmlA6 (one isolate), aadA2-aadA12-aadA23 (one isolate), aadA2-aadA28-dfrA12 (one isolate), aadA7-aac(3)-Id (one isolate), and arr-3-aadA16-dfrA27 (one isolate). Twelve isolates did not carry any of the gene cassettes. The class 2 integron-containing strain had only the aadA1-dfrA1-sat2 gene cassette arrangements. A total of 11 plasmid replicon types were detected in all 36 CMY-2-producing E. coli isolates (Table 3.). The most common plasmid replicon was FIB (83.3%), followed by I1 (75.0%) and Y (75.0%).
Virulence factors
Distributions of virotypes are shown in Table 3. The most prevalent virulence genes in pigs were LT (22 isolates, 71.0%), F18 (16 isolates, 51.6%), and STb (16 isolates, 51.6%), while stx2 (four isolates, 80.0%) followed by stx1 (three isolates, 60.0%) were most frequently detected in humans. The stx2 gene was also detected in pigs (two isolates, 6.5%). ETEC (17 isolates, 54.8%) was most prevalent pathotype in pigs, followed by ETEC/STEC (11 isolates, 35.5%). But, in humans, STEC (four isolates, 80.0%) was the most prevalent pathotype; it was also identified in pigs (one isolate, 3.2%).
Multi-locus sequence typing
Our collection of 36 CMY-2-producing E. coli isolates showed a high diversity of sequence types (ST) (Fig. 1). For isolates from human and pigs, we determined 4, and 8 different STs, respectively (Table 1). ST88 was present in E. coli from both pigs (11 isolates) and humans (one isolate). But several STs were only present in E. coli from pigs: ST100 (seven isolates), ST10 (six isolates), ST1 (two isolates), ST641 (two isolates), ST602 (one isolate), ST953 (one isolate), and ST1642 (one isolate). Also, ST410, ST131, and ST1308 were only observed in two, one, and one E. coli isolates, respectively, from humans. A population snapshot of 36 CMY-2-producing E. coli isolates, diagrammed based on a minimal spanning tree using optimized eBURST (goeBURST), based on PHYLOViZ software (www.phyloviz.net).

Minimum spanning tree based on sequence type of 36 CMY-2-producing E. coli isolated from human and pigs in Korea. Every circle represents a ST (the ST number is shown in the circle), and the size of the circle represents the number of isolates
Discussion
In our study, all third-generation cephalosporin-resistant E. coli were identified multidrug-resistant and were nonsusceptible to β-lactam antimicrobials like penicillins. These results indicate that third-generation cephalosporin-resistant E. coli show co-association of resistance to other classes of antimicrobials and high MDR rates. Also, although high resistance frequencies about non-beta-lactam antimicrobials were identified for aminoglycosides (98.6%), phenicols (91.7%), tetracycline (99.9%), quinolone (86.1%), and folate pathway inhibitors (80.6%) in pigs, only aminoglycoside resistance (90.3%) showed high frequency in humans. In animals, antimicrobial agents are used in large amounts to promote animal growth as well as to prevent and treat diseases [33, 34]. Therefore, the widespread use and misuse of antimicrobials in animal has resulted in the emergence of antimicrobial-resistant bacteria and it can get transferred to humans.
ESBL and pAmpC β-lactamase genes emerge when third-generation cephalosporins are overused and misused for prevention and treatment, which is a common mechanism of resistance to third-generation cephalosporins. In particular, the CMY-2-encoding gene is the most dominant pAmpC β-lactamase gene in E. coli isolates from both human and food producing animals. In Europe, the CMY-2-encoding gene has been detected in third-generation cephalosporin-resistant E. coli isolated in humans as well as in pigs [6, 35, 36]. In china, the prevalence of CMY-2-producing E. coli has been reported from food and domestic animals [37, 38]. Moreover, in Korea, CMY-2-encoding gene have been observed among animal and human E. coli isolates [5, 39, 40]. In this study, TEM-1 and OXA-1, enzymes conferring β-lactam resistance, were detected in 27 (75.0%) and 4 (11.1%) isolates, respectively. These genes are not ESBL or pAmpC β-lactamases, but can be transformed into ESBL by mutations that alter the amino acid sequence around the active site [41].
The CMY-2 gene can co-exist with other classes of antimicrobials genes in mobile genetic elements, and thus CMY-2-producing E. coli are commonly MDR [7, 42]. In this study, all 36 CMY-2-producing E. coli isolates carried a variety of antimicrobial resistance genes including sul1, sul2, catA1, cmlA, aac(6’)-Ib, aac(3)-II, tetA, tetB, tetE, and qnrS. The tetB gene was identified in CMY-2-producing E. coli isolates in both pigs and humans, which is consistent with the findings of Koga et al. and Endimiani et al. [7, 43]. The sul1 and sul2 genes, which encode sulfonamide-resistant dihydropteroate synthase, were identified in 15 (41.7%) and 27 (75.0%) CMY-2-producing E. coli isolates, respectively, and the aac(3)-II and aac(6’)-Ib genes, which encode aminoglycoside adenylyltransferase, were detected in 11 (30.6%) and 7 (19.4%) CMY-2-producing E. coli isolates, respectively. These genes have already been reported as major determinants of sulfonamides (sul1 and sul2) and gentamicin (aac(3)-II and aac(6’)-Ib) resistance in Gram-negative bacteria [44]. Although chloramphenicol is banned in food-producing animals because of its suspected carcinogenicity [45], we found that 18 (50.0%) and one (2.8%) of 31 CMY-2-producing E. coli isolated from pigs carried the cmlA and catA1 genes, respectively; these genes encodes a specific chloramphenicol transporter. Also, florfenicol which has been shown to have a spectrum of activity similar to that of chloramphenicol and used in veterinary medicine is related to chloramphenicol and can select for cross-resistance among bacterial pathogens [46, 47]. Therefore, although chloramphenicol is already prohibited worldwide in food animals, there is a reservoir of chloramphenicol resistance in bacteria from food animals, which can disseminate on transferable plasmids, remains a concern as chloramphenicol is a useful antibiotic for the treatment of bacterial infections in humans [48, 49]. qnrS genes, which encodes plasmid-mediated quinolone resistance, were identified in both pigs (32.3%) and humans (40.0%). Previous studies reported that the PMQR genes in β-lactamases-producing-E. coli were detected at high levels [50]. The presence of PMQR genes in β-lactamases-producing Enterobacteriaceae may be due to common carriage on a plasmid [51].
Integrons are genetic elements that transfer antimicrobial resistance gene, and they play an important role in conferring resistance to multiple antimicrobials [52]. In recent years, integrons have been found in β-lactamase-producing isolates of Gram-negative bacteria [53, 54]. In this study, the aadA gene was the most prevalent gene cassette of the integrons followed by the dfrA gene. These genes are also frequently detected in gene cassettes of integrons isolated from humans and pigs in Korea [55, 56]. Because β-lactamase-producing isolates harboring the aadA or dfrA or both genes showed higher antimicrobial resistance rates [57, 58], the selection of antimicrobials for the treatment of colibacillosis remains a serious concern. Also, the most common plasmid replicons were IncF plasmids including FIB (83.3%), FIC (52.8%), and FIA (33.3%). IncF plasmids are associated with important role in the worldwide emergence and spread of virulence and antimicrobial resistance determinants including extended-spectrum β-lactamases and pAmpC genes (CMY and DHA) among pathogenic E. coli [59].
For diagnosis and preventative measures for colibacillosis, detection of E. coli virulence factors is important [60]. In this study, LT, STb, F18, and F4 were detected in 22 (71.0%), 16 (51.6%), 16 (51.6%) and 11 (35.3%) CMY-2- producing E. coli isolates from pigs, respectively. The LT gene play a significant role in producing heat-labile enterotoxins and causing diseases [61].ST genes damage vessels and cause edema leading to high mortality in pigs [62]. Further, adhesive fimbriae gene including F4, F5, F6, F18, and F41 play important roles in allowing pathogenic E. coli to attach to the epithelial cells and intestinal mucosa and in causing diseases [58]. Interestingly, inactivated vaccines targeting F4 and F18 are being used in Korea [63]. The use of these vaccines could cause antigenic variations and would account for the prevalence of F4 and F18, in pigs. The stx2 gene was also detected in isolates from both pigs (two isolates, 6.5%) and humans (four isolates, 80.0%). The stx gene is associated with edema disease in swine and hemolytic-uremic syndrome in humans [64,65,66] and the receptor for stx2 is globotriosyl ceramide, which is found in both humans and pigs.
MLST help to identify the phylogenetic relationships among deep lineages, providing a view of the population structure of bacterial species [67]. In this study, we found eleven STs, including eight STs (ST1, ST10, ST88, ST100, ST602, ST641, ST953, and ST1642) from pigs and four STs (ST88, ST131, ST410, and ST1308) from humans. The most prevalent STs in pigs were ST88 (12 isolates), ST100 (seven isolates), and ST10 (six isolates), which are the predominant ETEC type, and are important pig pathogens in the many country (Canada, Germany, Thailand, and United States (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli). In particular, ST88 was reported in both humans and pigs and it has been previously described in association with antimicrobial resistant gene like AmpC [68]. These ST is related to strains pathogenic and antimicrobial resistance and emergence of similar ST might indicates transmission between pigs and humans [69, 70].
Conclusions
In this study, we genetically analyzed, characterized, and investigated the prevalence and relationship of third-generation cephalosporin resistance and CMY-2-producing E. coli isolated from humans and pigs in Korea suffering from diarrhea. To our knowledge, this is the first study to investigate the molecular characteristics and relationship between third-generation cephalosporin-resistant and CMY-2-producing E. coli isolated from humans and pigs in Korea. Third-generation cephalosporin resistant bacteria can get transferred to humans through the food chain and lead to treatment failure of serious infections. Therefore, a critical need for comprehensive surveillance of third-generation cephalosporin resistance is necessary to preserve the usefulness of third-generation cephalosporins in both humans and pigs.
Availability of data and materials
Not applicable.
References
Schroeder CM, Meng J, Zhao S, DebRoy C, Torcolini J, Zhao C, McDermott PF, Wagner DD, Walker RD, White DG. Antimicrobial resistance of Escherichia coli O26, O103, O111, O128, and O145 from animals and humans. Emerg Infect Dis. 2002;8(12):1409–14.
Fairbrother JM, Gyles CL, Colibacillosis. In: Zimmerman JJ, Karriker LA, Ramirez A, Schwartz KJ, Stevenson GW, editors. Disease of Swine. 10th ed; p. 723–747. 2012.
Moulin G, Cavalié P, Pellanne I, Chevance A, Laval A, Millemann Y, Colin P, Chauvin C. A comparison of antimicrobial usage in human and veterinary medicine in France from 1999 to 2005. J Antimicrob Chemother. 2008;62(3):617–25.
Nachimuthu R, Kannan VR, Bozdogan B, Krishnakumar V, Manohar P. CTX-M-type ESBL-mediated resistance to third-generation cephalosporins and conjugative transfer of resistance in Gram-negative bacteria isolated from hospitals in Tamil Nadu, India. Access Microbiol. 2020;3(3):000142.
Shin SW, Jung M, Won HG, Belaynehe KM, Yoon IJ, Yoo HS. Characteristics of Transmissible CTX-M- and CMY-Type β-Lactamase-Producing Escherichia coli Isolates Collected from Pig and Chicken Farms in South Korea. J Microbiol Biotechnol. 2017;27(9):1716–23.
Pietsch M, Irrgang A, Roschanski N, Brenner Michael G, Hamprecht A, Rieber H, Käsbohrer A, Schwarz S, Rösler U, Kreienbrock L, et al. Whole genome analyses of CMY-2-producing Escherichia coli isolates from humans, animals and food in Germany. BMC Genomics. 2018;19(1):601.
Koga VL, Maluta RP, da Silveira WD, Ribeiro RA, Hungria M, Vespero EC, Nakazato G, Kobayashi RKT. Characterization of CMY-2-type beta lactamase-producing Escherichia coli isolated from chicken carcasses and human infection in a city of South Brazil. BMC Microbiol. 2019;19(1):174.
Hernández-Fillor RE, Brilhante M, Marrero-Moreno CM, Baez M, Espinosa I, Perreten V. Characterization of Third-Generation Cephalosporin-Resistant Escherichia coli Isolated from Pigs in Cuba Using Next-Generation Sequencing. Microb Drug Resist. 2021;27(7):1003–10.
Jacoby GA. AmpC beta-Lactamases. Clin Microbiol Rev. 2009;22(1):161–82.
Philippon A, Arlet G, Jacoby GA. Plasmid-determined AmpC-type beta-lactamases. Antimicrob Agents Chemother. 2002;46(1):1–11.
Do KH, Byun JW, Lee WK. Antimicrobial resistance, adhesin and toxin genes of porcine pathogenic Escherichia coli following the ban on antibiotics as the growth promoters in feed. Pak Vet J. 2021;41(4):519–23.
Kim YJ, Kim JH, Hur J, Lee JH. Isolation of Escherichia coli from piglets in South Korea with diarrhea and characteristics of the virulence genes. Can J Vet Res. 2010;74(1):59–64.
Fairbrother JM, Nadeau E. Escherichia coli: on-farm contamination of animals. Rev Sci Tech. 2006;25(2):555–69.
Shim JB, Seo KW, Kim YB, Jeon HY, Lim SK, Lee YJ. Molecular characteristics of extended-spectrum and plasmid-mediated AmpC b-lactamase-producing Escherichia coli isolated from commercial layer in Korea. Poult Sci. 2019;98(2):949–56.
Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial disk and dilution susceptibility tests for bacteria isolated from Animals. 5th ed. Clinical and Laboratory Standards Institute; Wayne, PA, 2018. CLSI standard VET01.
Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing, M100-S29. Clinical and Laboratory Standards Institute, Wayne, PA. 2019.
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–81.
Dallenne C, Da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65(3):490–5.
Wang M, Tran JH, Jacoby GA, Zhang Y, Wang F, Hooper DC. Plasmid-mediated quinolone resistance in clinical isolates of Escherichia coli from Shanghai. China Antimicrob Agents Chemother. 2003;47(7):2242–8.
Jiang Y, Zhou Z, Qian Y, Wei Z, Yu Y, Hu S, Li L. Plasmid-mediated quinolone resistance determinants qnr and aac(6’)-Ib-cr in extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae in China. J Antimicrob Chemother. 2008;61(5):1003–6.
Wang M, Guo Q, Xu X, Wang X, Ye X, Wu S, Hooper DC, Wang M. New plasmid-mediated quinolone resistance gene, qnrC, found in a clinical isolate of proteus mirabilis. Antimicrob Agents Chemother. 2009;53(5):1892–7.
Cavaco LM, Hasman H, Xia S, Aarestrup FM. qnrD, a novel gene conferring transferable quinolone resistance in Salmonella enterica serovar Kentucky and Bovismorbificans strains of human origin. Antimicrob Agents Chemother. 2009;53(2):603–8.
Yamane K, Wachino J, Suzuki S, Kimura K, Shibata N, Kato H, Shibayama K, Konda T, Arakawa Y. New plasmid-mediated fluoroquinolone efflux pump, QepA, found in an Escherichia coli clinical isolate. Antimicrob Agents Chemother. 2007;51(9):3354–60.
Sandvang D, Aarestrup FM. Characterization of aminoglycoside resistance genes and class 1 integrons in porcine and bovine gentamicin-resistant Escherichia coli. Microb Drug Resist. 2000;6(1):19–27.
Van TT, Chin J, Chapman T, Tran LT, Coloe PJ. Safety of raw meat and shellfish in Vietnam: an analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int J Food Microbiol. 2008;124(3):217–23.
Sandvang D, Aarestrup FM, Jensen LB. Characterisation of integrons and antibiotic resistance genes in Danish multiresistant Salmonella enterica Typhimurium DT104. FEMS Microbiol Lett. 1998;160(1):37–41.
Maynard C, Fairbrother JM, Bekal S, Sanschagrin F, Levesque RC, Brousseau R, Masson L, Larivière S, Harel J. Antimicrobial resistance genes in enterotoxigenic Escherichia coli O149:K91 isolates obtained over a 23-year period from pigs. Antimicrob Agents Chemother. 2003;47(10):3214–21.
Sengeløv G, Agersø Y, Halling-Sørensen B, Baloda SB, Andersen JS, Jensen LB. Bacterial antibiotic resistance levels in Danish farmland as a result of treatment with pig manure slurry. Environ Int. 2003;28(7):587–95.
Ng LK, Mulvey MR, Martin I, Peters GA, Johnson W. Genetic characterization of antimicrobial resistance in Canadian isolates of Salmonella Serovar Typhimurium DT104. Antimicrob Agents Chemother. 1999;43(12):3018–21.
Sáenz Y, Briñas L, Domínguez E, Ruiz J, Zarazaga M, Vila J, Torres C. Mechanisms of resistance in multiple-antibioticresistant Escherichia coli strains of human, animal, and food Origins. Antimicrob Agents Chemother. 2004;48(10):3996–4001.
Do KH, Byun JW, Lee WK. Prevalence of O-serogroups, virulence genes, and F18 antigenic variants in Escherichia coli isolated from weaned piglets with diarrhea in Korea during 2008–2016. J Vet Sci. 2019;20(1):43–50.
Johnson TJ, Wannemuehler YM, Johnson SJ, Logue CM, White DG, Doetkott C, Nolan LK. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl Environ Microbiol. 2007;73(6):1976–83.
Food and Drug Administration (FDA). Antimicrobials Sold or Distributed for Use in Food-Prodcing Animals. Silver Spring, MD. 2020.
Wegener HC. Antibiotics in animal feed and their role in resistance development. Curr Opin Microbiol. 2003;6(5):439–45.
Ewers C, Bethe A, Semmler T, Guenther S, Wieler LH. Extended-spectrum b-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin Microbiol Infect. 2012;18(7):646–55.
Ewers C, de Jong A, Prenger-Berninghoff E, El Garch F, Leidner U, Tiwari SK, Semmler T. Genomic Diversity and Virulence Potential of ESBL- and AmpC-β-Lactamase-Producing Escherichia coli Strains From Healthy Food Animals Across Europe. Front Microbiol. 2021;12: 626774.
Guo YF, Zhang WH, Ren SQ, Yang L, Lü DH, Zeng ZL, Liu YH, Jiang HX. IncA/C Plasmid-Mediated Spread of CMY-2 in Multidrug-Resistant Escherichia coli from Food Animals in China. PLoS ONE. 2014;9(5): e96738.
Tang F, Wang J, Li D, Gao S, Ren J, Ma L, Liu F, Zhuge X, Yan G, Lu Y, et al. Comparative genomic analysis of 127 Escherichia coli strains isolated from domestic animals with diarrhea in China. BMC Genomics. 2019;20(1):212.
Hong JS, Song W, Park HM, Oh JY, Chae JC, Jeong S, Jeong SH. Molecular Characterization of Fecal Extended-Spectrum β-Lactamase- and AmpC β-Lactamase-Producing Escherichia coli From Healthy Companion Animals and Cohabiting Humans in South Korea. Front Microbiol. 2020;11:674.
Kim YK, Lee WK, Song KE. Complete genome of the multidrug-resistant Escherichia coli strain KBN10P04869 isolated from a patient with acute myeloid leukemia. Korean J Microbiol. 2018;54(4):442–4.
Bajpai T, Pandey M, Varma M, Bhatambare GS. Prevalence of TEM, SHV, and CTX-M Beta-Lactamase genes in the urinary isolates of a tertiary care hospital. Avicenna J Med. 2017;7(1):12–6.
Seo KW, Shim JB, Lee YJ. Emergence of CMY-2-Producing Escherichia coli in Korean Layer Parent Stock. Microb Drug Resist. 2019;25(3):462–8.
Endimiani A, Hilty M, Perreten V. CMY-2-Producing Escherichia coli in the Nose of Pigs. Antimicrob Agents Chemother. 2012;56(8):4556–7.
Dessie HK, Bae DH, Lee YJ. Characterization of integrons and their cassettes in Escherichia coli and Salmonella isolates from poultry in Korea. Poult Sci. 2013;92(11):3036–43.
Mbodi FE, Nguku P, Okolocha E, Kabir J. Determination of chloramphenicol residues in commercial chicken eggs in the federal capital territory, Abuja, Nigeria. Food Addit Contam Part A Chem Anal Control Expo Risk Assess. 2014;31(11):1834–9.
White DG, Hudson C, Maurer JJ, Ayers S, Zhao S, Lee MD, Bolton L, Foley T, Sherwood J. Characterization of Chloramphenicol and Florfenicol Resistance in Escherichia coli Associated with Bovine Diarrhea. J Clin Microbiol. 2000;38(12):4593–8.
Schwarz S, Kehrenberg C, Doublet B, Cloeckaert A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol Rev. 2004;28(5):519–42.
Eliakim-Raz N, Lador A, Leibovici-Weissman Y, Elbaz M, Paul M, Leibovici L. Efficacy and safety of chloramphenicol: joining the revival of old antibiotics? Systematic review and meta-analysis of randomized controlled trials. J Antimicrob Chemother. 2014;70(4):979–96.
Livingston RJ, Butterworth JW, Belt P. Reaction or infection: topical chloramphenicol treatment. Ann R Coll Surg Engl. 2013;95(1):e20–1.
Seo KW, Lee YJ. Detection of plasmid-mediated quinolone resistance genes in b-lactamase-producing Escherichia coli isolates from layer hens. Poult Sci. 2019;98(3):1480–7.
Xue G, Li J, Feng Y, Xu W, Li S, Yan C, Zhao H, Sun H. High prevalence of plasmid-mediated quinolone resistance determinants in Escherichia coli and Klebsiella pneumoniae isolates from pediatric patients in China. Microb Drug Resist. 2017;23(1):107–14.
Fluit AC, Schmitz FJ. Resistance integrons and superintegrons. Clin Microbiol Infect. 2004;10(4):272–88.
Ramatla T, Mileng K, Ndou R, Mphuti N, Syakalima M, Lekota KE, Thekisoe OMM. Molecular Detection of Integrons, Colistin and β-lactamase Resistant Genes in Salmonella enterica Serovars Enteritidis and Typhimurium Isolated from Chickens and Rats Inhabiting Poultry Farms. Microorganisms. 2022;10(2):313.
Seo KW, Lee YJ. The occurrence of CTX-M–producing E. coli in the broiler parent stock in Korea. Poult Sci. 2021;100(2):1008–15.
Kang HY, Jeong YS, Oh JY, Tae SH, Choi CH, Moon DC, Lee WK, Lee YC, Seol SY, Cho DT, et al. Characterization of antimicrobial resistance and class 1 integrons found in Escherichia coli isolates from humans and animals in Korea. J Antimicrob Chemother. 2005;55(5):639–44.
Yang YY, Suh MH. Analysis of Integrons and Antimicrobial Resistances of Multidrug Resistant Escherichia coli Isolated in Korea. J Bacteriol Virol. 2019;49(4):176–90.
Liu LT, Wan LH, Song XH, Xiong Y, Jin SJ, Zhou LM. Relevance of class 1 integrons and extended-spectrum β-lactamases in drug-resistant Escherichia coli. Mol Med Rep. 2013;8(4):1251–5.
Sung JY, Oh JE. Distribution and characterization of integrons in Enterobacteriaceae isolates from chickens in Korea. J Microbiol Biotechnol. 2014;24(7):1008–13.
Villa L, García-Fernández A, Fortini D, Carattoli A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J Antimicrob Chemother. 2010;65(12):2518–29.
Fairbrother JM, Nadeau E, Gyles CL. Escherichia coli in postweaning diarrhea in pigs: an update on bacterial types, pathogenesis, and prevention strategies. Anim Health Res Rev. 2005;6(1):17–39.
Mirhoseini A, Amani J, Nazarian S. Review on pathogenicity mechanism of enterotoxigenic Escherichia coli and vaccines against it. Microb Pathog. 2018;117:162–9.
Nguyen UV, Coddens A, Melkebeek V, Devriendt B, Goetstouwers T, Poucke MV, Peelman L, Cox E. High susceptibility prevalence for F4+ and F18+ Escherichia coli in Flemish pigs. Vet Microbiol. 2017;202:52–7.
Do KH, Park HE, Byun JW, Lee WK. Virulence and antimicrobial resistance profiles of Escherichia coli encoding mcr gene from diarrhoeic weaned piglets in Korea during 2007–2016. J Glob Antimicrob Resist. 2020;20:324–7.
Joseph A, Cointe A, Mariani Kurkdjian P, Rafat C, Hertig A. Shiga toxin-associated hemolytic uremic syndrome: A narrative review. Toxins (Basel). 2020;12(2):67.
Jung J, Kim H, Jo A, Kim J, Lee W, Byun J. Enrichment media for Stx2e production in Shiga toxin-producing Escherichia coli. J Biomed Transl Res. 2017;18:103–7.
Lee JB, Han D, Lee HT, Wi SM, Park JH, Jo JW, Cho YJ, Hahn TW, Lee S, Kang B, et al. Pathogenic and phylogenetic characteristics of non-O157 Shiga toxin-producing Escherichia coli isolates from retail meats in South Korea. J Vet Sci. 2018;19(2):251–9.
Jaureguy F, Landraud L, Passet V, Diancourt L, Frapy E, Guigon G, Carbonnelle E, Lortholary O, Clermont O, Denamur E, et al. Phylogenetic and genomic diversity of human bacteremic Escherichia coli strains. BMC Genomics. 2008;9:560.
Crémet L, Caroff N, Giraudeau C, Dauvergne S, Lepelletier D, Reynaud A, Corvec S. Occurrence of ST23 complex phylogroup A Escherichia coli isolates producing extended-spectrum AmpC β-lactamase in a French hospital. Antimicrob Agents Chemother. 2010;54(5):2216–8.
Nicolas-Chanoine MH, Blanco J, Leflon-Guibout V, Demarty R, Alonso MP, Caniça MM, Park YJ, Lavigne JP, Pitout J, Johnson JR. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J Antimicrob Chemother. 2008;61(2):273–81.
Do KH, Seo K, Lee WK. Antimicrobial resistance, virulence genes, and phylogenetic characteristics of pathogenic Escherichia coli isolated from patients and swine suffering from diarrhea. BMC Microbiol. 2022;22(1):199.
Acknowledgements
The pathogen resources from humans for this study were provided by the National Culture Collection for Pathogens, the Gyeongsang National University Hospital Branch of the NCCP, and the Kyungpook National University Hospital Branch of the NCCP.
Funding
This research was supported by "Regional Innovation Strategy (RIS)" through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (MOE) (2021RIS-001).
Author information
Authors and Affiliations
Contributions
KWS, KHD, and WKL conceived and designed the study. KWS, KHD, MKS, and WKL (Woo-Kon Lee) contributed in comprehensive research and sample collection. KWS wrote the paper. KHD, and WKL participated in manuscript editing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests. The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Prevalence of β-lactamase-encoding genes in 103 third-generation cephalosporin-resistant E. coli isolates.
Additional file 2.
Antibiotic resistance rates in 103 third-generation cephalosporin-resistant E. coli isolated from humans and pigs in Korea.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Seo, KW., Do, KH., Shin, MK. et al. Comparative genetic characterization of CMY-2-type beta-lactamase producing pathogenic Escherichia coli isolated from humans and pigs suffering from diarrhea in Korea. Ann Clin Microbiol Antimicrob 22, 7 (2023). https://doi.org/10.1186/s12941-023-00559-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-023-00559-1