
Abstract
Background
Artemisinin resistance in Plasmodium falciparum threatens global malaria elimination efforts. To contain and then eliminate artemisinin resistance in Eastern Myanmar a network of community-based malaria posts was instituted and targeted mass drug administration (MDA) with dihydroartemisinin-piperaquine (three rounds at monthly intervals) was conducted. The prevalence of artemisinin resistance during the elimination campaign (2013–2019) was characterized.
Methods
Throughout the six-year campaign Plasmodium falciparum positive blood samples from symptomatic patients and from cross-sectional surveys were genotyped for mutations in kelch-13—a molecular marker of artemisinin resistance.
Result
The program resulted in near elimination of falciparum malaria. Of 5162 P. falciparum positive blood samples genotyped, 3281 (63.6%) had K13 mutations. The prevalence of K13 mutations was 73.9% in 2013 and 64.4% in 2019. Overall, there was a small but significant decline in the proportion of K13 mutants (p < 0.001). In the MDA villages there was no significant change in the K13 proportions before and after MDA. The distribution of different K13 mutations changed substantially; F446I and P441L mutations increased in both MDA and non-MDA villages, while most other K13 mutations decreased. The proportion of C580Y mutations fell from 9.2% (43/467) before MDA to 2.3% (19/813) after MDA (p < 0.001). Similar changes occurred in the 487 villages where MDA was not conducted.
Conclusion
The malaria elimination program in Kayin state, eastern Myanmar, led to a substantial reduction in falciparum malaria. Despite the intense use of artemisinin-based combination therapies, both in treatment and MDA, this did not select for artemisinin resistance.
Similar content being viewed by others

Introduction
Artemisinin combination therapies (ACTs) are the first-line treatment for P. falciparum malaria globally. In the Greater Mekong Subregion (GMS) artemisinin resistance has emerged over the past two decades. This is characterized by slow parasite clearance [1] which facilitates the selection of partner drug resistance, resulting in ACT failures [2,3,4,5]. Artemisinin resistance in P. falciparum threatens recent gains in malaria control and compromises malaria elimination efforts in the countries of the GMS and beyond [6]. Recently artemisinin-resistant P. falciparum parasites have also been reported in several African countries [7,8,9,10]. Greater efforts are needed to eliminate P. falciparum malaria in these foci of multidrug resistance before it becomes untreatable [11].
Myanmar has the large majority of malaria cases in the GMS, and most of the artemisinin resistant P. falciparum parasites. Access to health services is very difficult in the remote areas of Myanmar where malaria is most prevalent [12, 13]. To address this, in recent years several thousand community-based health workers (CHWs) have been trained and equipped with rapid diagnostic test (RDT), antimalarials, and long-lasting insecticidal nets (LLINs). Improved community access to free early diagnosis and treatment has dramatically reduced the burden of malaria in those areas where these measures have been deployed [14, 15]. However, concerns have been raised that the intense usage of artemisinin, in particular the use of these drugs in mass drug administration MDA [16,17,18] could fuel the emergence and spread of drug resistance in the parasite population, although the basis for this concern has been contested [19]. Here we describe the impact of the elimination campaign on the presence of artemisinin resistant P. falciparum parasites.
Background
In 2013 and 2014, a large community-based malaria elimination programme was rolled out in five large rural areas (“townships”) of Kayin State in Eastern Myanmar where multi-drug resistant P. falciparum parasites were prevalent [3]. There were two geographically separate programmes -one led by the malaria elimination task force (METF) who were operational in Central Kayin State, and the other led by Medical Action Myanmar (MAM) who were operational in South Kayin State. A large network of CHWs was gradually introduced by both programmes to cover the remote villages in the target areas. Long-lasting insecticide treated bed-nets were distributed, and the prevalence of malaria was assessed by ultrasensitive polymerase chain reaction (uPCR) [20,21,22]. Selected communities with a high prevalence of asymptomatic malaria, identified by uPCR, (hereafter referred to as “hotspots”), were targeted with mass drug administration (MDA) in order to accelerate the elimination of P. falciparum parasites [21, 22]. As a result of these interventions, the incidence of P. falciparum malaria declined dramatically throughout the elimination campaign areas [21, 23].
Methods
Study design
We conducted a retrospective analysis of the changing prevalence of the kelch-13 molecular markers of artemisinin resistance in P. falciparum during the malaria elimination programmes in Kayin State, East Myanmar. This included the results from previously published work [20, 21, 24, 25].
Study sites
The P. falciparum samples included in this study were collected from the community-based malaria post network, as well as cross sectional surveys malaria surveys conducted by MAM and METF. The METF program provided malaria services in the townships of Hpapun, Hlaing Bwe, Myawaddy, Kawkareik, while the MAM program provided malaria services in Kyainseikgyi township. Early diagnosis (rapid diagnostic tests) and malaria treatments were provided in the community by trained CHWs. In total there were 1477 villages in the target areas with an estimated total population of 467,535 persons i.e. 0.9% of the population of Myanmar [21, 22].
Malaria diagnosis and treatment
Malaria posts were operated by trained CHWs who diagnosed symptomatic cases of malaria using RDT (PF/PV SD Bioline, Alere, Korea Inc.). Patients with P. falciparum malaria were treated with fixed dose combination tablets of 20 mg artemether and 120 mg lumefantrine (AL) (Coartem, Novartis) at an approximate dose of 1.5/9 mg/kg taken twice a day for 3 days after a meal or with oily food. They were also given a single dose of primaquine (PQ) (Remedica Ltd) of 0.25 mg base/kg in the METF program and 0.75 mg base/kg in the MAM program. In the METF program, P. vivax was treated with chloroquine (Remedica, Ltd) 25 mg base/kg for 3 days [22]. In the MAM program P. vivax was treated with 3 days chloroquine and 14 days primaquine 0.25 mg base/kg per day [21].
Malaria surveys and mass drug administrations
Villages were surveyed to identify high P. falciparum malaria prevalence (“hotspots”) using uPCR for the detection of malaria parasitaemia as described previously [21, 22, 26]. In total, 315 malaria surveys were conducted by METF and MAM between 2013 and 2017. In the METF program, a village was defined as a malaria “hotspot” if the uPCR P. falciparum prevalence was ≥ 20%, or the combined prevalence of P. falciparum and P. vivax was ≥ 40%. In the MAM program, a village was defined as a “hotspot” if the P. falciparum prevalence was > 10%, or the combined prevalence of P. falciparum and P. vivax was > 30%.
The METF program treated all identified hotspots with MDA, while the MAM program conducted a cluster randomized controlled trial in which only hotspots randomized to the intervention arm received MDA. The MDA consisted of three consecutive treatment rounds administered at one-month intervals. In each round, a weight-based regimen of dihydroartemisinin 7 mg per kg and piperaquine 55 mg per kg (DP) (Eurartesim, Sigma-Tau) was given once daily for 3 days and a single low-dose of PQ was provided on the first day [22]. All consenting individuals in the village were eligible to receive MDA except for pregnant women, children under 6 months of age, and breastfeeding mothers. These interventions and results have been reported elsewhere [20, 21].
MDA intervention was conducted in 68 hotspots and 38,617 DP treatments [3] and single low dose primaquine were administered between 2014 and 2018. Asymptomatic prevalence of P. falciparum was substantially reduced as well as the village-level incidence of symptomatic falciparum malaria were substantially reduced following MDA [20, 21].
Specimen collection
Samples of P. falciparum DNA were collected during cross-sectional malaria surveys [21, 22], and from village malaria posts and extracted from dried blood spots on filter paper (Whatman 3MM) taken from symptomatic individuals diagnosed with P. falciparum malaria by RDT. The collected blood spots were labeled with the village malaria post identification code (with corresponding GPS coordinates) and were then shipped to the main offices in Mae Sot, Thailand (METF), and Yangon, Myanmar (MAM) where they were stored in a dry, temperature-controlled room until molecular analysis at Mahidol University, Thailand, and the Texas Biomedical Research Institute, USA.
Laboratory methods
We obtained P. falciparum kelch (K13) gene sequence data using two approaches; Sanger sequencing and Illumina sequencing. The detailed laboratory procedures are described in the supplementary materials (Text 1).
Kelch13 mutations classification
For this analysis, P. falciparum genotyped results are grouped into four categories: wild type (WT), K13 mutations definitely associated with slow parasite clearance, K13 mutations definitely not associated with slow parasite clearance, and K13 mutations where the association has not been established [27].
Data analysis
Statistical analyses were performed using Stata (Statistical Software: Release 17. StataCorp LLC) and maps were produced using GIS software (ArcGIS version 10.5). Pre- and post-MDA allele frequencies were compared using chi-square tests.
Ethics committee reviews
The METF program was approved by the Department of Medical Research (Lower Myanmar) 73/Ethics 2014 and OxTREC (reference no. 1017-13). The other studies that provided samples were approved by the Department of Medical Research (Myanmar) Ethics/DMR/2015/113E and Ethics/DMR/2015/109E.
Results
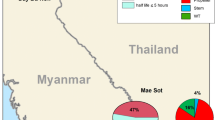
A total of 5162 samples of P. falciparum DNA were collected from 543 sites between 2013 and 2019, of which 12.5% (647) were collected during malaria surveys, and 87.5% [4] were collected at the malaria posts during routine case diagnosis and treatment. The majority of isolates came from Hpapun (66.2%), followed by Kyainseikgyi (27.6%), Myawaddy (4.7%), Hlaingbwe (1.3%), and Kawkareik (0.2%) (Fig. 1).

Location of collected P.falciparum genotyped specimen
Falciparum malaria incidence fell dramatically (a 15-fold decline) during the intervention period from 21.2 per 1000 persons-year in 2014 to 2.6 infections per 1000 persons-year in 2019 (Fig. 2). The overall proportion of K13 mutants in P. falciparum isolates was 63.6%. The highest proportion was observed in 2013 (73.9%) and the lowest was in 2018 (56.6%). Overall there was a small but significant decline in the proportion of K13 mutants among the P. falciparum isolates (Chi-square for trend; p < 0.0001) (Fig. 3). F446I was the most common mutation, present in 22.4% (1156/5162) of all genotyped isolates. The prevalence of F446I increased from 7.4% (21/283) [95% CI 4.9–11.1] in 2013 to a peak of 33.9% (353/1040) [95% CI 31.1–36.9] in 2016, and then decreased to 18.1% (27/149) [95% CI 12.7–25.1] in 2019. P441L was the second most common mutation accounting for 9.6% (496/5,162) of all genotypes. Its prevalence increased substantially from 1.4% (4/283) [95% CI 0.5–3.7] in 2013 to 27.5% (41/149) [95% CI 20.9–35.2] in 2019. In contrast, the prevalence of the previously dominant C580Y mutation [25] was 30.4% (86/283) [95% CI, 25.3–36.0] in 2013 but then fell steadily to 0.7% (1/149) [95% CI 0.1–4.6] by 2019 (Table 1, Fig. 4).

Annual symptomatic P.falciparum/Pmix incidence per thousand persons per year from 2014 to 2019 diagnosed and treated by malaria posts

Annual proportion of P. falciparum isolates with K13 mutations during the six-year programmes. Circled areas are proportional to the square root of the sample size

Proportion of K13 mutations from 2013 to 2019. The y-axis represents K13 mutants proportion with 95% confidence intervals while the x-axis denotes the years from 2013 to 2019. #K13 mutations where the association with parasite clearance has not yet been established. The uncharacterized mutations are A621V, A626S, C469F, C469Y, C542Y, D109Y, D452E, D584V, E208K, E321K, F614L, G533A, G533D, G533S, G718S, I205T, K189T, K438N, K479I, K586E, M562I, N264I, N490H, N525Y, R265P, R528S, R529G, R575K, S423N, T192I, T535M, V193E, V494I, V534G, W611C
Kelch13 mutations distribution by area
Hpapun township
K13 mutations were present in 57.6% (1969/3417) of the isolates collected in Hpapun from 2013 to 2019. The prevalence of K13 mutations was lowest (47.9%; 45/94) in 2013 and the highest (70.4%; 19/27) in 2015 (Fig. 5A). A total of 31 different K13 mutations were identified, of which 4 mutations (F446I, P441L, R561H, G449A) accounted for 46.3% (1,581/3,417) of all genotypes (Additional file 3: Table S1A). The F446I mutation was most common (15.3%) and declined in proportion slightly from 22.3% (21/94) [95% CI 15.0–31.9] in 2013 to 18.4% (26/141) [95% CI 12.9–25.7] in 2019. The P441L mutation was not detected in 2013 and was infrequent in 2014 (2.4%, 6/254) [95%CI 1.1–5.2] but then increased to become the most common K13 mutation (29.1%; 91/896) [95% CI 22.2–37.1] by 2019. C580Y was detected at low frequencies which declined from 3.2% (3/94) [95% CI 1.0–9.4] in 2013 to 0.7% (1/141) [95% CI 0.1–4.9] in 2019. (Additional file 1: Fig. S1A).

Proportion of K13 mutations from 2013 to 2019. A Hpapun, B Kyainseikgyi and C Myawaddy, Kawkareik and Hlaingbwe. #K13 mutations where the association with parasite clearance has not yet been established. The uncharacterized mutations are A621V, A626S, C469F, C469Y, C542Y, D109Y, D452E, D584V, E208K, E321K, F614L, G533A, G533D, G533S, G718S, I205T, K189T, K438N, K479I, K586E, M562I, N264I, N490H, N525Y, R265P, R528S, R529G, R575K, S423N, T192I, T535M, V193E, V494I, V534G, W611C
Kyainseikgyi township
In Kyainseikgyi K13 mutants accounted for the majority of the genotyped isolates (74.2%; 1057/1,425) collected from 2015 to 2017, during the implementation of an MDA cluster randomized trial. The prevalence of K13 mutations decreased slightly from 70.8% (379/535) [95% CI 66.8–74.5] in 2015 to 66.6% (285/428) [95% CI 42.0–70.9] in 2017 (Additional file 1: Fig. S1B). Overall F466I was the dominant mutation, accounting for 43.9% (625/1425) of the K13 mutant genotypes. The C580Y mutation was the 2nd most common detected mutation (9.6%; 137/1425) (Additional file 3: Table S1B).
Myawaddy, Hlaingbwe and Kawkareik
In these 3 townships, K13 mutations were present in 79.7% (255/320) of isolates. The proportion of C580Y mutants was highest in 2013, accounting for 43.9% (83/189) [95% CI 37.0–51.1] of isolates but it then declined to 24.0% (12/50) [95% CI 14.1–37.7] by 2018 (Fig. 5C). By 2019 malaria had declined substantially. Only 7 K13 mutations were identified of which 6 were uncharacterized mutations (Additional file 3: Table S1C).
Impact of mass drug administration on the proportion of K13 mutations
A total of 1280 genotypes were collected from 69 MDA villages. Of these, 36.5% (467/1280) were collected before MDA and 63.5% (813/1280) after MDA was administered (Table 2, Additional file 2: Fig. S2). Collection of samples before MDA was done over a shorter period (median 7 months; IQR 2–12 months) than after MDA (median 14 months; IQR 7–21 months).
Comparing samples gathered before and after MDA the proportion of F446I mutations increased from 10.7% (50/467) to 23.0% (187/813), (p < 0.001), the proportion of P441L mutations increased from 6.0% (28/467) to 10.7% (87/813) (p = 0.039) and the proportion of C580Y mutants decreased from 9.2% (43/467) to 2.3% (19/813) (p < 0.001), (Table 2). However, overall the proportion of isolates with K13 mutations was similar before MDA (60.3%; 282/467) and after MDA (56.3%; 458/813) (p = 0.158). In 487 villages where MDA was not delivered (which by design had less malaria than the MDA villages) the overall proportion of P. falciparum isolates with K13 mutants did not change significantly between 2013 (70.1%, 150/214) and 2019 (73%, 88/121) (p = 0.610). As in the MDA villages, the proportion of F446I mutants increased from 9.8%, (21/214) in 2013 to 22.3%, (27/121) in 2019 (p < 0.001), the proportion of P441L mutations increased from 1.4% (3/67) in 2013 to 30.6% (37/121) in 2019 (p < 0.001) and the proportion of C580Y mutations decreased from 28.5% (61/214) in 2013 to 0.8% (1/121) in 2019 (p < 0.001). (Fig. 6). There was no difference in the prevalence of K13 mutations in villages where MDA was deployed to those in contemporaneous areas where MDA was not deployed.

Proportion of K13 mutations comparison in MDA and non-MDA villages from 2013 to 2019. #Uncharacterized mutations are K13 mutations where the association with parasite clearance has not yet been established
Discussion
Artemisinin resistance in Plasmodium falciparum was first identified in western Cambodia nearly 20 years ago, and it is now widespread in the Greater Mekong subregion [1, 25]. Mutations mainly in the propeller region of the K13 gene cause artemisinin resistance [28], and this manifests as slow parasite clearance following treatment [2]. Other genetic factors also contribute to the phenotype of reduced ring stage antiparasitic activity and thus slow parasite clearance following treatment with artemisinin drugs [29]. Resistance to the artemisinin component of ACTs leaves the partner drugs with a higher residual load and thus a greater probability of selecting partner drug resistance [30, 31]. This has been documented both for mefloquine and piperaquine containing ACTs [3, 5, 32]. Among several independent emergences with different K13 mutations a single dominant lineage with slow parasite clearance (K13 C580Y) was selected and spread widely over Cambodia and surrounding areas in Vietnam, Laos and East Thailand [33]. This dominant parasite lineage was linked with high dihydroartemisinin-piperaquine treatment failure rates [4].
Meanwhile in Myanmar, which has the vast majority of malaria cases in the Greater Mekong subregion (GMS), different lineages emerged and then spread over wide distances [33]. This compromised the artesunate-mefloquine combination [3] but so far has not compromised the national first-line recommended antimalarial treatment artemether-lumefantrine (although there are few recent data) [34]. When artemisinin resistance was recognized first it seemed that the best containment strategy was to try and eliminate all falciparum malaria from the affected regions. To complete this rapidly mass drug administrations would be needed in focal areas with malaria hotspots, and this would require use of dihydroartemisinin-piperaquine and single low-dose primaquine. This was regarded as a risk for the further selection and spread of artemisinin resistance. The falciparum malaria elimination campaign in Eastern Myanmar described in this study resulted in a rapid decline in P. falciparum malaria incidence (Fig. 2) [20, 21, 23]. Critically, while there were changes in the proportions of different K13 genotypes, there was no overall variation in the prevalence of K13 mutations despite the widespread deployment of artemisinin-based combination therapy both in treatment and MDA campaigns.
The parasite lineage with the K13 C580Y allele dominated initially on the Thailand-Myanmar border in 2013 [3, 34, 35], but this declined over the six years study period and has now almost disappeared in this region. Interestingly a C580Y mutant lineage of different origin also dominated in the Eastern GMS and also has declined in prevalence relative to other K13 genotypes in recent years [25]. Meanwhile, in Eastern Myanmar the proportions of parasites with F446I and P441L mutations increased. In addition, the G449A and R561H mutations also became established in the area, albeit to a lesser extent. Other K13 mutations were identified at low frequencies. The F446I mutation confers less slowing of parasite clearance [2], but the reasons why some mutations increase in relative prevalence is not well characterized—and may relate to other genetic factors increasing relative fitness which are unrelated to artemisinin resistance. Among uncharacterized mutations in Kelch13 gene, 68% of these mutants were reported in Africa, Asia, Americas and Oceania according to the WWARN artemisinin Molecular Surveyor (http://www.wwarn.org/molecular/surveyor/k13).
In the villages where MDA was conducted, we found no evidence that MDA had selected for artemisinin resistance in the few remaining parasites. It is commonly stated that MDA selects for drug-resistant parasites, but this depends on how effective the MDA is. Poorly implemented MDA with low coverage or inadequate regimens might fail to eliminate the parasites. This failure might create a selective pressure where resistant strains survive and multiply, potentially leading to an increased of artemisinin resistance in the long run.
In this elimination campaign MDA interventions resulted in a significant reduction in P. falciparum incidence through high MDA coverage with an effective treatment regimen among targeted hotspots [20, 21]. MDA generally encounters relatively low parasite densities in infections that have already been controlled by host immunity, and this increases the probability of successful parasite clearance. During MDA, all parasite carriers are treated, and the number of P. falciparum infections with high parasite biomass is reduced, resulting in fewer courses of ACT treatment needed to treat symptomatic higher burden infections. This reduces the risk of resistance selection [36]. The deployment of different ACT regimens for routine treatment and for MDA is an additional factor which reduces the chance of selecting resistant parasites [37]. The high level of participation of the villagers in the elimination programme, facilitated by effective community engagement [38], and the absence of other sources of antimalarials, likely played an additional role in preventing the increase of artemisinin resistance in this area.
This study has several limitations. It is a largely observational experience. There were no K13 data on the period before the establishment of the malaria post/ community health worker network in Eastern Myanmar. Other molecular markers of antimalarial drug resistance were not assessed, although molecular markers of lumefantrine resistance (other than Pfmdr1 amplification) are not well established [25, 39]. MDA interventions quickly followed establishment of the malaria posts in the “hotspot” villages, and so there was shorter follow-up time and there were fewer P. falciparum specimens collected in pre-MDA period than in the post-MDA period. This may have resulted in an underestimate of the presence of some mutant alleles before MDA. Most of the specimens were collected from febrile patients who presented to the malaria posts with a positive malaria HRP2 antigen based rapid diagnostic test. Theoretically, this means that infections caused by parasites that carry histidine rich proteins deletion mutation (HRP2) could have been missed. However, the rate of PfHRP2 deletion has been reported to be very low in Myanmar [40, 41].
Continuing surveillance of genetic markers for both artemisinin and partners drugs resistance is warranted in this area.
Conclusion
This elimination campaign, was successful in reducing the incidence of P. falciparum. Despite the intense use of artemisinin-based combination therapies, both in treatment and MDA, it did not select for further artemisinin resistance.
Availability of data and materials
All relevant data are within the manuscript. Upon reasonable request via the MORU website or from MORU data sharing committee datasharing@tropmedres.ac the raw data set will be available.
References
Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, et al. Artemisinin resistance in Plasmodium falciparum malaria. New Engl J Med. 2009;361(5):455–67.
Ashley EA, Dhorda M, Fairhurst RM, Amaratunga C, Lim P, Suon S, et al. Spread of artemisinin resistance in plasmodium falciparum Malaria. New Engl J Med. 2014;371(5):411–23.
Phyo AP, Ashley EA, Anderson TJC, Bozdech Z, Carrara VI, Sriprawat K, et al. Declining efficacy of artemisinin combination therapy against P. falciparum malaria on the Thai-Myanmar border (2003–2013): the role of parasite genetic factors. Clin Infect Dis. 2016;63(6):784–91.
Amato R, Lim P, Miotto O, Amaratunga C, Dek D, Pearson RD, et al. Genetic markers associated with dihydroartemisinin–piperaquine failure in Plasmodium falciparum malaria in Cambodia: a genotype–phenotype association study. Lancet Infect Dis. 2017;17(2):164–73.
Amaratunga C, Lim P, Suon S, Sreng S, Mao S, Sopha C, et al. Dihydroartemisinin–piperaquine resistance in Plasmodium falciparum malaria in Cambodia: a multisite prospective cohort study. Lancet Infect Dis. 2016;16(3):357–65.
White NJ. Triple artemisinin-containing combination anti-malarial treatments should be implemented now to delay the emergence of resistance. Malar J. 2019;18(1):338.
Balikagala B, Fukuda N, Ikeda M, Katuro OT, Tachibana SI, Yamauchi M, et al. Evidence of artemisinin-resistant malaria in Africa. New Engl J Med. 2021;385(13):1163–71.
Ikeda M, Kaneko M, Tachibana SI, Balikagala B, Sakurai-Yatsushiro M, Yatsushiro S, et al. Artemisinin-resistant plasmodium falciparum with high survival rates, Uganda, 2014–2016. Emerg Infect Dis. 2018;24(4):718–26.
Sene SD, Pouye MN, Martins RM, Diallo F, Mangou K, Bei AK, et al. Identification of an in vitro artemisinin-resistant Plasmodium falciparum kelch13 R515K mutant parasite in Senegal. Front Parasitol. 2023;2:1076759.
Conrad MD, Asua V, Garg S, Giesbrecht D, Niaré K, Smith S, et al. Evolution of Partial resistance to artemisinins in malaria parasites in Uganda. N Engl J Med. 2023;389(8):722–32.
Ndwiga L, Kimenyi KM, Wamae K, Osoti V, Akinyi M, Omedo I, et al. A review of the frequencies of Plasmodium falciparum Kelch 13 artemisinin resistance mutations in Africa. Int J Parasitol Drugs Drug Resist. 2021;16:155–61.
WHO. World malaria report 2015. Geneva: WHO; 2015. p. 243.
Parker DM, Carrara VI, Pukrittayakamee S, McGready R, Nosten FH. Malaria ecology along the Thailand-Myanmar border. Malaria J. 2015;14(1):388.
NMCP. (NMCP M). Annual VBDC presentation. 2021.
WHO. World malaria report 2021. Geneva: WHO; 2021. p. 263.
von Seidlein L, Greenwood BM. Mass administrations of antimalarial drugs. Trends Parasitol. 2003;19(10):452–60.
WHO. Consideration of mass drug administration for the containment of artemisinin-resistant malaria in the Greater Mekong subregion, Report of a consensus meeting, 27–28 September 2010. Geneva: WHO; 2011.
Poirot E, Skarbinski J, Sinclair D, Kachur SP, Slutsker L, Hwang J. Mass drug administration for malaria. Cochrane Db Syst Rev. 2013;2013(12):CD008846.
White NJ. Does antimalarial mass drug administration increase or decrease the risk of resistance? Lancet Infect Dis. 2017;17(1):e15-20.
Landier J, Parker DM, Thu AM, Lwin KM, Delmas G, Nosten FH, et al. Effect of generalised access to early diagnosis and treatment and targeted mass drug administration on Plasmodium falciparum malaria in Eastern Myanmar: an observational study of a regional elimination programme. Lancet. 2018;391(10133):1916–26.
McLean ARD, Indrasuta C, Khant ZS, Phyo AK, Maung SM, Heaton J, et al. Mass drug administration for the acceleration of malaria elimination in a region of Myanmar with artemisinin-resistant falciparum malaria: a cluster-randomised trial. Lancet Infect Dis. 2021;21(11):1579–89.
Parker DM, Landier J, Thu AM, Lwin KM, Delmas G, Nosten FH, et al. Scale up of a Plasmodium falciparum elimination program and surveillance system in Kayin State. Myanmar Wellcome Open Res. 2017;2:98.
Rae JD, Landier J, Simpson JA, Proux S, Devine A, Maude RJ, et al. Longitudinal trends in malaria testing rates in the face of elimination in eastern Myanmar: a 7-year observational study. BMC Public Health. 2021;21(1):1725.
Bonnington CA, Phyo AP, Ashley EA, Imwong M, Sriprawat K, Parker DM, et al. Plasmodium falciparum Kelch 13 mutations and treatment response in patients in Hpa-Pun District, Northern Kayin State, Myanmar. Malaria J. 2017;16(1):480.
Imwong M, Dhorda M, Tun KM, Thu AM, Phyo AP, Proux S, et al. Molecular epidemiology of resistance to antimalarial drugs in the Greater Mekong subregion: an observational study. Lancet Infect Dis. 2020;20(12):1470–80.
Imwong M, Nguyen TN, Tripura R, Peto TJ, Lee SJ, Lwin KM, et al. The epidemiology of subclinical malaria infections in South-East Asia: findings from cross-sectional surveys in Thailand-Myanmar border areas, Cambodia, and Vietnam. Malar J. 2015;14(1):381.
Amaratunga C, Andrianaranjaka VH, Ashley E, Bethell D, Björkman A, Bonnington CA, et al. Association of mutations in the Plasmodium falciparum Kelch13 gene (Pf3D7_1343700) with parasite clearance rates after artemisinin-based treatments—a WWARN individual patient data meta-analysis. BMC Med. 2019;17(1):1.
Wicht KJ, Mok S, Fidock DA. Molecular mechanisms of drug resistance in plasmodium falciparum malaria. Annu Rev Microbiol. 2020;74(1):431–54.
Zhu L, Tripathi J, Rocamora FM, Miotto O, van der Pluijm R, Voss TS, et al. The origins of malaria artemisinin resistance defined by a genetic and transcriptomic background. Nat Commun. 2018;9(1):5158.
Dhorda M, Amaratunga C, Dondorp AM. Artemisinin and multidrug-resistant Plasmodium falciparum—a threat for malaria control and elimination. Curr Opin Infect Dis. 2021;34(5):432–9.
Hanboonkunupakarn B, White NJ. Advances and roadblocks in the treatment of malaria. Br J Clin Pharmacol. 2022;88(2):374–82.
Witkowski B, Duru V, Khim N, Ross LS, Saintpierre B, Beghain J, et al. A surrogate marker of piperaquine-resistant Plasmodium falciparum malaria: a phenotype–genotype association study. Lancet Infect Dis. 2017;17(2):174–83.
Imwong M, Suwannasin K, Kunasol C, Sutawong K, Mayxay M, Rekol H, et al. The spread of artemisinin-resistant Plasmodium falciparum in the Greater Mekong subregion: a molecular epidemiology observational study. Lancet Infect Dis. 2017;17(5):491–7.
Tun KM, Imwong M, Lwin KM, Win AA, Hlaing TM, Hlaing T, et al. Spread of artemisinin-resistant Plasmodium falciparum in Myanmar: a cross-sectional survey of the K13 molecular marker. Lancet Infect Dis. 2015;15(4):415–21.
Anderson TJC, Nair S, McDew-White M, Cheeseman IH, Nkhoma S, Bilgic F, et al. Population parameters underlying an ongoing soft sweep in southeast Asian malaria parasites. Mol Biol Evol. 2017;34(1):131–44.
White NJ. The consequences of treating asymptomatic malaria Parasitemia. Clin Infect Dis. 2017;64(5):654–5.
Boni MF, Smith DL, Laxminarayan R. Benefits of using multiple first-line therapies against malaria. Proc National Acad Sci. 2008;105(37):14216–21.
König A, Kajeechiwa L, Thwin MM, Nosten S, Tun SW, Tangseefa D, et al. Community engagement for malaria elimination in contested areas of the Karen/Kayin State, Myanmar: a case study on the Malaria Elimination Task Force. Wellcome Open Res. 2018;3:22.
Price RN, Uhlemann AC, van Vugt M, Brockman A, Hutagalung R, Nair S, et al. Molecular and pharmacological determinants of the therapeutic response to Artemether-Lumefantrine in multidrug-resistant plasmodium falciparum Malaria. Clin Infect Dis. 2006;42(11):1570–7.
Li P, Xing H, Zhao Z, Yang Z, Cao Y, Li W, et al. Genetic diversity of Plasmodium falciparum histidine-rich protein 2 in the China-Myanmar border area. Acta Trop. 2015;152:26–31.
Lê HG, Kang JM, Lee J, Yoo WG, Myint MK, Lin K, et al. Genetic variations in histidine-rich protein 2 and histidine-rich protein 3 of Myanmar Plasmodium falciparum isolates. Malaria J. 2020;19(1):388.
Acknowledgements
The authors would like to acknowledge the hard work and support of all members of the METF and MAM, collaborators and colleagues who have contributed to this work. NJW is a Principal Research Fellow funded by the Wellcome Trust (092956/Z/10/C). For the purpose of Open Access, the author has applied a CC BY public copyright license to any Author Accepted Manuscript version arising from this submission.
Funding
The work was supported by the Regional Artemisinin Initiative RAI and RAI2E (Global Fund against AIDS, Tuberculosis and Malaria), the Wellcome Trust (220211) and the Bill & Melinda Gates Foundation (OPP1117507). APP is funded through the Wellcome Trust (214208/Z/18/Z). TJA is supported by NIH (R01 AI048071). The funders had no role in study design, data collection and analysis, the decision to publish or the preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
APP, AMT, FN, NJW and FS contributed to the conceptualization of the study. KS and WW contributed to the specimen collection. AA, AR, XL, TJA, OM, MI performed the specimen analysis. AMT, APP, CP, NJW and FS contributed to the analysis. AMT, APP and FS wrote the original draft of the manuscript. AMT, FN, NJW and FS contributed to the writing and editing of the manuscript. EAA, ARDM, APP, JDR, JL, DMP, TA, CP review the manuscript. All authors read, edited and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
No competing interests were disclosed.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Proportion of K13 mutations by townships from 2013 and 2019. (A) Hpapun (B) Kyainseikgyi (C) Myawaddy, Hlaingbwe and Kawkareik. The y-axis indicates K13 mutant proportion with 95% confidence intervals.
Additional file 2: Figure S2.
Comparison of the proportion K13 mutations before and after MDA. # Uncharacterized mutations are K13 mutations where the association with parasite clearance has not yet been established.
Additional file 3:
Table S1A. K13 mutations frequency in Hpapun township from 2013 to 2019. Table S1B. K13 mutations frequency in Kyainseikgyi township from 2015 to 2017. Table S1C. K13 mutations frequency in Myawaddy, Kawkareik and Hlaingbwe townships from 2013 to 2019. Table S2. Status of uncharacterized K13 mutants in other contexts.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Thu, A.M., Phyo, A.P., Pateekhum, C. et al. Molecular markers of artemisinin resistance during falciparum malaria elimination in Eastern Myanmar. Malar J 23, 138 (2024). https://doi.org/10.1186/s12936-024-04955-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-024-04955-6