
Abstract
Background
Since the advent of the Green Revolution, pesticides have played an important role in the global management of invertebrate pests including vector mosquitoes. Despite optimal efficacy, insects often display insensitivity to synthetic insecticides owing to prolonged exposure that may select for resistance development. Such insecticide insensitivity may regress national and regional coordination in mosquito vector management and indeed malaria control. In Botswana, prolonged use of synthetic insecticides against malaria vectors have been practiced without monitoring of targeted mosquito species susceptibility status.
Methods
Here, susceptibility status of a malaria vector (Anopheles arabiensis), was assessed against World Health Organization-recommended insecticides, across three malaria endemic districts. Adult virgin female mosquitoes (2–5 days old) emerging from wild-collected larvae were exposed to standardized insecticide-impregnated papers with discriminating doses.
Results
The results showed resistance dynamics were variable in space, presumably as a result of spatial differences in insecticide use across malaria endemic districts and the types of insecticides used in the country. Overall, there was a reduced susceptibility of An. arabiensis for the pyrethroid lambda-cyhalothrin and for dichloro diphenyl trichloroethane [DDT], which have similar modes of action and have been used in the country for many years. The Okavango district exhibited the greatest reduction in susceptibility, followed by Ngamiland and then Bobirwa, reflective of national intervention strategy spraying intensities. Vector mosquitoes were, however, highly susceptible to carbamates and organophosphates irrespective of region.
Conclusions
These results provide important findings of vector susceptibility to insecticides recommended for vector control. The results highlight the need to implement insecticide application regimes that more effectively including regionally integrated resistance management strategies for effective malaria control and elimination.
Similar content being viewed by others

Background
Over decades, arthropods have been controlled using synthetic pesticides with dramatic reduction on target pest populations and their associated impacts of society [1, 2]. While effective, their prolonged and widespread use has unintentionally resulted in increased prevalence of pesticide resistance, with agricultural and medical implications [3, 4]. Pesticide resistance is typically a result of injudicious pesticide use and mounting pressure on population genetics (e.g. through natural selection and evolution) [5, 6]. Indeed, pesticide resistance is triggered by many genetic, [7] operational [8, 9] as well as biological [10] factors. For instance, selection pressure due to excessive use of insecticides may result in behavioral adaptation of vectors and subsequently, gene mutation expression, leading to temporal and spatial intra-specific heterogeneity [7]. While well-assessed in certain taxa and regions, pesticide resistance is dynamic in space and time and requires continuous evaluation. There are, however, regions where resistance has not been assessed. Furthermore, without empirical evidence for optimal efficacy, synthetic pesticidal active ingredients continue to be used within same localities.
Malaria in humans is an infectious disease, spread by various mosquito species in the Anopheles genus, which serve as bridge vector hosts for the Plasmodium spp. parasites that cause the disease. Given the role of mosquitoes in malarial transmission dynamics, their management is a crucial component of integrated malaria control strategies [11, 12]. Mitigation of the spread of malaria is typically reliant of vector monitoring and control, which most often involves the use of insecticides [13], although several other complementary approaches are also widely explored [14, 15]. With regards to insecticide resistance, there are four common synthetic insecticides that are capable of conferring resistance to insects; namely the organochlorines, organophosphates, carbamates and pyrethroids [16]. These insecticides are target site specific with organophosphates and carbamates inhibiting activity of the neuro-synaptic enzymes whilst the organochlorines and the pyrethroids target the sodium ion channels [17, 18]. Mosquitoes have shown alterations in their genetics (e.g. acetylcholinesterase genes), with consequent reduction in the binding efficiency with insecticides and hence reduced efficacy [19]. According to Williamson et al. [20], organochlorines and pyrethroid resistance emanates from point mutations in the voltage-gated sodium channels resulting in knockdown resistance (KDR). Cross resistance as a consequence, may occur when a resistance mechanism, also confers resistance to another insecticide [21], thus further occurring between pesticides from different chemical classes [22]. Evidence of multiple insecticide resistance in mosquito vectors, including Anopheles species, have been reported from many regions [23, 24]. Furthermore, Anopheles malaria vectors have also been shown to develop adaptive escape behaviours, through either learning or based on insecticide avoidance and/or repellency, creating further challenges for control and elimination of these vectors and associated infections [25, 26].
Insecticide resistance is a consistently worsening situation in Africa, requiring urgent intervention for effective control of malaria vector species [27, 28]. Malaria is the most prevalent mosquito-borne disease in the sub-Saharan Africa and is carried by various species of Anopheline mosquitoes. Botswana, situated in the warm subtropics of southern Africa is no exception, with malaria cases reported annually (~ 0.01% /1000 population) and even spreading to non-endemic parts of the country [29]. However, regardless of the pronged insecticide use for vector control in Botswana [30], mosquito susceptibility investigations are scant. For many decades lamda-cyhalothrin (pyrethroid) and dichloro diphenyl trichloroethane (DDT; organochlorine) have been the main insecticides used for mitigation against malaria vectors in Botswana [30], although recently (2019) pirimiphos-methyl (organophosphate) was deployed for indoor residual spraying (IRS) use across all malaria endemic districts [13]. Similarly, since the 1940s, DDT has been used in the country for IRS and later complimented by pyrethroids long-lasting insecticide nets (LLINs) and microbial larviciding (Bacillus thuringiensis serovar israelensis) [13, 31]. However, insecticide susceptibility status of malaria vectors is currently unexplored in Botswana. This is regardless of the country having been using these insecticides for > 70 years [30, 32], a time-scale that will likely have promoted resistance development. This may subsequently regress nationwide or regional planning initiatives on malaria elimination achievement targets by 2023 [33].
As part of a larger project on mosquito control in the region, here we conducted a baseline assessment on Anopheles arabiensis insecticide susceptibility for World Health Organization (WHO) recommended and currently used pesticides across three malaria endemic regions in Botswana [34]. Anopheles arabiensis is the biggest contributor to malaria in the region [35] and is widely distributed across malaria endemic and even non-endemic parts of the country [36,37,38]. Specifically, An. arabiensis susceptibility status to eleven registered insecticide products was assessed, comprising four classes of pesticides and determined their knockdown times (KDT50) and differences in susceptibility patterns across malaria endemic districts. It was hypothesized that (i) locally used insecticides and those with a similar mode of action, had differed efficacy on An. arabiensis, with (ii) knockdown times differed across recommended insecticides based on their mode of action and intensity of use, and (iii) that An. arabiensis susceptibility status will differ in space owing to differences in insecticide use.
Methods
Mosquito collection and maintenance
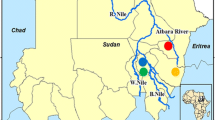
Mosquito larvae were collected from stagnant pools across human settlements in malaria endemic districts (Okavango (Mohembo): 18. 2876° S, 021. 7898° E, Ngamiland (Shorobe): 19. 7625° S, 023. 6774° E and Bobirwa (Mothabaneng): 22. 1051° S, 028. 5253° E) in austral summer season between February to March of 2016 and 2017 (Fig. 1). Each sampled district was represented by a village undergoing a deployment of chemical intervention (IRS and/or LLINs) through the country’s national malaria vector control programme. Site selection was based on proximity to human settlements. The collection was done using a 1000 µm mesh net with larvae transferred to a netted 3 L aerated container holding ~ 1.5 L 50:50 habitat and matured tap water, and transported in cooler boxes to the laboratory for further processing. In the laboratory, rearing containers were housed in climate chambers (HPP 260, Memmert GmbH + Co.KG, Germany) set at 25 °C ± 2 and 65% ± 10 relative humidity (RH) under a 12:12 light:dark photocycle. The larvae were fed with fish food (Sera: Vipan family, Randburg, South Africa) ad libitum daily and the water was exchanged with matured tap water every two days to prevent the built up of scum. Eclosed adults were fed with 10% sugar solution ad libitum soaked in a piece of cotton wool placed over the net. Adults were identified using gross morphology [39, 40] and confirmed as An. arabiensis following Bass et al. [41]. Adult mosquitoes were morphologically sexed upon eclosion using differences in antenna [40] with virgin females retained for use in bioassays.

Map showing (a) location of Botswana in Africa and (b) malaria endemic districts in the northern part of the country: [Okavango (1), Ngamiland (2), Chobe (3), Tutume (4), Boteti (5) and Bobirwa (6)], with study site villages (black up-pointing triangle) in Okavango (Mohembo), Ngamiland (Shorobe) and Bobirwa (Mothabaneng)
Insecticide susceptibility bioassays
The insecticide susceptibility bioassays were performed in accordance with the standard WHO procedure [34]. Virgin female mosquitoes (2 to 5 days old) were exposed (by contact) to surfaces impregnated by a discriminating insecticide doses for a period of 1 h to assess mortality [34]. The bioassays were performed with eleven insecticide types across three districts (n = 1100 per sampling site) grouped in four common recommended classes, namely; (i) organophosphates (malathion 5%), (ii) organochlorines (DDT 4.0%, dieldrin 0.4%, dieldrin 4%), (iii) carbamates (propoxur 0.1%, bendiocarb 0.1%) and (iv) pyrethroids (lambda-cyhalothrin 0.05%, permethrin 0.75%, deltamethrin 0.5%, cyfluthrin 0.15% and etofenprox 0.5%). A batch of 20 virgin female mosquitoes were transferred into a total of 5 holding tubes (n = 100 per insecticide), four of which (green dotted) were replications to be exposed to insecticides while the other was used as a control. A set of exposure tubes, four lined with impregnated insecticide (red dotted) and a control tube consisted of a paper lined with risella oils (organochlorines), silicone oils (pyrethroids) and olive oils (organophosphates and carbamates). The holding tubes and the exposure tubes were fastened together, and mosquitoes were transferred to the exposure tubes. Mosquitoes that were damaged during the transfer were replaced before the exposure period. After a successful transfer, the holding tubes were removed leaving the exposure tubes upside down on the slides. For every 15 min (up to 60 min), the knockdown mortalities were observed and recorded. At the elapse of 1 h, the mosquitoes were transferred back to the holding tubes then kept for the next 24 h with a cotton wool (soaked in 10% sugar solution) placed on top for feeding purposes. Mosquitoes that survived 24 h post this treatment may have developed resistance [34].
Data analysis
Mosquito mortality for insecticide susceptibility testing was analysed as percentage following corrected control mortality [34, 42, 43]. The formula was ignored if control mortality was below 5%, but was used only when the control mortality was between 5 and 20%. However, if control mortality was more than 20%, the experiment was discarded. Overall, mosquito efficacy to tested insecticides were defined as susceptible (≥ 98% mortality), suspected resistance (90- 97% mortality) and resistant (< 90% mortality) [29]. The knockdown time 50% (KDT50) of total tested female mosquitoes for each insecticide were pooled together and subjected to probit analysis from the Statistical Package for the Social Sciences (SPSS) software (Version 24) [43, 44].
Results
Insecticide susceptibility bioassays
Insecticide resistance (< 90% mortality) was recorded in Okavango and Ngamiland mainly for the pyrethroid pesticidal group (Table 1). Okavango showed prominent resistance to pyrethroids; lambda-cyhalothrin 0.05%, permethrin 0.75%, deltamethrin 0.5% and cyfluthrin 0.15% (78.8%, 78.8%, 81.3% and 83.8% mortality respectively) while Ngamiland mosquitoes exhibited resistance to lambda-cyhalothrin 0.05% and permethrin 0.75% (81.3% and 83.8% mortality respectively). Suspected resistance (90–97% mortality) to organochlorines was confirmed in all study sites (districts) (Table 1). Okavango recorded suspected resistance to DDT 4.0%, and dieldrin dosages (0.4 and 4%) at 96%, 93% and 95% mortality respectively. Similarly, Ngamiland exhibited suspected resistance to DDT 4.0%, and dieldrin dosages (0.4 and 4%) at 97.5%, 95% and 96.3% mortality, respectively, while Bobirwa mosquitoes yielded suspected resistance of 97.5% mortality to dieldrin (0.4%). Moreover, dieldrin (0.4%) showed suspected resistance across all districts tested. Suspected resistance was further observed in pyrethroids across districts sampled. Okavango mosquitoes reported suspected resistance of 97.5% mortality to etofenprox (0.5%). Ngamiland mosquitoes displayed suspected resistance to deltamethrin (0.5%), cyfluthrin (0.15%) and etofenprox (0.5%) at 92.5%, 97.5% and 96.3% mortality respectively. Mosquitoes in Bobirwa showed suspected resistance of 95%, 95% and 96.3% mortality to lambda-cyhalothrin (0.05%), permethrin (0.75%) and deltamethrin (0.5%) respectively. Susceptibility (≥ 98% mortality) to organophosphate (malathion (5%) and carbamates; propoxur (0.1%) and bendiocarb (0.1%) was recorded in mosquitoes from all the districts (Table 1). Bobirwa mosquitoes showed susceptibility to organochlorines; DDT (4.0%) and dieldrin (4%) both at 98.8% mortality. Similarly, they yielded susceptibility to pyrethroids; cyfluthrin (0.15%) and etofenprox (0.5%) both at 98.8% mortality.
Knockdown time (KDT50)
Carbamates [propoxur (0.1%) and bendiocarb (0.1%)] and organochlorines [dieldrin (0.4 and 4%)] had the highest mosquito KDT50 across sampled districts (Fig. 2). The lowest value (32.753 min) was recorded in Bobirwa [dieldrin (4%)] and the highest (47.994 min) in Okavango [propoxur (0.1%)]. DDT 4% was the only organochlorine which had the lowest KDT50 with the lowest value (25.721 min) reported in Ngamiland and highest in Bobirwa (31.229 min). Organophosphate [malathion (5%)] showed an intermediate KDT50 with consistent values for Okavango (39.073 min), Ngamiland (39.294 min) and Bobirwa (38.352 min). The pyrethroids; lambda-cyhalothrin 0.05%, permethrin 0.75% and deltamethrin 0.5% reported the lowest KDT50 compared to other classes of insecticides (Fig. 2). In particular, deltamethrin 0.5% scored the lowest value (17.28 min) in Bobirwa while lambda-cyhalothrin (0.05%) had the highest (23.559 min) for the same district. Conversely, amongst the pyrethroids, cyfluthrin (0.15%) and etofenprox (0.5%) had the highest mosquito KDT50 with the highest score (39.137 min) reported in Bobirwa [cyfluthrin (0.15%)] and the lowest (25.798 min) in Okavango [cyfluthrin (0.15%)]. Overall, the locally used insecticides (DDT and lambda-cyhalothrin yielded lower KDT50 than carbamates and the organophosphate tested (Fig. 2).

Mean ± 95% CL summary results of knockdown time (KDT50) (minutes) of field collected Anopheles arabiensis F1 progeny (n = 100 per insecticide) from malaria endemic districts tested against different classes of insecticides. Pyrethroids (etofenprox 0.5%, cyfluthrin 0.15%, deltamethrin 0.5%, permethrin 0.75%, lambda-cyhalothrin 0.05%), Carbamates (bendiocarb 0.1%, propoxur 0.1%), Organochlorines (dieldrin 4%, dieldrin 0.4%, dichloro-diphenyl-trichloroethane [DDT]) 4.0%), Organophosphates (malathion 5%)
Discussion
The study highlighted the general reduced efficacy of organochlorines and pyrethroids on An. arabiensis across malaria endemic areas. Results showed high resistance development for lambda-cyhalothrin (0.05%), permethrin (0.75%), deltamethrin (0.5%) and cyfluthrin (0.15%) in Okavango and for only lambda-cyhalothrin (0.05%) and permethrin (0.75%) in Ngamiland. It further showed that resistance dynamics were variable in space and likely as a result of differing insecticide intensities and nature of application regimes across districts [31]. In Botswana, a combination of vector control strategy using LLINs (pyrethroid based) and IRS (pyrethroid and DDT) have been practiced continuously over decades with first intervention (DDT) rolled out in Okavango, Ngamiland and Chobe districts in mid-1940s [13, 30]. Okumu and Moore [8] suggested that the two intervention strategies used together, may promote the evolution of insecticide resistance in mosquito populations due to increased pesticide selection pressure. Little information is, however, available regarding extent and difference among national vector control and even domestic insecticide use, across the three districts and its role in resistance development. What is known is that the Okavango and Ngamiland districts are formally considered more problematic for malaria and likely receive more intensive and frequent use of intervention strategies [45].
Botswana has been able to effectively reduce malaria cases over the years through chemical based intervention strategies [13, 31]. Parallel to this achievement, this study demonstrated compromised vector efficacy to insecticides used for IRS and LLINs, similar to observations in other parts of the world [46]. The reduced mosquito sensitivity to insecticides observed in this study may be due to the prolonged (> 70 years) use of IRS (DDT and lambda-cyhalothrin) and massive area-wide distribution of LLINs (pyrethroid impregnated) [32]. Moreover, lambda-cyhalothrin (pyrethroid) and DDT (organochlorine) are insecticides of similar mode of action [47]. Insensitivity to pyrethroids not currently registered for vector control in the country vis permethrin, deltamethrin, cyfluthrin and etofenprox was recorded. One of the reasons for this could be the prolonged use of lambda-cyhalothrin [13], with the same mode of action as these pesticides. Okavango showed the most prominent insecticide resistance which may be associated with extensive pesticidal usage since 1940s from the national vector control programme and household interventions, as an area of targeted malaria ‘hotspot’ compared to Ngamiland and Bobirwa [31, 32]. Indeed, the results showed that Bobirwa only had cases of suspected resistance. This may be associated with (i) the vector insecticidal intervention intensification strategy post 2012 in Bobirwa and its categorization as a malaria ‘hotspot’ area [31] and (ii) the interventions are less frequent/intense than other malaria endemic areas (e.g. Okavango and Ngamiland) [48]. Moreover, Simon et al. [31] reported public defiance in Bobirwa toward national intervention strategies (e.g. IRS), likely ‘delaying’ An. arabiensis resistance in the area. However, the data are only based on samples from one location (village) per district. As such, future work should consider monitoring susceptibility status of malaria vectors in more exhaustive human settlements receiving unique chemical intervention to establish other bio-physical factors contributing to insecticide resistance.
The results showed entire susceptibility to the organophosphate (malathion) and carbamates (propoxur, bendiocarb), irrespective of region. The country’s national vector control strategies are based on insecticides (pyrethroids and DDT) that target one site (voltage-gated sodium channel proteins), which may facilitate selection pressure for possible mutation. Therefore, it may be logical, from the perspective above to use insecticides with different modes of action (e.g. organophosphates and carbamates) on rotational/alternation to improve efficacy while simultaneously managing insecticide resistance [49]. To err on the side of caution regarding the deployment of insecticides in microhabitats (e.g. human habitation structures), abiotic factors shown to influence mosquitocides’ efficacy [50, 51] should also be considered during application. For example, temperature can interact with mosquito chemical intervention approaches (LLINs and IRS) within structures of ‘unstandardized’ thermal condition [50]. As such, assessing temperature coefficient (TC) of pesticides prior to regional use is recommended, as insecticides with positive TC are likely less efficacious at elevated temperatures [51, 52]. Future work on monitoring and evaluation of other Anopheles vectors [35] and mechanisms of resistance is warranted (although see Kgoroebutswe et al. [38]).
Assessment and selection of pesticides based on their time of action for vector control is an essential component that has a bearing in management of insecticide resistance. The KDT50 determines the time that enables 50% of mosquito population to be knocked down by an insecticide. Although it may be necessary to opt for insecticides that are fast in action (shorter KDT), this can be overruled if induced insecticide resistance is observed. The results demonstrated that pyrethroids generally reported shorter KDT50 than other classes tested. This is in keeping with Wakeling et al. [53] that this group of insecticide is fast in action. Regardless of their ability to knockdown mosquitoes within a short period of time, this group appeared not to be efficacious for An. arabiensis, at least in Okavango and Ngamiland, indicating potential development of pyrethroid resistance. In contrast, malathion, propoxur and bendiocarb were generally observed to be slow to action (high KDT50) across the study sites with mosquito vector susceptible to their discriminating dozes. Therefore, it may follow that, if mosquitoes do not show resistance, a fast-acting insecticide may be given priority of choice but with pyrethroid-resistance areas (as in Okavango), slow-acting insecticides (e.g. organophosphates and carbamates) may serve as alternatives. Hence, it may be important that vector response assessment on both the KDT50 and insecticide susceptibility status be carefully considered and merged appropriately for future insecticide selection and subsequently managing resistance. Furthermore, alternative vector control strategies [54], applied in an integrated and area-wide approach may help bridge resistance development.
Though pyrethroids were observed to be fast in action, An. arabiensis displayed a compromised sensitivity to the insecticides, which has implications for future vector control strategies using this pesticidal group. This baseline assessment work advocates for continuous monitoring of insecticide resistance to all potential mosquito vectors in the country before conclusive recommendations on susceptibility status are made. Moreover, malaria elimination in the country and region is a priority that necessitates efforts in vector management to monitor insecticide resistance. This may be achieved using integrated approaches that complement the current vector management strategies in minimizing resistance and at the same time delivering environmental benefits [55].
Conclusion
While requiring further investigation, the results suggest that An. arabiensis, one of the important malaria vectors in the country [35], may be showing a genetic drift towards resistance as reported in other southern African neighbouring countries [56, 57]. The current study adds to other reports on insecticide resistance in Africa and may be extended to other disease vectors on fine-to large-scale susceptibility to insecticides for both the endemic and non-endemic districts across diverse landscapes [43, 58]. With the resistance reported here, Botswana should integrate the current national intervention strategies with other approaches of vector management in minimizing resistance and simultaneously considering environmental benefits. This may include novel complimentary non-chemical ‘bio-friendly’ approaches targeting both immature and adult vector life stages [59, 60]. Furthermore, public records of governmental, industrial and private insecticidal use and availability should be considered to aid delimit drivers of resistance development. This may help integrated vector management, frameworks for pest decision-making, continued insecticide resistance monitoring, and the implementation of insecticide resistance management strategies while maintaining biodiversity and essential ecosystem services [61]. The country needs to be more conservative with the continuous use of pyrethroids especially in the Okavango delta, where insecticide resistance was evident and biodiversity sustenance is key for sustainable livelihoods and the tourism economy [62].
Availability of data and materials
The datasets during and/or analysed during the current study available from the corresponding author on reasonable request.
References
Cooper J, Dobson H. The benefits of pesticides to mankind and the environment. Crop Prot. 2007;26:1337–48.
Aktar W, Sengupta D, Chowdhury A. Impact of pesticides use in agriculture: their benefits and hazards. Interdiscip Toxicol. 2009;2:1–12.
Jin T, Zeng L, Lin Y, Lu Y, Liang G. Insecticide resistance of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in mainland China. Pest Manag Sci. 2011;67:370–6.
Liu N. Insecticide resistance in mosquitoes: impact, mechanisms, and research directions. Annu Rev Entomol. 2015;60:537–59.
Antonio-Nkondjio C, Sonhafouo-Chiana N, Ngadjeu CS, Doumbe-Belisse P, Talipouo A, Djamouko-Djonkam L, et al. Review of the evolution of insecticide resistance in main malaria vectors in Cameroon from 1990 to 2017. Parasit Vectors. 2017;10:472.
Sokhna C, Ndiath MO, Rogier C. The changes in mosquito vector behaviour and the emerging resistance to insecticides will challenge the decline of malaria. Clin Microbiol Infect. 2013;19:902–7.
Main BJ, Lee Y, Collier TC, Norris LC, Brisco K, Fofana A, et al. Complex genome evolution in Anopheles coluzzii associated with increased insecticide usage in Mali. Mol Ecol. 2015;24:5145–57.
Okumu FO, Moore SJ. Combining indoor residual spraying and insecticide-treated nets for malaria control in Africa: a review of possible outcomes and an outline of suggestions for the future. Malar J. 2011;10:208.
Sougoufara S, Harry M, Doucouré S, Sembène PM, Sokhna C. Shift in species composition in the Anopheles gambiae complex after implementation of long-lasting insecticidal nets in Dielmo. Senegal Med Vet Entomol. 2016;30:365–8.
Yayo AM, Ado A, Safiyanu M, Hemingway J. Selection for and biochemical characterization of DDT resistance in laboratory strains of Anopheles arabiensis. Afr J Biotechnol. 2020;19:344–52.
Ogunah JA, Lalah JO, Schramm KW. Malaria vector control strategies. What is appropriate towards sustainable global eradication? Sustain Chem Pharm. 2020;18:100339.
Ogunmodede AF. Mosquitoes and their medical importance. WJMBS. 2020;1:115–20.
Kgoroebutswe TK, Makate N, Fillinger U, Mpho M, Segoea G, Sangoro PO, et al. Vector control for malaria elimination in Botswana: progress, gaps and opportunities. Malar J. 2020;19:301.
Gueye CS, Newby G, Gosling RD, Whittaker MA, Chandramohan D, Slutsker L, et al. Strategies and approaches to vector control in nine malaria-eliminating countries: a cross-case study analysis. Malar J. 2016;15:2.
Macias VM, McKeand S, Chaverra-Rodriguez D, Hughes GL, Fazekas A, Pujhari S, et al. Cas9-mediated gene-editing in the malaria mosquito Anopheles stephensi by ReMOT Control. G3. 2020;10:1353–60.
Zhu W, Schmehl DR, Mullin CA, Frazier JL. Four common pesticides, their mixtures and a formulation solvent in the hive environment have high oral toxicity to honey bee larvae. PLoS ONE. 2014;9:e77547.
Tiwari S, Stelinski LL, Rogers ME. Biochemical basis of organophosphate and carbamate resistance in Asian citrus psyllid. J Econ Entomol. 2012;105:540–8.
Zhang Y, Du Y, Jiang D, Behnke C, Nomura Y, Zhorov BS, et al. The receptor site and mechanism of action of sodium channel blocker insecticides. J Biol Chem. 2016;291:20113–24.
Accrobessy K, Olé ML, Dorkenoo MA, Ossè RA, Akinro BA, Sidick A, et al. Status and genes involved in insecticide resistance in Anopheles gambiae sibling species in Lomé, (Togo), West Africa. J Mosq Res. 2017;7:84–95.
Williamson MS, Denholm I, Bell CA, Devonshire AL. Knockdown resistance (kdr) to DDT and pyrethroid insecticides maps to a sodium channel gene locus in the house fly (Musca domestica). Mol Gen Genet. 1993;240:17–22.
Yunta C, Hemmings K, Stevenson B, Koekemoer LL, Matambo T, Pignatelli P, et al. Cross-resistance profiles of malaria mosquito P450s associated with pyrethroid resistance against WHO insecticides. Pestic Biochem Physiol. 2019;161:61–7.
Ranson H, N’Guessan R, Lines J, Moiroux N, Nkuni Z, Corbel V. Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control? Trends Parasitol. 2011;27:91–8.
Djouaka RJ, Atoyebi SM, Tchigossou GM, Riveron JM, Irving H, Akoton R, et al. Evidence of a multiple insecticide resistance in the malaria vector Anopheles funestus in South West Nigeria. Malar J. 2016;15:565.
Nongley FN, Mugenzi LM, Terence E, Niang A, Wondji MJ, Tchoupo M, et al. Elevated Plasmodium sporozoite infection and multiple insecticide resistance in the principal malaria vectors Anopheles funestus and Anopheles gambiae in a forested locality close to the Yaoundé airport. Cameroon Wellcome Open Res. 2020;5:146.
Sougoufara S, Doucouré S, Sembéne PMB, Harry M, Sokhna C. Challenges for malaria vector control in sub-Saharan Africa: resistance and behavioral adaptations in Anopheles populations. J Vector Borne Dis. 2017;54:4–15.
Kreppel KS, Viana M, Main BJ, Johnson PCD, Govella NJ, Lee Y, et al. Emergence of behavioural avoidance strategies of malaria vectors in areas of high LLIN coverage in Tanzania. Sci Rep. 2020;10:14527.
Ranson H, Lissenden N. Insecticide resistance in African Anopheles mosquitoes: a worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016;32:187–96.
Letchuman S, Thanthrige SM, Shafras M, Premarathne AD. Fundamental biological mechanism and resistance of insect repellent which make worse the liability of malaria in emerging nations. Entomol Ornithol Herpetol. 2020;9:1–5.
Moakofhi K, Edwards JK, Motlaleng M, Namboze J, Butt W, Obopile M, et al. Advances in malaria elimination in Botswana: a dramatic shift to parasitological diagnosis, 2008–2014. Publ Health Action. 2018;8:S34–8.
Makate NM. A review of insecticide resistance status in Botswana. In: Insecticides Resistance. S Trdan, Ed. IntechOpen, Chapt. 13, 2016.
Simon C, Moakofhi K, Mosweunyane T, Libril HB, Nkomo B, Motlaleng M, et al. Malaria control in Botswana, 2008–2012: the path towards elimination. Malar J. 2013;12:458.
Chihanga S, Haque U, Chanda E, Mosweunyane T, Moakofhi K, Jibril HB, et al. Malaria elimination in Botswana, 2012–2014: achievements and challenges. Parasit Vectors. 2016;9:99.
Lover AA, Harvard KE, Lindawson AE, Gueye CS, Shretta R, Gosling R, et al. Regional initiatives for malaria elimination: building and maintaining partnerships. PLoS Med. 2017;14:e1002401.
WHO. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes. Geneva: World Health Organization; 2016.
Tawe L, Ramatlho P, Waniwa K, Muthoga CW, Makate N, Ntebela DS, et al. Preliminary survey on Anopheles species distribution in Botswana shows the presence of Anopheles gambiae and Anopheles funestus complexes. Malar J. 2017;16:106.
Chirebvu E, Chimbari MJ. Characterization of an indoor-resting population of Anopheles arabiensis (Diptera: Culicidae) and the implications on malaria transmission in Tubu village in Okavango subdistrict, Botswana. J Med Entomol. 2016;53:569–76.
Bango ZA, Tawe L, Muthoga CW, Paganotti GM. Past and current biological factors affecting malaria in the low transmission setting of Botswana: a review. Infect Genet Evol. 2020;85:104458.
Kgoroebutswe TK, Ramatlho P, Reeder S, Makate N, Paganotti GM. Distribution of Anopheles mosquito species, their vectorial role and profiling of knock-down resistance mutations in Botswana. Parasitol Res. 2020;119:1201–8.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). S Afr Inst M Res. 1987;1987:55.
Jupp PG. Mosquitoes of Southern Africa: culicinae and toxorhynchitinae. Johannesburg: Ekogilde Publishers; 1996.
Bass C, Williamson MS, Wilding CS, Donnelly MJ, Field LM. Identification of the main malaria vectors in the Anopheles gambiae species complex using a TaqMan real-time PCR assay. Malar J. 2007;6:155.
Abbott WS. Abbott’s formula. A method of computing the effectiveness of an insecticide. J Am Mosquito Contr. 1987;3:302–3.
Matowo NS, Munhenga G, Tanner M, Coetzee M, Feringa WF, Ngowo HS, et al. Fine-scale spatial and temporal heterogeneities in insecticide resistance profiles of the malaria vector, Anopheles arabiensis in rural south-eastern Tanzania. Wellcome Open Res. 2017;2:96.
Rahimi S, Vatandoost H, Abai MR, Raeisi A, Hanafi-Bojd AA. Status of resistant and knockdown of West Nile vector, Culex pipiens complex to different pesticides in Iran. J Arthropod-Borne Dis. 2019;13:284–96.
Motlaleng M, Edwards J, Namboze J, Butt W, Moakofhi K, Obopile M, et al. Driving towards malaria elimination in Botswana by 2018: progress on case-based surveillance, 2013–2014. Publ Health Action. 2018;8:S24–8.
Dhiman S, Yadav K, Rabha B, Goswami D, Hazarika S, Tyagi V. Evaluation of insecticides susceptibility and malaria vector potential of Anopheles annularis s.l. and Anopheles vagus in Assam. India PLoS One. 2016;11:e0151786.
Sanchez-Bayo FP. Insecticides mode of action in relation to their toxicity to non-target organisms. J Environ Anal Toxicol. 2012;S4:2.
Ministry of Health. Botswana malaria indicator survey 2012 report. Gaborone, Botswana: National Malaria Program, Ministry of Health; 2012.
Sparks TC, Nauen R. IRAC: mode of action classification and insecticide resistance management. Pestic Biochem Phys. 2015;121:122–8.
Glunt KD, Blanford JI, Paaijmans KP. Chemicals, climate, and control: increasing the effectiveness of malaria vector control tools by considering relevant temperatures. PLoS Pathog. 2013;9:e1003602.
Abiodun GJ, Maharaj R, Witbooi P, Okosun KO. Modelling the influence of temperature and rainfall on the population dynamics of Anopheles arabiensis. Malar J. 2016;15:364.
Glunt KD, Oliver SV, Hunt RH, Paaijmans KP. The impact of temperature on insecticide toxicity against the malaria vectors Anopheles arabiensis and Anopheles funestus. Malar J. 2018;17:131.
Wakeling EN, Neal AP, Atchison WD. Pyrethroids and their effects on ion channels. In: Pesticides-advances in chemical and botanical pesticides. Soundararajan RP, editor. IntechOpen. 2012;3: https://doi.org/10.5772/50330.
Wilson AL, Courtenay O, Kelly-Hope LA, Scott TW, Takken W, Torr SJ, et al. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl Trop Dis. 2020;14:e0007831.
Benelli G, Jeffries CL, Walker T. Biological control of mosquito vectors: past, present, and future. Insects. 2016;7:52.
Munhenga G, Masendu HT, Brooke BD, Hunt RH, Koekemoer LK. Pyrethroid resistance in the major malaria vector Anopheles arabiensis from Gwave, a malaria-endemic area in Zimbabwe. Malar J. 2008;7:247.
Nardini L, Christian RN, Coetzer N, Koekemoer LL. DDT and pyrethroid resistance in Anopheles arabiensis from South Africa. Parasit Vectors. 2013;6:229.
Padonou GG, Sezonlin M, Ossé R, Aizoun N, Oké-Agbo F, Oussou O, et al. Impact of three years of large scale Indoor Residual Spraying (IRS) and Insecticide Treated Nets (ITNs) interventions on insecticide resistance in Anopheles gambiae s.l. in Benin. Parasit Vectors. 2012;5:72.
Sarwar M, Salman M. Success stories of eco-friendly organically acceptable insecticides as natural products discovery. Int J Materials Chem Phys. 2015;1:392–8.
Muema JM, Bargul JL, Njeru SN, Onyango JO, Imbahale SS. Prospects for malaria control through manipulation of mosquito larval habitats and olfactory-mediated behavioural responses using plant-derived compounds. Parasit Vectors. 2017;10:184.
Chanda E, Ameneshewa B, Bagayoko M, Govere JM, Macdonald MB. Harnessing integrated vector management for enhanced disease prevention. Trends Parasitol. 2017;33:30–41.
Ramberg L, Hancock P, Lindholm M, Meyer T, Ringrose S, Sliva J, et al. Species diversity of the Okavango delta. Botswana Aquat Sci. 2006;68:310–37.
Acknowledgements
We thank the Botswana International University of Science and Technology (BIUST) for providing funding and infrastructure for the study. Also, we thank the Ministry of Health and Wellness (Botswana) for the research permit (PPME 13/18/1 PS V (353) and collaboration. Further, we sincerely acknowledge the contributions made by Tshepang Kabelo, Kamogelo Mmereke, Rebaone Motswagole and Nonofo Gotcha in data collection and processing.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Project conceptualization and management: MB RJW CN. Data curation: MB. Formal analysis: MB RJW CN. Investigation: MB CN. Visualization and validation: MB RJW CN. Writing—original draft: MB. Writing, review and editing: MB RJW CN. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Buxton, M., Wasserman, R.J. & Nyamukondiwa, C. Spatial Anopheles arabiensis (Diptera: Culicidae) insecticide resistance patterns across malaria-endemic regions of Botswana. Malar J 19, 415 (2020). https://doi.org/10.1186/s12936-020-03487-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-020-03487-z