
Abstract
Background
Polyhydroxybutyrate (PHB) has emerged as a promising eco-friendly alternative to traditional petrochemical-based plastics. In the present study, we isolated and characterized a new strain of Salinicola salarius, a halophilic bacterium, from the New Suez Canal in Egypt and characterized exclusively as a potential PHB producer. Further genome analysis of the isolated strain, ES021, was conducted to identify and elucidate the genes involved in PHB production.
Results
Different PHB-producing marine bacteria were isolated from the New Suez Canal and characterized as PHB producers. Among the 17 bacterial isolates, Salinicola salarius ES021 strain showed the capability to accumulate the highest amount of PHB. Whole genome analysis was implemented to identify the PHB-related genes in Salinicola salarius ES021 strain. Putative genes were identified that can function as phaCAB genes to produce PHB in this strain. These genes include fadA, fabG, and P3W43_16340 (encoding acyl-CoA thioesterase II) for PHB production from glucose. Additionally, phaJ and fadB were identified as key genes involved in PHB production from fatty acids. Optimization of environmental factors such as shaking rate and incubation temperature, resulted in the highest PHB productivity when growing Salinicola salarius ES021 strain at 30°C on a shaker incubator (110 rpm) for 48 h. To maximize PHB production economically, different raw materials i.e., salted whey and sugarcane molasses were examined as cost-effective carbon sources. The PHB productivity increased two-fold (13.34 g/L) when using molasses (5% sucrose) as a fermentation media. This molasses medium was used to upscale PHB production in a 20 L stirred-tank bioreactor yielding a biomass of 25.12 g/L, and PHB of 12.88 g/L. Furthermore, the produced polymer was confirmed as PHB using Fourier-transform infrared spectroscopy (FTIR), gas chromatography-mass spectroscopy (GC–MS), and nuclear magnetic resonance spectroscopy (NMR) analyses.
Conclusions
Herein, Salinicola salarius ES021 strain was demonstrated as a robust natural producer of PHB from agro-industrial wastes. The detailed genome characterization of the ES021 strain presented in this study identifies potential PHB-related genes. However, further metabolic engineering is warranted to confirm the gene networks required for PHB production in this strain. Overall, this study contributes to the development of sustainable and cost-effective PHB production strategies.
Similar content being viewed by others


Background
The increasing production of conventional plastics has resulted in the accumulation of tons of plastic waste in the environment, enduring for centuries due to their slow decomposition. These plastics derived from non-renewable resources like fossil fuels [1] have inspired researchers to explore environmentally friendly alternatives. The significance of bioplastics extends beyond their biocompatibility, renewability, non-toxicity, and biodegradability, as they can also be derived from various waste products including cellulose and polysaccharides [2].
Polyhydroxyalkanoates (PHAs) are polyester bioplastics that have garnered substantial attention as promising commercial and green alternatives for fossil fuel plastics due to their safe components and their high biodegradability [3]. PHAs can be fully degraded to carbon dioxide and water via microbial metabolic processes [4]. Polyhydroxybutyrate (PHB) is a short-chain length type of PHA and is formed by the condensation of 3-hydroxybutyric acid [5]. The PHB synthesis operon (phaCAB) is involved in the formation of PHB, playing a pivotal role in the conversion of acetyl-CoA to PHB [6]. PHB can be produced by biotechnological processes under controlled conditions. Consequently, the production or degradation of PHB does not impose any adverse impact on the environment.
Various bacterial strains can produce PHB as intracellular particles, like synthetic polymers. PHAs exhibit physico-chemical characteristics including flexibility and toughness [7]. Despite the inherent and environmental advantages of PHB over petrochemical-derived plastics, its industrial production is constrained by the high cost of the involved raw materials, which can constitute up to 40–50% of the total production cost [8].
Advancements in PHB production involve manipulating microbial strains, using mixed microbial cultures, diversifying substrates, and optimizing processes, including scaling up production. Researchers are employing synthetic biology strategies to enhance PHB production [9, 10]. However, these advancements also contribute to increased cost competitiveness, complexities associated with scale-up, and the challenge of ensuring high PHB yields. To overcome these challenges, continuous investigations are essential to identify naturally high PHB-producing strains, low-cost raw materials and optimize fermentation processes.
Investigating industrial or agricultural waste feedstocks as carbon sources for PHB production via microorganisms presents a promising avenue to reduce waste disposable and PHB production costs [11]. Among agro-industrial wastes, salted whey (SW), a major by-product of cheese manufacturing comprising 70–90% of milk content, and molasses, a by-product of the sugar manufacturing process containing approximately 50% (w/w) of total sugars are noteworthy candidates. Sucrose is the main sugar component in sugarcane molasses, which comprises glucose and fructose [12]. Molasses has previously been utilized as a substrate for PHB production by Bacillus sp. and Priestia sp. [13, 14].
Therefore, the aim of this study was to produce PHB naturally using different isolates of marine bacteria from the New Suez Canal in Egypt and to characterize the genetic pathways involved in PHB production in the most promising PHB-producing isolate. In addition, this study aimed to optimize the production conditions in small flasks and in a 20 L stirred-tank bioreactor and to perform a structural analysis of the produced PHB by FTIR, GC–MS, and NMR spectroscopy.
Methods
Isolation of PHB-producing bacteria from seawater
Marine samples were aseptically collected in a sterilized clean bottle from the New Suez Canal, Qantara City, Ismailia, Egypt. Each marine sample was serially diluted and plated into Sucrose/Yeast extract agar medium (SYA) (KH2PO4 1 g/L, MgSO4 0.5 g/L, yeast extract 4 g/L, and sucrose 20 g/L) [15] supplemented with 3% NaCl (SYANa) and incubated at 30˚C for 24 h. Thereafter, several bacterial colonies with distinct morphologies were selected and sub-cultured on SYA. The pure bacterial colonies were streaked on SYA slants, incubated at 30°C overnight, and then stored at 4°C for further use.
Screening the isolated bacteria for PHB production
The isolated bacterial colonies were stained with an alcoholic solution of Sudan Black B (SBB 0.02%). PHB-producing colonies exhibited a bluish-black color while non-PHB-producing colonies exhibited a white color [16]. Bacterial smears were prepared from the pure isolates grown on SYANa plates. Smears were stained with Sudan Black-B (SBB) solution for 10 min [17], followed by counterstaining with 0.5% aqueous safranine for 5s. Subsequently, the slides were examined under the light microscope (Leica MZ 16) (100x). Additionally, culture plates were subjected to SBB staining for 30 min, followed by light microscope examination.
Nile Blue-A stain was also used to stain bacterial smears prepared from the isolated strains according to the method described by Ostle and Holt [18]. The stained smears were examined under the fluorescence microscope (Leica CH 9435 Herrburg) at a wavelength of 490 nm. PHB granule-producing bacterial isolates exhibited a bright yellowish-orange fluorescence under the microscope. The PHB-producing isolates were selected and stored at -20°C for further use.
Submerged fermentation for PHB production
The PHB production was conducted in 250 ml Erlenmeyer flasks, each containing 100 ml of sterilized Sucrose/Yeast extract broth medium (SYB) supplemented with 3% NaCl (SYBNa). The flasks were inoculated with 1 ml of the bacterial inoculum (17 isolates) and then incubated at 25°C for 48 h. To assess the effect of temperature and shaking rate on the PHB production, the flasks were incubated at 25 and 30°C under both static and shaking conditions (110 rpm, Biobock Saewfc, France) for 48 h, using the selected isolate (ES021).
The dry cell weight (biomass, g/L) and PHB concentration (g/L) were determined at the end of the fermentation process to estimate the percentage of PHB yield.
Whole bacterial genome sequencing of the most promising PHB-producing isolate
Genomic DNA was extracted from the isolate ES021 that showed the highest levels of PHB production using Gentra puregene yeast/bacteria DNA purification kit (Qiagen, cat. no. 158567) according to the manufacturer’s instructions. The DNA libraries were prepared using Illumina Truseq DNA nano kit (cat no. 20015964) and sequenced on an Illumina Miseq with a coverage of approximately 50x. The quality of paired end sequencing reads was checked using FastQC tool [19]. The reads were then de-novo assembled using Unicycler (v0.4.8). QUAST tool (v5.0.2) was used to check the quality of genome assembly. The genome completeness of the assembled contigs was comprehensively assessed using Benchmarking Universal Single-Copy Orthologue (BUSCO v5.5.0) database bacteria and Gammaprotebacteria _0db10 [20]. The annotation of the assembled genome contigs was performed using RAST tool kit (RASTtk) [21]. Functional annotation was conducted using PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center on-line platform (www.bv-brc.org) and KEGG pathway database. For publication, the NCBI Prokaryotic Genome Annotation Pipeline was used later for the genome annotation [22]. The genome features were visualized through a circus plot using the Proksee platform (https://proksee.ca) [23].
For comparative genomics, the genetic distance between the genome sequence of ES021 strain and genomic sequences of closely related bacteria was calculated using Mash/MinHash provided by PATRIC genomic comparative analysis tool.
Phylogenetic analysis was performed using Molecular Evolutionary Genetics Analysis Version 11 (MEGA 11) to align the 16S rRNA sequences employing Muscle algorithm (Gap open: -400, gap extend: 0, and UPGMA clustering) [24]. Subsequently, the Maximum Likelihood (ML) method was utilized for constructing the 16S rRNA tree. Evolutionary distances were computed using the Kimura 2-parameter method [25], and the phylogeny was tested through 1000 bootstrap replicates. The resulting tree was visualized using Itol v6 [26]. The average nucleotide identity based on blast algorithm (ANIb) in JSpecies web server (www.ribocon.com) was implemented to align the whole genome sequences and generate an ANI matrix. Heatmap.2 function in R v4.3.2 was used to construct a dendrogram.
Optimization of PHB production process parameters using agro-industrial wastes
Optimization of the fermentation process parameters for maximum PHB production on SW and molasses using the bacterial strain Salinicola salarius ES021 was conducted in batches in 250 ml Erlenmeyer flask containing the fermentation medium (100 ml working volume). The effect of fermentation factors: sugar concentration, NaCl concentration, temperature, pH, shaking rate, inoculum size, and incubation period were investigated. The inoculum was prepared using SYBNa.
Raw materials
Salted whey (SW) containing 2% either, 4% salt, and 5% sugar, was obtained from Dairy Department, Faculty of Agriculture, Cairo University, Giza, Egypt. SW was clarified according to Gupte and Nair [27].
Sugarcane molasses containing 40.5% sugar, was obtained from Sugar and Integrated Industries Company, Hawamdia, Giza, Egypt. Molasses was clarified according to Panda et al. [28].
Effect of fermentation media on PHB production
Different media formulations were prepared from SW to study the impact of salt, sugar concentrations, and yeast extract (4 g/L) supplementation as well as mineral salts (KH2PO4 1 g/L and MgSO4 0.5 g/L) on PHB production as shown in Table 1. In M1 and M2 media, sugar and salt concentrations were maintained at 4 and 5%, respectively, with or without yeast extract and minerals. In M3 and M4, the salt concentration was adjusted to 3%, mirroring the salt concentration in SYBNA with and without yeast extract and minerals. M5 and M6 featured a reduced salt concentration of 2% to explore the impact of lower salt levels on PHB production. Constant sugar concentrations were maintained across most media formulations, except in M7 and M8, where sugar concentrations were decreased to 2% to assess their effect on PHB production. The fermentation media were prepared in 250 ml Erlenmeyer flasks, each containing 100 ml of the sterilized SW media, and inoculated with 1% inoculum from the 48-h-old culture of ES021 strain. The flasks were incubated at 30°C for 48 h. PHB concentration, bacterial dry cell weight (DCW), and PHB yield % were measured at the end of fermentation.
Effect of incubation temperature on PHB production
Fermentation medium M4 supplemented with 3% sugar and 5% salt concentration was incubated at five distinct temperatures (25, 27.5, 30, 32.5, and 35°C). The PHB production was estimated following a 48-h growth period of the ES021 strain.
pH values
The pH values of the fermentation medium M4 were adjusted at 6.2, 6.5, 7, 7.3, and 7.6. All flasks were inoculated with a 1% inoculum of the ES021 strain and incubated on a shaking incubator (110 rpm) for 48 h at 30°C. Thereafter, PHB yield % was quantified.
Inoculum size
To assess the impact of inoculum size on PHB production, different inoculum sizes (0.5, 1, 1.5, 2, 2.5, and 3 % v/v) were supplemented to the fermentation medium M4 (pH = 7). The flasks were incubated at 30°C with continuous shaking at 110 rpm for 48 h.
Shaking rate
To determine the effect of shaking rate, liquid medium M4 (pH = 7) was inoculated with 1% (inoculum size v/v) and incubated at 30°C under static conditions as well as with shaking at 110, 130, 150, and 190 rpm for 48 h.
Incubation period
The impact of incubation duration on the PHB production was assessed by inoculating the liquid medium M4 (pH = 7) with a 1% inoculum (v/v). The flasks were then incubated at 30 °C with shaking (150 rpm) for varying incubation periods, including 12, 24, 36, 48, 60, 72, 84, and 96 h.
Production of PHB from molasses and mixture of SW and molasses
Molasses was clarified to achieve sucrose concentration of 5 and 10% as a carbon source and supplemented with 3% NaCl to be utilized for PHB production using the ES021 strain. Furthermore, a combination of SW M4 medium and molasses were used as a fermentation medium for PHB production via the ES021 strain in a ratio of 1:1 and 2:1, resulting in sugar concentrations of 10 and 15%, respectively. All batches were carried out in conical flasks, each containing 100 ml volume, maintained at 30°C with shaking (150 rpm) for 48 h and an inoculum size of 1% (v/v).
Scaling up PHB production in a 20 L bioreactor:
To enhance the biomass and PHB accumulation by Salinicola salarius ES021 strain, batch cultivation was carried out in a 20 L stirred-tank bioreactor containing 15 L of optimized media (molasses 5% sugar, 3% NaCl, KH2PO4 1 g/L, yeast extract 4 g/L, and MgSO4 0.50 g/L) inoculated with a 1% of bacterial inoculum and maintained for a duration of 48 h. The operating conditions, including temperature, pH, aeration, and agitation speed were maintained at 30°C, 7.0, 2.5 vvm, and 150 rpm, respectively.
Determination of PHB dry weight, dry cell weight, and PHB yield
The concentration (dry weight) of extracted PHB was determined as g/L using the method described by Williamson and Willinson, as well as Hahn et al. [29, 30]. The total biomass dry cell weight (DCW) was estimated as g/L according to the method described by Kunioka and Nakamura as well as Hawas et al. [31, 32]. The efficiency of PHB production via the bacterial strain was expressed as PHB yield (%) and calculated as the ratio of PHB concentration (dry weight, g/L) to DCW (g/L), multiplied by 100.
Biopolymer chemical analysis
The structure of extracted PHB was analyzed using FTIR and GC-MS, and the produced spectrum was compared with the PHB standard (Sigma cat no: 29435-48-1). NICOLET 380 FTIR (Thermo Scientific, China) was applied to identify the functional groups in the produced polymer [33]. To determine the chemical composition of the produced polymer, GC-TSQ mass spectrometer (Thermo Scientific, USA) was used [34]. NMR analysis of the extracted PHB was performed according to the method described by Kanzariya et al. [35]. 1H NMR spectra were recorded on a Varian Mercury VX-300 NMR spectrometer. H1 spectra were run at 300 MHz in deuterated dimethylsulphoxide (DMSO-d6).
Statistical analysis
All experiments were conducted in triplicates, and all data were recorded as means of three replicates. The factors were considered significant if the P value is ≤ 0.05 using analysis of variance (ANOVA). Duncan’s multiple range test was used to test the significance of variance treatments (P < 0.05). The statistical analysis and figures were conducted using GraphPad Prism 7, Python v.3.9, and R v4.3.2.
Results and discussion
Isolation and screening of PHB- producing bacteria
Several bacterial isolates were isolated from the New Suez Canal, in Egypt. A total number of 36 bacterial isolates were screened for PHB production using both rapid-plate screening (Fig. 1A) and Sudan Black B staining of bacterial cells. Among the screened isolates, only 17 isolates were found to be PHB producers based on rapid-plate screening and Sudan Black B staining. Microscopic analysis revealed the presence of lipophilic black granules in stained bacterial cells (Fig. 1B), confirming PHB granule production. The isolates were further purified, Gram-stained, and examined using a light microscope (100x). Most isolates belonged to the genus Bacillus (15), with two identified as Gram-negative short rods. Confirmation of PHB production was achieved through Nile blue A staining, followed by fluorescence microscope examination (Fig. 1C). Sudan Black B and Nile blue A are lipophilic dyes characterized by their high selective binding to lipid-rich structures including PHB, allowing their visualization under microscope [36,37,38,39,40].

Photomicrograph of the isolate ES021 showing the produced PHB granules inside the cells A Rapid screening of PHB production by Sudan Black-B staining (culture staining) showing black spots indicative of PHB granules. B Sudan Black-B staining of a bacterial smear (slide staining) showing PHB granules (black spots) surrounding the cells. C Nile blue A staining of a bacterial smear (slide staining)
Submerged fermentation for PHB production
The batch culture technique was used to evaluate the PHB production in the selected 17 isolates. Figure 2 shows the percentage of isolates that produced different amounts of PHB.

The percentage of bacterial isolates that produce different PHB quantities ranging from 0.23 to 0.72 g/L
The results revealed that the two short-rods G-negative isolates produced the highest amount of PHB (0.71-0.72 g/L). Consequently, the isolate ES021 was selected to enhance PHB production in SYBNa medium by investigating the influence of incubation temperature (25 and 30°C) and shaking rate (static and 110 rpm) on the PHB productivity. At the end of the experiment, PHB concentration (g/L), DCW, and PHB yield % were estimated.
The incubation temperature was shown to affect the PHB production, with the the PHB amount (g/L) increasing from 0.72 to 0.77 g/L when the incubation temperature was raised from 25 to 30°C (Fig. 3A). Notably, incubation at 25°C favored biomass production, while incubation at 30°C was more promotive to PHB production, resulting in a 6.94% increase. Therefore, the PHB yield was enhanced when growing the isolate ES021 at 30°C. Consistently, previous studies reported PHB production by various species of G-bacteria is temperature dependent. Hawas et al reported a higher amount of PHB production by Pseudomonas sp. at 30°C (3.5 g/L) compared to 25°C (0.600 g/L) [32], Mostafa et al used Pseudodonghicola xiamenensis to accumulate the highest amount of PHB (15.54 g/L) at 30°C [2].

The effect of incubation temperature (A) and shaking rate (B) on the PHB productivity by the isolate ES021
Furthermore, growing the isolate ES021 under shaking conditions at 110 rpm demonstrated an improvement in PHB productivity. The PHB amount increased from 0.77 to 0.80 g/L at 30°C under shaking conditions (110 rpm) compared to static conditions. These findings highlight the importance of shaking for enhanced PHB production. Previous studies have shown that PHB yield significantly increased when growing Bacillus sp. at 37°C for 48 h under shaking at 150 rpm [37]. Similarly, PHB production by Bacillus mycoides DFC1 strain was influenced by incubation temperature and shaking rate, resulting in increased PHB amount (1.8 g/L) and biomass (3.2 g/L) with a yield of 57.20 % under incubation temperature at 37°C and shaking at 120 rpm for 48 h [41].
Genome characterization
To reveal the genetic features and identify the pathways involved in PHB production, whole genome sequencing of the isolate ES021 was conducted at 50× coverage. The whole genome was assembled using Unicycler into 24 contigs with a genome length of 3,823,256 bp and an average GC content of 62.67%. The N50 length (the shortest sequence length at 50% of the genome) is 382,245 bp, and the L50 count (the smallest number of contigs whose length sum produces N50 is 4). The completeness of genome assembly was evaluated using BUSCO database specific to bacteria and Gammaprotebacteria database, which encompasses 4085 genomes. BUSCO assessment revealed 123 complete single-copy orthologs, accounting for 99.2% completeness and only one missing in the Salinicola salarius ES021 strain (0.8%) (Fig. 4). These outcomes establish the high-quality nature of the Salinicola salarius ES021 genome, rendering it well suited for the precise identification of PHA and PHB synthesis genes.

BUSCO assessment of the genome assembly completeness using the bacteria and Gammaprotebacteria_odb10 database. Single-copy orthologs (123, 99.2%) were found in the Salinicola salarius ES021 genome matched with 124 orthologs in bacteria and Gammaprotebacteria database. Only one missing gene was identifed in the Salinicola salarius ES021 genome
The isolate ES021 was identified using the NCBI blast tool as Salinicola salarius and deposited in DDBJ/ENA/GenBank under the accession JARJLD000000000. The genome of Salinicola salarius was annotated using RASTtk applying the genetic code 11. This bacterium belongs to the Halomonadaceae family. This genome has 3530 protein-coding sequences (CDS), 60 transfer RNA (tRNA) genes, and 4 ribosomal RNA (rRNA) genes. The genome annotation included 926 hypothetical proteins and 2604 proteins with functional assignments. The proteins with functional assignments included 979 proteins with Enzyme Commission (EC) numbers, 823 with Gene Ontology (GO) assignments, and 741 proteins that were mapped to KEGG pathways.
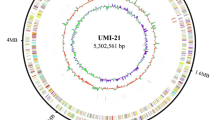
A circos plot of the genome annotation distributions is shown in Fig. 5A. This shows from outer to inner rings, the contigs, CDS on the forward strand, CDS on the reverse strand, RNA genes, GC content, and GC skew. The identified subsystems and the number of corresponding genes within each subsystem are shown in Fig. 5B, providing insights into the functional categories of the genome.

Genetic distribution of the genetic features and subsystems in the genome of Salinicola salarius strain ES021. A Circular graphical distribution of the genome annotation showing RNA genes, GC content, and GC skew. B An overview of the subsystems detected in this genome showing the number of subsystems and number of genes involved
The annotated genes have homology to known transporters, virulence factors, drug targets, and antibiotic resistance genes. The specialty genes and the specific source database where homology was found are shown in (Table 2). The list of annotated genes, their functions, sequences, and locations are shown in Additional file 2: Data Set 1.
Phylogenetic analysis of the 16S rRNA genes from Salinicola, Halomonas, and Chromohalobacter strains, conducted using the Maximum Likelihood (ML) algorithm, elucidated the phylogenetic relationships among these genera. As shown in Fig. 6A, the analysis revealed the placement of Salinicola salarius ES021 strain in relation to closely related strains within these genera.

Phylogenetic analysis of Salinicola salarius ES021 strain in relation to other taxonomic groups based on the 16S rRNA gene and whole genome sequences. A Maximum Likelihood (ML) tree based on 16S rRNA gene analysis depicts the phylogenetic relationships among closely related species. Bootstrap values exceeding 75% (based on 1000 replicates) are represented as bubbles on the tree. B The Average Nucleotide Identity (ANI) dendrogram of whole genome sequences. ANI similarity values are color-coded, ranging from 75% (shown in red) to 100% (shown in blue)
Salinicola salarius ES021 strain exhibited the highest similarity to the 16S rRNA gene of Salinicola salarius 1A01339 strain, demonstrating a close phylogenetic relationship between these two strains.
The alignment of whole genome sequences and determination of ANI% can enhance depth and robustness of phylogenetic analysis compared to relying solely on the 16S rRNA gene-based phylogenetic analysis. The complete genome sequence of Salinicola salarius ES021 strain was aligned with 18 genomes belonging to Salinicola, Halomonas, and Chromohalobacter strains. ANI analysis showed that Salinicola salarius ES021 strain is closely related to Salinicola salarius DSM 18044 (ANI > 97%) (Fig. 6B). Furthermore, Salinicola salarius ES021 strain showed a close genetic relationship to Salinicola socius strains (ANI > 85%).
Therefore, the implementation of 16S rRNA and ANI analyses consistently supports the taxonomic placement of Salinicola salarius ES021 strain as a strain of Salinicola salarius.
Mash/MinHash was used to estimate the genetic distance between Salinicola salarius ES021 strain and other sequenced Salinicola species (Additional file 1). The genetic distance between ES021 strain and Salincola salarius DSM 18044 strain is 0.01 showing a significant difference between both strains.
The genetic pathways for PHB and medium-chain-length PHA (MCL-PHA) production in Salinicola salarius ES021 strain were analyzed and predicted (Fig. 6). Interestingly, the PHA synthesis operon phaCAB does not exist in the ES021 strain. In most bacteria, PHB production proceeds in three steps mediated by the genes located in the phaCAB operon [42, 43]. These genes encode three enzymes including β-ketothiolase (PhaA), acetoacetyl-CoA reductase (PhaB), and PHA synthase (PhaC) [44, 45]. However, alternative genes that can function as phaCAB genes were predicted in Salinicola salarius strain ES021 leading to the construction of distinct PHB and MCL-PHA synthetic pathways (Fig. 7). List of putative genes that were predicted to be involved in the PHB and MCL-PHA production is shown in Table 5.

Overview of the PHB and MCL-PHA synthetic pathways in Salinicola salarius ES021 strain through EMP pathway, De novo fatty acid synthesis pathway, and fatty acid β-oxidation pathway
Acetyl CoA is the precursor of PHB synthesis, which is produced through the oxidation of pyruvate by the pyruvate dehydrogenase enzyme. Pyruvate is mainly produced through glucose metabolism via the Embden-Meyerhof-Parnas (EMP) pathway, pentose phosphate (PP) pathway, and Entner-Doudoroff (ED) pathway. The three pathways are detected in this ES021 strain using the KEGG pathway. Additionally, the genome of ES021 strain was found to harbor genes encoding enzymes involved in sucrose and starch metabolism. These enzymes include UTP-glucose-1-phosphate uridylyltransferase, glucokinase and alpha-amylase. Both UTP-glucose-1-phosphate uridylyltransferase and glucokinase play pivotal role in sucrose synthesis and degradation, while alpha-amylase is related to starch metabolism.
For PHB production from glucose in the ES021 strain, the enzyme 3-ketoacyl-CoA thiolase encoded by the fadA gene was speculated to be involved in the condensation of two acetyl-CoA molecules into acetoacetyl-CoA. 3-ketoacyl-CoA thiolase has been reported previously to catalyze the condensation reaction between two acyl-CoAs and acetyl-CoAs [46]. fadA gene was identified as a second copy of phaA gene in the halophilic marine bacterium Cobetia sp. MC34 [47]. The amino acid sequence of this gene has 95.7% similarity to the phaA gene in Cobetia marina DSM 4741T strain and 42.4% to the phaA gene in Halomonas sp. SF2003 [48].
The reduction of acetoacetyl-CoA to (S)-3-hydroxybutyryl-CoA in the ES021 strain is catalyzed by fabG that can function as phaB encoding 3-oxoacyl-ACP reductase. In the cyanobacterium Synechocystis sp., FabG has been shown to have the same function as PhaB and can replace PhaB as an acetoacetyl-CoA reductase for PHB production [49, 50]. In vivo studies showed that FabG in Pseudomonas aeruginosa can work as β-ketoacyl-CoA reductase for PHB production in E. coli [51]. Furthermore, the structures of both FabG and PhaB from Synechocystis sp. were shown to be very similar [52]. This indicates that FabG could act as an acetoacetyl-CoA reductase (phaB) in Salinicola salarius ES021 strain to produce PHB.
The (S)-3-hydroxybutyryl-CoA is converted into (R)-3-hydroxybutyryl-CoA by hydroxybutyryl-CoA epimerase enzyme. The conversion of 3-hydroxybutyryl-CoA into (R)-3-hydroxybutyrate was predicted in this study to be mediated by acyl-CoA thioesterase II that can function as PHB synthase (phaC) catalyzing the removal of CoA-SH from 3HB. PHB synthase is the key enzyme for PHB synthesis. It catalyzes the condensation of PHB monomers to PHB [53, 54]. In E. coli, thioesterase II was reported to be capable of hydrolyzing 3-hydroxybutyryl-CoA turning it into 3-hydroxybutyrate, which is the monomer of PHB [55, 56].
For PHB and MCL-PHA production from fatty acids, the acetyl-CoA is converted into malonyl-CoA by acetyl-CoA carboxylase carbonyl transferase to enter the De novo fatty acid synthesis pathway. A cascade of metabolic reactions is mediated in this pathway through the action of enzymes encoded by fadBDG genes (Fig. 6). In the fatty acid β-oxidation pathway, acyl-CoA acts as a precursor to produce both MCL-PHA and PHB. Acyl-CoA can be converted into acetyl-CoA through the action of enzymes encoded by phaJ and fadAB genes. The produced acetyl-CoA can mediate the production of PHB as aforementioned. MCL-PHA can be produced from acyl-CoA by converting it into 3-hydroxyacyl-CoA. Acyl-CoA thioesterase II enzyme can convert 3-hydroxyacyl-CoA into PHA through the removal of CoA-SH (Table 3).
Fermentation media
Salted whey (SW) served as the fermentation medium for PHB production, with different formulations tested involving different sugar and salt concentrations. Yeast extract and minerals were introduced into the salted whey to investigate their impact on PHB productivity. Table 4 presents the PHB quantities (g/L), dry cell weight (DCW, g/L), and PHB yield (%) by the ES021 strain under different SW medium conditions. The results reveal that the PHB production capacity of the ES021 strain is influenced by sugar concentration, salt (NaCl) levels, and the addition of yeast extract and minerals to the SW medium. Elevated salt concentrations exhibited a notable effect on PHB production, with a decrease of over 9% observed in M1 when salt concentration increased from 2 to 4%. Notably, the highest PHB production (3.46 g/L) was achieved in the M4 culture medium containing 5% lactose, supplemented with yeast extract and mineral salts (Table 4). Further investigation into the effect of sugar levels indicated that increasing lactose quantity in the fermentation medium stimulated PHB production. The optimal level for PHB production was found to be 5% lactose, resulting in a PHB amount of 2.56 g/L, with DCW reaching 4.56 g/L and a PHB yield of 56.14%. Thus, fermentation media containing 2% and 5% lactose levels were applied to study the influence of yeast extract and mineral salt supplementation on PHB productivity. The addition of yeast extract and mineral salt significantly influenced the conversion of lactose into PHB, irrespective of sugar or salt concentration. The highest PHB amount (3.46 g/L) was achieved when yeast extract and mineral salts were added to the SW medium containing 5% lactose and 3% NaCl (M4). Therefore, M4 was selected to study the impact of other growth parameters on PHB production. Whey from cheese, produced globally in substantial quantities, is a promising raw material for PHB production. The ES021 strain demonstrated the ability to accumulate a significant amount of PHB from lactose and SW, highlighting its potential in utilizing whey as a cost-effective substrate for PHB manufacturing. Numerous studies have documented PHB production from whey by indigenous microorganisms. Elmashi et al. and Szacherska et al. reported PHB production from whey using Bacillus cereus and Pseudomonas sp., respectively [57, 58]. Whey also necessitates a costly treatment prior to disposal, making it an attractive raw material for PHB manufacturing. ES021 strain has demonstrated a high capability to accumulate a significant amount of PHB from lactose and SW.
Incubation temperature
The ES021 strain was allowed to grow at five different temperatures (25, 27.5, 30, 32.5, and 35°C) to optimize PHB production. Given that the optimal temperature for Salincola salarius growth is 30°C, this temperature, along with two points below and higher, was selected for further PHB production assessment. Fig. 8 and Additional file 3: Table S1 illustrate that the incubation of the ES021 strain at 30°C resulted in the highest PHB production, reaching 3.46 g/L after 48 hours at pH=6.2. Lower yields were observed at 25°C, and the difference in PHB production at 30°C compared to 25°C was statistically significant (P < 0.0001). This finding aligns with previous studies demonstrating the significant impact of incubation temperature on PHB production [12, 59, 60]. For instance, Mostafa et al. achieved the highest PHB production by cultivating P. xiamenensis at 35°C for 96 hours [2].

Effect of different incubation temperatures on PHB production by the ES021 strain. The Incubation period was for 48 h under shaking at 110 rpm with an inoculum size of 1% in the fermentation medium M4. **** P < 0.0001. Data are presented as values of three independent replicates. The error bars represent the standard error of the mean
pH value
The ES021 strain demonstrated the highest PHB production at 30°C and pH 7, yielding 5.16 g/L of PHB with a corresponding PHB yield of 66.32% after 48 h (Fig. 9 and Additional file 3: Table S2). This finding agrees with previous studies suggesting that a pH of 7.5 is optimal for PHB production [61]. Additionally, studies on Rhizobium strains cultured in yeast extract mannitol broth indicated an increase in PHB production with the elevation of pH to 7 (from 0.01 to 0.5 g/L culture) [62].

Effect of different pH values on PHB production by the ES021 strain. Incubation was at 30°C for 48 h under shaking at 110 rpm with an inoculum size of 1% in the fermentation medium M4. ****P < 0.0001. Data are presented as values of three independent replicates. The error bars represent the standard error of the mean
Inoculum size
Various inoculum sizes (0.5, 1, 1.5, 2, and 2.5%) were examined to determine the optimum size for maximizing PHB production. As depicted in Fig. 10 and Additional file 3: Table S3, the ES021 strain exhibited significantly higher PHB levels (5.16 g/L, P < 0.0001) at an inoculum size of 1% compared to other sizes. Abdel Kareem et al. showed that increasing the inoculum size to 4 and 5% resulted in lower PHB amounts, when using whey as a fermentation medium [34].

Effect of different inoculum sizes on PHB production by the ES021 strain. Incubation was at 30°C for 48 h under shaking at 110 rpm at pH = 7 in the fermentation medium M4. **** P < 0.0001. Data are presented as values of three independent replicates. The error bars represent the standard error of the mean
Shaking rate
To assess the impact of shaking rate on PHB production (Fig. 11 and Additional file 3: Table S4), fermentation experiments were conducted under both static and shaking conditions (110, 130, 150, and 190 rpm). Shaking the culture during fermentation proved to be advantageous for achieving higher PHB production levels. The ES021 strain exhibited a significant increase in PHB production under shaking conditions. Elevating the shaking rate from 110 to 150 rpm resulted in an increase in PHB production. These findings align with previous research demonstrating that increasing the shaking rate enhances PHB production in bacteria [63].

Effect of shaking rate on PHB production by the ES021 strain. Incubation temperature was 30°C for 48 h in the fermentation medium M4 at pH = 7 and an inoculum size of 1%. *** P < 0.001. Data are presented as values of three independent replicates. The error bars represent the standard error of the mean
Incubation period
Figure12 and Additional file 3: Table S5 show the effect of various incubation periods on PHB production by the ES021 strain. The PHB production increased in a time dependent manner. A significantly higher amount of PHB (5.39 g/L) was obtained after 48 h. However, the production levels of PHB decreased to 2.95 and 1.21 after 72 and 96 h, respectively. Accordingly, these results revealed that the maximal PHB accumulation by the ES021 strain was at 48 h. Consistently, Bonartseva et al. showed that the observed maximal PHB accumulation by Rhizobium sp. was at 48 h [64]. This was also co-related entirely with previous studies showing that optimal time course for PHB production was at 48 h in Pseudomonas sp., Bacillus sp., and Rhizobium alti [12, 65].

Effect of the incubation period on PHB production by the ES021 strain. Incubation temperature was at 30°C and shaking at 150 rpm in the fermentation medium M4 at pH = 7 and an inoculum size of 1%. **P value < 0.01. Data are presented as values of three independent replicates. The error bars represent the standard error of the mean
Production of PHB from molasses and a mixture of salted whey and molasses
Molasses, an economical industrial by-product compared to glucose, was used in this study as a carbon source by the ES021 strain for PHB synthesis (Table 5). PHB productivity exhibited a significant increase when utilizing molasses containing 5% sucrose (13.34 g/L with a PHB yield of 87.25%). Conversely, PHB production amount and yield decreased when employing a mixture of SW and molasses (10 and 15%). Belal reported PHB production from molasses ranging from 0.7 to 2.2 g/L at inoculum sizes of 1 and 2%, respectively [63]. Page et al. and Beaulieu et al. demonstrated the use of molasses as the sole carbon source to produce PHB by Azotobacter vinelandii and Alcaligenes eutrophus, respectively [66, 67]. Gouda et al. obtained a maximum PHB production with sugarcane molasses and glucose as sole carbon sources (40.8 and 39.9 per mg cell dry matter, respectively). The 3% molasses resulted in higher biomass, while the maximum yield of PHB (46.2% per mg cell dry matter) was achieved with 2% molasses [68].
Scaling up PHB production in a 20 L bioreactor
To improve the PHB and biomass production of salinicola salaries ES021 strain, a batch fermentation was carried out in a 20 L stirred-tank bioreactor containing 15 L of the modulated medium (molasses 5%, NaCl 3%, yeast extract 4 g/L, MgSO4 0.5 g/L, KH2PO4 1 g/L). The temperature, pH, aeration, and agitation speed were maintained at 30°C, 7.0, 2.5 vvm, and 150 rpm, respectively. At 48 h of fermentation, PHB was accumulated by ES021 strain in an amount of 12.88 g/L, and the DCW was 25.12 g/L. Previous studies have demonstrated that the PHB production and biomass can be significantly enhanced in a 20 L stirred-tank bioreactor [69, 70].
FTIR analysis
The extracted polymer from the ES021 strain was used for determination of IR spectra. The FTIR spectrum of the produced PHB (Fig. 13) shows significant peaks at different wavelengths representing the PHB features. The FTIR spectrum of the produced polymer using sucrose as substrate was compared with the PHB standard and FTIR spectrum described by Brinda et al. [71]. The extracted polymer showed peaks at 3442, 2976 and 2934, 1731 and intense peaks at 1133 and 1053 cm-1. This indicates the presence of O–H stretching of alcohol, C–H stretching of alkanes, and C=O and C–O stretching of ester, respectively. Additionally, the range (899–511 cm-1) in the FTIR spectrum refers to the zone of carbon fingerprinting located in the PHB polymer [72].

FTIR spectrum of PHB produced by the ES021 strain
GC–MASS analysis
Table 6 shows the biodegradable compounds obtained from the produced polymer by the ES021 strain. GC–MS analysis of the PHB standard exhibited the twenty-three different biodegradable compounds existed in the produced polymer as shown in (Additional file 3: Table S6). The major detected compounds were 9-Octadecenoic acid (Z)-, methyl ester with a molecular weight of 296 in both standard and ES021 strain. The results were analyzed and compared with the standard and those described by Abdelrhman et al. and Okwuobi and Ogunjobiand and confirmed that the produced polymer is PHB [40, 73].
NMR analysis
The 1H NMR spectrum of the produced PHB exhibited a doublet signal for a methyl group (–CH3) at a chemical shift of δ = 1.17–1.23 ppm. Signals arising from a methylene group attached to a carbonyl group (–CH2) were observed at a chemical shift of δ = 2.5–2.55 ppm. Multiple signals observed at a chemical shift of δ = 5.07–5.19 ppm were identified as originating from a methine group (-CH). All these signals collectively validate the structural composition of P(3-HB) (Fig. 14).

1H NMR spectrum of the PHB extracted from Salinicola salarius ES021 strain showing signals at chemical shift of δ = 1.17–1.23 ppm (CH3), 2.5–2.55 ppm (CH2), and 5.07–5.19 ppm (CH)
Conclusions
Salinicola salarius ES021 strain is a potential and efficient natural-PHB producer from different carbon sources. The genome of the ES021 strain was characterized showing the genetic features and the putative PHB-related genes. Salinicola salarius ES021 strain has a unique genetic pathway for PHB production. However, further genetic engineering and construction of knockout mutants will be needed to confirm the involvement of the putative PHB-related genes in this strain. Optimization studies of the growth conditions were conducted in this study showing the effects of fermentation media, temperature, aeration, pH, inoculum size, and incubation period on the PHB production. Salincola salarius showed an enhanced PHB production on molasses. This indicates that Salinicola salarius ES021 strain can produce high amounts of PHB from different agro-industrial wastes such as salted whey and molasses.
Availability of data and materials
The datasets generated or analyzed during this study are available and included in this publication (and its Additional files). The genomic raw sequence data are accessible with the following link https://www.ncbi.nlm.nih.gov/sra/PRJNA943367.
Abbreviations
- PHB:
-
Poly(3-hydroxybutrate)
- FTIR:
-
Fourier-transform infrared spectroscopy
- GC-MS:
-
Gas chromatography-mass spectroscopy
- PHAs:
-
Polyhydroxyalkanoates
- SW:
-
Salted whey
- SYA:
-
Sucrose/Yeast extract agar
- SYANa:
-
Sucrose/Yeast extract agar medium supplemented with 3% Nacl
- SBB:
-
Sudan Black B
- SYB:
-
Sucrose/yeast extract broth
- SYBNa:
-
Sucrose/yeast extract broth medium supplemented with 3% NaCl
- CDS:
-
Protein-coding sequences
- NCBI :
-
National Center for Biotechnology Information
- RASTtk:
-
RAST tool kit
- DCW:
-
Dry cellweight
- tRNA:
-
Transfer RNA
- rRNA:
-
Ribosomal RNA
- EC:
-
Enzyme Commission
- GO:
-
Gene Ontology
- MCL-PHA:
-
Medium-chain-length PHA
- EMP:
-
Embden-Meyerhof-Parnas pathway
- PP:
-
Pentose phosphate pathway
- ED:
-
Entner-Doudoroff pathway
References
Khattab AM, Esmael ME, Farrag AA, Ibrahim MI. Structural assessment of the bioplastic (poly-3-hydroxybutyrate) produced by Bacillus flexus Azu-A2 through cheese whey valorization. Int J Biol Macromol. 2021;190:319–32.
Mostafa YS, Alrumman SA, Alamri SA, Otaif KA, Mostafa MS, Alfaify AM. Bioplastic (poly-3-hydroxybutyrate) production by the marine bacterium Pseudodonghicola xiamenensis through date syrup valorization and structural assessment of the biopolymer. Sci Rep. 2020. https://doi.org/10.1038/s41598-020-65858-5.
Zhou W, Bergsma S, Colpa DI, Euverink GJ, Krooneman J. Polyhydroxyalkanoates (PHAs) synthesis and degradation by microbes and applications towards a circular economy. J Environ Manage. 2023;341: 118033.
Koller M, Mukherjee A. A new wave of industrialization of PHA biopolyesters. Bioengineering. 2022;9:74.
Quispe MM, Lopez OV, Boina DA, Stumbé JF, Villar MA. Glycerol-based additives of poly (3-hydroxybutyrate) films. Polym Testing. 2021;93: 107005.
Nagajothi K, Murugesan AG. Polyhydroxy butyrate biosynthesis by Azotobacter chroococcum MTCC 3858 through groundnut shell as lignocellulosic feedstock using resource surface methodology. Sci Rep. 2023;13:10743.
Das M, Maiti SK. Recent progress and challenges in cyanobacterial autotrophic production of polyhydroxybutyrate (PHB), a bioplastic. J Environ Chem Eng. 2021;9: 105379.
Sakthiselvan P, Madhumathi R. Kinetic evaluation on cell growth and biosynthesis of polyhydroxybutyrate (PHB) by Bacillus safensis EBT1 from sugarcane bagasse. Eng Agric Environ Food. 2018;11:145–52.
Ray S, Jin JO, Choi I, Kim M. Recent trends of biotechnological production of polyhydroxyalkanoates from C1 carbon sources. Front Bioeng Biotechnol. 2023;10: 907500.
Sabapathy PC, Devaraj S, Meixner K, Anburajan P, Kathirvel P, Ravikumar Y, Zabed HM, Qi X. Recent developments in Polyhydroxyalkanoates (PHAs) production—a review. Biores Technol. 2020;306: 123132.
Nielsen C, Rahman A, Rehman AU, Walsh MK, Miller CD. Food waste conversion to microbial polyhydroxyalkanoates. Microb Biotechnol. 2017;10:1338–52.
Belal EB, Farid MA. Production of poly-β-hydroxybutyric acid (PHB) by Bacillus cereus. Int J Curr Microbiol Appl Sci. 2016;5:442–60.
Dalsasso RR, Pavan FA, Bordignon SE, de Aragão GM, Poletto P. Polyhydroxybutyrate (PHB) production by Cupriavidus necator from sugarcane vinasse and molasses as mixed substrate. Process Biochem. 2019;85:12–8.
Jung HJ, Kim SH, Shin N, Oh SJ, Hwang JH, Kim HJ, Kim YH, Bhatia SK, Jeon JM, Yoon JJ, Yang YH. Polyhydroxybutyrate (PHB) production from sugar cane molasses and tap water without sterilization using novel strain, Priestia sp. YH4. Int J Biol Macromol. 2023;250: 126152.
Bormann EJ, Leissner M, Beer B. Growth-associated production of poly (hydroxybutyric acid) by Azotobacter beijerinckii from organic nitrogen substrates. Appl Microbiol Biotechnol. 1998;49:84–8.
Liu M, González JE, Willis LB, Walker GC. A novel screening method for isolating exopolysaccharide-deficient mutants. Appl Environ Microbiol. 1998;64:4600–2.
Gerhardt P, Murray RG, Costilow RN, Nester EW, Wood WA, Krieg NR, Phillips GB. Manual of methods for general bacteriology. Washington, DC: American Society for Microbiology; 1981.
Ostle AG, Holt JG. Nile blue A as a fluorescent stain for poly-beta-hydroxybutyrate. Appl Environ Microbiol. 1982;44:238–41.
FastQC A. Quality control tool for high throughput sequence data. BibSonomy. https://www.bibsonomy.org/bibtex/f230a919c34360709aa298734d63dca3.2015. Accessed 17 Mar 2022.
Simão FA, Waterhouse RM, Ioannidis P, Kriventseva EV, Busco EZ. Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics. 2015. https://doi.org/10.1093/bioinformatics/btv351.:3210-2.
Brettin T, Davis JJ, Disz T, Edwards RA, Gerdes S, Olsen GJ, Olson R, Overbeek R, Parrello B, Pusch GD, Shukla M. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci Rep. 2015;5:8365.
Tatusova T, DiCuccio M, Badretdin A, Chetvernin V, Ciufo S, Li W. Prokaryotic genome annotation pipeline. In The NCBI Handbook. 2nd edn 2013. National Center for Biotechnology Information (US).
Grant JR, Enns E, Marinier E, Mandal A, Herman EK, Chen CY, Graham M, Van Domselaar G, Stothard P. Proksee: in-depth characterization and visualization of bacterial genomes. Nucleic Acids Res. 2023. https://doi.org/10.1093/nar/gkad326.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32(5):1792–7.
Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111–20.
Letunic I, Bork P. Interactive tree of life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47(W1):W256–9.
Gupte AM, Nair JS. β-galactosidase production and ethanol fermentation from whey using Kluyveromyces marxianus NCIM 3551. J Sci Ind Res (India). 2010;69:855–9.
Panda T, Kundu S, Majumdar SK. Studies on citric acid production by Aspergillus niger using treated Indian cane molasses. Process Biochem. 1984;19:183–7.
Williamson DH, Wilkinson JF. The isolation and estimation of the poly-β-hydroxy-butyrate inclusions of Bacillus species. Microbiology. 1958;19:198–209.
Hahn SK, Chang YK, Kim BS, Chang HN. Optimization of microbial poly (3-hydroxybutyrate) recover using dispersions of sodium hypochlorite solution and chloroform. Biotechnol Bioeng. 1994;44:256–61.
Kunioka M. New bacterial copolyesters produced in Alcaligenes eutrophus from organic acids. Polym Commun. 1988;29:174–6.
Hawas JM, El-Banna TE, Belal EB, El-Aziz AA. Production of bioplastic from some selected bacterial strains. Int J Curr Microbiol App Sci. 2016;5:10–22.
Sindhu R, Ammu B, Binod P, Deepthi SK, Ramachandran KB, Soccol CR, Pandey A. Production and characterization of poly-3-hydroxybutyrate from crude glycerol by Bacillus sphaericus NII 0838 and improving its thermal properties by blending with other polymers. Braz Arch Biol Technol. 2011;54:783–94.
Abd El-Kareem MS, Rabbih MA, Selim ET, Elsherbiny EA, El-Khateeb AY. Application of GC/EIMS in combination with semi-empirical calculations for identification and investigation of some volatile components in basil essential oil. Int J Anal Mass Spectrom Chromatogr. 2016;4:14–25.
Kanzariya R, Gautam A, Parikh S, Shah M, Gautam S. Structure analysis and thermal stability of PHB recovered from sugar industry waste. Biotechnol Genet Eng Rev. 2023. https://doi.org/10.1080/02648725.2023.2192076.
Upadhayay V, Verma S, Kuila A. Production of polyhydroxy butyrate (PHB) from Eichhornia crassipes through microbial fermentation process. Plant Sci Today. 2019;6(sp1):541–50.
Getachew A, Woldesenbet F. Production of biodegradable plastic by polyhydroxybutyrate (PHB) accumulating bacteria using low cost agricultural waste material. BMC Res Notes. 2016;9:1–9.
Bhagowati P, Pradhan S, Dash HR, Das S. Production, optimization and characterization of polyhydroxybutyrate, a biodegradable plastic by Bacillus spp. Biosci Biotechnol Biochem. 2015;79:1454–63.
Chetia J. Isolation and characterization of PHA producing bacteria from sewage samples of Assam. Int J Recent Technol Eng. 2019;8:10686–92.
Abdelrhman SA, Barakat OS, Elsayed TR. Biopolymer production by some marine bacterial strains isolated from Egypt. Egypt J Chem. 2022;65:513–23.
Aarthi N, Ramana KV. Identification and characterization of polyhydroxybutyrate producing Bacillus cereus and Bacillus mycoides strains. Int J Environ Sci. 2011;1:744.
Raberg M, Voigt B, Hecker M, Steinbüchel A. A closer look on the polyhydroxybutyrate-(PHB-) negative phenotype of Ralstonia eutropha PHB-4. PLoS ONE. 2014;9: e95907.
Haywood GW, Anderson AJ, Chu L, Dawes EA. Characterization of two 3-ketothiolases possessing differing substrate specificities in the polyhydroxyalkanoate synthesizing organism Alcaligenes eutrophus. FEMS Microbiol Lett. 1988;52:91–6.
Yanti NA, Sembiring L, Margino S, Ahmad SW. Bacterial production of poly-β-hydroxybutyrate (PHB): converting starch into bioplastics. In: Kuddus M, editor. Bioplastics for sustainable development. Singapore: Springer Singapore; 2021. p. 259–76. https://doi.org/10.1007/978-981-16-1823-9_9.
Celik PA, Barut D, Enuh BM, Gover KE, Yaman BN, Mutlu MB, Cabuk A. A novel higher polyhydroxybutyrate producer Halomonas halmophila 18H with unique cell factory attributes. Biores Technol. 2023;372: 128669.
Peoples OP, Sinskey AJ. Poly-β-hydroxybutyrate biosynthesis in Alcaligenes eutrophus H16: characterization of the genes encoding β-ketothiolase and acetoacetyl-CoA reductase. J Biol Chem. 1989;264:15293–7.
Schubert P, Steinbüchel A, Schlegel H. Cloning of the Alcaligenes eutrophus genes for synthesis of poly-beta-hydroxybutyric acid (PHB) and synthesis of PHB in Escherichia coli. J Bacteriol. 1988;170:5837–47.
Liu L, Zhou S, Deng Y. The 3-ketoacyl-CoA thiolase: an engineered enzyme for carbon chain elongation of chemical compounds. Appl Microbiol Biotechnol. 2020;104:8117–29.
Christensen M, Jablonski P, Altermark B, Irgum K, Hansen H. High natural PHA production from acetate in Cobetia sp. MC34 and Cobetia marina DSM 4741T and in silico analyses of the genus specific PhaC2 polymerase variant. Microb Cell Fact. 2021;20:1–23.
Zhang H, Liu Y, Yao C, Cao X, Tian J, Xue S. FabG can function as PhaB for poly-3-hydroxybutyrate biosynthesis in photosynthetic cyanobacteria Synechocystis sp. PCC 6803. Bioengineered. 2017;8:707–15.
Koch M, Forchhammer K. Polyhydroxybutyrate: a useful product of chlorotic cyanobacteria. Microbial Physiol. 2021;31:67–77.
Nomura CT, Taguchi K, Gan Z, Kuwabara K, Tanaka T, Takase K, Doi Y. Expression of 3-ketoacyl-acyl carrier protein reductase (fabG) genes enhances production of polyhydroxyalkanoate copolymer from glucose in recombinant Escherichia coli JM109. Appl Environ Microbiol. 2005;71:4297–306.
Braunegg G, Lefebvre G, Genser KF. Polyhydroxyalkanoates, biopolyesters from renewable resources: physiological and engineering aspects. J Biotechnol. 1998;65:127–61.
Mascarenhas J, Aruna K. Screening of polyhydroxyalkonates (PHA) accumulating bacteria from diverse habitats. J Glob Biosci. 2017;6:4835–48.
Perez-Zabaleta M, Sjöberg G, Guevara-Martínez M, Jarmander J, Gustavsson M, Quillaguamán J, Larsson G. Increasing the production of (R)-3-hydroxybutyrate in recombinant Escherichia coli by improved cofactor supply. Microb Cell Fact. 2016;15:1.
Guevara-Martínez M, Perez-Zabaleta M, Gustavsson M, Quillaguamán J, Larsson G, van Maris AJ. The role of the acyl-CoA thioesterase “YciA” in the production of (R)-3-hydroxybutyrate by recombinant Escherichia coli. Appl Microbiol Biotechnol. 2019;103:3693–704.
Elmashi TR, Eldanasoury M, Khattab AM. Production of bio plastic (polyhydroxyalkanoates) from cheese whey by isolated Bacillus cereus. Al-Azhar J Agric Res. 2023;0(0):0–0. https://doi.org/10.21608/ajar.2022.142504.1062.
Szacherska K, Moraczewski K, Czaplicki S, Oleskowicz-Popiel P, Mozejko-Ciesielska J. Effect of short-and medium-chain fatty acid mixture on polyhydroxyalkanoate production by Pseudomonas strains grown under different culture conditions. Front Bioeng Biotechnol. 2023;10: 951583.
Sangkharak K, Prasertsan P. Optimization of polyhydroxybutyrate production from a wild type and two mutant strains of Rhodobacter sphaeroides using statistical method. J Biotechnol. 2007;132:331–40.
Pozo C, Martınez-Toledo MV, Rodelas B, Gonzalez-Lopez J. Effects of culture conditions on the production of polyhydroxyalkanoates by Azotobacter chroococcum H23 in media containing a high concentration of alpechın (wastewater from olive oil mills) as primary carbon source. J Biotechnol. 2002;97:125–31.
Narayanan M, Kandasamy S, Kumarasamy S, Gnanavel K, Ranganathan M, Kandasamy G. Screening of polyhydroxybutyrate producing indigenous bacteria from polluted lake soil. Heliyon. 2020. https://doi.org/10.1016/j.heliyon.2020.e05381.
Aslim B, Yuksekdag ZN, Beyatli Y. Determination of PHB growth quantities of certain Bacillus species isolated from soil. Turk Electr J Biotechnol. 2002;52:24–30.
Belal EB, Farid MA. Production of poly-β-hydroxybutyric acid (PHB) by Rhizobium elti and Pseudomonas stutzeri. Curr Res J Biol Sci. 2013;5:273–84.
Bonartseva GA, Myshkina VL, Zagreba ED. Poly-β-hydroxybutyrate content in cells of various Rhizobium species during growth with different carbon and nitrogen sources. Microbiology (New York, NY). 1994;63:45–8.
Lathwal P, Nehra K, Singh M, Jamdagni P, Rana JS. Optimization of culture parameters for maximum polyhydroxybutyrate production by selected bacterial strains isolated from rhizospheric soils. Pol J Microbiol. 2015;64:7.
Page WJ. Production of poly-β-hydroxybutyrate by Azotobacter vinelandii UWD in media containing sugars and complex nitrogen sources. Appl Microbiol Biotechnol. 1992;38:117–21.
Beaulieu M, Beaulieu Y, Melinard J, Pandian S, Goulet J. Influence of ammonium salts and cane molasses on growth of Alcaligenes eutrophus and production of polyhydroxybutyrate. Appl Environ Microbiol. 1995;61:165–9.
Gouda MK, Swellam AE, Omar SH. Production of PHB by a Bacillus megaterium strain using sugarcane molasses and corn steep liquor as sole carbon and nitrogen sources. Microbiol Res. 2001;156:201–7.
Ali I, Jamil N. Enhanced biosynthesis of poly (3-hydroxybutyrate) from potato starch by Bacillus cereus strain 64-INS in a laboratory-scale fermenter. Prep Biochem Biotechnol. 2014;44:822–33.
Gamal RF, Abdelhady HM, Khodair TA, El-Tayeb TS, Hassan EA, Aboutaleb KA. Semi-scale production of PHAs from waste frying oil by Pseudomonas fluorescens S48. Braz J Microbiol. 2013;44:539–49.
Devi AB, Nachiyar CV, Kaviyarasi T, Samrot AV. Characterization of polyhydroxybutyrate synthesized by Bacillus cereus. Int J Pharm Pharm Sci. 2015;7:140–4.
Suhazsini P, Keshav R, Narayanan S, Chaudhuri A, Radha P. A study on the synthesis of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) by Bacillus megaterium utilizing cheese whey permeate. J Polym Environ. 2020;28:1390–405.
Okwuobi PN, Ogunjobi AA. Production and analysis of polyhydroxyalkanoate (PHA) by Bacillus megaterium using pure carbon substrates. World Appl Sci J. 2013;28:1336–40.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). The authors did not receive support from any organization for the submitted work and have no relevant financial to declare.
Author information
Authors and Affiliations
Contributions
SAA performed the experimental work (isolation of PHB producing bacteria; extraction of the polymer and estimation of the PHB amount), also organized the data, conducted the statistical analysis, figures, and manuscript writing. OSB performed the conceptualization of this research work, planned the methodology, manuscript writing and editing. MNA conducted the whole genome analysis, statistical analysis, figures, and manuscript writing and editing. All authors commented on previous versions of the manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
The genetic distance between Salinicola salarius ES021 strain and other sequenced Salinicola species and other information related to other sequenced Salinicola species.
Additional file 2.
The list of annotated genes, their functions, sequences, and locations in Salinicola salarius ES021 strain.
Additional file 3: Table S1.
Effect of different incubation temperatures on PHB production by ES021. Table S2. Effect of different pH values on PHB production by ES021. Table S3. Effect of different inoculum sizes on PHB production by ES021. Table S4. Effect of different shaking rates on PHB production by ES021. Table S5. Effect of different incubation periods on PHB production by ES021. Table S6. GCMS analysis of PHB standard showing chemical composition of biodegradable polymer.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Abdelrahman, S.A., Barakat, O.S. & Ahmed, M.N. Genetic characterization of a novel Salinicola salarius isolate applied for the bioconversion of agro-industrial wastes into polyhydroxybutyrate. Microb Cell Fact 23, 56 (2024). https://doi.org/10.1186/s12934-024-02326-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-024-02326-z