
Abstract
Background
Mucositis is one of the most relevant gastrointestinal inflammatory conditions in humans, generated by the use of chemotherapy drugs, such as 5-fluoracil (5-FU). 5-FU-induced mucositis affects 80% of patients undergoing oncological treatment causing mucosal gut dysfunctions and great discomfort. As current therapy drugs presents limitations in alleviating mucositis symptoms, alternative strategies are being pursued. Recent studies have shown that the antimicrobial pancreatitis-associated protein (PAP) has a protective role in intestinal inflammatory processes. Indeed, it was demonstrated that a recombinant strain of Lactococcus lactis expressing human PAP (LL-PAP) could prevent and improve murine DNBS-induced colitis, an inflammatory bowel disease (IBD) that causes severe inflammation of the colon. Hence, in this study we sought to evaluate the protective effects of LL-PAP on 5-FU-induced experimental mucositis in BALB/c mice as a novel approach to treat the disease.
Results
Our results show that non-recombinant L. lactis NZ9000 have antagonistic activity, in vitro, against the enteroinvasive gastrointestinal pathogen L. monocytogenes and confirmed PAP inhibitory effect against Opportunistic E. faecalis. Moreover, L. lactis was able to prevent histological damage, reduce neutrophil and eosinophil infiltration and secretory Immunoglobulin-A in mice injected with 5-FU. Recombinant lactococci carrying antimicrobial PAP did not improve those markers of inflammation, although its expression was associated with villous architecture preservation and increased secretory granules density inside Paneth cells in response to 5-FU inflammation.
Conclusions
We have demonstrated for the first time that L. lactis NZ9000 by itself, is able to prevent 5-FU-induced intestinal inflammation in BALB/c mice. Moreover, PAP delivered by recombinant L. lactis strain showed additional protective effects in mice epithelium, revealing to be a promising strategy to treat intestinal mucositis.
Similar content being viewed by others


Background
Mucositis is a gastrointestinal inflammatory disorder caused by radiotherapy or chemotherapeutic agents in oncology patients [1,2,3]. This enfeebling condition has been reported in 80% of patients undergoing clinical treatment with 5-fluorouracil (5-FU), a drug commonly prescribed for treating several types of cancer, including gastrointestinal, breast, pancreas, head and neck [2, 4]. As 5-FU present nonspecific cytotoxicity to cells with high replication rate, it inhibits the proliferation of both cancer and normal cells, such as the enterocytes lining the digestive tract [5, 6]. Hence, this process often leads to mucosal alterations, characterized by leukocyte infiltration, ulcers, villus shortening and decreased villus/crypt ratio, favoring systemic translocation of harmful bacteria colonizing the gut, increasing thus the possibility of infections and fatal consequences [7,8,9,10,11]. Moreover, 5-FU-induced mucositis has been associated with mucosal gut dysfunctions, affecting patients’ food consumption, provoking vomiting, abdominal pain, and diarrhea which may cause dehydration [1, 3].
Up to date, mucositis treatment relies mostly on mucosal coatings, cryotherapy, antibiotics and analgesics administration. However, no current therapy has been efficient to alleviate the disease [12, 13]. Thus, alternative strategies are currently being investigated. Contemporary studies have described promising achievements with the administration of lactic acid bacterium (LAB), instead of drugs [14,15,16,17,18]. These bacteria are considered to be probiotics, a term defined as “live microorganisms administered in adequate amounts that confer a beneficial health effect on the host” [19].
Species like Streptococcus thermophilus and several Lactobacilli have been shown to reduce intestinal inflammation caused by chemotherapy drugs injection, such as 5-FU, irinotecan or methotrexate, in rats [14,15,16]. Moreover, administration of yogurts containing L. johnsonii or L. bulgaricus and S. thermophilus, seems to be useful to restore intestinal barrier function in these animals [17].
The use of recombinant LAB strains, such as the model Lactococus lactis for delivering biologically active molecules with anti-inflammatory properties has also been explored as an alternative therapy for the treatment of mucositis and other gastrointestinal inflammatory disorders, as inflammatory bowel diseases (IBD) [20,21,22,23]. In 2006, a biological confinement strategy to contain the dissemination of genetically engineered L. lactis strain expressing IL-10 was carried out in phase I clinical trial with IBD patients, suggesting the feasibility of mucosal therapy using recombinant lactococci [24]. This trial provided novel possibilities for testing genetically modified L. lactis to treat similar intestinal disorders, such as mucositis. In fact, recent studies have designed recombinant strains of L. lactis to produce anti-inflammatory proteins, involved in the maintenance of epithelial barrier integrity, such as Trefoil factor 1 (TFF-1), which has been promising to treat mucositis in clinical trial as well [25, 26]. Therefore, other key elements in the host-microbiota relationship are being glimpsed as potential candidates to be cloned in L. lactis.
Delivery of antimicrobial peptides (AMP) that protects the host by killing harmful bacteria has been shown to prevent inflammation in colitis mice models [27, 28]. This positive effect was associated with host epithelial cell surface protection against pro-inflammatory bacteria in the intestinal mucosa [27, 28]. As AMP have been recently considered to be a very effective approach to fight inflammation, various types are being explored in basic research such as Reg3A, also known as pancreatitis associated protein (PAP). It has been extensively studied due to its protective effect in the intestinal inflammatory process [29,30,31,32,33,33]. This protein belongs to the RegIII gene sub-family, which encodes proteins involved in the regulation of epithelial cell proliferation and antimicrobial activity in several organs, including intestines [29,30,31,32,32]. In the small intestine, PAP is mainly produced by Paneth cells that are located in mucosal crypts and exerts bactericidal activity against Gram-positive bacteria species that might pose risk of infection to the host [32, 33]. Recently, our research group, constructed and confirmed the expression of human PAP by recombinant L. Lactis NZ9000 using the inducible Nisin Control Expression System (NICE) [34]. This work evaluated the therapeutic effect of this strain in mice model of dinitrobenzenosulfonic acid (DNBS)-induced colitis. We found out that PAP delivered by lactococci revealed to be anti-inflammatory [34]. As this strategy have shown to be useful in the treatment of IBD, we sought to investigate LL-PAP protective role in another important inflammatory gastrointestinal disorder for which conventional therapy is not enough, as mucositis, using the 5-FU intestinal mucositis experimental mouse model.
Methods
Bacterial strains and growth conditions
Lactococcus lactis NZ9000 strain harboring pSEC:PAP vector (LL-PAP) and L. lactis NZ9000 strain [35] carrying pSEC vector without the open reading frame of PAP (LL) [36], were grown in M17 medium (Difco) supplemented with 0.5% glucose (GM17) at 30 °C without shaking. Recombinant strains were selected by the addition of chloramphenicol (Cm, 10 µg/mL). For nisin-induced PAP expression, LL-PAP was cultivated until the optical density at 600 nm reached 0.6. Afterwards, 10 ng/mL of nisin (Sigma) were added to the medium and cultures were maintained at 30 °C for 2 h before experimentation. For in vitro antagonistic assays, L. lactis strains were grown in brain–heart infusion (BHI) medium containing Cm (10 µg/mL) at 30 °C, as well as Listeria monocytogenes ATCC 15313 and Enterococcus faecalis ATCC 19433, which was grown in BHI containing or not Cm (10 µg/mL) at 37 °C without shaking.
Antagonistic activity assay against pathogenic Listeria monocytogenes and Enterococcus faecalis
Antimicrobial activity of PAP secreted by LL-PAP strain was assessed against the food-borne pathogen L. monocytogenes or the commensal opportunistic E. faecalis using a previously described method [37]. Briefly, LL and LL-PAP were grown separately in BHI medium containing Cm (10 µg/mL) and, after OD600 reached 0.6, 10 ng/mL of nisin were added. Cultures were then centrifuged in an OD600 of 1.0, at 4000 g for 10 min and supernatants were sterilized using 0.20 µm Millipore filters (Sarstedt, Nümbrecht, Germany). L. monocytogenes or E. faecalis were then inoculated separately into filter-sterilized lactococcal supernatant or in BHI medium containing Cm (10 µg/mL) and nisin (10 ng/mL) at an initial OD600 of 0.1. Cultures were incubated at 37 °C and after 2 and 4 h respectively, serial dilutions were seeded in BHI agar plates that were maintained at 37 °C for 24 h. Thereafter, number of viable L. monocytogenes or F. faecalis was estimated through counting of bacterial colony forming units (CFU).
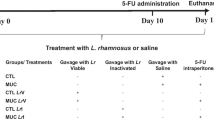
Animals, bacterial administration and experimental groups
Conventional female BALB/c mice between 6 and 8 weeks of age were obtained at Federal University of Minas Gerais (UFMG–Belo Horizonte, Brazil) and the study was approved by the Brazilian Ethics Committee on Animal Use (CEUA). Mice were kept in a temperature-controlled room with ad libitum access to water and standard chow diet 24 h prior to experiments. Animals were fed daily orally by drinking 5 mL of water; or 5 mL of M17 medium supplemented with Cm (10 µg/mL) and nisin (10 ng/mL) (M17/Cm/Nisin); or with 5 mL of M17/Cm/Nisin containing 2.5 × 109 CFU/mL of either LL or LL-PAP strains for 13 days. In order to induce mucositis, mice received a single intraperitoneal injection of 5-fluoracil (300 mg/kg) on day 10 and were euthanized on day 14. An injection of saline (NaCl 0.9%) was used as a control. For experimentation, BALB/c mice were divided into eight groups, each containing 6–9 animals. Animals from group 1–4 were injected with 0.9% saline on day 10 (noninflamed groups); group 1 received water, group 2 were fed with M17/Cm/Nisin medium; group 3 were administered with LL culture, group 4 received L. lactis expressing PAP. Mouse from group 5–8 were injected with 5-FU on day 10 (inflamed groups): group 5 received water; group 6 received M17/Cm/Nisin medium; group 7 were administered with LL culture and finally group 8 received LL-PAP strain.
Intestinal histology and morphology
After euthanasia, the distal portion of the small bowel (ileum) from the animals was collected, and, after washing, rolls were prepared for histomorphological analysis. Rolls were fixed with 10% buffered formaldehyde. Material was then embedded in paraffin, and a 4 µm section of each sample was placed on a glass slide and stained with hematoxylin and eosin (HE). Slides of each experimental group were photographed using a digital camera (Moticam 2500, China) coupled to an optical microscope (Olympus Optical Co., Japan). The histological score was determined using a previously described method [10], which measures the intensity of both mononuclear and polymorphonuclear cells infiltrate in the lamina propria, changes in mucosal architecture and presence of ulceration. For each parameter it was used the ranking values: absent (0), mild (1), moderate (2) and severe (3). For morphological analysis, ten images from the ileum of each animal were randomly captured and analyzed through ImageJ software. Granular density inside Paneth cells was determined by measuring the intracellular area occupied by secretory granules. Villus height and crypt depth was measured vertically from the tip of a villus to the base of the adjacent crypt. Villus height⁄crypt height ratio from the intestinal epithelium was also obtained.
Determination of intestinal myeloperoxidase and eosinophil peroxidase activity
The extent of neutrophil accumulation in the small bowel was assessed by determination of myeloperoxidase (MPO) activity, as described previously [38]. Briefly, a 3 cm portion of mice intestines were removed and 100 mg of intestine were weighted and homogenized with 1 mL of PBS and centrifuged at 12,000g for 10 min. The supernatant was discarded, and the erythrocytes were lysed. The samples were then centrifuged, the supernatant was discarded, and the pellet was suspended in 1 mL of 0.05 M Na3PO4, frozen three times in liquid nitrogen, and centrifuged at 4 °C at 12,000g for 10 min. The supernatant was used in the enzymatic assay by the addition of an equal amount substrate (2.9 mmol of tetramethylbenzidine in DMSO). The reaction was stopped with 50 µL of 1 M H2SO4, and the absorbance was read at 450 nm. Results were expressed as the relative unit that denotes activity of MPO related with casein-elicited murine peritoneal neutrophils processed in the same way.
The extent of tissue eosinophil infiltration was assessed by measurement of eosinophil peroxidase (EPO) activity, as previously described [39]. Briefly, 100 mg of intestine were weighted, homogenized with 1.9 mL of PBS and centrifuged at 12,000g for 10 min. Supernatant was discarded, and pellet was resuspended in 1.9 mL of 0.5% hexadecyltrimethyl ammonium bromide diluted in PBS. After being frozen three times in liquid nitrogen, samples were centrifuged at 4 °C, 12,000g during 10 min. To test EPO activity, the obtained supernatant was mixture with a substrate (1:1) containing 1.5 mmol/L of o-phenylenediamine, 6.6 mmol/L of H2O2 and 0.075 mmol/L of Tris–HCl (pH 8). Reaction was stopped with 50 µL of 1 M H2SO4, and absorbance was measured at 492 nm.
Secretory IgA
Levels of secretory IgA were determined by enzyme linked immunosorbent assay (ELISA) in small bowel intestinal fluids. Microtitre plates (Nunc-Immuno Plates, MaxiSorp) were coated with goat anti-mouse antibody (Southern Biotechnology, Birmingham, AL, USA) in carbonate-bicarbonate buffer (0.1 M Na2CO3/NaHCO3—pH 9.6) for 18 h at 4 °C. Wells were washed with washing solution (saline 0.9% plus 0.05% tween 20) and blocked with 200 µL of 0.05% casein in PBS for 1 h at room temperature. Intestinal fluids previously centrifuged at 432g for 20 min were added to the plate and diluted in PBS-0.25% casein (two times until dilution 1:80). After incubation of 1 h at room temperature, plate was washed and biotin conjugated anti-mouse IgA antibody (Southern Biotechnology) in PBS-0.25% casein (1:10.000) was added to the wells. After incubation of 1 h at 37 °C, peroxidase-streptavidin goat anti-mouse IgA (Southern Biotechnology, Birmingham, AL, USA) was added; plate was incubated for 1 h more and, then, coated with 100 µL/well of orthophenylenediamine (OPD) (1 mg/mL) (Sigma, St. Louis, MO, USA) and 0.04% H2O2 substrates. Color was developed at room temperature and reaction was stopped by the addition of 20 µL/well of 2 N H2SO4. Absorbance was measured at 492 nm using a Bio-Rad Model 450 Microplate Reader. Results were expressed as concentration (µg/mL), according to the standard curve.
Statistical analysis
Differences between groups were statistically evaluated by one way analysis of variance (ANOVA). Bonferroni test was applied to calculate statistical significance across groups. Non-parametric Mann–Whitney test was used for data sets based on scores or percentages. All data was processed using GraphPad Prism 5.0 software. P values under 0.05 were considered significant.
Results
L. lactis supernatant inhibits in vitro growth of L. monocytogenes
The antagonistic activity of LL or LL-PAP culture supernatants was assessed against the food-borne pathogen L. monocytogenes. After 2 h, it was not observed any statistical differences in CFU counts of L. monocytogenes inoculated in BHI + Cm + Nisin medium or in the supernatant of LL and LL-PAP, both containing Cm and Nisin (Fig. 1a). However, after 4 h, it was observed a twofold decrease in CFU counts of L. monocytogenes grown either in LL + Cm + Nisin or in LL-PAP + Cm + Nisin culture supernatants when compared to counts obtained for the pathogen inoculated in BHI + Cm + Nisin medium. Presence of PAP in the supernatant did not reduce L. monocytogenes counts (Fig. 1b).

Inhibitory activity of bacterial supernantants from L. lactis and L. lactis-PAP against pathogenic L. monocytogenes, after 2 (a) and 4 h (b) incubation. *p < 0.05
PAP-producing L. lactis strain supernatant inhibits in vitro of E. faecalis
The inhibitory activity of LL or LL-PAP culture supernatants was evaluated against the opportunistic commensal E. faecalis. After 2 h, it was observed a statistical differences in CFU counts of E. faecalis inoculated in BHI + Cm + Nisin medium compared to supernatant of both LL and LL-PAP cultures, 1.6- and 2-fold respectively (Fig. 2a). Moreover, after 4 h, it was observed a 2.2-fold decrease in CFU counts of E. faecalis grown in LL-PAP + Cm + Nisin culture supernatants when compared to counts obtained for the enterococci inoculated in LL + Cm + Nisin culture. LL supernatant did not reduce E. faecalis counts after 4 h (Fig. 2b).

Inhibitory activity of bacterial supernantants from L. lactis and L. lactis-PAP against pathogenic E. faecalis, after 2 (a) and 4 h (b) incubation. **p < 0.003; *p < 0.05
L. lactis administration prevents 5-FU-induced mucosal inflammation in the ileum
Mice injected with 5-FU showed significant decreases in body weight (10 and 17% loss) from groups fed with water or M17 + Cm + Nisin medium as expected (Fig. 3). Treatment with L. lactis strains did not improve mouse weight as groups receiving LL or LL-PAP cultures lost approximately 10% of their initial body weight after 5-FU injection (Fig. 3b). Histological analysis revealed mucosal pattern within normal limits in all groups injected with 0.9% saline (Fig. 4a). 5-FU injection in mice receiving water or M17 + Cm + Nisin medium caused lesions in the small intestine characterized by an inflammatory cellular infiltrate in lamina propria, as well as in the submucosa and muscular layers, with increased number of histopathological parameters (Fig. 4b). Mice immunized with LL prevented 5-FU-induced mucosal inflammation in the ileum, showing reduced infiltration by polymorphonuclear neutrophils, ulceration and reduced alterations of intestinal mucosal architecture. Treatment with LL-PAP strain did not improve the histological score (Fig. 4b).

Time-course of body weight for mice injected with saline or 5-fluoracil receiving water, M17 + Cm + Nisin medium or L. lactis and L. lactis-PAP strains (a). Weight loss observed after 5-FU injection and differences across groups (b).*p < 0.05; ***p < 0.0001

Representative images from mucosal histopathology (a) and histopathological scores obtained for experimental groups (b). ***p < 0.0001; **p < 0.003
Delivery of human PAP by L. lactis improves villous architecture preservation and Paneth cells activity in ileum
Morphometric analysis was carried out to evaluate epithelial integrity. A decrease was observed in villus height (Fig. 5a), villus/crypt ratio (Fig. 5c) and granular density inside Paneth cells (Fig. 6) after 5-FU injection in mice receiving water or M17 + Cm + Nisin medium. No statistical differences in crypt depth were observed across groups (Fig. 5b). Inflamed mice treated with LL-PAP showed increased villus height (Fig. 5a), vilus/crypt ratio (Fig. 5c) and granular density within Paneth cells (Fig. 6a, b) when compared to mice treated with LL.

Morphometric analysis of villus height (a), crypts depth (b) and villus height/crypt depth ratio (c). ***p < 0,0001; **p < 0.003; *p < 0.005

Representative images from Paneth cells morphology (a) and microscopic morphometric analysis of Paneth cell secretory granules (b). *p > 0.05
L. lactis treatment reduces neutrophilic and eosinophilic infiltration in the ileum
To evaluate whether treatment with LL or LL-PAP strain would have an effect in reducing polymorphonuclear cells infiltration in the small bowel of mice, MPO and EPO activity were measured in cell lysates of the ileum. As expected, 5-FU administration increased both intestinal MPO and EPO activity in mice administered with water or M17 + Cm + Nisin medium (Figs. 7, 8). Treatment with LL culture demonstrated to reduce infiltration those enzymes activity when compared to mice that were given M17 + Cm + Nisin medium. Expression of PAP by L. lactis did not reduce neither EPO nor MPO activity (Figs. 7, 8).

Differences across experimental groups in myeloperoxidase activity from cell lysates of the ileum. ***p < 0.0001; *p > 0.05

Differences across experimental groups in eosinophil peroxidase activity from cell lysates of the ileum. **p < 0.003; *p > 0.05
L. lactis reduces IgA secretion in the small bowel
Secretory IgA (sIgA) response was also measured in the small bowel of mice as it plays an important role in mucosal protection. sIgA secretion increased in mice injected with 5-FU receiving water or M17 + Cm + Nisin medium, as expected (Fig. 9). Group of animals immunized with LL or LL-PAP strain demonstrated reduced levels of IgA when compared to M17-treated group (Fig. 9). However, LL-PAP administration did not caused a significant reduction of sIgA when compared to LL-treated mice (Fig. 9).

Evaluation of IgA secretion in the small bowel from inflamed and noninflamed mice receveing water, M17 medium or L. lactis and L. lactis-PAP strains. ***p < 0.0001; **p < 0.003
Discussion
PAP anti-microbial role against Gram-positive pathogenic bacteria has been demonstrated in previous studies [29,30,31,32,33,33]. Hence, we decided to investigate whether recombinant PAP expressed by Lactococcus lactis would have an inhibitory in vitro effect against Gram-positive foodborne pathogen Listeria monocytogenes and opportunistic commensal E. faecalis. Interestingly, we found that the bacterial supernatant from either L. lactis NZ9000 or L. lactis expressing PAP cultures could statistically inhibit both L. monocytogenes and E. faecalis growth. Moreover, expression of PAP seems to enhance this inhibitory effect against E. faecalis. In our work we did not add trypsin, which is necessary for PAP N-terminal cleavage and activation [31,32,33], to the medium as a recent work demonstrated that intact unprocessed PAP protein was capable of binding and killing Gram-positive bacteria [29,30,, 30]. Indeed, although we used non-concentrated supernatant from LL-PAP, it inhibited E. faecalis growth revealing PAP anti-microbial activity against this Gram-positive species. Moreover, L. lactis antagonism against both pathogens could be caused by expression of some molecules as lactic acid, bacteriocins or metabolites naturally produced by this LAB species. Actually, Charlier and colleagues [40] had tested the inhibitory effect of seventy-five strains of L. lactis, including L. lactis NZ9000 strain, against Gram-positive pathogenic Staphylococcus aureus. They demonstrated that medium acidification was not involved in the inhibition of S. aureus in early growth phases, suggesting that further experimentation is required to characterize the molecular bases of L. lactis NZ9000 antagonistic activity against this pathogen [40].
Besides its antimicrobial activity, PAP has been reported to be involved in maintaining intestinal homeostasis [32]. Indeed, a recent report from our research group showed that L. lactis expressing PAP was capable to efficiently prevent colitis in mice [34]. Therefore, we supposed that this strain could also display protective effects in other inflammatory disorders of the gastrointestinal tract, as mucositis in which dysbiosis have been implicated as key factor for inducing intestinal inflammation [41,42,43,44,45]. The medicament 5-FU commonly used in oncology generate as an adverse effect mucositis, causing weight loss to patients as around 80% of their epithelial cells population are destroyed by apoptosis thus leading to lower absorption of nutrients [46]. In our study, pretreatment with both L. lactis NZ9000 and L. lactis-PAP strains during 13 days had no influence on weight recovery of inflamed mice. Differently, Bowen and colleagues have shown that VSL#3 probiotic mixture could reduce weight loss in mice with mucositis after a long period of administration (28 days) [15]. Therefore, we believe that longer treatment duration is required to recover weight of mice injected with 5-FU.
A common feature of mucositis patients receiving 5-FU is the presence of inflammatory cells infiltrating into the lamina propria [2, 4]. Another characteristic of mucositis disorder is altered intestinal morphology, such as villus shortening and decreased villus/crypt ratio, with loss of the mucosal barrier integrity [7,8,9,10,11]. Thus, all this parameters were evaluated in this work in order to test the anti-inflammatory activity of both LL and LL-PAP strains. Histological analysis revealed that the administration of L. lactis NZ9000 in inflamed mice decreased the histopathological scores, with less polymorphonuclear infiltration and ulceration observed in samples. Surprisingly, PAP expression by L. lactis did not improve these parameters. Despite LL-PAP did not improve histopathological scores, morphometric analysis showed that it was able to better preserve intestinal villous architecture. Moreover, LL-PAP was able to preserve and improve Paneth cells activity, thus augmenting antimicrobial gastrointestinal function, as these cells are associated with the secretion of antimicrobial peptides. This was a very interesting result because PAP has also been reported to stimulate the proliferation of epithelial cells in the colon, including Paneth cells [47]. Mucositis is characterized by a very acute mucosal inflammation, in which the recruitment of neutrophils plays a major role on the pathophysiology process through the release of reactive oxygen species and inflammatory mediators [2]. Thus, we measured the activity of MPO enzyme, strictly produced by neutrophils, to estimate the cell influx into intestinal lamina propria from non-inflamed and inflamed mice receiving water, medium or LL and LL-PAP strains. Interestingly, we demonstrated that LL or LL-PAP treated mice reduced MPO activity after 5-FU injection. This result reveals the anti-inflammatory capacity of L. lactis NZ9000 strain to exempt neutrophil recruitment to the tissue. Moreover, recent work demonstrated that the increased influx of eosinophils is also an important event for the pathogenesis of mucositis [45]. Thus, we measured the activity of EPO enzyme, naturally produced by eosinophils, to estimate eosinophil influx into intestinal lamina propria across experimental groups. We showed that L. lactis NZ9000 strain expressing or not PAP was able to reduce eosinophil infiltration. A similar result was described by Holvoet colleagues [48] when another strain, L. lactis NCC2287, was used for the treatment of Eosinophilic esophagitis in mice. They showed that lactococci administration in mice significantly decreased esophageal eosinophilia, elicited by epicutaneous sensitization with Aspergillus fumigatus protein extract, reiterating the beneficial effects of L. lactis in another severe inflammatory disease [48].
Levels of secretory IgA were also determined as it is crucial to prevent pathogens to penetrate the epithelial barrier and, thus, to contain inflammatory processes [49]. We found that animals with mucositis that did not receive lactococci treatment had increased sIgA levels in the small bowel. This effect was expected, as the amount of this immunoglobulin increases during intestinal inflammatory process as a defense mechanism of the host [49, 50]. Interestingly, L. lactis NZ9000 administration decreased IgA secretion in mouse intestinal lumen. Again, no improvements were obtained with the use of L. lactis secreting PAP strain.
Altogether, our results have shown that L. lactis NZ9000 strain carrying pSEC vector without the cDNA of PAP was able to prevent 5-FU-induced intestinal inflammation in BALB/c mice. This was a very surprising and intriguing result because L. lactis NZ9000 strain is a derivative of L. lactis MG1363 used as starter cultures for cheddar cheese production [51, 52], and has not been reported as improving host health. Actually, few studies have reported beneficial effects of this species [48, 50, 53, 54]. Ballal and colleagues found that L. lactis I-1631 ameliorated colitis in T-bet−/− Rag2−/− mice and other two studies have demonstrated that either NCDO2118 sub. lactis or FC sub. cremoris presents anti-inflammatory properties in inflamed mice receiving chemical agent Dextran Sulphate Sodium (DSS) [48, 50, 53].
Furthermore, PAP expression by L. lactis subtly alleviate mucositis damage, as it did not show to decrease markers of inflammation such as, ulceration, pro-inflammatory cells infiltrate and IgA levels, but preserved architecture and increased secretory granules density inside Paneth cells in response to 5-FU inflammation.
Conclusion
In conclusion, we have demonstrated that the allochthonous bacterium, L. lactis NZ9000, derived from dairy L. lactis MG1363 reveals to be a promising tool to prevent chemotherapy drug 5-Fluoracil-induced mucositis. Moreover, we opened the doors for future studies investigating possible factors involved in L. lactis NZ9000 anti-inflammatory effects. As beneficial effects has been demonstrated by the recombinant LL-PAP strain, further studies should be considered, such as biological confinement strategies preventing its dissemination into nature, in order to make it a safe approach to be tested in humans.
Abbreviations
- 5-FU:
-
5-fluoracil
- AMP:
-
antimicrobial peptides
- DNBS:
-
dinitrobenzenosulfonic acid
- EPO:
-
eosinophil peroxidase
- IBD:
-
inflammatory bowel diseases
- LAB:
-
lactic acid bacteria
- NICE:
-
Nisin Control Expression System
- PAP:
-
pancreatitis-associated protein
- sIgA:
-
secretory immunoglobulin A
- TFF-1:
-
trefoil factor 1
References
Murphy BA, Beaumont JL, Isitt J, Garden AS, Gwede CK, Trotti AM, Meredith RF, Epstein JB, Le Q-T, Brizel DM. Mucositis-related morbidity and resource utilization in head and neck cancer patients receiving radiation therapy with or without chemotherapy. J Pain Symptom Manage. 2009;38:522–32.
Sonis ST. The pathobiology of mucositis. Nat Rev Cancer. 2004;4:277–84.
Trotti A, Bellm LA, Epstein JB, Frame D, Fuchs HJ, Gwede CK, Komaroff E, Nalysnyk L, Zilberberg MD. Mucositis incidence, severity and associated outcomes in patients with head and neck cancer receiving radiotherapy with or without chemotherapy: a systematic literature review. Radiother Oncol. 2003;66:253–62.
Chang C-T, Ho T-Y, Lin H, Liang J-A, Huang H-C, Li C-C, Lo H-Y, Wu S-L, Huang Y-F, Hsiang C-Y. 5-Fluorouracil induced intestinal mucositis via nuclear factor-κB activation by transcriptomic analysis and in vivo bioluminescence imaging. PLoS ONE. 2012;7:e31808.
Longley DB, Harkin DP, Johnston PG. 5-Fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003;3:330–8.
Parker WB, Cheng YC. Metabolism and mechanism of action of 5-fluorouracil. Pharmacol Ther. 1990;48:381–95.
Soares PMG, Mota JMSC, Souza EP, Justino PFC, Franco AX, Cunha FQ, Ribeiro RA, Souza MHLP. Inflammatory intestinal damage induced by 5-fluorouracil requires IL-4. Cytokine. 2013;61:46–9.
Maioli TU, de Melo Silva B, Dias MN, Paiva NC, Cardoso VN, Fernandes SO, Carneiro CM, dos Santos Martins F, de Vasconcelos Generoso S. Pretreatment with Saccharomyces boulardii does not prevent the experimental mucositis in Swiss mice. J Negat Results Biomed. 2014;13:6.
de Vasconcelos Generoso S, Rodrigues NM, Trindade LM, Paiva NC, Cardoso VN, Carneiro CM, de Matos Ferreira AV, Faria AMC, Maioli TU. Dietary supplementation with omega-3 fatty acid attenuates 5-fluorouracil induced mucositis in mice. Lipids Health. 2015;14:1.
Antunes MM, Leocádio PCL, Teixeira LG, Leonel AJ, Cara DC, Menezes GB, de Vasconcelos Generoso S, Cardoso VN, Alvarez-Leite JI, Correia MITD. Pretreatment with l-citrulline positively affects the mucosal architecture and permeability of the small intestine in a murine mucositis model. JPEN J Parenter Enteral Nutr. 2016;40(2):279–86.
Leocádio PCL, Antunes MM, Teixeira LG, Leonel AJ, Alvarez-Leite JI, Machado DCC, Generoso SV, Cardoso VN, Correia MITD. l-Arginine pretreatment reduces intestinal mucositis as induced by 5-FU in mice. Nutr Cancer. 2015;67:486–93.
Sonis ST. Mucositis: the impact, biology and therapeutic opportunities of oral mucositis. Oral Oncol. 2009;45:1015–20.
Plevová P. Prevention and treatment of chemotherapy- and radiotherapy-induced oral mucositis: a review. Oral Oncol. 1999;35:453–70.
Tooley KL, Howarth GS, Lymn KA, Lawrence A, Butler RN. Oral ingestion of Streptococcus thermophilus diminishes severity of small intestinal mucositis in methotrexate treated rats. Cancer Biol Ther. 2006;5:593–600.
Bowen JM, Stringer AM, Gibson RJ, Yeoh ASJ, Hannam S, Keefe DMK. VSL#3 probiotic treatment reduces chemotherapy-induced diarrhea and weight loss. Cancer Biol Ther. 2007;6:1449–54.
Smith CL, Geier MS, Yazbeck R, Torres DM, Butler RN, Howarth GS. Lactobacillus fermentum BR11 and fructo-oligosaccharide partially reduce jejunal inflammation in a model of intestinal mucositis in rats. Nutr Cancer. 2008;60:757–67.
Southcott E, Tooley KL, Howarth GS, Davidson GP, Butler RN. Yoghurts containing probiotics reduce disruption of the small intestinal barrier in methotrexate-treated rats. Dig Dis Sci. 2008;53:1837–41.
Prisciandaro LD, Geier MS, Butler RN, Cummins AG, Howarth GS. Probiotic factors partially improve parameters of 5-fluorouracil-induced intestinal mucositis in rats. Cancer Biol Ther. 2011;11:671–7.
WHO. Guidelines for the evaluation of probiotics in food: joint FAO/WHO Working Group meeting, London, 30 April–1 May; 2002.
Zurita-Turk M, del Carmen S, Santos AC, Pereira VB, Cara DC, Leclercq SY, dM de LeBlanc A, Azevedo V, Chatel J-M, LeBlanc JG. Lactococcus lactis carrying the pValac DNA expression vector coding for IL-10 reduces inflammation in a murine model of experimental colitis. BMC Biotechnol. 2014;14:73.
del Carmen S, Martín Rosique R, Saraiva T, Zurita-Turk M, Miyoshi A, Azevedo V, de Moreno de LeBlanc A, Langella P, Bermúdez-Humarán LG, LeBlanc JG. Protective effects of lactococci strains delivering either IL-10 protein or cDNA in a TNBS-induced chronic colitis model. J Clin Gastroenterol. 2014;48(Suppl 1):S12–7.
LeBlanc JG, Aubry C, Cortes-Perez NG, de Moreno de LeBlanc A, Vergnolle N, Langella P, Azevedo V, Chatel J-M, Miyoshi A, Bermúdez-Humarán LG. Mucosal targeting of therapeutic molecules using genetically modified lactic acid bacteria: an update. FEMS Microbiol Lett. 2013;344:1–9.
Saraiva TDL, Morais K, Pereira VB, de Azevedo M, Rocha CS, Prosperi CC, Gomes-Santos AC, Bermudez-Humaran L, Faria AMC, Blottiere HM, et al. Milk fermented with a 15-lipoxygenase-1-producing Lactococcus lactis alleviates symptoms of colitis in a murine model. Curr Pharm Biotechnol. 2015;16:424–9.
Braat H, Rottiers P, Hommes DW, Huyghebaert N, Remaut E, Remon J-P, van Deventer SJH, Neirynck S, Peppelenbosch MP, Steidler L. A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin Gastroenterol Hepatol. 2006;4:754–9.
Caluwaerts S, Vandenbroucke K, Steidler L, Neirynck S, Vanhoenacker P, Corveleyn S, Watkins B, Sonis S, Coulie B, Rottiers P. AG013, a mouth rinse formulation of Lactococcus lactis secreting human trefoil factor 1, provides a safe and efficacious therapeutic tool for treating oral mucositis. Oral Oncol. 2010;46:564–70.
Limaye SA, Haddad RI, Cilli F, Sonis ST, Colevas AD, Brennan MT, Hu KS, Murphy BA. Phase 1b, multicenter, single blinded, placebo-controlled, sequential dose escalation study to assess the safety and tolerability of topically applied AG013 in subjects with locally advanced head and neck cancer receiving induction chemotherapy. Cancer. 2013;2013(119):4268–76.
Wong CCM, Zhang L, Li ZJ, Wu WKK, Ren SX, Chen YC, Ng TB, Cho CH. Protective effects of cathelicidin-encoding Lactococcus lactis in murine ulcerative colitis. J Gastroenterol Hepatol. 2012;27:1205–12.
Seo E, Weibel S, Wehkamp J, Oelschlaeger TA. Construction of recombinant E. coli Nissle 1917 (EcN) strains for the expression and secretion of defensins. Int J Med Microbiol. 1917;2012(302):276–87.
Cash HL, Whitham CV, Behrendt CL, Hooper LV. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science. 2006;313:1126–30.
Medveczky P, Szmola R, Sahin-Tóth M. Proteolytic activation of human pancreatitis-associated protein is required for peptidoglycan binding and bacterial aggregation. Biochem J. 2009;420:335–44.
Mukherjee S, Partch CL, Lehotzky RE, Whitham CV, Chu H, Bevins CL, Gardner KH, Hooper LV. Regulation of C-type lectin antimicrobial activity by a flexible N-terminal prosegment. J Biol Chem. 2009;284:4881–8.
Vaishnava S, Yamamoto M, Severson KM, Ruhn KA, Yu X, Koren O, Ley R, Wakeland EK, Hooper LV. The antibacterial lectin RegIII promotes the spatial segregation of microbiota and host in the intestine. Science. 2011;334:255–8.
Mukherjee S, Zheng H, Derebe MG, Callenberg KM, Partch CL, Rollins D, Propheter DC, Rizo J, Grabe M, Jiang QX. Antibacterial membrane attack by a pore-forming intestinal C-type lectin. Nature. 2014;505:103–7.
Breyner et al. Manuscript in preparation; 2016.
de Ruyter PG, Kuipers OP, de Vos WM. Controlled gene expression systems for Lactococcus lactis with the food-grade inducer nisin. Appl Environ Microbiol. 1996;62:3662–7.
Bermúdez-Humarán LG, Langella P, Miyoshi A, Gruss A, Guerra RT. Montes de Oca-Luna R, Le Loir Y. Production of human papillomavirus type 16 E7 protein in Lactococcus lactis. Appl Environ Microbiol. 2002;68:917–22.
Hütt P, Shchepetova J, Lõivukene K, Kullisaar T, Mikelsaar M. Antagonistic activity of probiotic lactobacilli and bifidobacteria against entero- and uropathogens. J Appl Microbiol. 2006;100:1324–32.
Souza DG, Pinho V, Cassali GD, Poole S, Teixeira MM. Effect of a BLT receptor antagonist in a model of severe ischemia and reperfusion injury in the rat. Eur J Pharmacol. 2002;440:61–9.
Strath M, Sanderson CJ. Production and functional properties of eosinophils from bone marrow cultures. J Cell Sci. 1985;74:207–17.
Charlier C, Even S, Gautier M, Le Loir Y. Acidification is not involved in the early inhibition of Staphylococcus aureus growth by Lactococcus lactis in milk. Int Dairy J. 2008;18:197–203.
Stringer AM, Gibson RJ, Bowen JM, Logan RM, Ashton K, Yeoh AS, Al-Dasooqi N, Keefe DM. Irinotecan-induced mucositis manifesting as diarrhoea corresponds with an amended intestinal flora and mucin profile. Int J Exp Pathol. 2009;90:489–99.
Vanhoecke B, De Ryck T, Stringer A, Van de Wiele T, Keefe D. Microbiota and their role in the pathogenesis of oral mucositis. Oral Dis. 2015;21:17–30.
Brandi G, Dabard J, Raibaud P, Di Battista M, Bridonneau C, Pisi AM. Morselli Labate AM, Pantaleo MA, De Vivo A, Biasco G. Intestinal microflora and digestive toxicity of irinotecan in mice. Clin Cancer Res. 2006;12:1299–307.
Thorpe DW, Stringer AM, Gibson RJ. Chemotherapy-induced mucositis: the role of the gastrointestinal microbiome and toll-like receptors. Exp Biol Med. 2013;238:1–6.
Pedroso SHSP, Vieira AT, Bastos RW, Oliveira JS, Cartelle CT, Arantes RME, Soares PMG, Generoso SV, Cardoso VN, Teixeira MM, et al. Evaluation of mucositis induced by irinotecan after microbial colonization in germ-free mice. Microbiology. 2015;161:1950–60.
Soares PMG, Mota JMSC, Gomes AS, Oliveira RB, Assreuy AMS, Brito GAC, Santos AA, Ribeiro RA, Souza MHLP. Gastrointestinal dysmotility in 5-fluorouracil-induced intestinal mucositis outlasts inflammatory process resolution. Cancer Chemother Pharmacol. 2008;63:91–8.
Lv Y, Yang X, Huo Y, Tian H, Li S, Yin Y, Hao Z. Adenovirus-mediated hepatocarcinoma-intestine-pancreas/pancreatitis-associated protein suppresses dextran sulfate sodium-induced acute ulcerative colitis in rats. Inflamm Bowel Dis. 2012;18:1950–60.
Holvoet S, Doucet-Ladevèze R, Perrot M, Barretto C, Nutten S, Blanchard C. Beneficial effect of Lactococcus lactis NCC 2287 in a murine model of eosinophilic esophagitis. Allergy. 2016;71(12):1753–61.
Schmucker DL, Owen RL, Outenreath R, Thoreux K. Basis for the age-related decline in intestinal mucosal immunity. J Immunol Res. 2003;10:167–72.
Luerce TD, Gomes-Santos AC, Rocha CS, Moreira TG, Cruz DN, Lemos L, Sousa AL, Pereira VB, de Azevedo M, Moraes K. Anti-inflammatory effects of Lactococcus lactis NCDO 2118 during the remission period of chemically induced colitis. Gut Pathog. 2014;6:33.
Fynan EF, Webster RG, Fuller DH, Haynes JR, Santoro JC, Robinson HL. DNA vaccines: protective immunizations by parenteral, mucosal, and gene-gun inoculations. Proc Natl Acad Sci USA. 1993;90:11478–82.
Upadhyay VK, Ravn P, Israelsen H, Sousa MJ, Kelly AL, McSweeney PL. Acceleration of proteolysis during ripening of Cheddar-type cheese using of a streptokinase-producing strain of Lactococcus. J Dairy Res. 2006;73:70–3.
Nishitani Y, Tanoue T, Yamada K, Ishida T, Yoshida M, Azuma T, Mizuno M. Lactococcus lactis subsp. cremoris FC alleviates symptoms of colitis induced by dextran sulfate sodium in mice. Int Immunopharmacol. 2009;9:1444–51.
Ballal SA, Veiga P, Fenn K, Michaud M, Kim JH, Gallini CA, Glickman JN, Quéré G, Garault P, Béal C, et al. Host lysozyme-mediated lysis of Lactococcus lactis facilitates delivery of colitis-attenuating superoxide dismutase to inflamed colons. Proc Natl Acad Sci USA. 2015;112:7803–8.
Authors’ contributions
RDOC performed in vitro analysis, animal experimentation regarding mucositis pre-treatment with the lactococci strains, interpreted the data regarding the immunological parameters that were assessed and was a major contributor in writing the manuscript. NMB, TUM, AMCF, PL, HCPF were major contributors in writing the manuscript. ZMG performed the EPO activity essays, analyzed and interpreted the respective data. NMR performed, analyzed and interpreted the Morphometric analysis from ileum slides. LLS performed, analysed and interpreted the secretory IgA quantification essay. DGS was responsible for conducting the EPO analysis and contributed to the respective interpretation of data and to manuscript writing. DCCM analyzed and interpreted the histological analysis and was a major contributor in writing the manuscript. JMC contributed to data interpretation and was a major contributor in writing the manuscript. LGBH was responsible for ceding the lactococci strains, contributed to data interpretation and was a major contributor in writing the manuscript. VACA and MSPA have equally contributed in the in vivo assay performance, analysis and interpretation of immunological data and as major contributors in writing the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors would like to thank CNPq and CAPES for their financial support and fellowships.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets generated or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
the Protocol no. 366/2012, related to the present project is in agreement with the Ethical Principles in Animal Experimentation, adopted by the Ethics Committee in Animal Experimentation (CEUA/UFMG), and was approved in 11/04/2013. This certificates expires in 11/04/2018.
Funding
The funding agencies Coordination for the Improvement of Higher Education Personnel (CAPES) and National Counsel of Technological and Scientific Development (CNPq) were responsible for giving financial support to this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Vasco Azevedo and Marcela S. de Azevedo equal contribution to senior authorship
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Carvalho, R.D., Breyner, N., Menezes-Garcia, Z. et al. Secretion of biologically active pancreatitis-associated protein I (PAP) by genetically modified dairy Lactococcus lactis NZ9000 in the prevention of intestinal mucositis. Microb Cell Fact 16, 27 (2017). https://doi.org/10.1186/s12934-017-0624-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-017-0624-x